
Wnt Lipidation and Modifiers in Intestinal Carcinogenesis and Cancer

Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
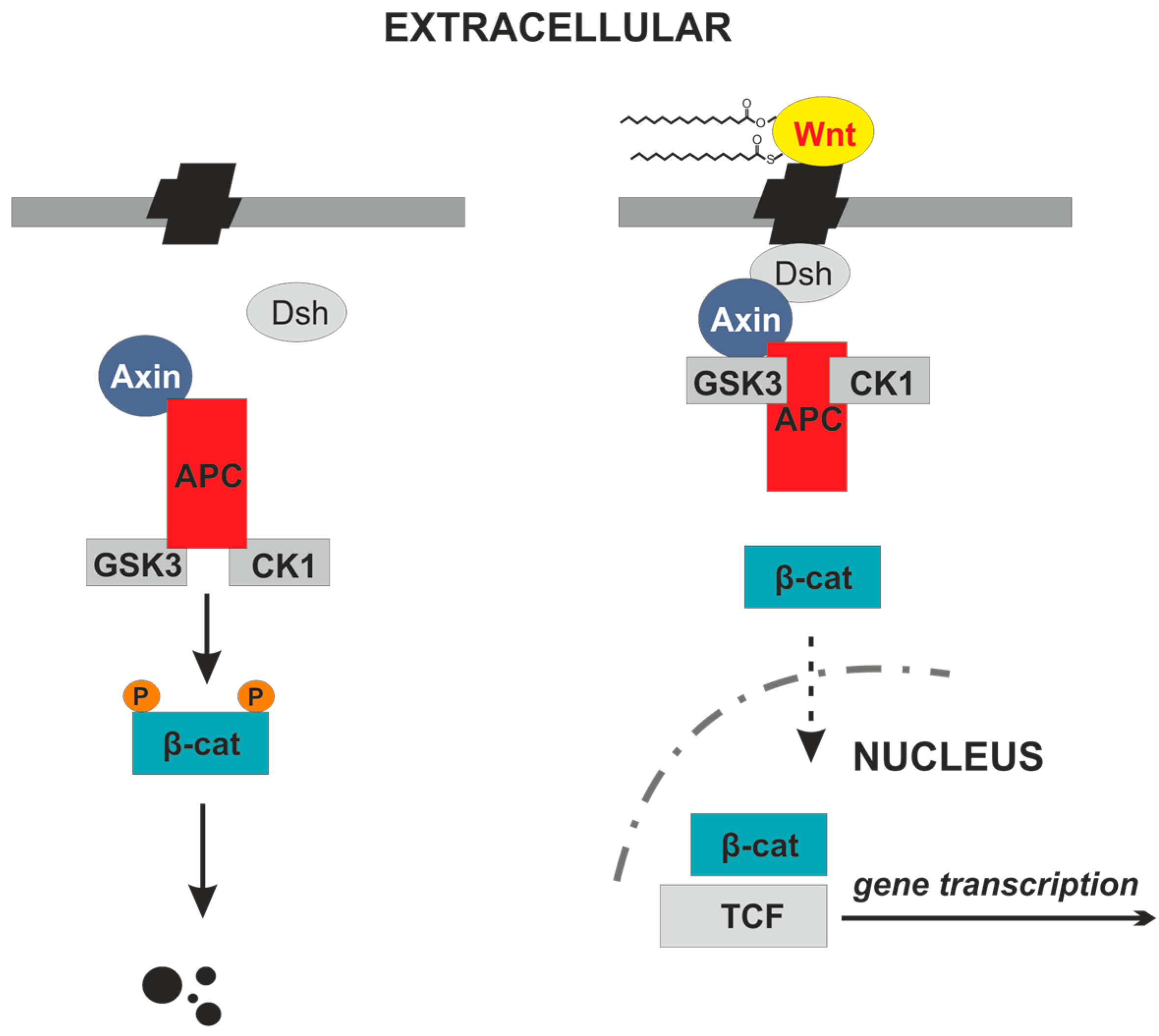
2. Wnt Proteins
3. Wnt Lipidation Is Essential for Secretion and Signaling Activity
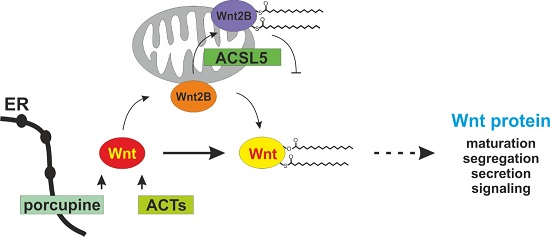
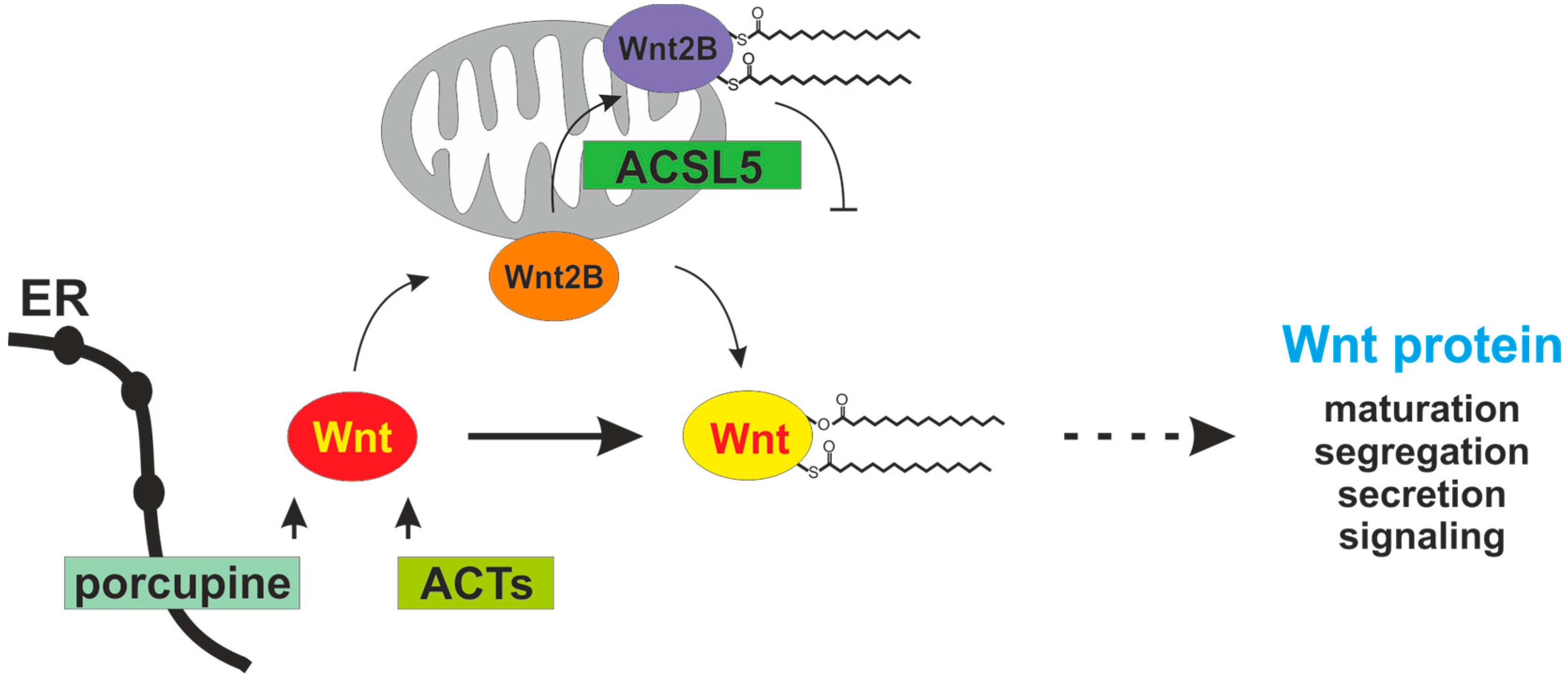
4. Porcupine and ACSL5 as Modifiers of Wnt Proteins
5. Wnt Lipidation and Signaling in Intestinal Carcinogenesis and Cancer
6. Therapeutic Modifiers of Wnt Lipidation
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alexandre, C.; Baena-Lopez, A.; Vincent, J.-P. Patterning and growth control by membrane-tethered Wingless. Nature 2014, 505, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. The intestinal crypt, a prototype stem cell compartment. Cell 2013, 154, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.-A.L.; Huang, S. Crosstalk of the Wnt/β-catenin pathway with other pathways in cancer cells. Genes Dis. 2016, 3, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; Kishida, S.; Yamamoto, H.; Murai, H.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the Wnt signaling pathway, forms a complex with GSK-3beta and beta-catenin and promotes GSK-3beta-dependent phosphorylation of beta-catenin. EMBO J. 1998, 17, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. Wnt signaling and cancer. Genes Dev. 2000, 14, 1837–1851. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.; Hawke, D.; Zheng, Y.; Xia, Y.; Meisenhelder, J.; Nika, H.; Mills, G.B.; Kobayashi, R.; Hunter, T.; Lu, Z. Phosphorylation of beta-catenin by AKT promotes beta-catenin transcriptional activity. J. Biol. Chem. 2007, 282, 11221–11229. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Ghosh, S.; Wang, Z.; Hunter, T. Downregulation of caveolin-1 function by EGF leads to the loss of E-cadherin, increased transcriptional activity of beta-catenin, and enhanced tumor cell invasion. Cancer Cell 2003, 4, 499–515. [Google Scholar] [CrossRef]
- Flentke, G.R.; Garic, A.; Hernandez, M.; Smith, S.M. CaMKII represses transcriptionally active β-catenin to mediate acute ethanol neurodegeneration and can phosphorylate β-catenin. J. Neurochem. 2014, 128, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Janda, C.Y.; Waghray, D.; Levin, A.M.; Thomas, C.; Garcia, K.C. Structural basis of Wnt recognition by frizzled. Science 2012, 337, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Moon, R.T.; Kohn, A.D.; de Ferrari, G.V.; Kaykas, A. Wnt and β-catenin signaling: Diseases and therapies. Nat. Rev. Genet. 2004, 5, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R. Cell signaling: Disarming Wnt. Nature 2015, 519, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Chi, S.; Xue, J.; Yang, J.; Li, F.; Liu, X. Emerging role and therapeutic implication of Wnt signaling pathways in autoimmune disease. J. Immunol. Res. 2016, 2016, 9392132. [Google Scholar] [CrossRef] [PubMed]
