
Proteomic Profiling of Cerebrospinal Fluid and Its Extracellular Vesicles from Extraventricular Drainage in Pediatric Pilocytic Astrocytoma, towards Precision Oncology
 , , , , ,
, , , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Size and Experimental Design
2.2. Patients and Sample Collection
2.3. Mass Spectrometry Profile
2.4. Western Blotting
2.5. ELISA Assay
2.6. Statistical Analysis
3. Results
3.1. Characterization of Extracellular Vesicles
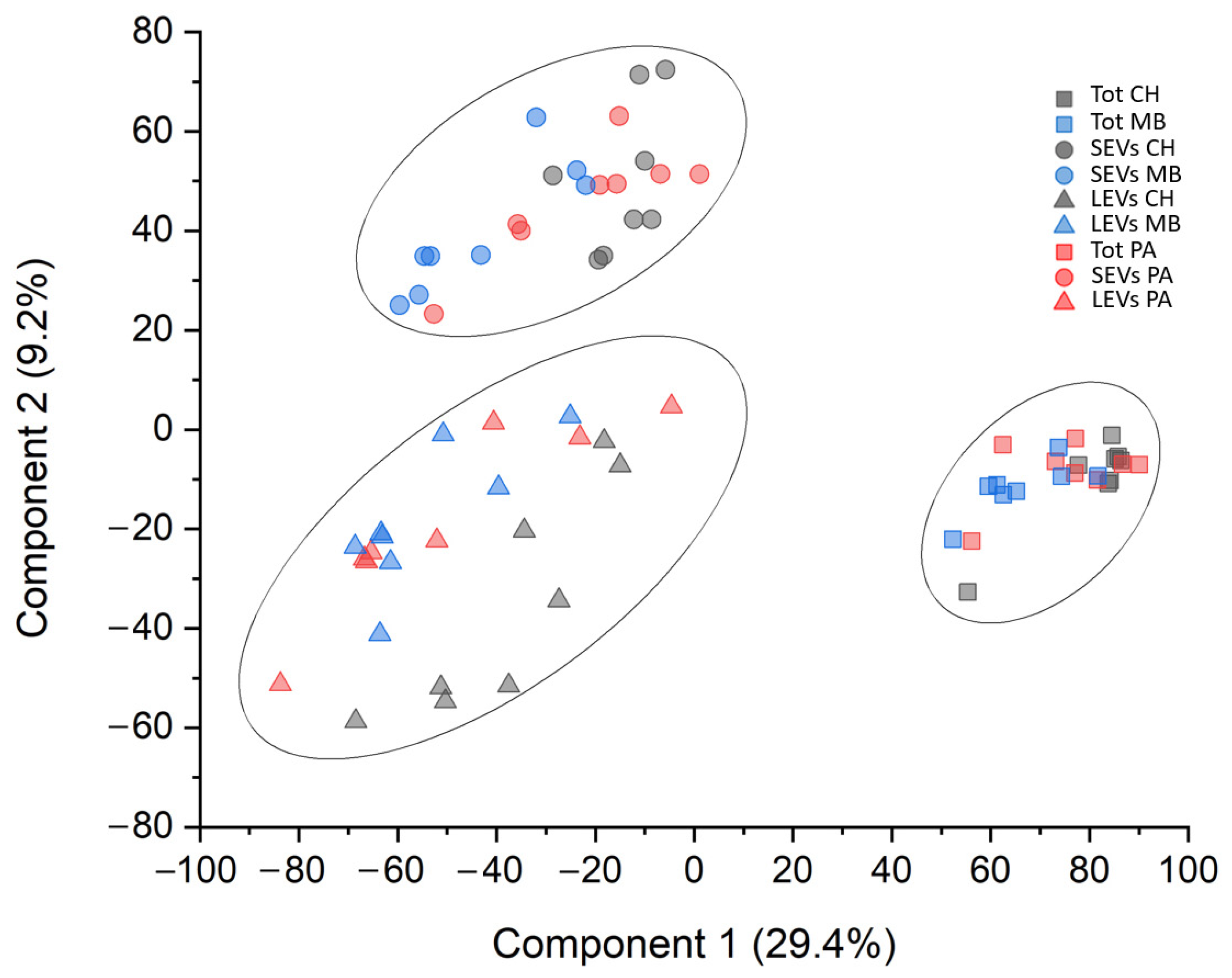
3.2. Protein Profile
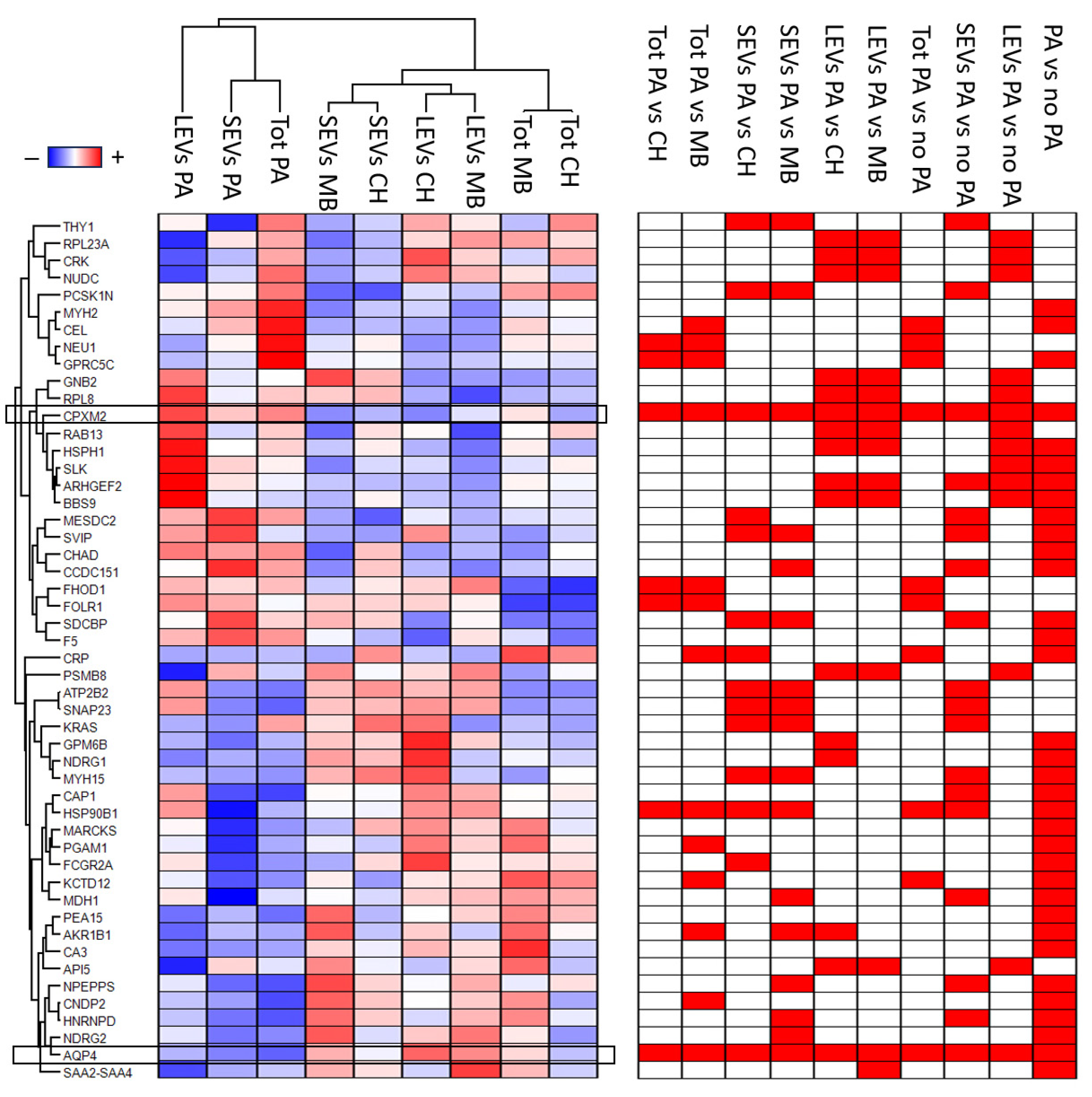
3.3. Gene Ontology Enrichment Analysis
3.4. Western Blot Analysis
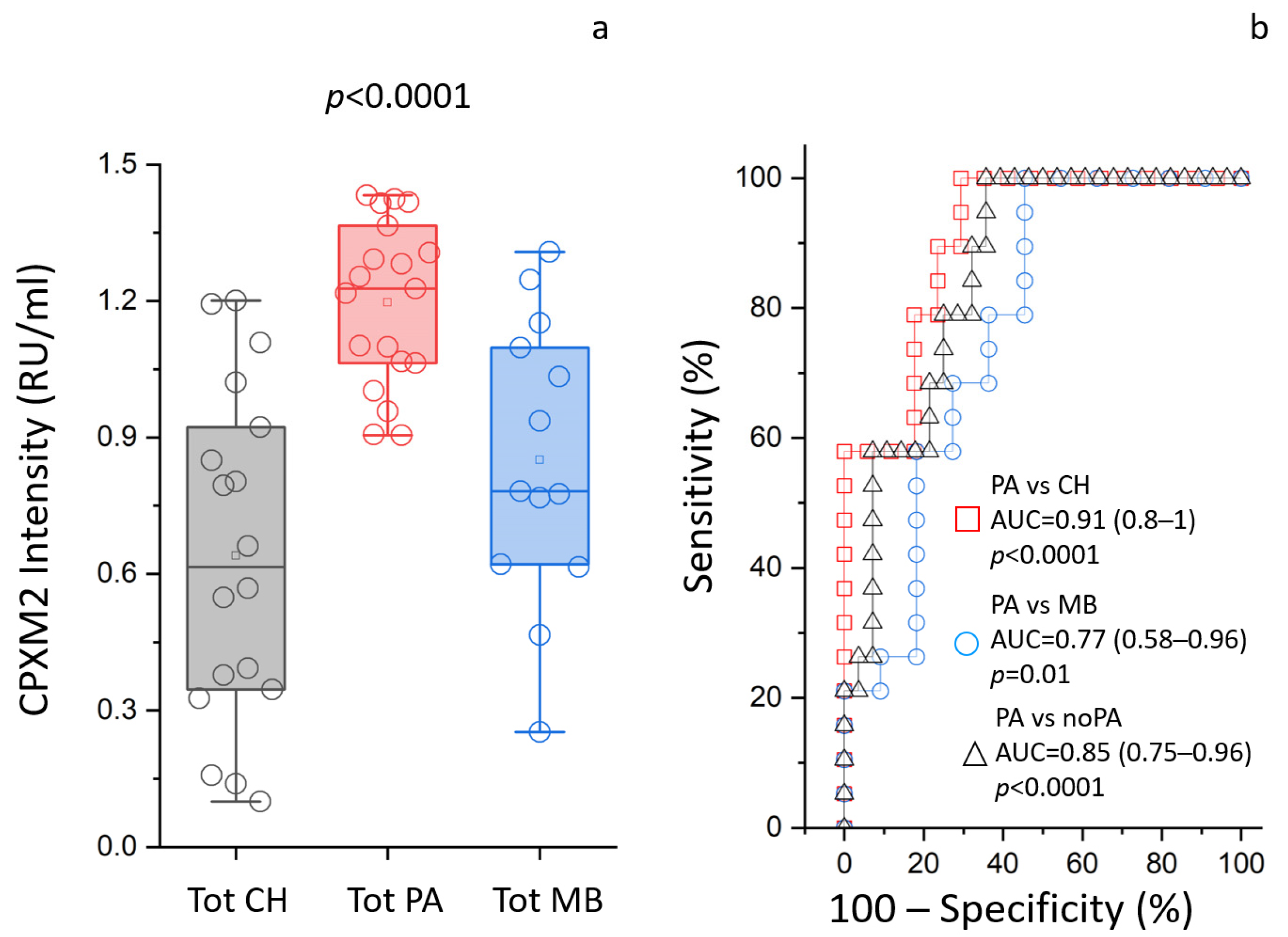
3.5. ELISA Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Damodharan, S.; Puccetti, D. Pediatric Central Nervous System Tumor Overview and Emerging Treatment Considerations. Brain Sci. 2023, 13, 1106. [Google Scholar] [CrossRef] [PubMed]
- Ganz, J.C. Low grade gliomas. Prog. Brain Res. 2022, 268, 271–277. [Google Scholar] [CrossRef]
- Ryall, S.; Tabori, U.; Hawkins, C. Pediatric low-grade glioma in the era of molecular diagnostics. Acta Neuropathol. Commun. 2020, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Salles, D.; Laviola, G.; Malinverni, A.C.M.; Stavale, J.N. Pilocytic Astrocytoma: A Review of General, Clinical, and Molecular Characteristics. J. Child. Neurol. 2020, 35, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Bauman, M.M.J.; Harrison, D.J.; Giesken, M.B.; Daniels, D.J. The evolving landscape of pilocytic astrocytoma: A bibliometric analysis of the top-100 most cited publications. Childs Nerv. Syst. 2022, 38, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Milde, T.; Rodriguez, F.J.; Barnholtz-Sloan, J.S.; Patil, N.; Eberhart, C.G.; Gutmann, D.H. Reimagining pilocytic astrocytomas in the context of pediatric low-grade gliomas. Neuro Oncol. 2021, 23, 1634–1646. [Google Scholar] [CrossRef] [PubMed]
