Peroxisome Proliferator-Activated Receptor and Vitamin D Receptor Signaling Pathways in Cancer Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
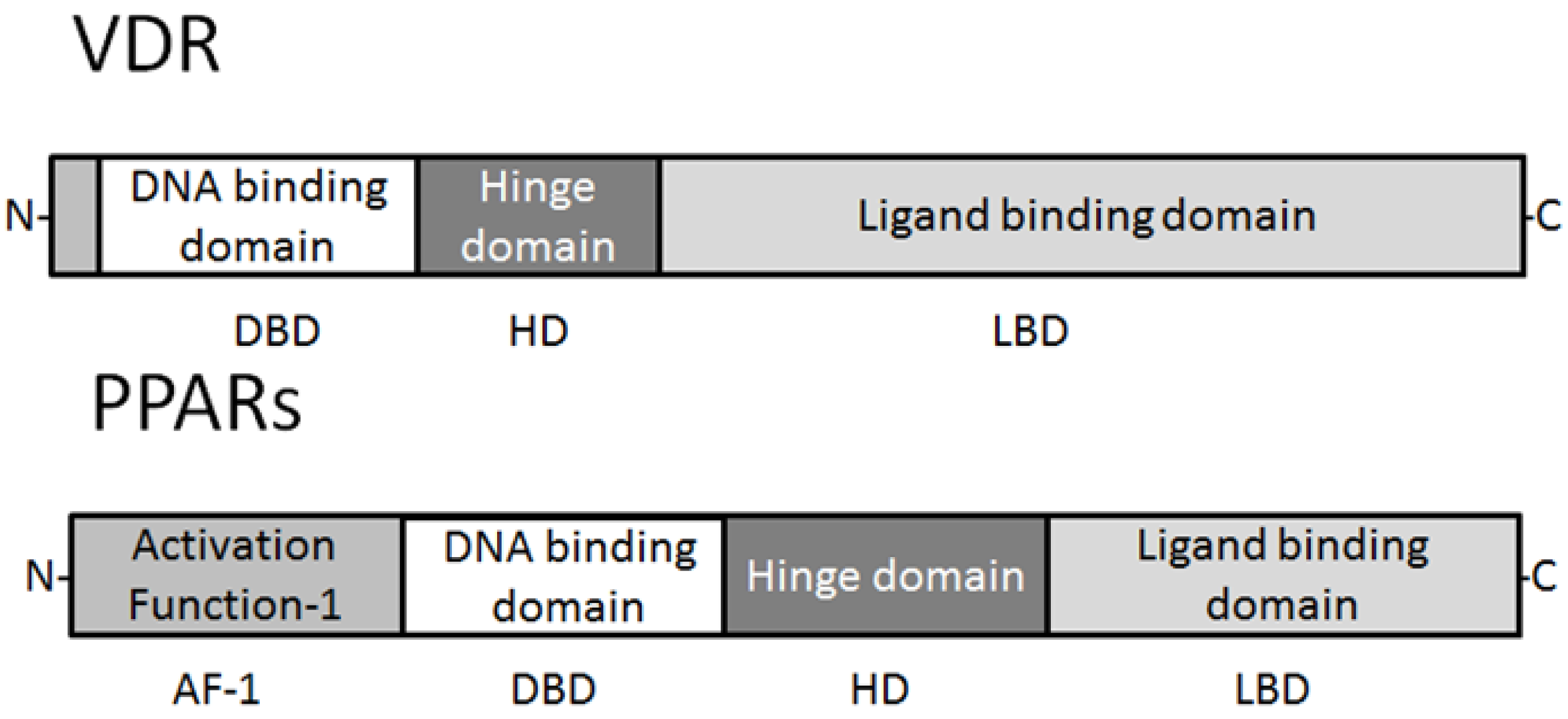
2. Expression and Characteristics of PPARs and Vitamin D Receptor


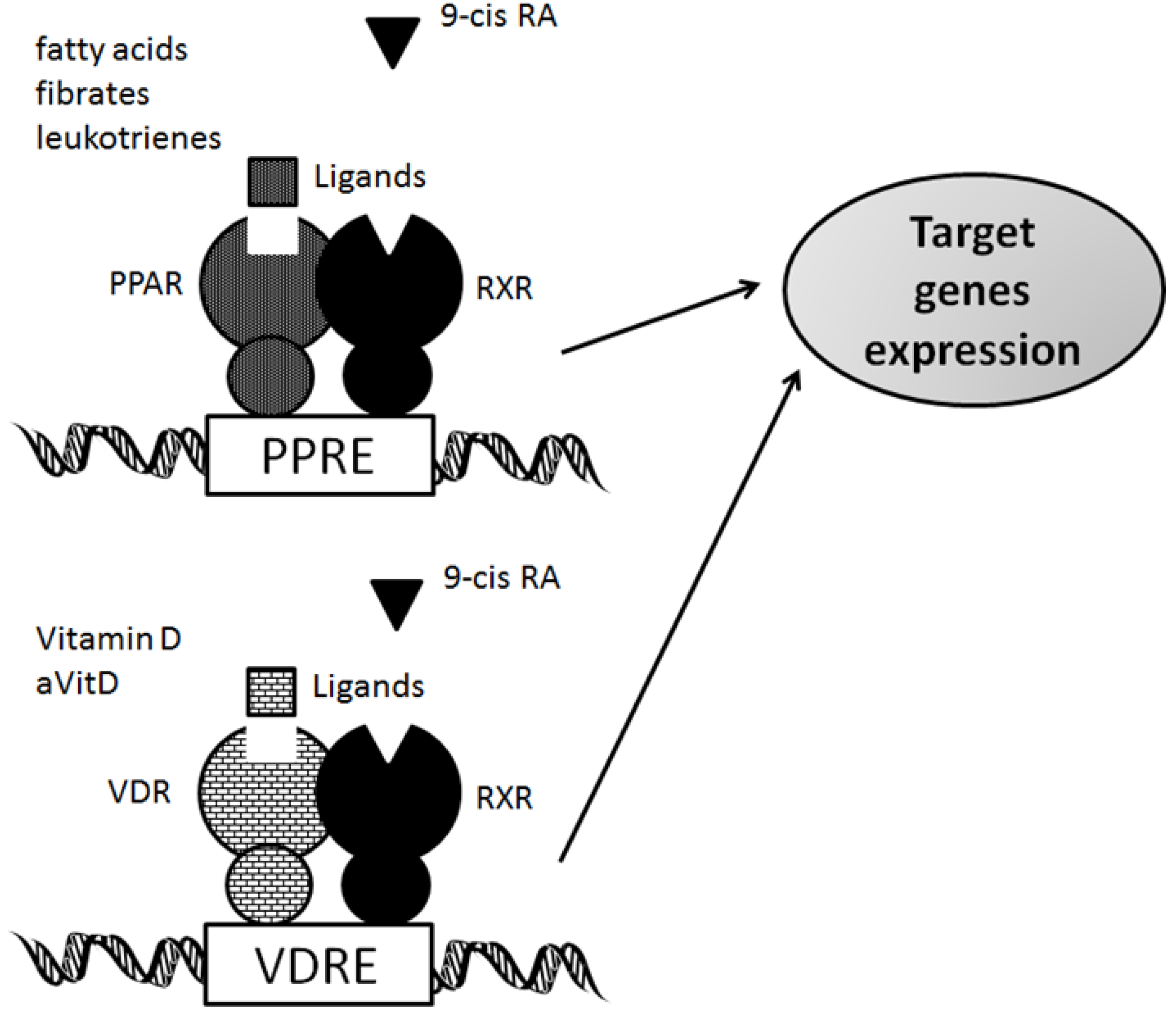
3. Functional Interplay of Vitamin D Receptor with PPARs
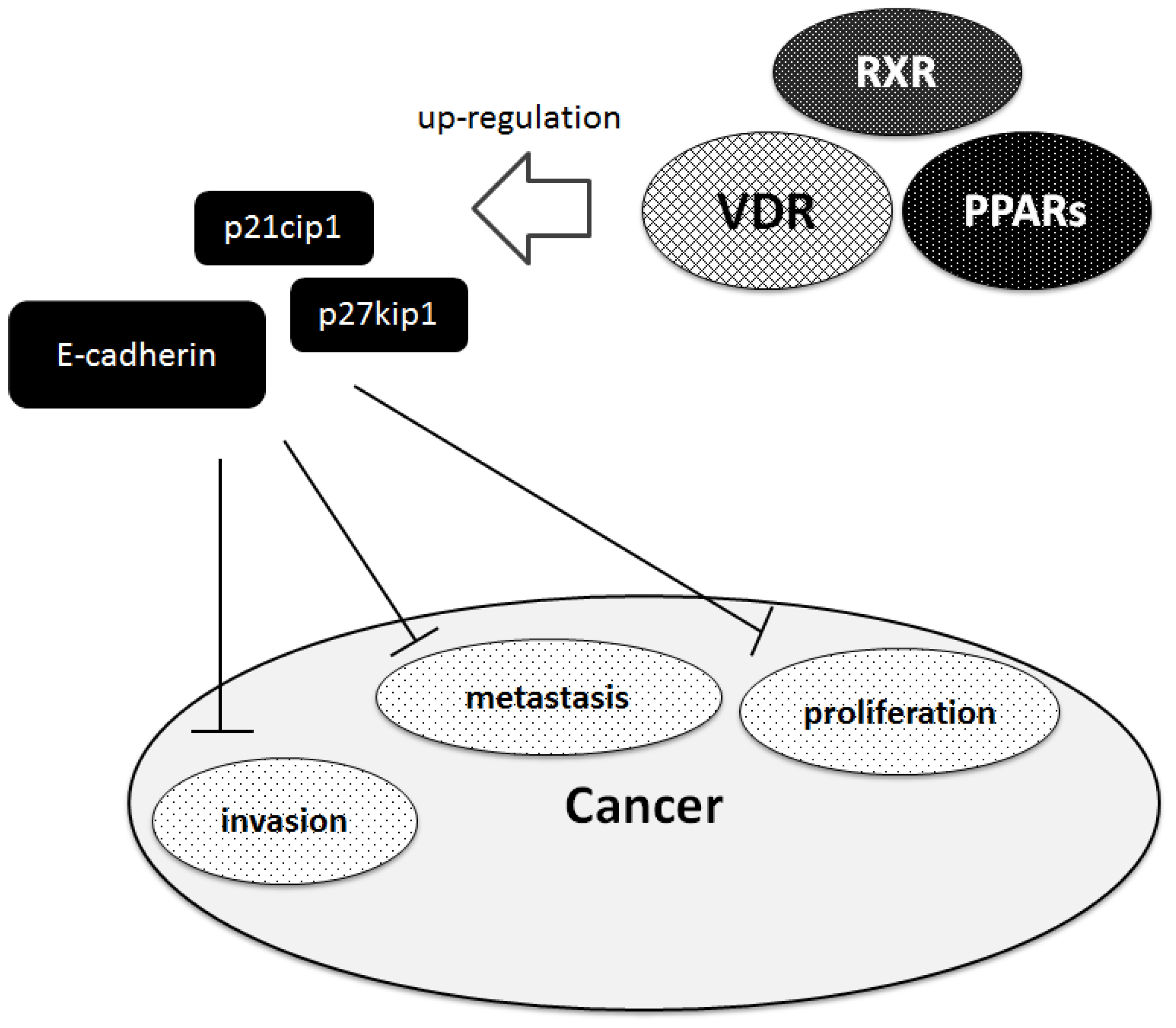
4. PPARs and Vitamin D Receptor in Cancer

5. Perspectives
6. Conclusions
Abbreviations
| aVitD | active form of vitamin D: 1,25(OH)2D3 |
| AP-1 | activator protein-1 |
| EMT | mesenchymal transition |
| mRNA | messenger RNA |
| PPAR | peroxisome proliferator-activated receptor |
| PPRE | PPAR response element |
| RA | retinoic acid |
| RARs | retinoic acid receptors |
| RE | response element |
| RXR | retinoid X receptor |
| TR | thyroid receptor |
| VDR | vitamin D receptor |
| VDREs | vitamin D response elements |
Acknowledgments
Conflicts of Interest
References
- Hu, X.; Funder, J.W. The evolution of mineralocorticoid receptors. Mol. Endocrinol. 2006, 20, 1471–1478. [Google Scholar]
- Pérez, E.; Bourguet, W.; Gronemeyer, H.; de Lera, A.R. Modulation of RXR function through ligand design. Biochim. Biophys. Acta 2012, 1821, 57–69. [Google Scholar] [CrossRef]
- Carlberg, C.; Dunlop, T.W. An integrated biological approach to nuclear receptor signaling in physiological control and disease. Crit. Rev. Eukaryot. Gene Expr. 2006, 16, 1–22. [Google Scholar] [CrossRef]
- Ditsch, N.; Mayr, D.; Lenhard, M.; Strauss, C.; Vodermaier, A.; Gallwas, J.; Stoeckl, D.; Graeser, M.; Weissenbacher, T.; Friese, K.; Jeschke, U. Correlation of thyroid hormone, retinoid X, peroxisome proliferator-activated, vitamin D and oestrogen/progesterone receptors in breast carcinoma. Oncol. Lett. 2012, 4, 665–671. [Google Scholar]
- Schulman, I.G. Nuclear receptors as drug targets for metabolic disease. Adv. Drug Deliv. Rev. 2010, 62, 1307–1315. [Google Scholar] [CrossRef]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef]