- Tury, A.; Tolentino, K.; Zou, Y. Altered expression of atypical RKC and Ryk in the spinal cord of a mouse model of amyotrophic lateral sclerosis. Dev. Neurobiol. 2014, 74, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; He, H.; Lv, R.; Zhang, M.; Huang, H.; An, Z.; Li, S. Preliminary mechanism on the methylation modification of Dkk-1 and Dkk-3 in hepatocellular carcinoma. Tumor Biol. 2015, 36, 1245–1250. [Google Scholar] [CrossRef] [PubMed]
- Gori, F.; Lerner, U.; Ohlsson, C.; Baron, R. A new WNT on the bone: WNT16, cortical bone thickness, porosity and fractures. BoneKEy Rep. 2015, 4, 669. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.R. The Wnts. Genome Biol. 2001, 3. [Google Scholar] [CrossRef]
- Coudreuse, D.; Korswagen, H.C. The making of Wnt: New insights into Wnt maturation, sorting and secretion. Development 2007, 134, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Kitagawa, Y.; Kadowaki, T. Drosophila segment polarity gene product porcupine stimulates the posttranslational N-glycosylation of wingless in the endoplasmic reticulum. J. Biol. Chem. 2002, 277, 12816–12823. [Google Scholar] [CrossRef] [PubMed]
- Gething, M.J.; Sambrook, J. Transport and assembly processes in the endoplasmic reticulum. Sem. Cell Biol. 1990, 1, 65–72. [Google Scholar]
- Burrus, L.W.; McMahon, A.P. Biochemical analysis of murine Wnt proteins reveals both shared and distinct properties. Exp. Cell Res. 1995, 220, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Port, F.; Basler, K. Wnt trafficking: New insights into Wnt maturation, secretion and spreading. Traffic 2010, 11, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Willert, K.; Brown, J.D.; Danenberg, E.; Duncan, A.W.; Weissman, I.L.; Reya, T.; Yates, J.R., 3rd; Nusse, R. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature 2003, 423, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Kurayoshi, M.; Yamamoto, H.; Izumi, S.; Kikuchi, A. Post-translational palmitoylation and glycosylation of Wnt-5a are necessary for its signalling. Biochem. J. 2007, 402, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Takada, R.; Satomi, Y.; Kurata, T.; Ueno, N.; Norioka, S.; Kondoh, H.; Takao, T.; Takada, S. Monounsaturated fatty acid modification of Wnt protein: Its role in Wnt secretion. Dev. Cell 2006, 11, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Bazan, J.F.; de Sauvage, F.J. Structural ties between cholesterol transport and morphogen signaling. Cell 2009, 138, 1055–1056. [Google Scholar] [CrossRef] [PubMed]