- Mustansir, F.; Mushtaq, N.; Darbar, A. Dabrafenib in BRAFV600E mutant pilocytic astrocytoma in a pediatric patient. Childs Nerv. Syst. 2020, 36, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, T.; Nazemi, K.J.; Crawford, J.; Kilburn, L.; Minturn, J.; Gajjar, A.; Gauvain, K.; Leary, S.; Dhall, G.; Aboian, M.; et al. Phase I study of vemurafenib in children with recurrent or progressive BRAF(V600E) mutant brain tumors: Pacific Pediatric Neuro-Oncology Consortium study (PNOC-002). Oncotarget 2020, 11, 1942–1952. [Google Scholar] [CrossRef]
- Kim, J.W.; Phi, J.H.; Kim, S.K.; Lee, J.H.; Park, S.H.; Won, J.K.; Choi, J.Y.; Kang, H.J.; Park, C.K. Comparison of the clinical features and treatment outcomes of pilocytic astrocytoma in pediatric and adult patients. Childs Nerv. Syst. 2023, 39, 583–591. [Google Scholar] [CrossRef]
- Reinhardt, A.; Stichel, D.; Schrimpf, D.; Sahm, F.; Korshunov, A.; Reuss, D.E.; Koelsche, C.; Huang, K.; Wefers, A.K.; Hovestadt, V.; et al. Anaplastic astrocytoma with piloid features, a novel molecular class of IDH wildtype glioma with recurrent MAPK pathway, CDKN2A/B and ATRX alterations. Acta Neuropathol. 2018, 136, 273–291. [Google Scholar] [CrossRef]
- Ali, R.H.; Almanabri, M.; Ali, N.Y.; Alsaber, A.R.; Khalifa, N.M.; Hussein, R.; Alateeqi, M.; Mohammed, E.M.A.; Jama, H.; Almarzooq, A.; et al. Clinicopathological analysis of BRAF and non-BRAF MAPK pathway-altered gliomas in paediatric and adult patients: A single-institution study of 40 patients. J. Clin. Pathol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, N.; Singh, A.; Kumar, P.; Kaushik, M. Protein kinases: Role of their dysregulation in carcinogenesis, identification and inhibition. Drug Res. 2023, 73, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Migliozzi, S.; Oh, Y.T.; Hasanain, M.; Garofano, L.; D’Angelo, F.; Najac, R.D.; Picca, A.; Bielle, F.; Di Stefano, A.L.; Lerond, J.; et al. Integrative multi-omics networks identify PKCdelta and DNA-PK as master kinases of glioblastoma subtypes and guide targeted cancer therapy. Nat. Cancer 2023, 4, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.I.; Nikonova, A.S.; Sun, D.; Golemis, E.A. Proliferative signaling by ERBB proteins and RAF/MEK/ERK effectors in polycystic kidney disease. Cell Signal 2020, 67, 109497. [Google Scholar] [CrossRef]