- Becker, J.; Delayre-Orthez, C.; Frossard, N.; Pons, F. Regulation of inflammation by PPARs: A future approach to treat lung inflammatory diseases? Fundam. Clin. Pharmacol. 2006, 20, 429–447. [Google Scholar] [CrossRef]
- Ringseis, R.; Eder, K. Influence of pharmacological PPARalpha activators on carnitine homeostasis in proliferating and non-proliferating species. Pharmacol. Res. 2009, 60, 179–184. [Google Scholar] [CrossRef]
- Wagner, K.D.; Wagner, N. Peroxisome proliferator-activated receptor beta/delta (PPARbeta/delta) acts as regulator of metabolism linked to multiple cellular functions. Pharmacol. Ther. 2010, 125, 423–435. [Google Scholar] [CrossRef]
- Cipolletta, D.; Feuerer, M.; Li, A.; Kamei, N.; Lee, J.; Shoelson, S.E.; Benoist, C.; Mathis, D. PPAR-γ is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature 2012, 486, 549–553. [Google Scholar]
- Giaginis, C.; Tsantili-Kakoulidou, A.; Theocharis, S. Peroxisome proliferator-activated receptors (PPARs) in the control of bone metabolism. Fundam. Clin. Pharmacol. 2007, 21, 231–244. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, X.; Chen, L.; Chen, J.; Hu, L.; Jiang, H.; Shen, X. Molecular determinants of magnolol targeting both RXRα and PPARγ. PLoS One 2011, 6, e28253. [Google Scholar]
- Temple, K.A.; Cohen, R.N.; Wondisford, S.R.; Yu, C.; Deplewski, D.; Wondisford, F.E. An intact DNA-binding domain is not required for peroxisome proliferator-activated receptor gamma (PPARgamma) binding and activation on some PPAR response elements. J. Biol. Chem. 2005, 280, 3529–3540. [Google Scholar]
- Clarke, S.D.; Thuillier, P.; Baillie, R.A.; Sha, X. Peroxisome proliferator-activated receptors: A family of lipid-activated transcription factors. Am. J. Clin. Nutr. 1999, 70, 566–571. [Google Scholar]