- Malinauskas, T.; Ariescu, A.R.; Lu, W.; Siebold, C.; Jones, E.Y. Modular mechanism of Wnt signaling inhibition by Wnt inhibiting factor 1. Nat. Struct. Mol. Biol. 2011, 18, 886. [Google Scholar] [CrossRef] [PubMed]
- Greco, V.; Hannus, M.; Eaton, S. Argosomes: A potential vehicle for the spread of morphogens through epithelia. Cell 2001, 106, 633–645. [Google Scholar] [CrossRef]
- Bartscherer, K.; Boutros, M. Regulation of Wnt protein secretion and its role in gradient formation. EMBO Rep. 2008, 9, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Farin, H.F.; Jordens, I.; Mosa, M.H.; Basak, O.; Korving, J.; Tauriello, D.V.F.; de Punder, K.; Angers, S.; Peters, P.J.; Maurice, M.M.; et al. Visualization of a short-range Wnt gradient in the intestinal stem-cell niche. Nature 2016, 530, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Wilder, E.; Klingensmith, J.; Zachary, K.; Perrimon, N. The segment polarity gene porcupine encodes a putative multimembrane protein involved in Wingless processing. Genes Dev. 1996, 10, 3116–3128. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K. A superfamily of membrane-bound O-acyltransferases with implications for Wnt signaling. Trends Biochem. Sci. 2000, 25, 111–112. [Google Scholar] [CrossRef]
- Galli, L.M.; Burrus, L.W. Differential palmit(e)oylation of Wnt1 on C93 and S224 residues has overlapping and distinct consequences. PLoS ONE 2011, 6, e26636. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Chaturvedi, D.; Cumberledge, S. Drosophila Wnt-1 undergoes a hydrophobic modification and is targeted to lipid rafts, a process that requires porcupine. J. Biol. Chem. 2004, 279, 33220–33227. [Google Scholar] [CrossRef] [PubMed]
- Tuladhar, R.; Lum, L. Fatty acyl donor selectivity in membrane bound O-acyltransferases and communal cell fate decision-making. Biochem. Soc. Trans. 2015, 43, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Gan, Q.F.; Golan-Lev, T.; Arora, P.; Yanuka, O.; Oren, Y.S.; Leikin-Frenkel, A.; Graf, M.; Garippa, R.; Boehringer, M.; et al. Selective elimination of human pluripotent stem cells by an oleate synthesis inhibitor discovered in a high-throughput screen. Cell Stem Cell 2013, 12, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.; Harryman-Samos, C.; Klingensmith, J.; Perrimon, N.; Nusse, R. Mutations in the segment polarity genes wingless and porcupine impair secretion of the wingless protein. EMBO J. 1993, 12, 5293–5302. [Google Scholar] [PubMed]