- Tripathy, A.; John, V.; Wadden, J.; Kong, S.; Sharba, S.; Koschmann, C. Liquid biopsy in pediatric brain tumors. Front. Genet. 2022, 13, 1114762. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, W.; Lin, F.; Huang, Q.; Zhong, J.; Gao, H.; Song, Y.; Liang, H. DNA methylation profile is a quantitative measure of biological aging in children. Aging 2019, 11, 10031–10051. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.D.; Young, A.M.; Karri, S.K. Biomarkers of pediatric brain tumors. Front. Pediatr. 2013, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Mirian, C.; Thastrup, M.; Mathiasen, R.; Schmiegelow, K.; Olsen, J.V.; Ostergaard, O. Mass spectrometry-based proteomics of cerebrospinal fluid in pediatric central nervous system malignancies: A systematic review with meta-analysis of individual patient data. Fluids Barriers CNS 2024, 21, 14. [Google Scholar] [CrossRef]
- Bruschi, M.; Petretto, A.; Cama, A.; Pavanello, M.; Bartolucci, M.; Morana, G.; Ramenghi, L.A.; Garre, M.L.; Ghiggeri, G.M.; Panfoli, I.; et al. Potential biomarkers of childhood brain tumor identified by proteomics of cerebrospinal fluid from extraventricular drainage (EVD). Sci. Rep. 2021, 11, 1818. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrugger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Lo Cicero, A.; Stahl, P.D.; Raposo, G. Extracellular vesicles shuffling intercellular messages: For good or for bad. Curr. Opin. Cell Biol. 2015, 35, 69–77. [Google Scholar] [CrossRef]
- Bruschi, M.; Ravera, S.; Santucci, L.; Candiano, G.; Bartolucci, M.; Calzia, D.; Lavarello, C.; Inglese, E.; Petretto, A.; Ghiggeri, G.; et al. The human urinary exosome as a potential metabolic effector cargo. Expert. Rev. Proteom. 2015, 12, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Morad, G.; Moses, M.A. Brainwashed by extracellular vesicles: The role of extracellular vesicles in primary and metastatic brain tumour microenvironment. J. Extracell. Vesicles 2019, 8, 1627164. [Google Scholar] [CrossRef] [PubMed]
