
Insights into the Human Microbiome and Its Connections with Prostate Cancer
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Evidence Acquisition
3. Gut Microbiome and Pca
4. Urinary Microbiome and Pca
5. Prostate Microbiome and PCa
6. Oral and Skin Microbiomes in Relation to PCa
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Drummond, F.J.; Carsin, A.-E.; Sharp, L.; Comber, H. Factors prompting PSA-testing of asymptomatic men in a country with no guidelines: A national survey of general practitioners. BMC Fam. Pract. 2009, 10, 3. [Google Scholar] [CrossRef]
- Franco, G.S.; Hardie, R.; Li, L.; Imai, C.; Sezgin, G.; Li, J.; McLeod, A.; Pearce, C.; Georgiou, A. Prostate-specific antigen testing of asymptomatic men in Australia: An observational study based on electronic general practice data. Med. J. Aust. 2021, 215, 228–229. [Google Scholar] [CrossRef] [PubMed]
- Biddle, C.; Brasel, A.; Underwood, W.; Orom, H. Experiences of Uncertainty in Men With an Elevated PSA. Am. J. Mens. Health 2017, 11, 24–34. [Google Scholar] [CrossRef]
- Schatteman, P.H.F.; Hoekx, L.; Wyndaele, J.J.; Jeuris, W.; Van Marck, E. Inflammation in Prostate Biopsies of Men without Prostatic Malignancy or Clinical Prostatitis. Eur. Urol. 2000, 37, 404–412. [Google Scholar] [CrossRef]
- Shinohara, D.B.; Vaghasia, A.M.; Yu, S.-H.; Mak, T.N.; Brüggemann, H.; Nelson, W.G.; De Marzo, A.M.; Yegnasubramanian, S.; Sfanos, K.S. A mouse model of chronic prostatic inflammation using a human prostate cancer-derived isolate of Propionibacterium acnes. Prostate 2013, 73, 1007–1015. [Google Scholar] [CrossRef]
- Alfano, M.; Canducci, F.; Nebuloni, M.; Clementi, M.; Montorsi, F.; Salonia, A. The interplay of extracellular matrix and microbiome in urothelial bladder cancer. Nat. Rev. Urol. 2016, 13, 77–90. [Google Scholar] [CrossRef]
- Javier-DesLoges, J.; McKay, R.R.; Swafford, A.D.; Sepich-Poore, G.D.; Knight, R.; Parsons, J.K. The microbiome and prostate cancer. Prostate Cancer Prostatic Dis. 2022, 25, 159–164. [Google Scholar] [CrossRef]
- Porter, C.M.; Shrestha, E.; Peiffer, L.B.; Sfanos, K.S. The microbiome in prostate inflammation and prostate cancer. Prostate Cancer Prostatic Dis. 2018, 21, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Estemalik, J.; Demko, C.; Bissada, N.F.; Joshi, N.; Bodner, D.; Shankar, E.; Gupta, S. Simultaneous Detection of Oral Pathogens in Subgingival Plaque and Prostatic Fluid of Men With Periodontal and Prostatic Diseases. J. Periodontol. 2017, 88, 823–829. [Google Scholar] [CrossRef]
- Roy, S.; Trinchieri, G. Microbiota: A key orchestrator of cancer therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef]
- De Marzo, A.M.; Platz, E.A.; Sutcliffe, S.; Xu, J.; Grönberg, H.; Drake, C.G.; Nakai, Y.; Isaacs, W.B.; Nelson, W.G. Inflammation in prostate carcinogenesis. Nat. Rev. Cancer 2007, 7, 256–269. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Sfanos, K.S.; Yegnasubramanian, S.; Nelson, W.G.; De Marzo, A.M. The inflammatory microenvironment and microbiome in prostate cancer development. Nat. Rev. Urol. 2018, 15, 11–24. [Google Scholar] [CrossRef]
- Golombos, D.M.; Ayangbesan, A.; O’Malley, P.; Lewicki, P.; Barlow, L.; Barbieri, C.E.; Chan, C.; DuLong, C.; Abu-Ali, G.; Huttenhower, C.; et al. The Role of Gut Microbiome in the Pathogenesis of Prostate Cancer: A Prospective, Pilot Study. Urology 2018, 111, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Liss, M.A.; White, J.R.; Goros, M.; Gelfond, J.; Leach, R.; Johnson-Pais, T.; Lai, Z.; Rourke, E.; Basler, J.; Ankerst, D.; et al. Metabolic Biosynthesis Pathways Identified from Fecal Microbiome Associated with Prostate Cancer. Eur. Urol. 2018, 74, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.T.; Dold, S.; Rehman, A.; Bird, J.K.; Steinert, R.E. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr. Res. 2021, 95, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Fujita, K.; Motooka, D.; Hatano, K.; Fukae, S.; Kawamura, N.; Tomiyama, E.; Hayashi, Y.; Banno, E.; Takao, T.; et al. The gut microbiota associated with high-Gleason prostate cancer. Cancer Sci. 2021, 112, 3125–3135. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Metabolism of Dietary and Microbial Vitamin B Family in the Regulation of Host Immunity. Front. Nutr. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Magnúsdóttir, S.; Ravcheev, D.; de Crécy-Lagard, V.; Thiele, I. Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes. Front. Genet. 2015, 6, 148. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.J. Intestinal flora and endogenous vitamin synthesis. Eur. J. Cancer Prev. 1997, 6, S43–S45. [Google Scholar] [CrossRef]
- Pieroth, R.; Paver, S.; Day, S.; Lammersfeld, C. Folate and Its Impact on Cancer Risk. Curr. Nutr. Rep. 2018, 7, 70–84. [Google Scholar] [CrossRef]
- Franko, A.; Shao, Y.; Heni, M.; Hennenlotter, J.; Hoene, M.; Hu, C.; Liu, X.; Zhao, X.; Wang, Q.; Birkenfeld, A.L.; et al. Human Prostate Cancer is Characterized by an Increase in Urea Cycle Metabolites. Cancers 2020, 12, 1814. [Google Scholar] [CrossRef]
- Bassett, J.K.; Severi, G.; Hodge, A.M.; Baglietto, L.; Hopper, J.L.; English, D.R.; Giles, G.G. Dietary intake of B vitamins and methionine and prostate cancer incidence and mortality. Cancer Causes Control 2012, 23, 855–863. [Google Scholar] [CrossRef]
- Engevik, M.A.; Morra, C.N.; Röth, D.; Engevik, K.; Spinler, J.K.; Devaraj, S.; Crawford, S.E.; Estes, M.K.; Kalkum, M.; Versalovic, J. Microbial Metabolic Capacity for Intestinal Folate Production and Modulation of Host Folate Receptors. Front. Microbiol. 2019, 10, 2305. [Google Scholar] [CrossRef]
- Price, A.J.; Travis, R.C.; Appleby, P.N.; Albanes, D.; Barricarte Gurrea, A.; Bjørge, T.; Bueno-de-Mesquita, H.B.; Chen, C.; Donovan, J.; Gislefoss, R.; et al. Circulating Folate and Vitamin B12 and Risk of Prostate Cancer: A Collaborative Analysis of Individual Participant Data from Six Cohorts Including 6875 Cases and 8104 Controls. Eur. Urol. 2016, 70, 941–951. [Google Scholar] [CrossRef]
- Cavalieri, E.; Chakravarti, D.; Guttenplan, J.; Hart, E.; Ingle, J.; Jankowiak, R.; Muti, P.; Rogan, E.; Russo, J.; Santen, R.; et al. Catechol estrogen quinones as initiators of breast and other human cancers: Implications for biomarkers of susceptibility and cancer prevention. Biochim. Biophys. Acta-Rev. Cancer 2006, 1766, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Bland, J.S. Prostate Cancer Risk Connection to Immunity, Hormones, and the Microbiome. Integr. Med. 2021, 20, 14–17. [Google Scholar]
- Nelles, J.L.; Hu, W.-Y.; Prins, G.S. Estrogen action and prostate cancer. Expert Rev. Endocrinol. Metab. 2011, 6, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Wirén, S.; Stocks, T.; Rinaldi, S.; Hallmans, G.; Bergh, A.; Stenman, U.-H.; Kaaks, R.; Stattin, P. Androgens and prostate cancer risk: A prospective study. Prostate 2007, 67, 1230–1237. [Google Scholar] [CrossRef]
- Komorowski, A.S.; Pezo, R.C. Untapped “-omics”: The microbial metagenome, estrobolome, and their influence on the development of breast cancer and response to treatment. Breast Cancer Res. Treat. 2020, 179, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Ervin, S.M.; Li, H.; Lim, L.; Roberts, L.R.; Liang, X.; Mani, S.; Redinbo, M.R. Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens. J. Biol. Chem. 2019, 294, 18586–18599. [Google Scholar] [CrossRef] [PubMed]
- Tetel, M.J.; de Vries, G.J.; Melcangi, R.C.; Panzica, G.; O’Mahony, S.M. Steroids, stress and the gut microbiome-brain axis. J. Neuroendocrinol. 2018, 30, e12548. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Ikegawa, S.; Alves, J.M.P.; Zhou, B.; Kobayashi, A.; Iida, T.; Mitamura, K.; Tanabe, G.; Serrano, M.; De Guzman, A.; et al. Clostridium scindens: A human gut microbe with a high potential to convert glucocorticoids into androgens. J. Lipid Res. 2013, 54, 2437–2449. [Google Scholar] [CrossRef]