- Biechele, S.; Cox, B.J.; Rossant, J. Porcupine homolog is required for canonical Wnt signaling and gastrulation in mouse embryos. Dev. Biol. 2011, 355, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.H.; Seaton, M.S.; Wallace, J.; Al-Harthi, L. Porcupine is not required for the production of the majority of Wnts from primary human astrocytes and CD8+ T cells. PLoS ONE 2014, 9, e92159. [Google Scholar] [CrossRef] [PubMed]
- Klaus, C.; Schneider, U.; Hedberg, C.; Schütz, A.K.; Bernhagen, J.; Waldmann, H.; Gassler, N.; Kaemmerer, E. Modulating effects of acyl-CoA synthetase 5-derived mitochondrial Wnt2B palmitoylation on intestinal Wnt activity. World J. Gastroenterol. 2014, 20, 14855–14864. [Google Scholar] [CrossRef] [PubMed]
- Gassler, N.; Roth, W.; Funke, B.; Schneider, A.; Herzog, F.; Tischendorf, J.J.W.; Grund, K.; Penzel, R.; Bravo, I.G.; Mariadason, J.; et al. Regulation of enterocyte apoptosis by acyl-CoA synthetase 5 splicing. Gastroenterology 2007, 133, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Meller, N.; Morgan, M.E.; Wong, W.P.S.; Altemus, J.B.; Sehayek, E. Targeting of acyl-CoA synthetase 5 decreases jejunal fatty acid activation with no effect on dietary long-chain fatty acid absorption. Lipids Health Dis. 2013, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- Klaus, C.; Kaemmerer, E.; Reinartz, A.; Schneider, U.; Plum, P.; Jeon, M.K.; Hose, J.; Hartmann, F.; Schnölzer, M.; Wagner, N.; et al. TP53 status regulates ACSL5-induced expression of mitochondrial mortalin in enterocytes and colorectal adenocarcinomas. Cell Tissue Res. 2014, 357, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Kirikoshi, H.; Terasaki, H.; Shiokawa, K. WNT2B2 mRNA, up-regulated in primary gastric cancer, is a positive regulator of the WNT-beta-catenin-TCF signaling pathway. Biochem. Biophys. Res. Commun. 2001, 289, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Struewing, I.T.; Tobotrek, A.; Mao, C.D. Mitochondrial and nuclear forms of Wnt13 are generated via alternative promoters, alternative RNA splicing, and alternative translation start sites. J. Biol. Chem. 2006, 281, 7282–7293. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; Ridgway, R.A.; van Es, J.H.; van de Wetering, M.; Begthel, H.; van den Born, M.; Danenberg, E.; Clarke, A.R.; Sansom, O.J.; Clevers, H. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 2009, 457, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, L.; de Sousa, E.M.; van der Heijden, M.; Cameron, K.; de Jong, J.H.; Borovski, T.; Tuynman, J.B.; Todaro, M.; Merz, C.; Rodermond, H.; et al. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat. Cell Biol. 2010, 12, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Horst, D.; Chen, J.; Morikawa, T.; Ogino, S.; Kirchner, T.; Shivdasani, R.A. Differential WNT activity in colorectal cancer confers limited tumorigenic potential and is regulated by MAPK signaling. Cancer Res. 2012, 72, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Weinberg, R.A. Epithelial-mesenchymal plasticity: A central regulator of cancer progression. Trends Cell Biol. 2015, 25, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Uchida, H.; Yamazaki, K.; Fukuma, M.; Yamada, T.; Hayashida, T.; Hasegawa, H.; Kitajima, M.; Kitagawa, Y.; Sakamoto, M. Overexpression of leucine-rich repeat-containing G protein-coupled receptor 5 in colorectal cancer. Cancer Sci. 2010, 101, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, K.M.; Spurlock, M.E. Palmitate activates the NF-kappaB transcription factor and induces IL-6 and TNF alpha expression in 3T3-L1 adipocytes. J. Nutr. 2005, 135, 1841–1846. [Google Scholar] [PubMed]
- Kaemmerer, E.; Peuscher, A.; Reinartz, A.; Liedtke, C.; Weiskirchen, R.; Kopitz, J.; Gassler, N. Human intestinal acyl-CoA synthetase 5 is sensitive to the inhibitor triacsin C. World J. Gastroenterol. 2011, 17, 4883–4889. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Varga, J.; Wang, T.C.; Greten, F.R. The gastrointestinal tumor microenvironment. Gastroenterology 2013, 145, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Myant, K.B.; Cammareri, P.; McGhee, E.J.; Ridgway, R.A.; Huels, D.J.; Cordero, J.B.; Schwitalla, S.; Kalna, G.; Ogg, E.L.; Athineos, D.; et al. ROS production and NF-κB activation triggered by RAC1 facilitate Wnt-driven intestinal stem cell proliferation and colorectal cancer initiation. Cell Stem Cell 2013, 12, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Gassler, N.; Herr, I.; Schneider, A.; Penzel, R.; Langbein, L.; Schirmacher, P.; Kopitz, J. Impaired expression of acyl-CoA synthetase 5 in sporadic colorectal adenocarcinomas. J. Pathol. 2005, 207, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Frequent up-regulation of WNT2 in primary gastric cancer and colorectal cancer. Int. J. Oncol. 