- D’Asti, E.; Chennakrishnaiah, S.; Lee, T.H.; Rak, J. Extracellular Vesicles in Brain Tumor Progression. Cell Mol. Neurobiol. 2016, 36, 383–407. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, A.; Roy, S.; Fleming, T.; Leong, H.S.; Schuurmans, C. The Emerging Role of Extracellular Vesicles in the Glioma Microenvironment: Biogenesis and Clinical Relevance. Cancers 2020, 12, 1964. [Google Scholar] [CrossRef] [PubMed]
- Akers, J.C.; Ramakrishnan, V.; Kim, R.; Skog, J.; Nakano, I.; Pingle, S.; Kalinina, J.; Hua, W.; Kesari, S.; Mao, Y.; et al. MiR-21 in the extracellular vesicles (EVs) of cerebrospinal fluid (CSF): A platform for glioblastoma biomarker development. PLoS ONE 2013, 8, e78115. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, M.; Kajana, X.; Petretto, A.; Bartolucci, M.; Pavanello, M.; Ghiggeri, G.M.; Panfoli, I.; Candiano, G. Weighted Gene Co-Expression Network Analysis and Support Vector Machine Learning in the Proteomic Profiling of Cerebrospinal Fluid from Extraventricular Drainage in Child Medulloblastoma. Metabolites 2022, 12, 724. [Google Scholar] [CrossRef] [PubMed]
- Kajana, X.; Spinelli, S.; Garbarino, A.; Balagura, G.; Bartolucci, M.; Petretto, A.; Pavanello, M.; Candiano, G.; Panfoli, I.; Bruschi, M. Identification of Central Nervous System Oncologic Disease Biomarkers in EVs from Cerebrospinal Fluid (CSF) of Pediatric Patients: A Pilot Neuro-Proteomic Study. Biomolecules 2023, 13, 1730. [Google Scholar] [CrossRef] [PubMed]
- Dell, R.B.; Holleran, S.; Ramakrishnan, R. Sample size determination. ILAR J. 2002, 43, 207–213. [Google Scholar] [CrossRef]
- Forshed, J. Experimental Design in Clinical ‘Omics Biomarker Discovery. J. Proteome Res. 2017, 16, 3954–3960. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, M.; Granata, S.; Santucci, L.; Candiano, G.; Fabris, A.; Antonucci, N.; Petretto, A.; Bartolucci, M.; Del Zotto, G.; Antonini, F.; et al. Proteomic Analysis of Urinary Microvesicles and Exosomes in Medullary Sponge Kidney Disease and Autosomal Dominant Polycystic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2019, 14, 834–843. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Gaugaz, F.Z. Fast and sensitive total protein and Peptide assays for proteomic analysis. Anal. Chem. 2015, 87, 4110–4116. [Google Scholar] [CrossRef] [PubMed]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Xie, Z.; London, A.B.K.; Yang, J.; Evangelista, J.E.; Lachmann, A.; Shu, I.; Torre, D.; Ma’ayan, A. KEA3: Improved kinase enrichment analysis via data integration. Nucleic Acids Res. 2021, 49, W304–W316. [Google Scholar] [CrossRef] [PubMed]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P.G. Blue silver: A very sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef]
- Lee, B.; Mahmud, I.; Pokhrel, R.; Murad, R.; Yuan, M.; Stapleton, S.; Bettegowda, C.; Jallo, G.; Eberhart, C.G.; Garrett, T.; et al. Medulloblastoma cerebrospinal fluid reveals metabolites and lipids indicative of hypoxia and cancer-specific RNAs. Acta Neuropathol. Commun. 2022, 10, 25. [Google Scholar] [CrossRef]
- Kumar, L.P.; Deepa, S.F.; Moinca, I.; Suresh, P.; Naidu, K.V. Medulloblastoma: A common pediatric tumor: Prognostic factors and predictors of outcome. Asian J. Neurosurg. 2015, 10, 50. [Google Scholar] [CrossRef]
- Iglesias Gonzalez, P.A.; Valdivieso, A.G.; Santa-Coloma, T.A. The G protein-coupled receptor GPRC5A-a phorbol ester and retinoic acid-induced orphan receptor with roles in cancer, inflammation, and immunity. Biochem. Cell Biol. 2023, 101, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.R.; Rossman, K.L.; Der, C.J. Rho guanine nucleotide exchange factors: Regulators of Rho GTPase activity in development and disease. Oncogene 2014, 33, 4021–4035. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.I.; Lechuga, S.; Marino-Melendez, A.; Naydenov, N.G. Unique and redundant functions of cytoplasmic actins and nonmuscle myosin II isoforms at epithelial junctions. Ann. N. Y Acad. Sci. 2022, 1515, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, R.; Roucourt, B.; Lembo, F.; Fares, J.; Carcavilla, A.M.; Restouin, A.; Zimmermann, P.; Ghossoub, R. Syntenin controls migration, growth, proliferation, and cell cycle progression in cancer cells. Front. Pharmacol. 2015, 6, 241. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Matta, A.; Masui, O.; Srivastava, G.; Kaur, J.; Thakar, A.; Shukla, N.K.; RoyChoudhury, A.; Sharma, M.; Walfish, P.G.; et al. Nuclear heterogeneous nuclear ribonucleoprotein D is associated with poor prognosis and interactome analysis reveals its novel binding partners in oral cancer. J. Transl. Med. 2015, 13, 285. [Google Scholar] [CrossRef] [PubMed]
