1. Introduction
Cancer poses a significant health challenge, ranking as the primary cause of death among individuals aged 40–79 for both males and females. Brain tumors, characterized by elevated mortality and morbidity, are a substantial health concern. More than 15,000 fatalities per year in the United States are attributed to malignant primary brain tumors [
1,
2]. The yearly incidence of primary malignant brain tumors stands at around 7 per 100,000 individuals, with an age-related increase. The five-year survival rate is approximately 36%. Glioblastomas account for roughly 49% of malignant brain tumors, while diffusely infiltrating lower-grade gliomas constitute 30% [
3,
4].
Glioblastoma multiforme (GBM), classified as a WHO grade 4 glioma, stands as the most prevalent malignant primary brain tumor and is acknowledged as the most lethal form of malignant brain tumor. The updated classification of CNS tumors designates GBM as the most aggressive adult tumor. In Jordan, there has been a notable 105.9% increase in the incidence rate of GBM between 1990 and 2019, with an age-standardized incidence rate of 4.4 per 100,000 and a prevalence rate of 15.8 per 100,000 [
5].
The current guidelines for treating individuals recently diagnosed with GBM involve maximum safe surgical removal, followed by a combination of radiotherapy and concurrent/adjuvant chemotherapy [
6]. Complete resection has demonstrated a greater likelihood of better survival and absence of progression compared to partial resection or biopsy [
7]. In the event of GBM recurrence, potential treatment choices encompass supportive care, reoperation, re-irradiation, systemic therapies, and combined modality therapy. The significance of reoperation in this context is currently not well defined [
8]. Current research indicates that the resectability of a tumor may be influenced by its biological characteristics. The primary impediments to the standard of care include various resistance mechanisms, the immunosuppressive microenvironment, and tumor infiltration [
9]. Advancing the standard of care in GBM management by exploring combined therapeutic strategies and delivery methods, encompassing immunotherapy, synthetic molecules, natural compounds, and the inhibition of glioma stem cells, can have the potential to enhance standard therapy in GBM management. [
10,
11].
A thorough comprehension of the interactions among multiple SNPs within a genomic context is crucial. Exploring the combined effects, referred to as epistasis, and their contribution to the variability in complex traits or diseases can advance our grasp of genetic factors. Moreover, examining the impact of SNPs across diverse populations, particularly among Arab Jordanians, may unveil population-specific associations, addressing potential gaps in personalized medicine and genomic risk assessment [
12].
Serine protease inhibitors (Serpins) play a vital role in the regulation of various biological processes, such as inflammation and the immune response. Situated in the 18q21 gene cluster,
Serpinb11 is a polymorphic gene/pseudogene that encodes for a Serpin lacking inhibitory properties [
13]. Previous studies suggest that variants of the
Serpinb11 gene affect its inhibitory serpin function and act with a non-inhibitory function [
14].
In the context of GBM, there is a vital need for a comprehensive and integrated understanding of the various genes and SNP components involved, as well as their interactions. In this study, we aim to elucidate the connection between the prognosis of GBM patients and the SNPs of the Serpinb11 gene with the integration of genomic analysis for Serpinb genes, shedding light on the relationship between genetic factors and overall survival in GBM patients of the Jordanian population.
4. Discussion
Glioblastoma Multiforme is a rapidly progressing tumor that affects the brain or spinal cord and is the most prevalent form of primary malignant brain tumor in adults. GBM patients have a 5-year survival rate of 7.2% and an average survival duration of 15 months [
23,
24]. In this study, we examined the rs4940595 (
Serpinb11) expression in 63 GBM patients. We aimed to investigate the association between rs4940595 (
Serpinb11) under four different inheritance models and genotypes (G/G, G/T, and T/T) and the survival and prognosis rates of GBM patients in Jordan. This study is the first to explore the rs4940595 (
Serpinb11) variant and its association with GBM, revealing a significant association in Jordan and worldwide.