- Nezbedova, P.; Brtko, J. 1alpha,25-dihydroxyvitamin D3 inducible transcription factor and its role in the vitamin D action. Endocr. Regul. 2004, 38, 29–38. [Google Scholar]
- Wolf, G. Is 9-cis-retinoic acid the endogenous ligand for the retinoic acid-X receptor? Nutr. Rev. 2006, 64, 532–538. [Google Scholar]
- Benz, V.; Kintscher, U.; Foryst-Ludwig, A. Sex-specific differences in type 2 diabetes mellitus and dyslipidemia therapy: PPAR agonists. Handb. Exp. Pharmacol. 2012, 214, 387–410. [Google Scholar]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef]
- Morosetti, R.; Servidei, T.; Mirabella, M.; Rutella, S.; Mangiola, A.; Maira, G.; Mastrangelo, R.; Koeffler, H.P. The PPARgamma ligands PGJ2 and rosiglitazone show a differential ability to inhibit proliferation and to induce apoptosis and differentiation of human glioblastoma cell lines. Int. J. Oncol. 2004, 25, 493–502. [Google Scholar]
- Yu, Z.; Schneider, C.; Boeglin, W.E.; Brash, A.R. Epidermal lipoxygenase products of the hepoxilin pathway selectively activate the nuclear receptor PPARalpha. Lipids 2007, 42, 491–497. [Google Scholar] [CrossRef]
- Kouroumichakis, I.; Papanas, N.; Zarogoulidis, P.; Liakopoulos, V.; Maltezos, E.; Mikhailidis, D.P. Fibrates: Therapeutic potential for diabetic nephropathy? Eur. J. Intern. Med. 2012, 23, 309–316. [Google Scholar] [CrossRef]
- Friedland, S.N.; Leong, A.; Filion, K.B.; Genest, J.; Lega, I.C.; Mottillo, S.; Poirier, P.; Reoch, J.; Eisenberg, M.J. The cardiovascular effects of peroxisome proliferator-activated receptor agonists. Am. J. Med. 2012, 125, 126–133. [Google Scholar] [CrossRef]
- Ibabe, A.; Herrero, A.; Cajaraville, M.P. Modulation of peroxisome proliferator-activated receptors (PPARs) by PPAR(alpha)- and PPAR(gamma)-specific ligands and by 17beta-estradiol in isolated zebrafish hepatocytes. Toxicol. In Vitro 2005, 19, 725–735. [Google Scholar] [CrossRef]