- Shin, J.-H.; Park, Y.-H.; Sim, M.; Kim, S.-A.; Joung, H.; Shin, D.-M. Serum level of sex steroid hormone is associated with diversity and profiles of human gut microbiome. Res. Microbiol. 2019, 170, 192–201. [Google Scholar] [CrossRef]
- Xu, H.; Cao, C.; Ren, Y.; Weng, S.; Liu, L.; Guo, C.; Wang, L.; Han, X.; Ren, J.; Liu, Z. Antitumor effects of fecal microbiota transplantation: Implications for microbiome modulation in cancer treatment. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wu, J.; Jin, D.; Wang, B.; Cao, H. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int. J. Cancer 2019, 145, 2021–2031. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, T.; Chen, X.; Ji, G.; Zhang, F. Microbiota transplantation: Targeting cancer treatment. Cancer Lett. 2019, 452, 144–151. [Google Scholar] [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Maleki Vareki, S.; Chanyi, R.M.; Abdur-Rashid, K.; Brennan, L.; Burton, J.P. Moving on from Metchnikoff: Thinking about microbiome therapeutics in cancer. Ecancermedicalscience 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Fijlstra, M.; Ferdous, M.; Koning, A.M.; Rings, E.H.H.M.; Harmsen, H.J.M.; Tissing, W.J.E. Substantial decreases in the number and diversity of microbiota during chemotherapy-induced gastrointestinal mucositis in a rat model. Support. Care Cancer 2015, 23, 1513–1522. [Google Scholar] [CrossRef]
- Bode, H.B.; Zeggel, B.; Silakowski, B.; Wenzel, S.C.; Reichenbach, H.; Müller, R. Steroid biosynthesis in prokaryotes: Identification of myxobacterial steroids and cloning of the first bacterial 2,3(S)-oxidosqualene cyclase from the myxobacterium Stigmatella aurantiaca. Mol. Microbiol. 2003, 47, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.-Y.; Yang, Y.-C.; Wang, C.-I.; Hsiao, P.-W.; Chiang, H.-I.; Chen, T.-W. Increase in Akkermansiaceae in Gut Microbiota of Prostate Cancer-Bearing Mice. Int. J. Mol. Sci. 2021, 22, 9626. [Google Scholar] [CrossRef] [PubMed]
- Sfanos, K.S.; Markowski, M.C.; Peiffer, L.B.; Ernst, S.E.; White, J.R.; Pienta, K.J.; Antonarakis, E.S.; Ross, A.E. Compositional differences in gastrointestinal microbiota in prostate cancer patients treated with androgen axis-targeted therapies. Prostate Cancer Prostatic Dis. 2018, 21, 539–548. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef]
- Reis Ferreira, M.; Andreyev, H.J.N.; Mohammed, K.; Truelove, L.; Gowan, S.M.; Li, J.; Gulliford, S.L.; Marchesi, J.R.; Dearnaley, D.P. Microbiota- and Radiotherapy-Induced Gastrointestinal Side-Effects (MARS) Study: A Large Pilot Study of the Microbiome in Acute and Late-Radiation Enteropathy. Clin. Cancer Res. 2019, 25, 6487–6500. [Google Scholar] [CrossRef]
- Nicolaro, M.; Portal, D.E.; Shinder, B.; Patel, H.V.; Singer, E.A. The human microbiome and genitourinary malignancies. Ann. Transl. Med. 2020, 8, 1245. [Google Scholar] [CrossRef]
- Sambi, M.; Bagheri, L.; Szewczuk, M.R. Current Challenges in Cancer Immunotherapy: Multimodal Approaches to Improve Efficacy and Patient Response Rates. J. Oncol. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Geier, M.S.; Howarth, G.S. Prebiotics: A Potential Treatment Strategy for the Chemotherapy-damaged Gut? Crit. Rev. Food Sci. Nutr. 2016, 56, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract—A role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Perez-Carrasco, V.; Soriano-Lerma, A.; Soriano, M.; Gutiérrez-Fernández, J.; Garcia-Salcedo, J.A. Urinary Microbiome: Yin and Yang of the Urinary Tract. Front. Cell. Infect. Microbiol. 2021, 11, 617002. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.E.; Hinman, F. Experiments with Induced Bacteriuria, Vesical Emptying and Bacterial Growth on the Mechanism of Bladder Defense to Infection. J. Urol. 1961, 86, 739–748. [Google Scholar] [CrossRef]
- Fowler, J.E. Secretory immunity of the prostate gland. Infection 1991, 19, S131–S137. [Google Scholar] [CrossRef]
