
Shifts in Backbone Conformation of Acetylcholinesterases upon Binding of Covalent Inhibitors, Reversible Ligands and Substrates


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
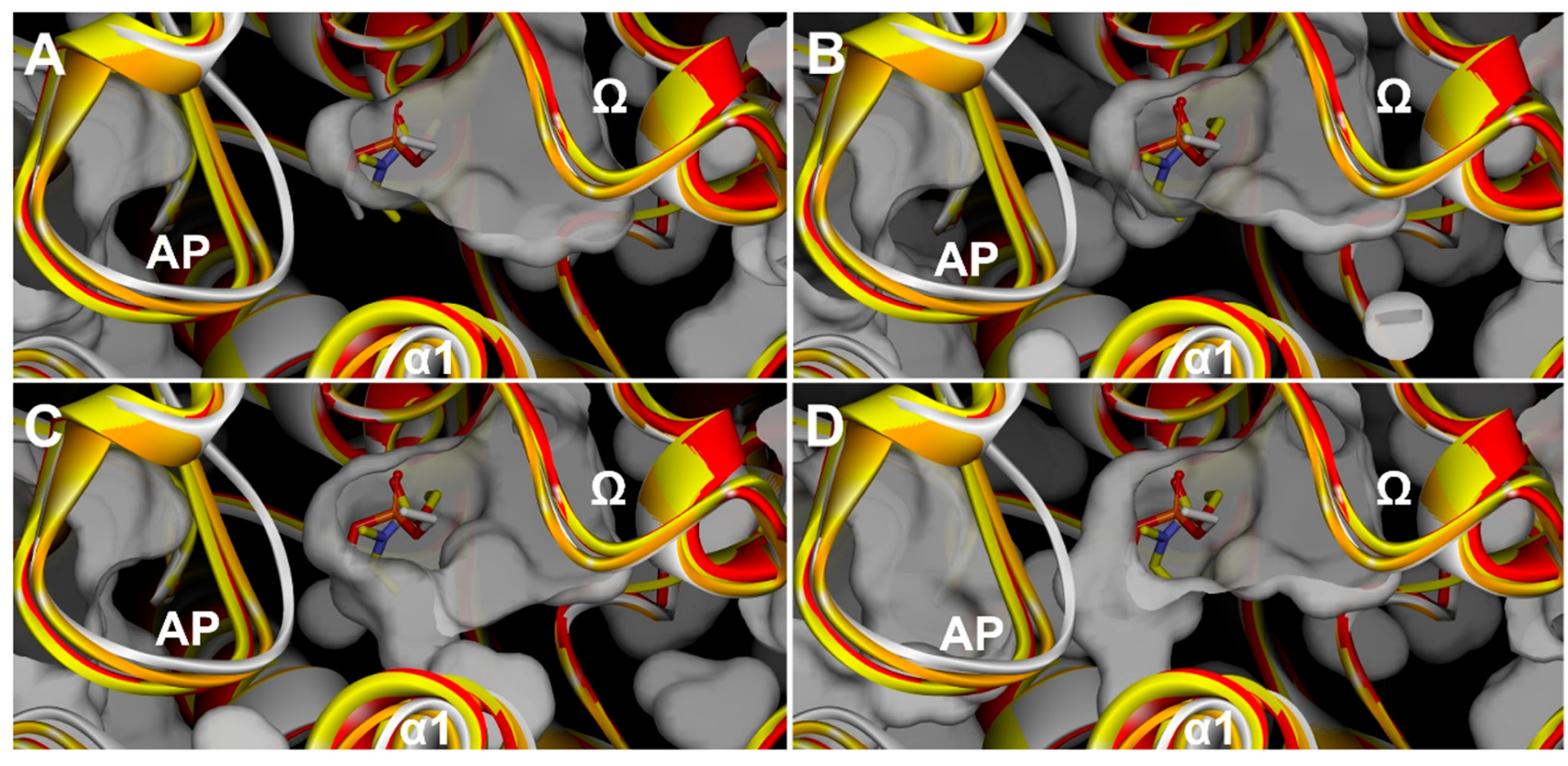
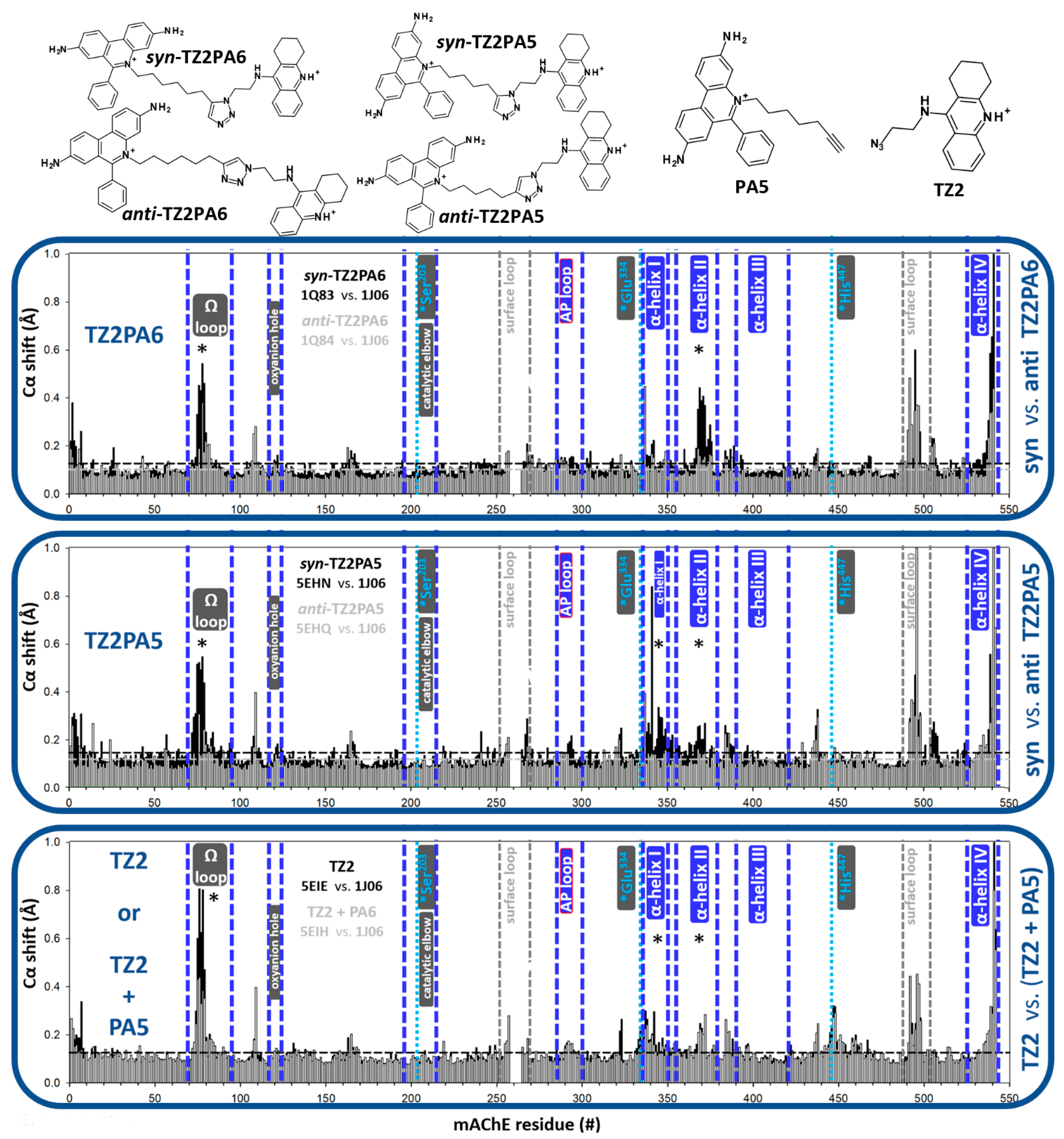
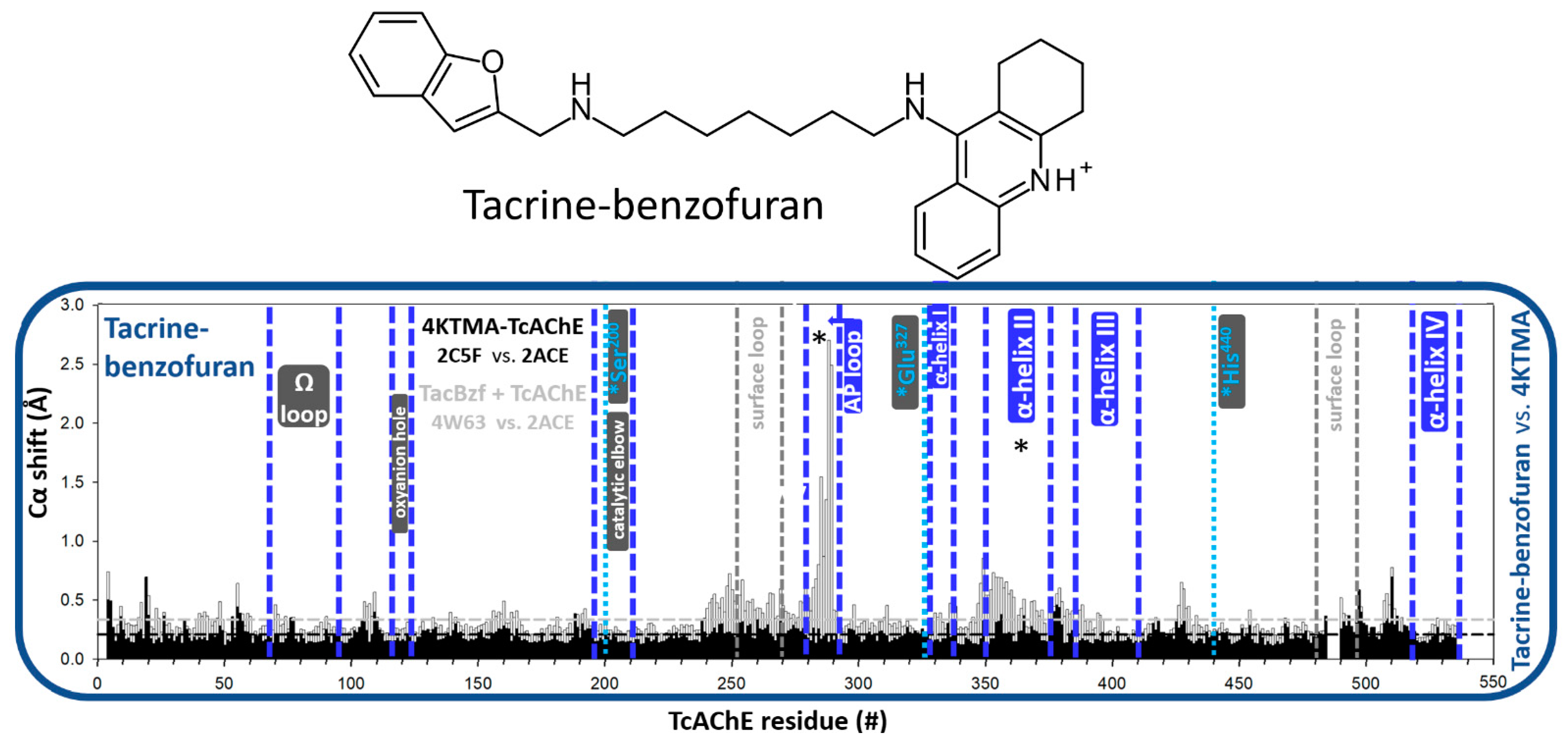
3.1. Effects of Covalent Binding on Backbone Conformation of hAChE
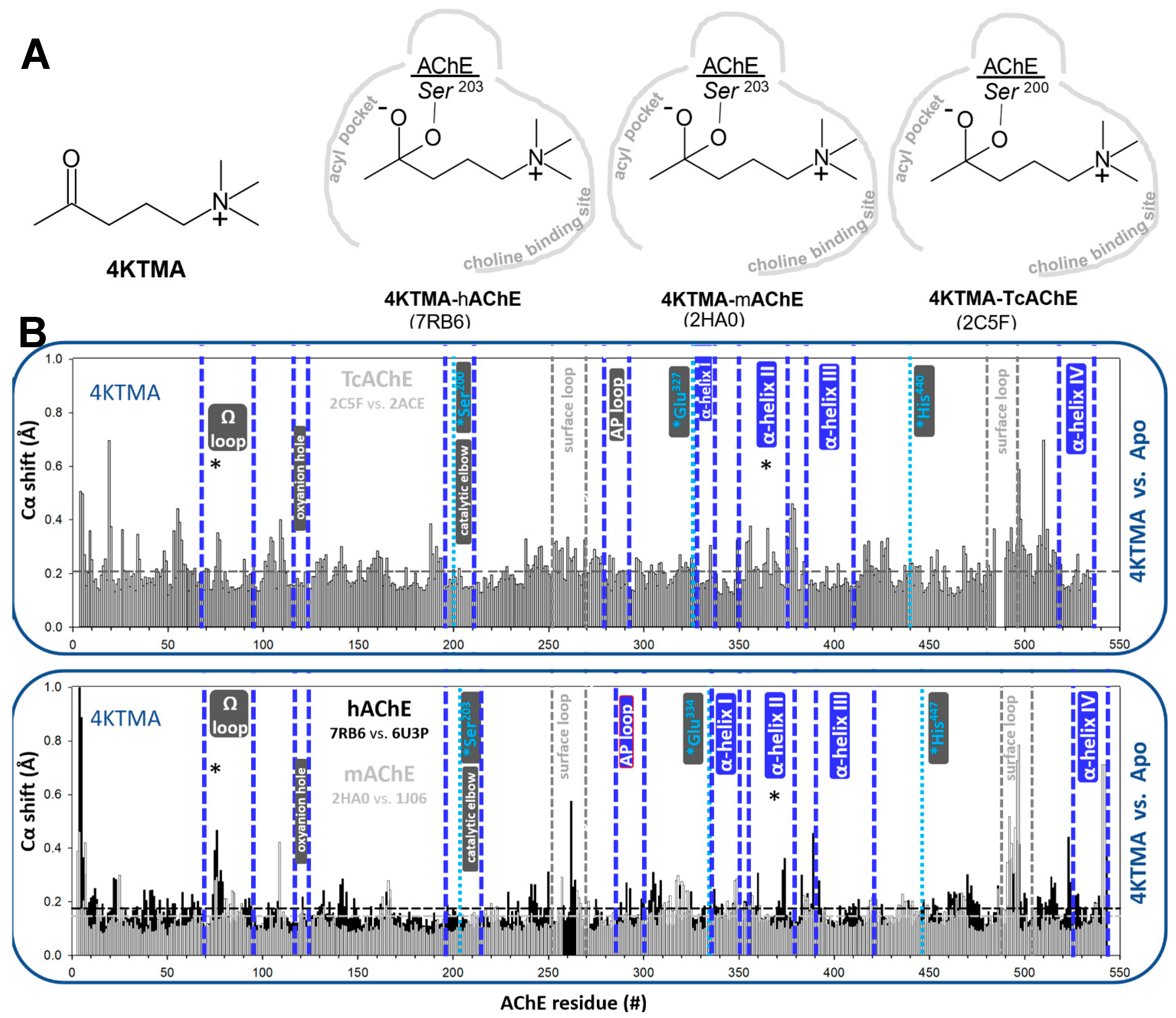
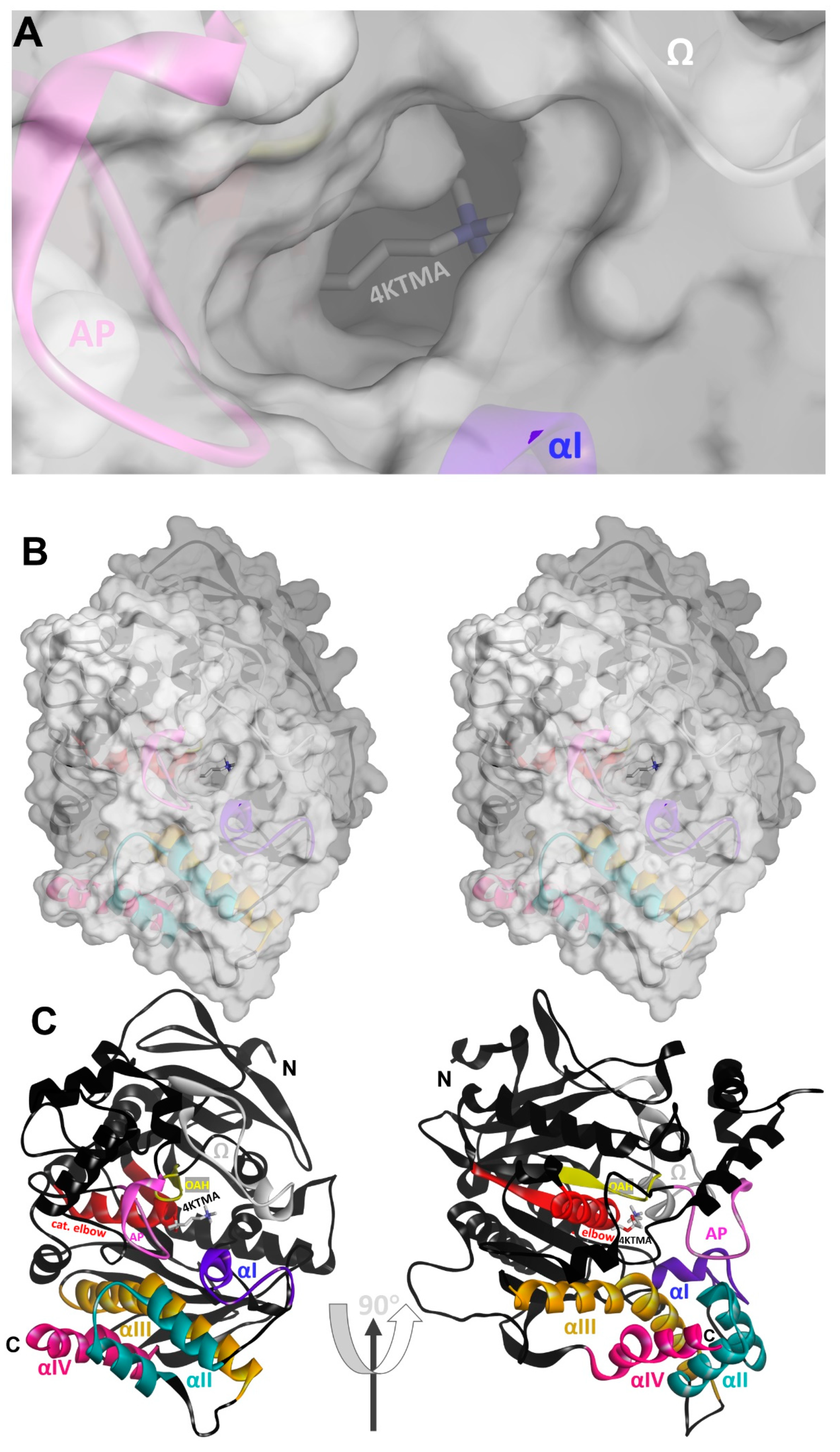
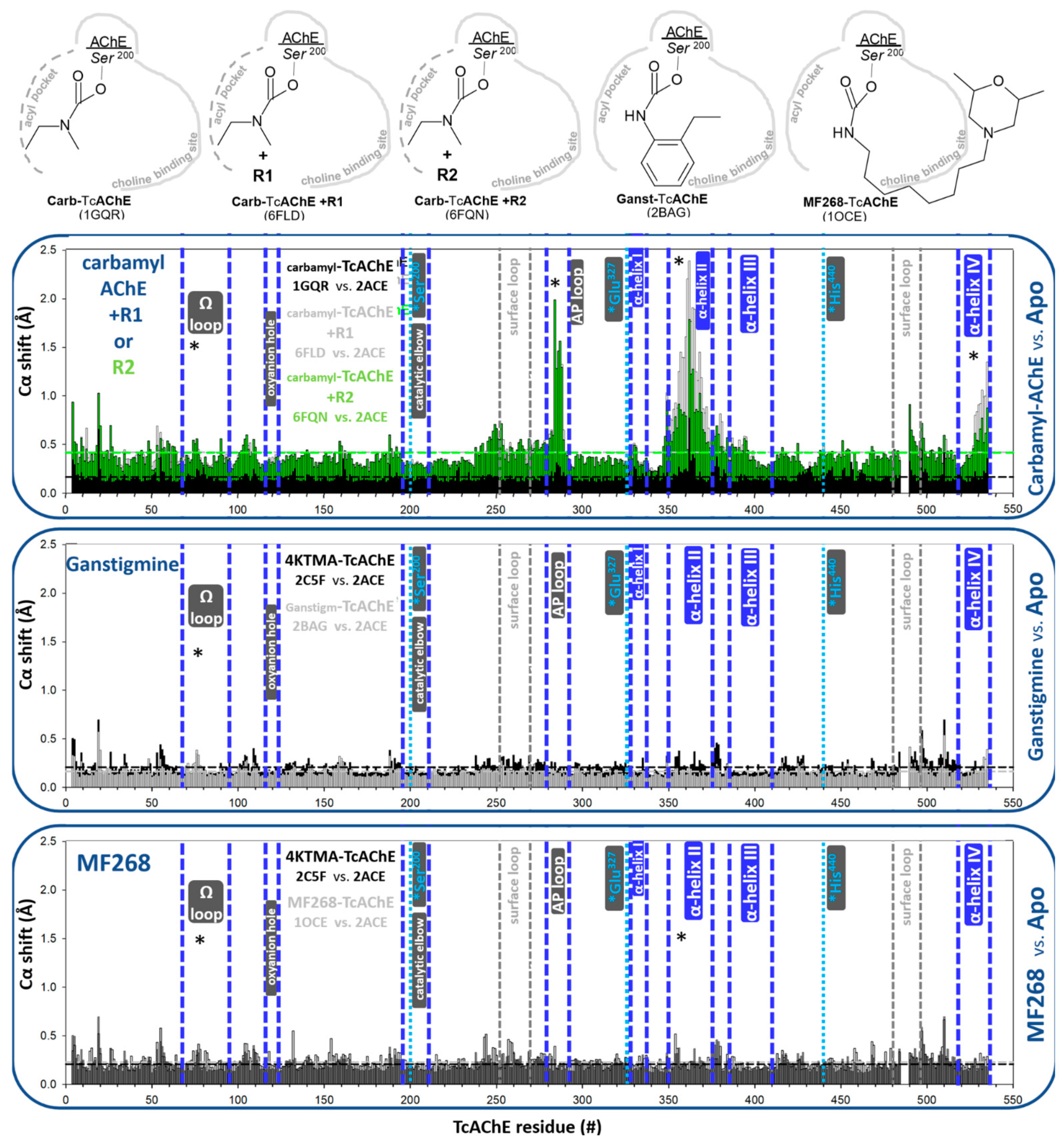
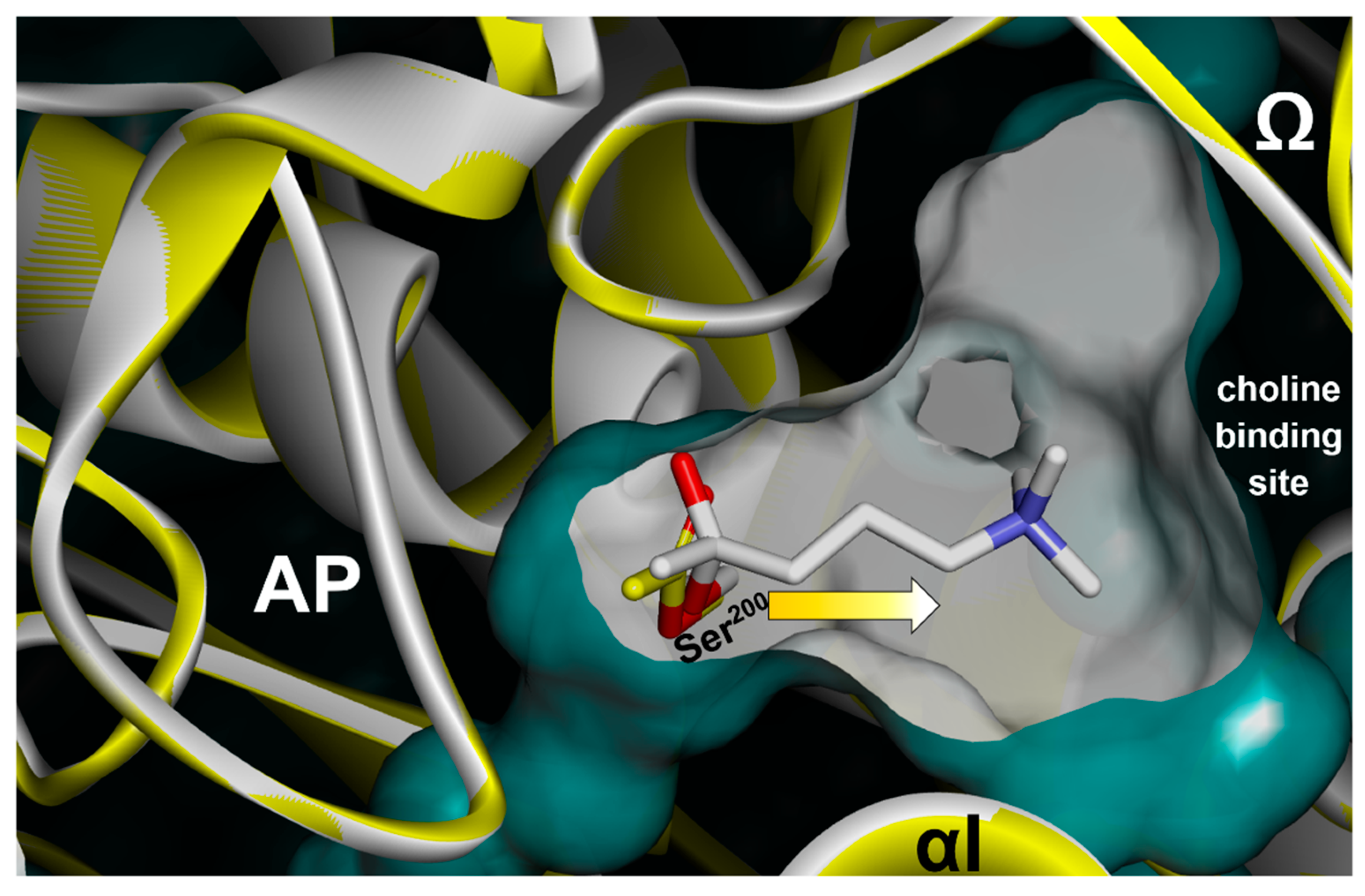
3.1.1. Reversible, Covalent Binding of the Substrate Analogue 4KTMA
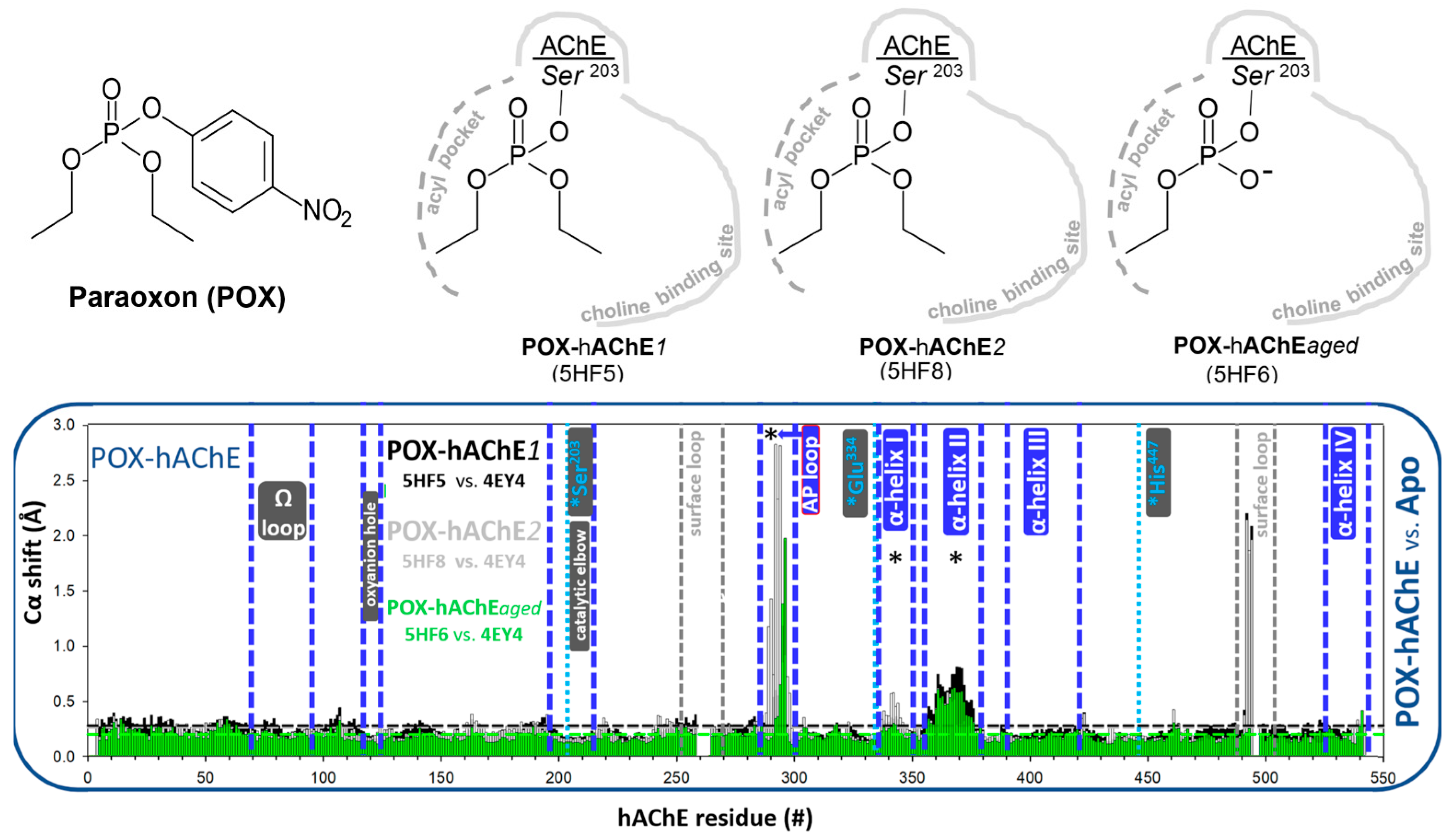
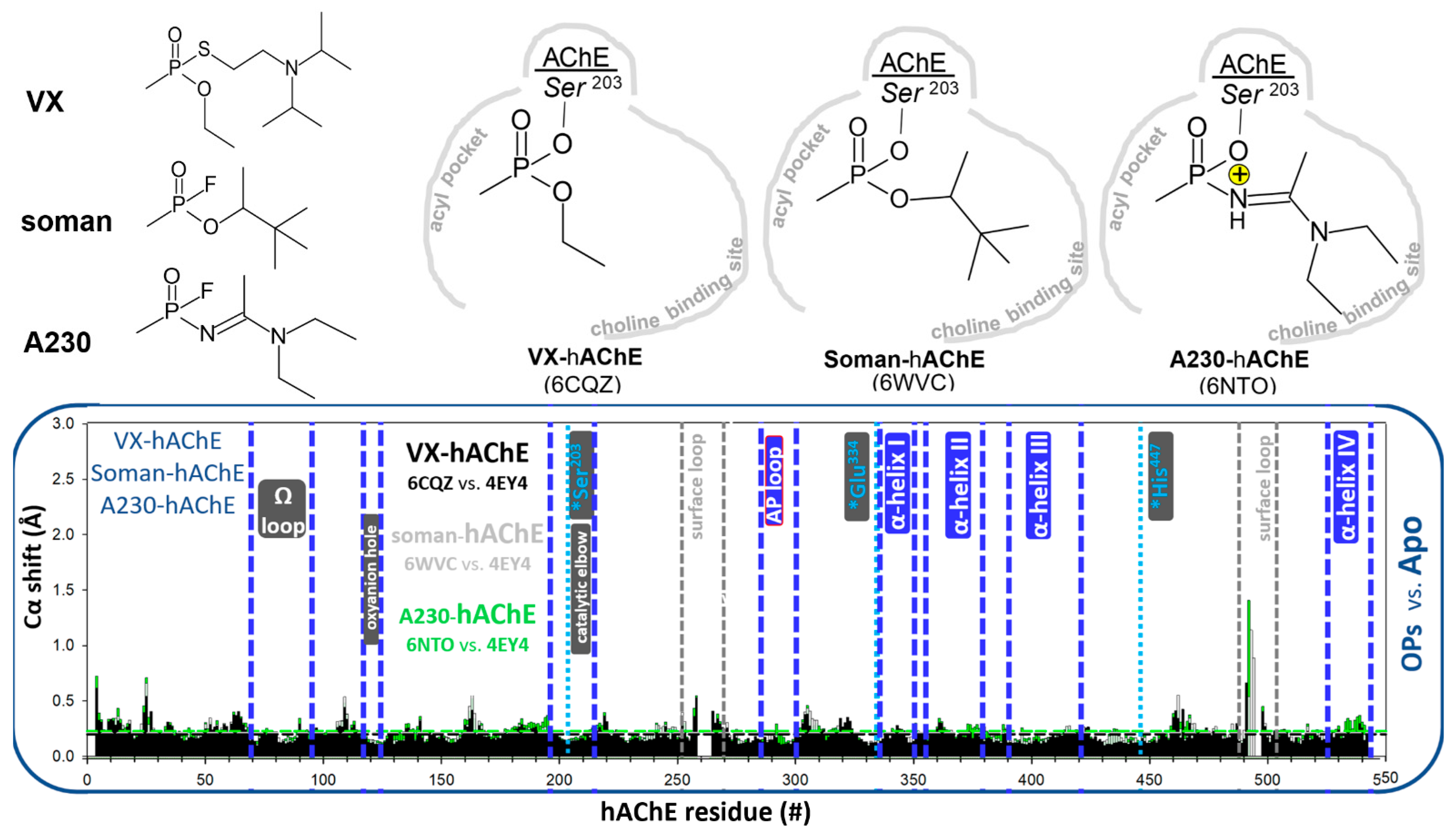
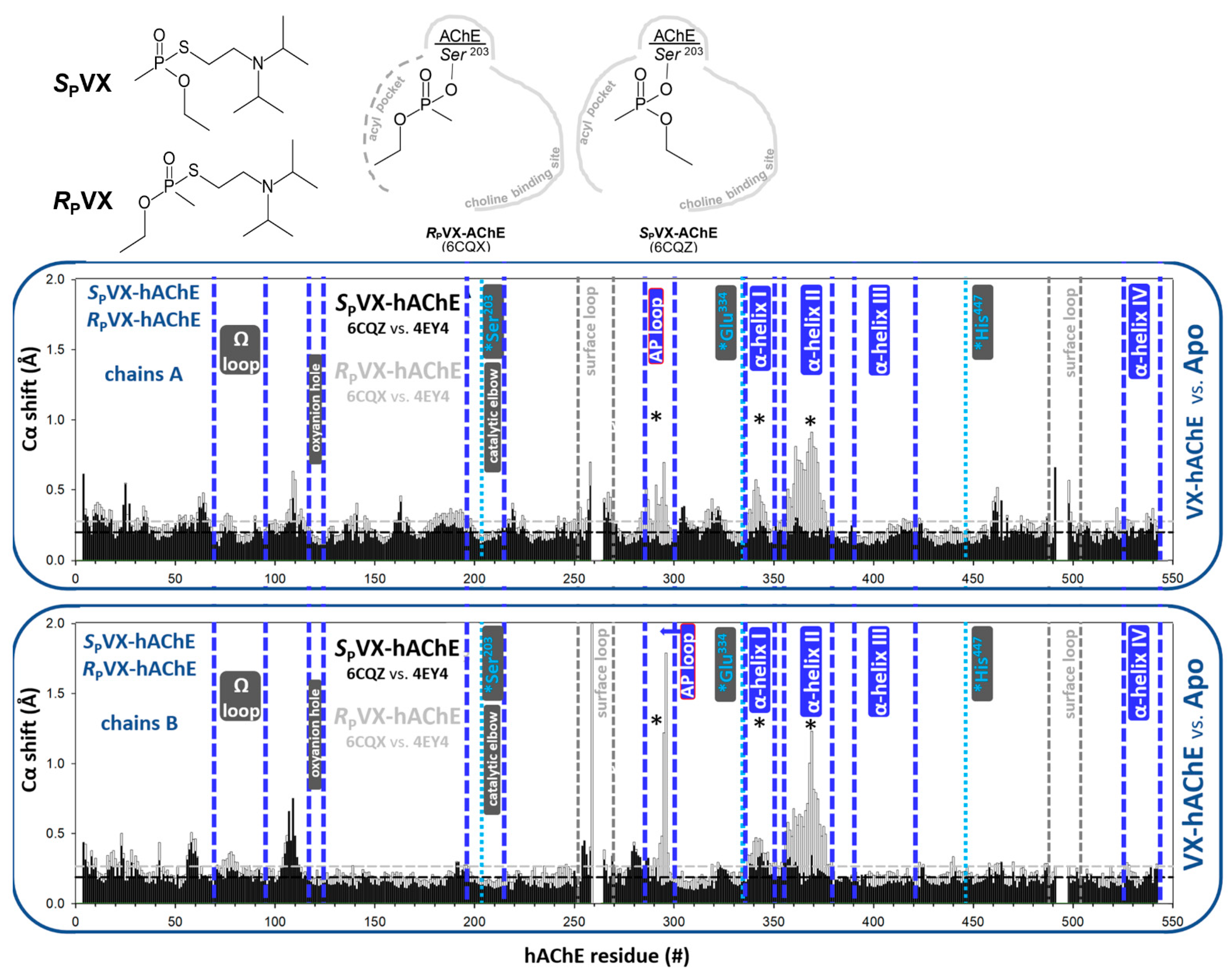
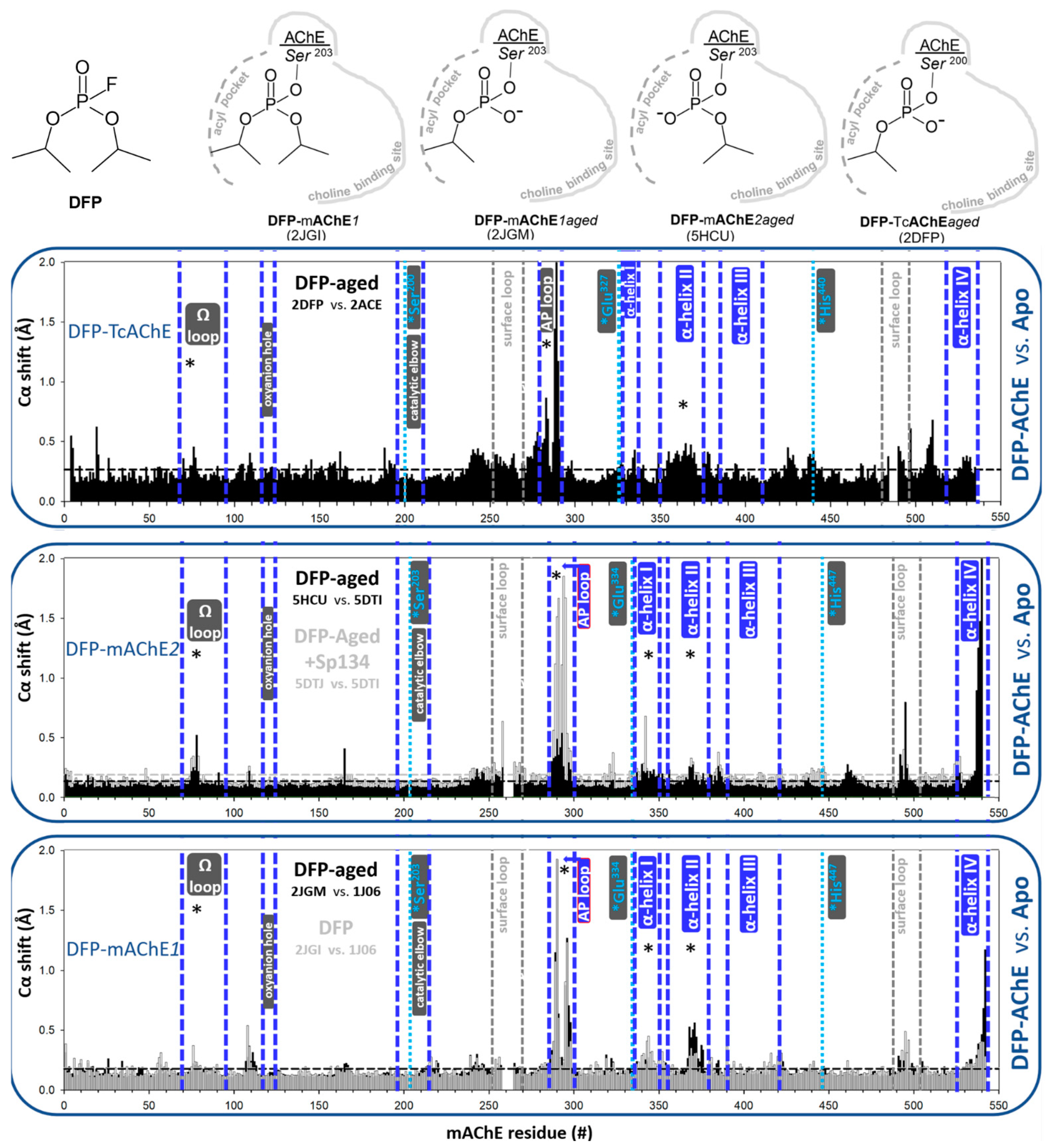
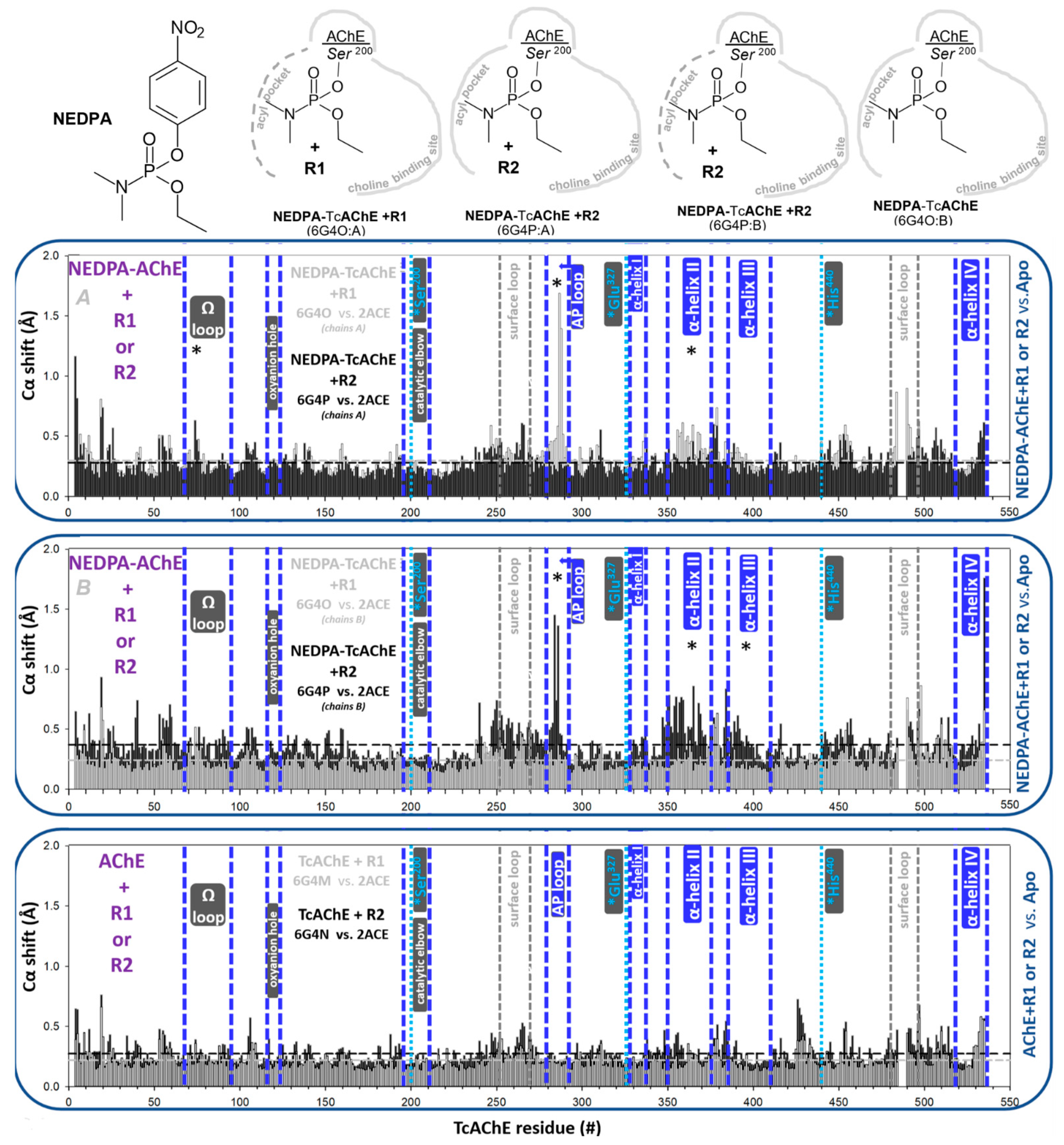
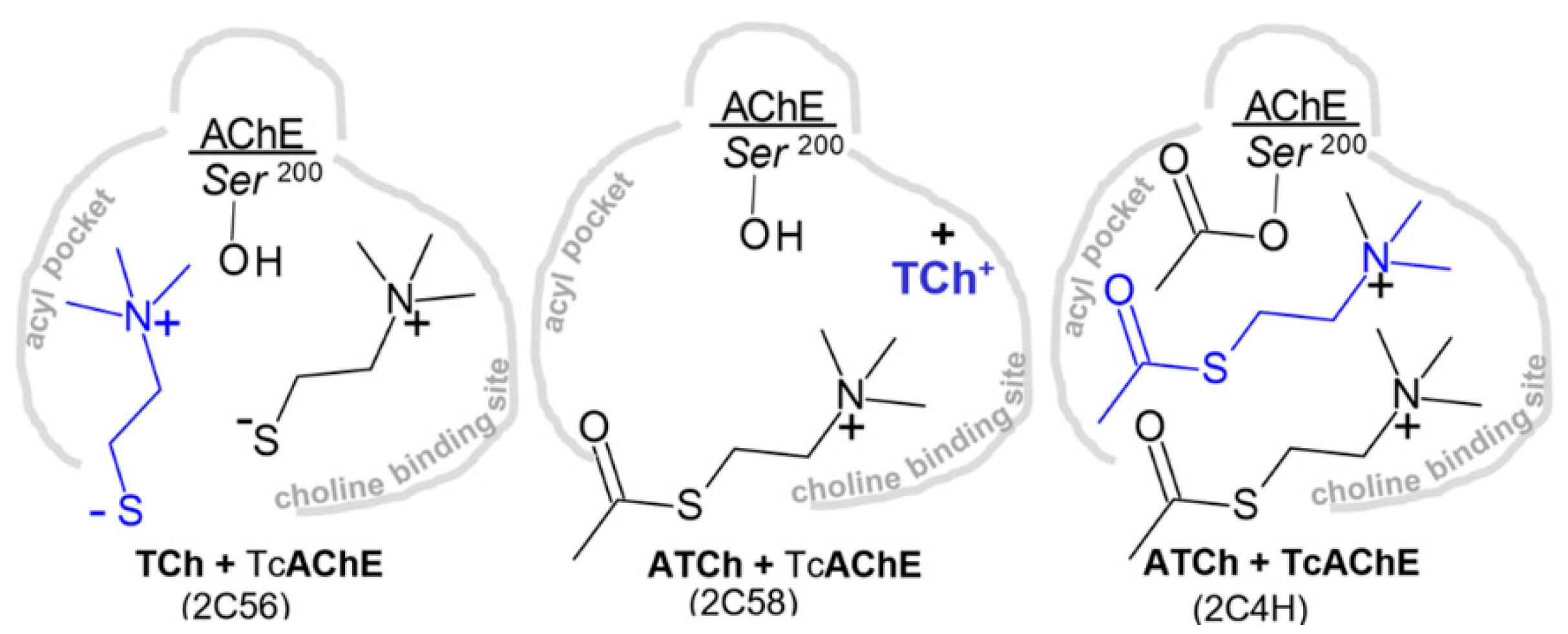
3.1.2. Covalent Conjugates Formed by OPs
3.1.3. Covalent Conjugates Formed by Carbamates
3.2. Effects of Reversible Binding on Backbone Conformation of hAChE