- Baker, P.R.; Lin, Y.; Schopfer, F.J.; Woodcock, S.R.; Groeger, A.L.; Batthyany, C.; Sweeney, S.; Long, M.H.; Iles, K.E.; Baker, L.M.; et al. Fatty acid transduction of nitric oxide signaling: multiple nitrated unsaturated fatty acid derivatives exist in human blood and urine and serve as endogenous peroxisome proliferator-activated receptor ligands. Biol. Chem. 2005, 280, 42464–42475. [Google Scholar] [CrossRef]
- Yu, S.; Reddy, J.K. Transcription coactivators for peroxisome proliferator-activated receptors. Biochim. Biophys. Acta 2007, 1771, 936–951. [Google Scholar]
- Waku, T.; Shiraki, T.; Oyama, T.; Morikawa, K. Atomic structure of mutant PPARgamma LBD complexed with 15d-PGJ2: Novel modulation mechanism of PPARgamma/RXRalpha function by covalently bound ligands. FEBS Lett. 2009, 583, 320–324. [Google Scholar] [CrossRef]
- Hansen, C.M.; Binderup, L.; Hamberg, K.J.; Carlberg, C. Vitamin D and cancer: Effects of 1,25(OH)2D3 and its analogs on growth control and tumorigenesis. Front. Biosci. 2001, 6, D820–D848. [Google Scholar] [CrossRef]
- Ebert, R.; Schütze, N.; Adamski, J.; Jakob, F. Vitamin D signaling is modulated on multiple levels in health and disease. Mol. Cell Endocrinol. 2006, 248, 149–159. [Google Scholar] [CrossRef]
- Jensen, T.J.; Henriksen, L.O.; Sølvsten, H.; Kragballe, K. Inhibition of the 1,25-dihydroxyvitamin D3-induced increase in vitamin D receptor (VDR) levels and binding of VDR-retinoid X receptor (RXR) to a direct repeat (DR)-3 type response element by an RXR-specific ligand in human keratinocyte cultures. Biochem. Pharmacol. 1998, 55, 767–773. [Google Scholar] [CrossRef]
- Shirazi, L.; Almquist, M.; Malm, J.; Wirfält, E.; Manjer, J. Determinants of serum levels of vitamin D: a study of life-style, menopausal status, dietary intake, serum calcium, and PTH. BMC Womens Health 2013. [Google Scholar] [CrossRef]
- Wacker, M.; Holick, M.F. Vitamin D—Effects on skeletal and extraskeletal health and the need for supplementation. Nutrients 2013, 5, 111–148. [Google Scholar] [CrossRef]