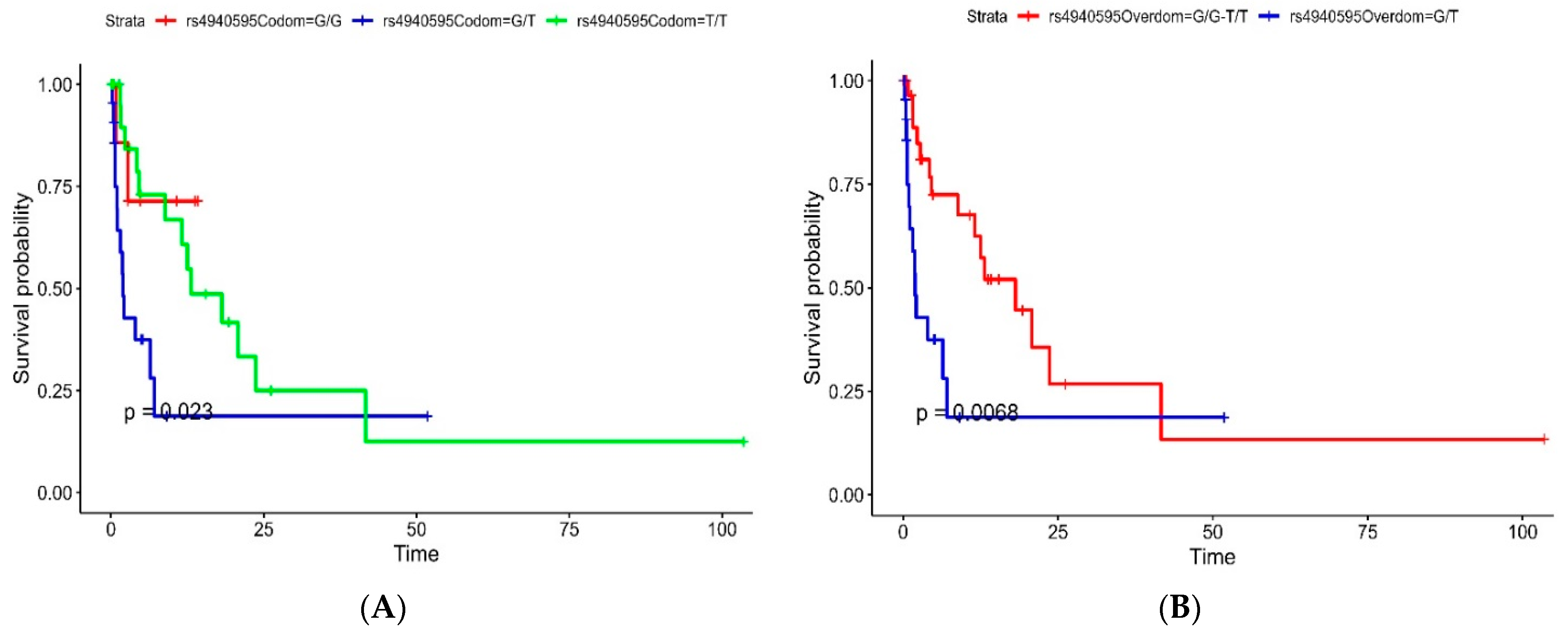
In our investigation, it was revealed that the codominant model of rs4940595 (
Serpinb11) exhibited a partial association with GBM survival with a trend toward a worse prognosis in the G/T genotype. Furthermore, a significant association with a worse prognosis was observed in the overdominant model of patients with the G/T genotype compared to those with the T/T and G/G genotypes. Situated within the 18q21 gene cluster,
Serpinb11 is a polymorphic gene/pseudogene responsible for encoding a non-inhibitory
Serpin [
13].
Serpinbs distinguish themselves in various aspects from other
Serpins. Unlike most
Serpins that function as extracellular proteins,
Serpinbs are primarily located within cytoplasmic or nuclear cell compartments. In these compartments, they are believed to play a role in safeguarding against indiscriminate proteolysis [
25,
26]. Our findings showed a significant association between the G/G genotype of
Serpinb11 and better survival outcomes. The availability of SNP genotyping makes it feasible to test their association with diseases and cancer, potentially revealing a serpin haplotype within clade B that is linked to conditions characterized by substantial changes in the balance between peptidases and inhibitors [
27]. Askew et al. indicated that variant residues within the
Serpinb11 framework negatively impacted serpin inhibitory function. Using sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) analysis, they showed that reactive site loop (RSL)-cleaved Serpinb11 failed to undergo the stressed-to-relaxed transition typically seen in inhibitory-type serpins [
14]. It has been shown previously that non-inhibitory serpins such as
SerpinB5/Maspin can lead to an increase in the sensitivity of tumor cells to cell death and apoptosis, in addition to preventing migration of tumor cells and cancer metastasis [
28].
Particularly,
Serpinb11 has been studied primarily in ovarian cancers; however, this is the first study to examine the role of
Serpinb11 in GBM. A study by Lee et al. investigated the anticancer effects of eupatilin as a potential therapeutic agent directed at
Serpinb11 in ovarian cancer cells, showing an inhibitory effect of eupatilin on
Serpinb11 expression [
29]. Eupatilin, a bioactive flavonoid, has gained attention for its anticancer effects [
30]. Recent studies suggest that eupatilin exerts its effects by modulating the cell cycle or inhibiting metastatic potential in various cancers, including gastric cancer, endometrial cancer, and glioma cells [
30,
31,
32]. In line with our findings in ovarian cancer, Park et al. showed that higher expression of
Serpinb11 was correlated with a poor prognosis in high-grade serous and clear cell carcinoma of the ovary [
33]. These findings suggest the potential utility of
Serpinb11 as a prognostic biomarker [
34].