- Ueda, N.; Kondo, M.; Takezawa, K.; Kiuchi, H.; Sekii, Y.; Inagaki, Y.; Soda, T.; Fukuhara, S.; Fujita, K.; Uemura, M.; et al. Bladder urothelium converts bacterial lipopolysaccharide information into neural signaling via an ATP-mediated pathway to enhance the micturition reflex for rapid defense. Sci. Rep. 2020, 10, 21167. [Google Scholar] [CrossRef]
- Krieger, J.N.; Rein, M.F. Zinc Sensitivity of Trichomonas vaginalis: In Vitro Studies and Clinical Implications. J. Infect. Dis. 1982, 146, 341–345. [Google Scholar] [CrossRef]
- Nelson, D.E.; Van Der Pol, B.; Dong, Q.; Revanna, K.V.; Fan, B.; Easwaran, S.; Sodergren, E.; Weinstock, G.M.; Diao, L.; Fortenberry, J.D. Characteristic Male Urine Microbiomes Associate with Asymptomatic Sexually Transmitted Infection. PLoS ONE 2010, 5, e14116. [Google Scholar] [CrossRef]
- Dong, Q.; Nelson, D.E.; Toh, E.; Diao, L.; Gao, X.; Fortenberry, J.D.; Van Der Pol, B. The Microbial Communities in Male First Catch Urine Are Highly Similar to Those in Paired Urethral Swab Specimens. PLoS ONE 2011, 6, e19709. [Google Scholar] [CrossRef] [PubMed]
- Davidsson, S.; Mölling, P.; Rider, J.R.; Unemo, M.; Karlsson, M.G.; Carlsson, J.; Andersson, S.-O.; Elgh, F.; Söderquist, B.; Andrén, O. Frequency and typing of Propionibacterium acnes in prostate tissue obtained from men with and without prostate cancer. Infect. Agent. Cancer 2016, 11, 26. [Google Scholar] [CrossRef]
- Simons, B.W.; Durham, N.M.; Bruno, T.C.; Grosso, J.F.; Schaeffer, A.J.; Ross, A.E.; Hurley, P.J.; Berman, D.M.; Drake, C.G.; Thumbikat, P.; et al. A human prostatic bacterial isolate alters the prostatic microenvironment and accelerates prostate cancer progression. J. Pathol. 2015, 235, 478–489. [Google Scholar] [CrossRef]
- Manente, L.; Gargiulo, U.; Gargiulo, P.; Dovinola, G. Propionibacterium acnes in urine and semen samples from men with urinary infection. Arch. Ital. Urol. Androl. 2022, 94, 62–64. [Google Scholar] [CrossRef]
- Shoskes, D.A.; Altemus, J.; Polackwich, A.S.; Tucky, B.; Wang, H.; Eng, C. The Urinary Microbiome Differs Significantly Between Patients With Chronic Prostatitis/Chronic Pelvic Pain Syndrome and Controls as Well as Between Patients With Different Clinical Phenotypes. Urology 2016, 92, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Khasriya, R.; Sathiananthamoorthy, S.; Ismail, S.; Kelsey, M.; Wilson, M.; Rohn, J.L.; Malone-Lee, J. Spectrum of Bacterial Colonization Associated with Urothelial Cells from Patients with Chronic Lower Urinary Tract Symptoms. J. Clin. Microbiol. 2013, 51, 2054–2062. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Guerra, G.; Lara-Oya, A.; Martínez-Egea, I.; Navarro-Marí, J.M.; Gutiérrez-Fernández, J. Urinary tract infection by Aerococcus sanguinicola. An emerging opportunistic pathogen. Rev. Clín. Esp. 2018, 218, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Pérez, E.; Aparicio-Gómez, J.A.; Gómez-Camarasa, C.; Gutiérrez-Fernández, J. A study of urinary tract infections by Streptococcus gallolyticus ssp. pasteurianus. Rev. Esp. Quimioter. 2019, 32, 189–191. [Google Scholar]
- Ruiz-Pino, M.; Foronda-García-Hidalgo, C.; Alarcón-Blanco, P.; Gutiérrez-Fernández, J. Male genitourinary infections by Corynebacterium glucuronolyticum. A review and clinical experience. Rev. Esp. Quimioter. 2019, 32, 479–484. [Google Scholar]
- Heras-Cañas, V.; Ros, L.; Sorlózano, A.; Gutiérrez-Soto, B.; Navarro-Marí, J.M.; Gutiérrez-Fernández, J. Especies de levaduras aisladas en muestras de orina en un hospital regional de España. Rev. Argent. Microbiol. 2015, 47, 331–334. [Google Scholar] [CrossRef]
- Moustafa, A.; Li, W.; Singh, H.; Moncera, K.J.; Torralba, M.G.; Yu, Y.; Manuel, O.; Biggs, W.; Venter, J.C.; Nelson, K.E.; et al. Microbial metagenome of urinary tract infection. Sci. Rep. 2018, 8, 4333. [Google Scholar] [CrossRef]
- Hiergeist, A.; Gessner, A. Clinical implications of the microbiome in urinary tract diseases. Curr. Opin. Urol. 2017, 27, 93–98. [Google Scholar] [CrossRef]