- Malley, R.C.; Muller, H.K.; Norval, M.; Woods, G.M. Dietary vitamin D alters the response of the skin to UVB-irradiation depending on the genetic background of the mice. Photochem. Photobiol. Sci. 2013, 12, 536–545. [Google Scholar]
- Luo, W.; Hershberger, P.A.; Trump, D.L.; Johnson, C.S. 24-Hydroxylase in cancer: Impact on vitamin D-based anticancer therapeutics. J. Steroid Biochem. Mol. Biol. 2013, 136, 252–257. [Google Scholar]
- Deeb, K.K.; Trump, D.L.; Johnson, C.S. Vitamin D signalling pathways in cancer: Potential for anticancer therapeutics. Nat. Rev. Cancer 2007, 7, 684–700. [Google Scholar] [CrossRef]
- Wood, R.J. Vitamin D and adipogenesis: New molecular insights. Nutr. Rev. 2008, 66, 40–46. [Google Scholar] [CrossRef]
- Mulholland, D.J.; Dedhar, S.; Coetzee, G.A.; Nelson, C.C. Interaction of nuclear receptors with the Wnt/beta-catenin/Tcf signaling axis: Wnt you like to know? Endocr. Rev. 2005, 26, 898–915. [Google Scholar] [CrossRef]
- Narvaez, C.J.; Simmons, K.M.; Brunton, J.; Salinero, A.; Chittur, S.V.; Welsh, J.E. Induction of STEAP4 correlates with 1,25-dihydroxyvitamin D3 stimulation of adipogenesis in mesenchymal progenitor cells derived from human adipose tissue. J. Cell Physiol. 2013, 228, 2024–2036. [Google Scholar] [CrossRef]
- Dunlop, T.W.; Väisänen, S.; Frank, C.; Molnár, F.; Sinkkonen, L.; Carlberg, C. The human peroxisome proliferator-activated receptor delta gene is a primary target of 1alpha,25-dihydroxyvitamin D3 and its nuclear receptor. J. Mol. Biol. 2005, 349, 248–260. [Google Scholar] [CrossRef]
- Alimirah, F.; Peng, X.; Yuan, L.; Mehta, R.R.; von Knethen, A.; Choubey, D.; Mehta, R.G. Crosstalk between the peroxisome proliferator-activated receptor γ (PPARγ) and the vitamin D receptor (VDR) in human breast cancer cells: PPARγ binds to VDR and inhibits 1α,25-dihydroxyvitamin D3 mediated transactivation. Exp. Cell. Res. 2012, 318, 2490–2497. [Google Scholar]