- Teng, X.; Aouacheria, A.; Lionnard, L.; Metz, K.A.; Soane, L.; Kamiya, A.; Hardwick, J.M. KCTD: A new gene family involved in neurodevelopmental and neuropsychiatric disorders. CNS Neurosci. Ther. 2019, 25, 887–902. [Google Scholar] [CrossRef] [PubMed]
- Behnam, M.; Motamedzadeh, A.; Aalinezhad, M.; Dadgostar, E.; Rashidi Noshabad, F.Z.; Pourfridoni, M.; Raei, M.; Mirzaei, H.; Aschner, M.; Tamtaji, O.R. The role of aquaporin 4 in brain tumors: Implications for pathophysiology, diagnosis and therapy. Mol. Biol. Rep. 2022, 49, 10609–10615. [Google Scholar] [CrossRef]
- Niu, G.; Yang, Y.; Ren, J.; Song, T.; Hu, Z.; Chen, L.; Hong, R.; Xia, J.; Ke, C.; Wang, X. Overexpression of CPXM2 predicts an unfavorable prognosis and promotes the proliferation and migration of gastric cancer. Oncol. Rep. 2019, 42, 1283–1294. [Google Scholar] [CrossRef]
- Zhao, X.; Li, R.; Wang, Q.; Wu, M.; Wang, Y. Overexpression of carboxypeptidase X M14 family member 2 predicts an unfavorable prognosis and promotes proliferation and migration of osteosarcoma. Diagn. Pathol. 2019, 14, 118. [Google Scholar] [CrossRef]
- Salles, D.; Santino, S.F.; Ribeiro, D.A.; Malinverni, A.C.M.; Stavale, J.N. The involvement of the MAPK pathway in pilocytic astrocytomas. Pathol. Res. Pract. 2022, 232, 153821. [Google Scholar] [CrossRef]
- Machicoane, M.; De Frutos, C.A.; Fink, J.; Rocancourt, M.; Lombardi, Y.; Garel, S.; Piel, M.; Echard, A. SLK-dependent activation of ERMs controls LGN-NuMA localization and spindle orientation. J. Cell Biol. 2014, 205, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Riman, S.; Rizkallah, R.; Kassardjian, A.; Alexander, K.E.; Luscher, B.; Hurt, M.M. Phosphorylation of the transcription factor YY1 by CK2alpha prevents cleavage by caspase 7 during apoptosis. Mol. Cell Biol. 2012, 32, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, D.; Liu, H.; Guo, C.; Zhao, R.; Zhang, Q.; Xu, C.; Qin, Z.; Feng, J.; Liu, Y.; et al. Comprehensive proteogenomic characterization of early duodenal cancer reveals the carcinogenesis tracks of different subtypes. Nat. Commun. 