- Tolani, M.A.; Suleiman, A.; Awaisu, M.; Abdulaziz, M.M.; Lawal, A.T.; Bello, A. Acute urinary tract infection in patients with underlying benign prostatic hyperplasia and prostate cancer. Pan Afr. Med. J. 2020, 36, 1–9. [Google Scholar] [CrossRef]
- Heyns, C.F. Urinary tract infection associated with conditions causing urinary tract obstruction and stasis, excluding urolithiasis and neuropathic bladder. World J. Urol. 2012, 30, 77–83. [Google Scholar] [CrossRef]
- Fan, C.-Y.; Huang, W.-Y.; Lin, K.-T.; Lin, C.-S.; Chao, H.-L.; Yang, J.-F.; Lin, C.-L.; Kao, C.-H. Lower Urinary Tract Infection and Subsequent Risk of Prostate Cancer: A Nationwide Population-Based Cohort Study. PLoS ONE 2017, 12, e0168254. [Google Scholar] [CrossRef]
- Lin, J.L.; Donegan, S.P.; Heeren, T.C.; Greenberg, M.; Flaherty, E.E.; Haivanis, R.; Su, X.; Dean, D.; Newhall, W.J.; Knapp, J.S.; et al. Transmission of Chlamydia trachomatis and Neisseria gonorrhoeae among Men with Urethritis and Their Female Sex Partners. J. Infect. Dis. 1998, 178, 1707–1712. [Google Scholar] [CrossRef]
- Marcus, J.L.; Kohn, R.P.; Barry, P.M.; Philip, S.S.; Bernstein, K.T. Chlamydia trachomatis and Neisseria gonorrhoeae Transmission From the Female Oropharynx to the Male Urethra. Sex. Transm. Dis. 2011, 38, 372–373. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Hayes, R.; Pfeiffer, R.; Viscidi, R.P.; Lee, F.K.; Wang, Y.F.; Reding, D.; Whitby, D.; Papp, J.R.; Rabkin, C.S. Sexually Transmissible Infections and Prostate Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2374–2381. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, S.; Zenilman, J.M.; Ghanem, K.G.; Jadack, R.A.; Sokoll, L.J.; Elliott, D.J.; Nelson, W.G.; De Marzo, A.M.; Cole, S.R.; Isaacs, W.B.; et al. Sexually Transmitted Infections and Prostatic Inflammation/Cell Damage as Measured by Serum Prostate Specific Antigen Concentration. J. Urol. 2006, 175, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Meader, E.; Gihawi, A.; Rallapalli, G.; Clark, J.; Kay, G.L.; Webb, M.; Manley, K.; Curley, H.; Walker, H.; et al. Microbiomes of Urine and the Prostate Are Linked to Human Prostate Cancer Risk Groups. Eur. Urol. Oncol. 2022. [Google Scholar] [CrossRef]
- Liss, M.; Lee, J.; White, J.; Johnson-Pais, T.; Lai, Z.; Troyer, D.; Leach, R.; Wickes, B. MP37-08 urinary microbiome as a biomarker for prostate cancer. J. Urol. 2022, 207, e611. [Google Scholar] [CrossRef]
- Garbas, K.; Zapała, P.; Zapała, Ł.; Radziszewski, P. The Role of Microbial Factors in Prostate Cancer Development—An Up-to-Date Review. J. Clin. Med. 2021, 10, 4772. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, E.; White, J.R.; Yu, S.-H.; Kulac, I.; Ertunc, O.; De Marzo, A.M.; Yegnasubramanian, S.; Mangold, L.A.; Partin, A.W.; Sfanos, K.S. Profiling the Urinary Microbiome in Men with Positive versus Negative Biopsies for Prostate Cancer. J. Urol. 2018, 199, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Alanee, S.; El-Zawahry, A.; Dynda, D.; Dabaja, A.; McVary, K.; Karr, M.; Braundmeier-Fleming, A. A prospective study to examine the association of the urinary and fecal microbiota with prostate cancer diagnosis after transrectal biopsy of the prostate using 16sRNA gene analysis. Prostate 2019, 79, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Meng, H.; Zhou, F.; Ni, X.; Shen, S.; Das, U.N. Urinary microbiota in patients with prostate cancer and benign prostatic hyperplasia. Arch. Med. Sci. 2015, 2, 385–394. [Google Scholar] [CrossRef]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.S. Contamination in Low Microbial Biomass Microbiome Studies: Issues and Recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef]
- Massari, F.; Mollica, V.; Di Nunno, V.; Gatto, L.; Santoni, M.; Scarpelli, M.; Cimadamore, A.; Lopez-Beltran, A.; Cheng, L.; Battelli, N.; et al. The Human Microbiota and Prostate Cancer: Friend or Foe? Cancers 2019, 11, 459. [Google Scholar] [CrossRef]
- Sfanos, K.S.; Isaacs, W.B.; De Marzo, A.M. Infections and inflammation in prostate cancer. Am. J. Clin. Exp. Urol. 2013, 1, 3–11. [Google Scholar]