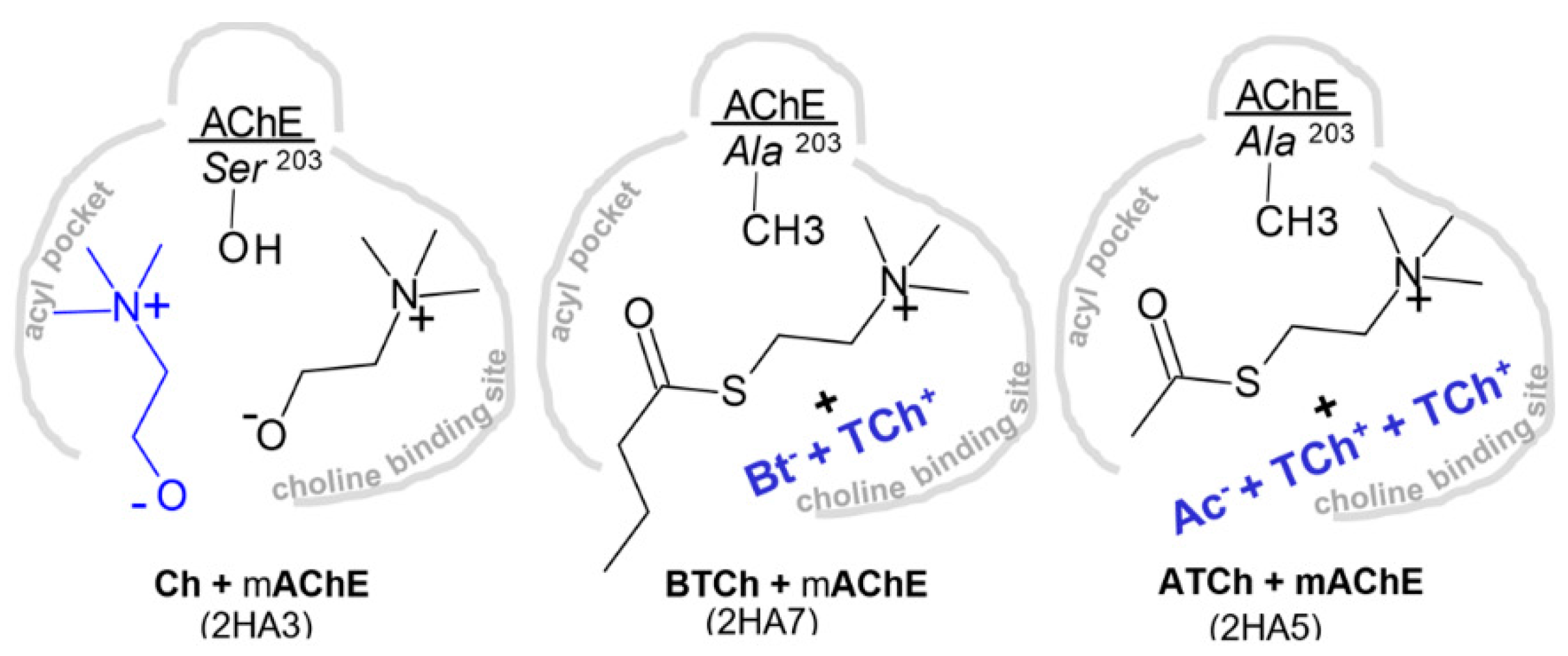
3.2.1. Reversible Binding of Substrates and Products
3.2.2. Reversible Binding of High-Affinity Inhibitors
4. Discussion
Supplementary Materials
Funding
Data Availability Statement
Conflicts of Interest
References
- Quinn, D.M. Acetylcholinesterase: Enzyme structure, reaction dynamics, and virtual transition states. Chem. Rev. 1987, 87, 955–979. [Google Scholar] [CrossRef]
- Sussman, J.L.; Harel, M.; Frolow, F.; Oefner, C.; Goldman, A.; Toker, L.; Silman, I. Atomic structure of acetylcholinesterase from Torpedo californica: A prototypic acetylcholine-binding protein. Science 1991, 253, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Radić, Z.; Taylor, P. Structure and Function of Cholinesterases. In Toxicology of Organophosphate and Carbamate Compounds; Gupta, R.C., Ed.; Elsevier: San Diego, CA, USA, 2006. [Google Scholar]