2023, 14, 1751. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, L.; Wan, D.; Zhou, L.; Zheng, S.; Lin, S.; Qiao, Y. Extracellular matrix and its therapeutic potential for cancer treatment. Signal Transduct. Target. Ther. 2021, 6, 153. [Google Scholar] [CrossRef] [PubMed]
- Czarniak, N.; Kaminska, J.; Matowicka-Karna, J.; Koper-Lenkiewicz, O.M. Cerebrospinal Fluid-Basic Concepts Review. Biomedicines 2023, 11, 1461. [Google Scholar] [CrossRef] [PubMed]
- Bonig, L.; Mohn, N.; Ahlbrecht, J.; Wurster, U.; Raab, P.; Puppe, W.; Suhs, K.W.; Stangel, M.; Skripuletz, T.; Schwenkenbecher, P. Leptomeningeal Metastasis: The Role of Cerebrospinal Fluid Diagnostics. Front. Neurol. 2019, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- Nakano, I.; Garnier, D.; Minata, M.; Rak, J. Extracellular vesicles in the biology of brain tumour stem cells--Implications for inter-cellular communication, therapy and biomarker development. Semin. Cell Dev. Biol. 2015, 40, 17–26. [Google Scholar] [CrossRef]
- Hallal, S.; Azimi, A.; Wei, H.; Ho, N.; Lee, M.Y.T.; Sim, H.W.; Sy, J.; Shivalingam, B.; Buckland, M.E.; Alexander-Kaufman, K.L. A Comprehensive Proteomic SWATH-MS Workflow for Profiling Blood Extracellular Vesicles: A New Avenue for Glioma Tumour Surveillance. Int. J. Mol. Sci. 2020, 21, 4754. [Google Scholar] [CrossRef]
- Sandri, B.J.; Kim, J.; Lubach, G.R.; Lock, E.F.; Ennis-Czerniak, K.; Kling, P.J.; Georgieff, M.K.; Coe, C.L.; Rao, R.B. Prognostic Performance of Hematological and Serum Iron and Metabolite Indices for Detection of Early Iron Deficiency Induced Metabolic Brain Dysfunction in Infant Rhesus Monkeys. J. Nutr. 2024, 154, 875–885. [Google Scholar] [CrossRef]