- Ragnarsdóttir, B.; Lutay, N.; Grönberg-Hernandez, J.; Köves, B.; Svanborg, C. Genetics of innate immunity and UTI susceptibility. Nat. Rev. Urol. 2011, 8, 449–468. [Google Scholar] [CrossRef]
- Miyake, M.; Ohnishi, K.; Hori, S.; Nakano, A.; Nakano, R.; Yano, H.; Ohnishi, S.; Owari, T.; Morizawa, Y.; Itami, Y.; et al. Mycoplasma genitalium Infection and Chronic Inflammation in Human Prostate Cancer: Detection Using Prostatectomy and Needle Biopsy Specimens. Cells 2019, 8, 212. [Google Scholar] [CrossRef]
- Banerjee, S.; Alwine, J.C.; Wei, Z.; Tian, T.; Shih, N.; Sperling, C.; Guzzo, T.; Feldman, M.D.; Robertson, E.S. Microbiome signatures in prostate cancer. Carcinogenesis 2019, 40, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Samanta, M.; Harkins, L.; Klemm, K.; Britt, W.J.; Cobbs, C.S. High Prevalence of Human Cytomegalovirus in Prostatic Intraepithelial Neoplasia and Prostatic Carcinoma. J. Urol. 2003, 170, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fierro, M.L.; Leach, R.J.; Gomez-Guerra, L.S.; Garza-Guajardo, R.; Johnson-Pais, T.; Beuten, J.; Morales-Rodriguez, I.B.; Hernandez-Ordoñez, M.A.; Calderon-Cardenas, G.; Ortiz-Lopez, R.; et al. Identification of viral infections in the prostate and evaluation of their association with cancer. BMC Cancer 2010, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, A.; Kalantari, M.; Simoneau, A.; Jensen, J.L.; Villarreal, L.P. Detection of human polyomaviruses and papillomaviruses in prostatic tissue reveals the prostate as a habitat for multiple viral infections. Prostate 2002, 53, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ramnarine, V.R.; Bell, R.; Volik, S.; Davicioni, E.; Hayes, V.M.; Ren, S.; Collins, C.C. Metagenomic and metatranscriptomic analysis of human prostate microbiota from patients with prostate cancer. BMC Genom. 2019, 20, 146. [Google Scholar] [CrossRef]
- Liang, W.; Ferrara, N. The Complex Role of Neutrophils in Tumor Angiogenesis and Metastasis. Cancer Immunol. Res. 2016, 4, 83–91. [Google Scholar] [CrossRef]
- Yang, L.; Pang, Y.; Moses, H.L. TGF-beta and immune cells: An important regulatory axis in the tumor microenvironment and progression. Trends Immunol. 2010, 31, 220–227. [Google Scholar] [CrossRef]
- Salachan, P.V.; Rasmussen, M.; Fredsøe, J.; Ulhøi, B.; Borre, M.; Sørensen, K.D. Microbiota of the prostate tumor environment investigated by whole-transcriptome profiling. Genome Med. 2022, 14, 9. [Google Scholar] [CrossRef]
- Han, I.-H.; Song, H.-O.; Ryu, J.-S. IL-6 produced by prostate epithelial cells stimulated with Trichomonas vaginalis promotes proliferation of prostate cancer cells by inducing M2 polarization of THP-1-derived macrophages. PLoS Negl. Trop. Dis. 2020, 14, e0008126. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, B.; Devi, A.; Maikhuri, J.P.; Rajender, S.; Gupta, G. Inflammation driven tumor-like signaling in prostatic epithelial cells by sexually transmitted Trichomonas vaginalis. Int. J. Urol. 2021, 28, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Gnanasekar, A.; Lee, A.; Li, W.T.; Haas, M.; Wang-Rodriguez, J.; Chang, E.Y.; Rajasekaran, M.; Ongkeko, W.M. Influence of Intratumor Microbiome on Clinical Outcome and Immune Processes in Prostate Cancer. Cancers 2020, 12, 2524. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, N.; Lee, C. Mysteries of TGF-Î2 Paradox in Benign and Malignant Cells. Front. Oncol. 2014, 4, 94. [Google Scholar] [CrossRef] [PubMed]
- Menzies, B.E. The role of fibronectin binding proteins in the pathogenesis of Staphylococcus aureus infections. Curr. Opin. Infect. Dis. 2003, 16, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Nakayama, K.; Yoshimura, F.; Okamoto, K.; Abe, N.; Yamamoto, K. Arg-gingipain Acts as a Major Processing Enzyme for Various Cell Surface Proteins in Porphyromonas gingivalis. J. Biol. Chem. 1998, 273, 29072–29076. [Google Scholar] [CrossRef] [PubMed]