- Franklin, M.C.; Rudolph, M.J.; Ginter, C.; Cassidy, M.S.; Cheung, J. Structures of paraoxon-inhibited human acetylcholinesterase reveal perturbations of the acyl loop and the dimer interface. Proteins 2016, 84, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; de Sousa, J.; de la Mora, E.; Dias, J.; Jean, L.; Sussman, J.L.; Silman, I.; Renard, P.Y.; Brown, R.; Weik, M.; et al. Structure-Based Optimization of Nonquaternary Reactivators of Acetylcholinesterase Inhibited by Organophosphorus Nerve Agents. J. Med. Chem. 2018, 61, 7630–7639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, Y.; Radić, Z.; Sulzenbacher, G.; Kim, E.; Taylor, P.; Marchot, P. Substrate and product trafficking through the active center gorge of acetylcholinesterase analyzed by crystallography and equilibrium binding. J. Biol. Chem. 2006, 281, 29256–29267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colletier, J.P.; Fournier, D.; Greenblatt, H.M.; Stojan, J.; Sussman, J.L.; Zaccai, G.; Silman, I.; Weik, M. Structural insights into substrate traffic and inhibition in acetylcholinesterase. EMBO J. 2006, 25, 2746–2756. [Google Scholar] [CrossRef] [PubMed]