- Sertznig, P.; Seifert, M.; Tilgen, W.; Reichrath, J. Activation of vitamin D receptor (VDR)- and peroxisome proliferator-activated receptor (PPAR)-signaling pathways through 1,25(OH)(2)D(3) in melanoma cell lines and other skin-derived cell lines. Dermato-endocrinology 2009, 1, 232–238. [Google Scholar]
- Sertznig, P.; Seifert, M.; Tilgen, W.; Reichrath, J. Peroxisome proliferator-activated receptor (PPAR) and vitamin D receptor (VDR) signaling pathways in melanoma cells: Promising new therapeutic targets? J. Steroid Biochem. Mol. Biol. 2010, 121, 383–386. [Google Scholar]
- Bambury, R.M.; Iyer, G.; Rosenberg, J.E. Specific PPAR gamma agonists may have different effects on cancer incidence. Ann. Oncol. 2013. [Google Scholar] [CrossRef]
- Terrasi, M.; Bazan, V.; Caruso, S.; Insalaco, L.; Amodeo, V.; Fanale, D.; Corsini, L.R.; Contaldo, C.; Mercanti, A.; Fiorio, E.; et al. Effects of PPARγ agonists on the expression of leptin and vascular endothelial growth factor in breast cancer cells. J. Cell Physiol. 2013, 228, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Grabacka, M.; Plonka, P.M.; Urbanska, K.; Reiss, K. Peroxisome proliferator-activated receptor alpha activation decreases metastatic potential of melanoma cells in vitro via down-regulation of Akt. Clin. Cancer Res. 2006, 12, 3028–3036. [Google Scholar]
- Wang, Q.M.; Jones, J.B.; Studzinski, G.P. Cyclin-dependent kinase inhibitor p27 as a mediator of the G1-S phase block induced by 1,25-dihydroxyvitamin D3 in HL60 cells. Cancer Res. 1996, 56, 264–267. [Google Scholar]
- Tosetti, F.; Ferrari, N.; De Flora, S.; Albini, A. Angioprevention: Angiogenesis is a common and key target for cancer chemopreventive agents. FASEB J. 2002, 16, 2–14. [Google Scholar] [CrossRef]
- Lee, H.J.; Liu, H.; Goodman, C.; Ji, Y.; Maehr, H.; Uskokovic, M.; Notterman, D.; Reiss, M.; Suh, N. Gene expression profiling changes induced by a novel Gemini Vitamin D derivative during the progression of breast cancer. Biochem. Pharmacol. 2006, 72, 332–343. [Google Scholar]