- Joshipura, K.J.; Rimm, E.B.; Douglass, C.W.; Trichopoulos, D.; Ascherio, A.; Willett, W.C. Poor Oral Health and Coronary Heart Disease. J. Dent. Res. 1996, 75, 1631–1636. [Google Scholar] [CrossRef]
- Michaud, D.S.; Izard, J.; Wilhelm-Benartzi, C.S.; You, D.-H.; Grote, V.A.; Tjønneland, A.; Dahm, C.C.; Overvad, K.; Jenab, M.; Fedirko, V.; et al. Plasma antibodies to oral bacteria and risk of pancreatic cancer in a large European prospective cohort study. Gut 2013, 62, 1764–1770. [Google Scholar] [CrossRef]
- Famili, P.; Cauley, J.A.; Greenspan, S.L. The Effect of Androgen Deprivation Therapy on Periodontal Disease in Men With Prostate Cancer. J. Urol. 2007, 177, 921–924. [Google Scholar] [CrossRef]
- Joshi, N.; Bissada, N.F.; Bodner, D.; MacLennan, G.T.; Narendran, S.; Jurevic, R.; Skillicorn, R. Association Between Periodontal Disease and Prostate-Specific Antigen Levels in Chronic Prostatitis Patients. J. Periodontol. 2010, 81, 864–869. [Google Scholar] [CrossRef]
- Noack, B.; Genco, R.J.; Trevisan, M.; Grossi, S.; Zambon, J.J.; Nardin, E. De Periodontal Infections Contribute to Elevated Systemic C-Reactive Protein Level. J. Periodontol. 2001, 72, 1221–1227. [Google Scholar] [CrossRef]
- Corbella, S.; Veronesi, P.; Galimberti, V.; Weinstein, R.; Del Fabbro, M.; Francetti, L. Is periodontitis a risk indicator for cancer? A meta-analysis. PLoS ONE 2018, 13, e0195683. [Google Scholar] [CrossRef]
- Nabil, F.; Bissada, N.A. Periodontal Treatment Improves Prostate Symptoms and Lowers Serum PSA in Men with High PSA and Chronic Periodontitis. Dentistry 2015, 5. [Google Scholar] [CrossRef]
- Boyapati, R.; Swarna, C.; Devulapalli, N.; Sanivarapu, S.; Katuri, K.; Kolaparthy, L. Unveiling the link between prostatitis and periodontitis. Contemp. Clin. Dent. 2018, 9, 524. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Michaud, D.S.; Lu, J.; Carter, H.B.; Platz, E.A. The association between clinically determined periodontal disease and prostate-specific antigen concentration in men without prostate cancer: The 2009–2010 National Health and Nutrition Examination Survey. Cancer Causes Control 2019, 30, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Kruck, S.; Hennenlotter, J.; Amend, B.; Geiger, M.; Filipova, E.; Stuhler, V.; Schubert, T.; Todenhofer, T.; Rausch, S.; Huettig, F.; et al. Chronic Periodontitis Does Not Impact Serum Levels of Prostate-specific Antigen. Anticancer Res. 2017, 37, 3163–3167. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.J.; Shannon, B.A.; McNeal, J.E.; Shannon, T.; Garrett, K.L. Proprionibacterium acnes associated with inflammation in radical prostatectomy specimens: A possiblew link to cancer evolution? J. Urol. 2005, 173, 1969–1974. [Google Scholar] [CrossRef]
- Alexeyev, O.A.; Marklund, I.; Shannon, B.; Golovleva, I.; Olsson, J.; Andersson, C.; Eriksson, I.; Cohen, R.; Elgh, F. Direct Visualization of Propionibacterium acnes in Prostate Tissue by Multicolor Fluorescent In Situ Hybridization Assay. J. Clin. Microbiol. 2007, 45, 3721–3728. [Google Scholar] [CrossRef]
- Drott, J.B.; Alexeyev, O.; Bergström, P.; Elgh, F.; Olsson, J. Propionibacterium acnes infection induces upregulation of inflammatory genes and cytokine secretion in prostate epithelial cells. BMC Microbiol. 2010, 10, 126. [Google Scholar] [CrossRef]
- Fassi Fehri, L.; Mak, T.N.; Laube, B.; Brinkmann, V.; Ogilvie, L.A.; Mollenkopf, H.; Lein, M.; Schmidt, T.; Meyer, T.F.; Brüggemann, H. Prevalence of Propionibacterium acnes in diseased prostates and its inflammatory and transforming activity on prostate epithelial cells. Int. J. Med. Microbiol. 2011, 301, 69–78. [Google Scholar] [CrossRef]


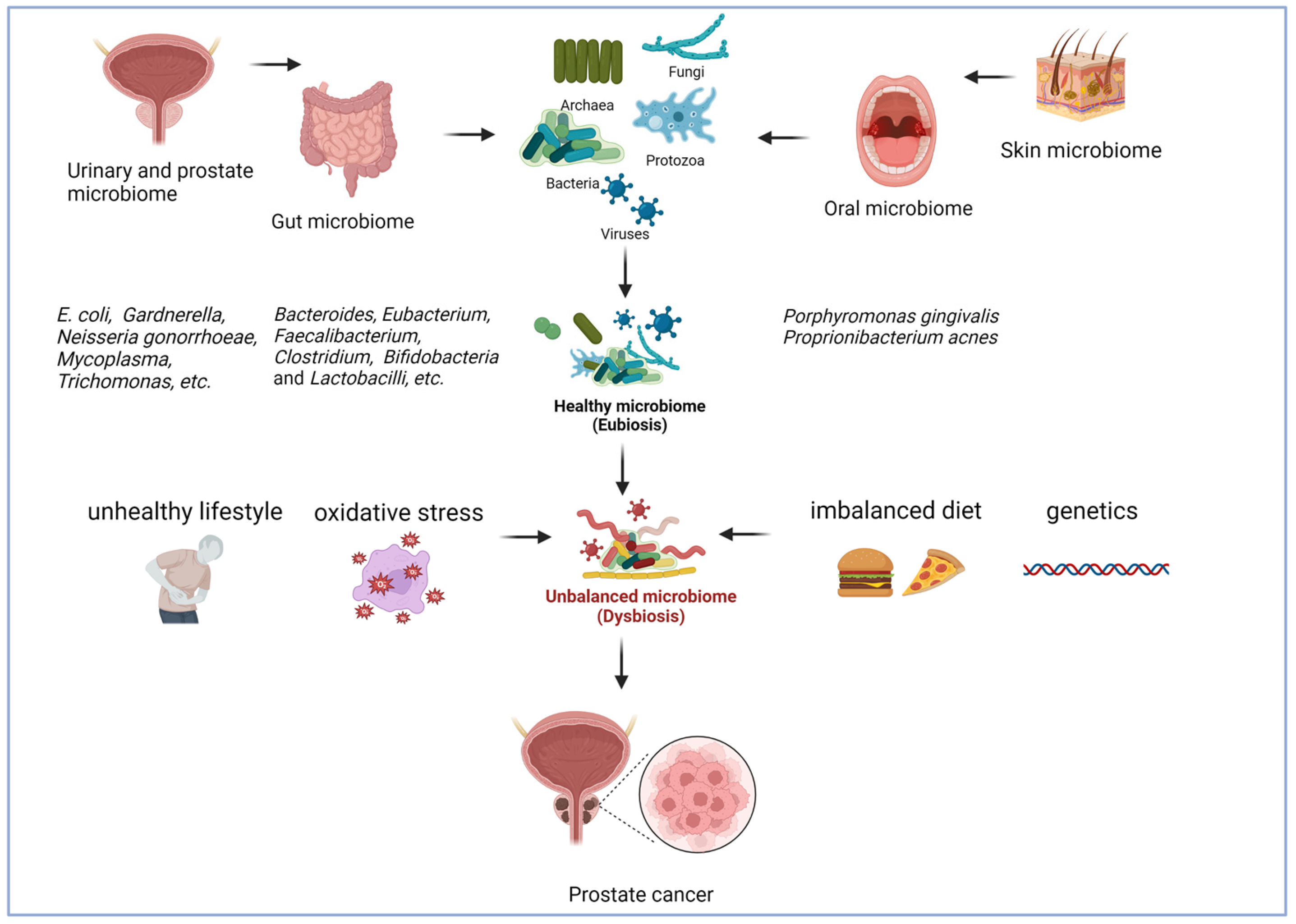{kind=link}
| Microorganisms | Isolation Site | Basic Involvement in Pca | Normal Tissue/BPH Samples | Pca Samples | Ref. |
|---|---|---|---|---|---|
| Bacteroides massiliensis | gut | processes complex molecules | low levels | high abundance | [17] |
| Eubacterium rectale | gut | major producer of butyrate | high concentration | low concentration | [17] |