In our bioinformatics analysis, we constructed a 5-gene risk score from the
Serpinb family, including
Serpinb3, Serpinb5, Serpinb6, Serpinb9, Serpinb11, and
Serpinb12. Our results of the
Serpinb 5-gene risk score revealed a trend toward better progression-free survival in low-risk patients. Furthermore, patients with higher expression of
Serpinb5 and
Serpinb6 were associated with significantly worse outcomes.
Serpinb5, referred to as Maspin (mammary serine protease inhibitor), was identified as a serine protease inhibitor and recognized as a tumor suppressor, and its loss has been observed in breast and prostate cancers, making it a promising diagnostic marker for monitoring tumor progression [
35,
36]. Several studies showed that Maspin (
Serpinb5) functions as a tumor suppressor gene, exerting inhibitory effects on angiogenesis, promoting cellular adhesion, and suppressing the migration of cancer cells [
37,
38]. In line with our findings, a bioinformatics study by He et al. showed that Serpinb5 expression was upregulated in lung adenocarcinoma and hypomethylated, with associations with poor survival in patients with higher Serpinb5 expression suggesting its role as a possible therapeutic target [
39]. Additionally, a study by Ma et al. showed that Serpinb5 mRNA expression was downregulated in glioma with a negative correlation with tumor grade compared to normal brain tissue [
37].
Furthermore,
Serpinb6, previously known as proteinase inhibitor 6 (
PI6), acts as a universal inhibitor of granule protease. Its expression is widespread, and it plays a role in inhibiting both metastasis and tumor progression [
40,
41]. A study by Burgener et al. showed that
Serpinb6 inhibits Cathepsin G in neutrophils and monocytes, preventing programmed necrosis [
42]. Consequently, Song et al. demonstrated the potential role of
Serpinb6 as a contributor to the regular functioning of CAR-T cells. However, additional research is necessary to validate this concept [
43]. Among the other Serpins studied in the literature,
Serpinb3 showed a suppressor of lysosomal-mediated cell death in glioblastoma cancer stem cells. Lauko et al. study illustrates that
Serpinb3 impedes the activity of cathepsin L released from lysosomes, resulting in enhanced resistance to radiation. Targeting this axis could represent a strategy to enhance the effectiveness of radiotherapy not only in glioblastoma but also in other cancer types [
44].
Additionally,
Serpinb12, previously identified as Yukopin, functions as a trypsin inhibitor and exhibits expression in various tissues, including the blood, kidney, liver, heart, and brain. Minimal expression of
Serpinb12 was observed in granular cells, Purkinje cells, and neurons/axons within the cerebellum, as well as in the axons and neuropil of the cerebral cortex [
45]. A study by Sun et al. revealed that the gene expression of
Serpinb12 was associated with a protective role in stage I-IIIA lung adenocarcinoma based on recurrence-free survival [
46]. Considering protective factors, it has been observed in previous reports that
Serpinb12 exhibits abnormal expression in the lungs. However, there is no additional clarification regarding its specific role [
45]. Additionally, animal studies suggest that
Serpinb12 has the potential to serve as a biomarker and may be employed for the early detection of ovarian cancer in women [
47].
Our study provides several strengths. First, our sample size is considered sufficient from a clinical perspective due to the low incidence rate of GBM. Second, we utilized a bioinformatics pipeline to explore the multi-omics characteristics of the Serpin B family, involving a GBM cohort from The Cancer Genome Atlas (TCGA-GBM). This adds a layer of complexity to the study, allowing for a comprehensive examination of gene expression, differentially expressed genes, functional enrichment, and pathway analysis.
However, our findings should be interpreted with caution in the context of several limitations. First, our study encountered substantial epidemiological and demographic constraints as it drew cases from a single-center tertiary hospital in North Jordan; thus, the generalizability of the findings is restricted to the Jordanian population. In our study, we have profiled the genotypes of specific SNPs. However, future large-scale multi-center studies incorporating gene sequencing for the whole exome and next-generation sequencing may reveal a significant association with the prevalence and survival rate of GBM cases. Furthermore, we emphasize the importance of conducting additional studies with larger and more generalized samples, representing a broader population. These studies should target multiple factors that were not well studied in this research, potentially providing a better representation of the truth. Integrating additional omics data, such as proteomics and epigenomics, could provide a more comprehensive understanding of the molecular landscape associated with glioblastoma. This could uncover additional therapeutic targets and biomarkers.

 ,
, 


{kind=link}
{kind=link}