- Akesson, J.; Hojjati, S.; Hellberg, S.; Raffetseder, J.; Khademi, M.; Rynkowski, R.; Kockum, I.; Altafini, C.; Lubovac-Pilav, Z.; Mellergard, J.; et al. Proteomics reveal biomarkers for diagnosis, disease activity and long-term disability outcomes in multiple sclerosis. Nat. Commun. 2023, 14, 6903. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Li, H.; Li, M.; Han, B.; Qi, Y.; Wang, H.; Gao, J. Label-free Quantitative Proteomic Analysis of Cerebrospinal Fluid and Serum in Patients With Relapse-Remitting Multiple Sclerosis. Front. Genet. 2022, 13, 892491. [Google Scholar] [CrossRef]
- Shama, A.; Soni, T.; Jawanda, I.K.; Upadhyay, G.; Sharma, A.; Prabha, V. The Latest Developments in Using Proteomic Biomarkers from Urine and Serum for Non-Invasive Disease Diagnosis and Prognosis. Biomark. Insights 2023, 18, 11772719231190218. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Discovery/Validation Cohort | Gender (M/F) | Age (Year) |
|---|---|---|---|
| Congenital hydrocephalus (CH) | 8/18 | 14/12 | 1 (0–22) |
| Pilocytic astrocytoma (PA) | 8/19 | 14/13 | 8 (3–15) |
| Medulloblastoma (MB) | 8/13 | 11/10 | 5 (0–15) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spinelli, S.; Kajana, X.; Garbarino, A.; Bartolucci, M.; Petretto, A.; Pavanello, M.; Verrina, E.; Candiano, G.; Panfoli, I.; Bruschi, M. Proteomic Profiling of Cerebrospinal Fluid and Its Extracellular Vesicles from Extraventricular Drainage in Pediatric Pilocytic Astrocytoma, towards Precision Oncology. Cancers 2024, 16, 1223. https://doi.org/10.3390/cancers16061223
Spinelli S, Kajana X, Garbarino A, Bartolucci M, Petretto A, Pavanello M, Verrina E, Candiano G, Panfoli I, Bruschi M. Proteomic Profiling of Cerebrospinal Fluid and Its Extracellular Vesicles from Extraventricular Drainage in Pediatric Pilocytic Astrocytoma, towards Precision Oncology. Cancers. 2024; 16(6):1223. https://doi.org/10.3390/cancers16061223
Chicago/Turabian StyleSpinelli, Sonia, Xhuliana Kajana, Andrea Garbarino, Martina Bartolucci, Andrea Petretto, Marco Pavanello, Enrico Verrina, Giovanni Candiano, Isabella Panfoli, and Maurizio Bruschi. 2024. "Proteomic Profiling of Cerebrospinal Fluid and Its Extracellular Vesicles from Extraventricular Drainage in Pediatric Pilocytic Astrocytoma, towards Precision Oncology" Cancers 16, no. 6: 1223. https://doi.org/10.3390/cancers16061223