- Axanova, L.S.; Chen, Y.Q.; McCoy, T.; Sui, G.; Cramer, S.D. 1,25-dihydroxyvitamin D(3) and PI3K/AKT inhibitors synergistically inhibit growth and induce senescence in prostate cancer cells. Prostate 2010, 70, 1658–1671. [Google Scholar] [CrossRef]
- Kouchi, Z.; Fujiwara, Y.; Yamaguchi, H.; Nakamura, Y.; Fukami, K. Phosphatidylinositol 5-phosphate 4-kinase type II beta is required for vitamin D receptor-dependent E-cadherin expression in SW480 cells. Biochem. Biophys. Res. Commun. 2011, 408, 523–529. [Google Scholar]
- Xiong, M.; Gong, J.; Liu, Y.; Xiang, R.; Tan, X. Loss of vitamin D receptor in chronic kidney disease: A potential mechanism linking inflammation to epithelial-to-mesenchymal transition. Am. J. Physiol. Renal Physiol. 2012, 303, F1107–F1115. [Google Scholar]
- Battaglia, S.; Maguire, O.; Thorne, J.L.; Hornung, L.B.; Doig, C.L.; Liu, S.; Sucheston, L.E.; Bianchi, A.; Khanim, F.L.; Gommersall, L.M.; et al. Elevated NCOR1 disrupts PPARalpha/gamma signaling in prostate cancer and forms a targetable epigenetic lesion. Carcinogenesis 2010, 31, 1650–1660. [Google Scholar]
- Karlic, H.; Varga, F. Impact of vitamin D metabolism on clinical epigenetics. Clin. Epigenet. 2011, 2, 55–61. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Matsuda, S.; Kitagishi, Y. Peroxisome Proliferator-Activated Receptor and Vitamin D Receptor Signaling Pathways in Cancer Cells. Cancers 2013, 5, 1261-1270. https://doi.org/10.3390/cancers5041261
Matsuda S, Kitagishi Y. Peroxisome Proliferator-Activated Receptor and Vitamin D Receptor Signaling Pathways in Cancer Cells. Cancers. 2013; 5(4):1261-1270. https://doi.org/10.3390/cancers5041261
Chicago/Turabian StyleMatsuda, Satoru, and Yasuko Kitagishi. 2013. "Peroxisome Proliferator-Activated Receptor and Vitamin D Receptor Signaling Pathways in Cancer Cells" Cancers 5, no. 4: 1261-1270. https://doi.org/10.3390/cancers5041261