- Radić, Z.; Pickering, N.A.; Vellom, D.C.; Camp, S.; Taylor, P. Three distinct domains in the cholinesterase molecule confer selectivity for acetyl- and butyrylcholinesterase inhibitors. Biochemistry 1993, 32, 12074–12084. [Google Scholar] [CrossRef] [PubMed]
- Luedtke, S.; Bojo, C.; Li, Y.; Luna, E.; Pomar, B.; Radić, Z. Backbone Conformation Shifts in X-ray Structures of Human Acetylcholinesterase upon Covalent Organophosphate Inhibition. Crystals 2021, 11, 1270. [Google Scholar] [CrossRef]
- Gerlits, O.; Blakeley, M.P.; Keen, D.A.; Radić, Z.; Kovalevsky, A. Room temperature crystallography of human acetylcholinesterase bound to a substrate analogue 4K-TMA: Towards a neutron structure. Curr. Res. Struct. Biol. 2021, 3, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, D.K.; Cheng, X.; Fajer, M.; Ho, K.Y.; Rohrer, J.; Gerlits, O.; Taylor, P.; Juneja, P.; Kovalevsky, A.; Radić, Z. Covalent inhibition of hAChE by organophosphates causes homodimer dissociation through long-range allosteric effects. J. Biol. Chem. 2021, 297, 1–14. [Google Scholar] [CrossRef]
- Rohrer, J.; Sidhom, M.; Han, J.; Radić, Z. Overlay-independent comparisons of X-ray structures reveal small, systematic conformational changes in liganded acetylcholinesterase. Period. Biol. 2016, 118, 319–328. [Google Scholar] [CrossRef] [Green Version]
- wwPDB Consortium. Protein Data Bank: The single global archive for 3D macromolecular structure data. Nucleic Acids Res. 2019, 47, D520–D528. Available online: https://pubmed.ncbi.nlm.nih.gov/30357364/ (accessed on 10 December 2021). [CrossRef] [PubMed] [Green Version]
- Cheung, J.; Rudolph, M.J.; Burshteyn, F.; Cassidy, M.S.; Gary, E.N.; Love, J.; Franklin, M.C.; Height, J.J. Structures of human acetylcholinesterase in complex with pharmacologically important ligands. J. Med. Chem. 2012, 55, 10282–10286. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Wong, L.; Radić, Z.; Tsigelny, I.; Brüggemann, R.; Hosea, N.A.; Berman, H.A. Analysis of cholinesterase inactivation and reactivation by systematic structural modification and enantiomeric selectivity. Chem. Biol. Interact. 1999, 119–120, 3–15. [Google Scholar] [CrossRef]
- Hörnberg, A.; Tunemalm, A.K.; Ekström, F. Crystal structures of acetylcholinesterase in complex with organophosphorus compounds suggest that the acyl pocket modulates the aging reaction by precluding the formation of the trigonal bipyramidal transition state. Biochemistry 2007, 46, 4815–4825. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.G.; Green, L.G.; Grynszpan, F.; Radić, Z.; Carlier, P.R.; Taylor, P.; Finn, M.G.; Sharpless, K.B. Click chemistry in situ: Acetylcholinesterase as a reaction vessel for the selective assembly of a femtomolar inhibitor from an array of building blocks. Angew. Chem. Int. Ed. Engl. 2002, 41, 1053–1057. [Google Scholar] [CrossRef]
- Zha, X.; Lamba, D.; Zhang, L.; Lou, Y.; Xu, C.; Kang, D.; Chen, L.; Xu, Y.; Zhang, L.; De Simone, A.; et al. Novel Tacrine-Benzofuran Hybrids as Potent Multitarget-Directed Ligands for the Treatment of Alzheimer’s Disease: Design, Synthesis, Biological Evaluation, and X-ray Crystallography. J. Med. Chem. 2016, 59, 114–131. [Google Scholar] [CrossRef] [PubMed]
- Baykov, S.V.; Mikherdov, A.S.; Novikov, A.S.; Geyl, K.K.; Tarasenko, M.V.; Gureev, M.A.; Boyarskiy, V.P. π–π Noncovalent Interaction Involving 1,2,4- and 1,3,4-Oxadiazole Systems: The Combined Experimental, Theoretical, and Database Study. Molecules 2021, 26, 5672. [Google Scholar] [CrossRef] [PubMed]
- Kryukova, M.A.; Sapegin, A.V.; Novikov, A.S.; Krasavin, M.; Ivanov, D.M. New Crystal Forms for Biologically Active Compounds. Part 1: Noncovalent Interactions in Adducts of Nevirapine with XB Donors. Crystals 2019, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Rydberg, E.H.; Brumshtein, B.; Greenblatt, H.M.; Wong, D.M.; Shaya, D.; Williams, L.D.; Carlier, P.R.; Pang, Y.P.; Silman, I.; Sussman, J.L. Complexes of alkylene-linked tacrine dimers with Torpedo californica acetylcholinesterase: Binding of Bis5-tacrine produces a dramatic rearrangement in the active-site gorge. J. Med. Chem. 2006, 49, 5491–5500. [Google Scholar] [CrossRef] [PubMed]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radić, Z. Shifts in Backbone Conformation of Acetylcholinesterases upon Binding of Covalent Inhibitors, Reversible Ligands and Substrates. Crystals 2021, 11, 1557. https://doi.org/10.3390/cryst11121557
Radić Z. Shifts in Backbone Conformation of Acetylcholinesterases upon Binding of Covalent Inhibitors, Reversible Ligands and Substrates. Crystals. 2021; 11(12):1557. https://doi.org/10.3390/cryst11121557
Chicago/Turabian StyleRadić, Zoran. 2021. "Shifts in Backbone Conformation of Acetylcholinesterases upon Binding of Covalent Inhibitors, Reversible Ligands and Substrates" Crystals 11, no. 12: 1557. https://doi.org/10.3390/cryst11121557