| Faecalibacterium prausnitzii | gut | produces butyrate from its utilization of acetate anti-inflammatory properties | high concentration | low concentration | [17] |
| Clostridium scindens | gut | converts glucocorticoids into androgens | present | could interfere with androgen deprivation therapy | [37] |
| Akkermansiaceae, Ruminococcaceae spp. | Gut | response to anti-PD1 immunotherapy | present | widely present; high concentration in men with androgen deprivation therapy | [47] |
| Clostridium, Roseburia, Phascolarctobacterium spp. | Gut | short-chain fatty acid producers | present | high prevalence in radiation enteropathy patients | [50] |
| Bifidobacteria and Lactobacilli | gut | increase the abundance and activity of specific gut microbial organisms | present | decreased | [52] |
| Microorganisms | Isolation Site | Basic Involvement in PCa | Normal Tissue/BPH Samples | PCa Samples | Ref. |
|---|---|---|---|---|---|
| Proprionibacterium acnes | urine | proinflammatory | present | increased in prostatitis, chronic inflammation, and PCa | [60] |
| Porphyromonas sp., Varibaculum sp., Peptoniphilus sp., and Fenollaria sp. | urine and prostate secretion | prognostic value for PCa | present | high in patients with high-risk PCa | [80] |
| Bradyrhizobium japonicum, Listeria monocytogenes, Methylobacterium radiotolerans, Pseudomonas aeruginosa, Stenotrophomonas maltophilia and Xanthomonas albilineans | urine | anticancer properties | present | low | [81] |
| Propionibacterium spp. and Staphylococcus spp. | Urine | infectious | absent | present | [81] |
| E. coli, Campylobacter concisus, Streptococcus pneumoniae, Nevskia ramosa, Staphylococcus aureus, Paraburkaholderia phymatum, Gardnerella vaginalis, Nitrobacter hamburgensis | urine | chronic inflammation, immune imbalance, DNA damage | absent | present | [81] |
| Neisseria gonorrhoeae and Chlamydia trachomatis | urine | infectious | absent | present | [77] |
| Streptococcus anginosus, Anaerococcus lactolyticus, Anaerococcus obesiensis, Actinobaculum schaalii, Varibaculum cambriense and Propionimicrobium lymphophilum | urine | pro-inflammatory | absent | abundant | [82] |
| Veillonella, Streptococcus, Bacteroides | urine | part of the normal flora of the mouth and gastrointestinal tract | low | abundant | [83] |
| Aecalibacterium, Lactobacilli, Actinetobacter | urine | abundant and important commensal bacteria | present | decreased | [83] |
| Microorganisms | Isolation Site | Basic Involvement in PCa | Normal Tissue/BPH Samples | PCa Samples | Ref. |
|---|---|---|---|---|---|
| Trichomonas vaginalis and Chlamydia trachomatis | prostate | M2 macrophage polarization affects mRNA expression for IL-6 and FGF-2 | absent | present | [81] |
| Mycoplasma genitalium | prostate | causes sexual transmitted diseases | absent | present | [89] |
| Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes | prostate | infection | absent | abundant | [90] |
| Polyomaviruses, human papillomaviruses (HPVs), human cytomegalovirus | prostate | infection | absent | present | [91] |
| Porphyromonas gingivalis | prostatic secretion | secreting Arg-gingipain, a collagen-degrading substance | absent | present | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munteanu, R.; Feder, R.-I.; Onaciu, A.; Munteanu, V.C.; Iuga, C.-A.; Gulei, D. Insights into the Human Microbiome and Its Connections with Prostate Cancer. Cancers 2023, 15, 2539. https://doi.org/10.3390/cancers15092539
Munteanu R, Feder R-I, Onaciu A, Munteanu VC, Iuga C-A, Gulei D. Insights into the Human Microbiome and Its Connections with Prostate Cancer. Cancers. 2023; 15(9):2539. https://doi.org/10.3390/cancers15092539
Chicago/Turabian StyleMunteanu, Raluca, Richard-Ionut Feder, Anca Onaciu, Vlad Cristian Munteanu, Cristina-Adela Iuga, and Diana Gulei. 2023. "Insights into the Human Microbiome and Its Connections with Prostate Cancer" Cancers 15, no. 9: 2539. https://doi.org/10.3390/cancers15092539