2001, 19, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Resh, M.D. Targeting protein lipidation in disease. Trends Mol. Med. 2012, 18, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, Y.; He, B.; Gong, Y. Development of small molecules targeting the Wnt signaling pathway in cancer stem cells for the treatment of colorectal cancer. Clin. Colorectal Cancer 2015, 14, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Noto, A.; Raffa, S.; de Vitis, C.; Roscilli, G.; Malpicci, D.; Coluccia, P.; di Napoli, A.; Ricci, A.; Giovagnoli, M.R.; Aurisicchio, L.; et al. Stearoyl-CoA desaturase-1 is a key factor for lung cancer-initiating cells. Cell Death Dis. 2013, 4, e947. [Google Scholar] [CrossRef] [PubMed]
- Herr, P.; Basler, K. Porcupine-mediated lipidation is required for Wnt recognition by WIs. Dev. Biol. 2012, 361, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covey, T.M.; Kaur, S.; Ong, T.T.; Proffitt, K.D.; Wu, Y.; Tan, P.; Virshup, D.M. PORCN moonlights in a Wnt-independent pathway that regulates cancer cell proliferation. PLoS ONE 2012, 7, e34532. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of porcupine by LGK974. Proc. Natl. Acad. Sci. USA 2013, 110, 20224–20229. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Heal, W.P.; Mann, D.J.; Tate, E.W. Protein myristoylation in health and disease. J. Chem. Biol. 2010, 3, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Frearson, J.A.; Brand, S.; McElroy, S.P.; Cleghom, L.A.; Smid, O.; Stojanovski, L.; Price, H.P.; Guther, M.L.; Torrie, L.S.; Robinson, D.A.; et al. N-myristoyltransferase inhibitors as new leads to treat sleeping sickness. Nature 2010, 464, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.G.; Frost, D.; Meta, M.; Qiao, X.; Yang, S.H.; Coffinier, C.; Young, S.G. A protein farnesyltransferase inhibitor ameliorates disease in a mouse model of progeria. Science 2006, 311, 1621–1623. [Google Scholar] [CrossRef] [PubMed]
- Ferri, N.; Paoletti, R.; Corsini, A. Lipid-modified proteins as biomarkers for cardiovascular disease: A review. Biomarkers 2005, 10, 219–237. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaemmerer, E.; Gassler, N. Wnt Lipidation and Modifiers in Intestinal Carcinogenesis and Cancer. Cancers 2016, 8, 69. https://doi.org/10.3390/cancers8070069
Kaemmerer E, Gassler N. Wnt Lipidation and Modifiers in Intestinal Carcinogenesis and Cancer. Cancers. 2016; 8(7):69. https://doi.org/10.3390/cancers8070069
Chicago/Turabian StyleKaemmerer, Elke, and Nikolaus Gassler. 2016. "Wnt Lipidation and Modifiers in Intestinal Carcinogenesis and Cancer" Cancers 8, no. 7: 69. https://doi.org/10.3390/cancers8070069