
Hypoxia, a Targetable Culprit to Counter Pancreatic Cancer Resistance to Therapy
 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Role of Hypoxia in PDAC Carcinogenesis
3. Impact of Hypoxic Stress on Antigen-Specific Cell-Mediated Cytotoxicity
4. Genomic Instability in the Context of Hypoxia Influencing the PDAC Immune Response
5. Hypoxia-Centered Combination Therapy in PDAC
5.1. Targeting Tumor Hypoxia in PDAC
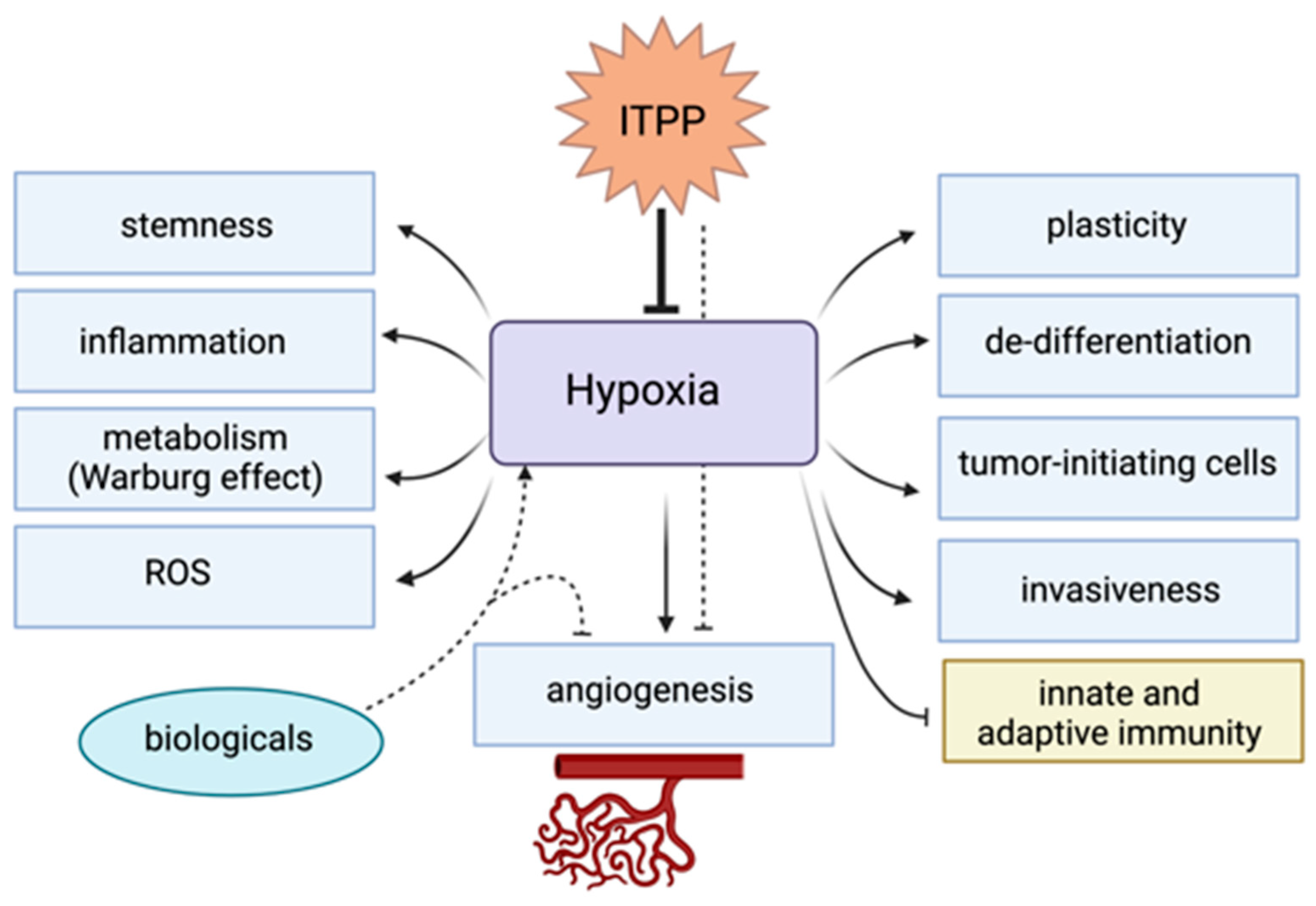
5.2. Antihypoxic Therapy Using Myo-Inositol-Trispyrophosphate (ITPP)
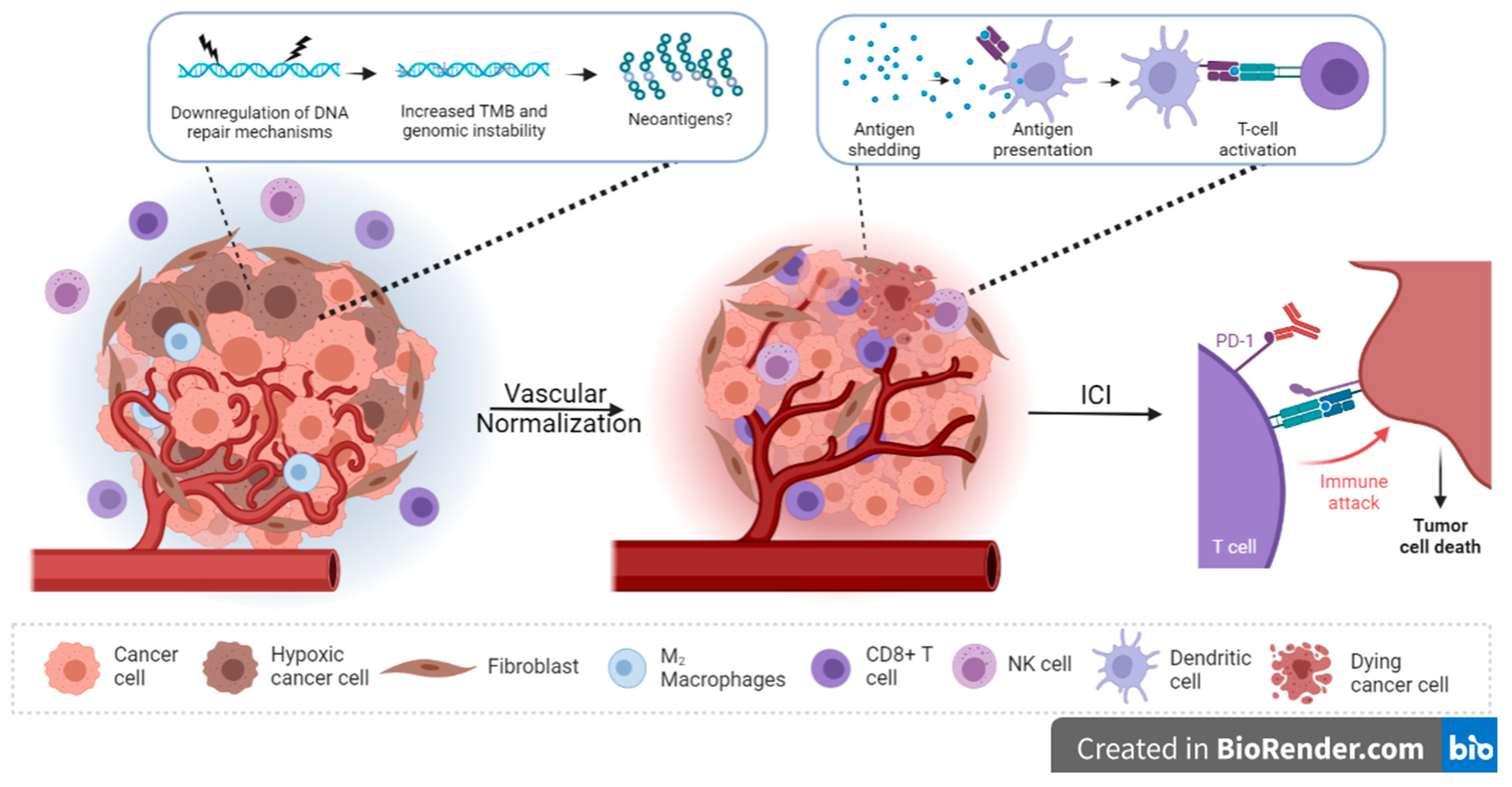
5.3. Preclincal Evidence Supporting the Manipulation of Hypoxia to Enhance Responses to Immunotherapy

{kind=link}
{kind=link}
| Target | Drug a | Mouse Model b | Combination Therapy | Immune Modulation | Efficacy of Combination | Ref. |
|---|---|---|---|---|---|---|
| HIF-1α | PX-478 | Panc-1, CF-PAC-1 or SU.86.86 subcutaneously implanted in the flank of female SCID mice | Fractionated radiation therapy, with or without the combined treatment with 5-fluorouracil or gemcitabine | NA | Significant increase in tumor regression, potentiating the antitumor activity of radiation and chemotherapy | [113] |
| HIF-1α | PX-478 | Panc02 subcutaneously implanted in the flank of C57BL/6 mice and immuno-incompetent nude (Nu/Nu) mice; Subcutaneous inoculation in the flank of Panc02 dying and dead cells and the supernatant from cells untreated or treated with Gem and/or PX-478 (vaccination) followed by subcutaneous implantation in the flank of surviving cells (challenge) | Gemcitabine | Increased cytotoxic CD3+/CD8+ T lymphocytes in the spleen and tumor tissues in mice compared to the single treatment | Significant reduction in tumor growth in immune-competent and incompetent mice with the single treatment; Increased tumor suppression effect in immune-competent but not in nude mice compared with the Gem single treatment; high vaccine efficacy, decreasing tumor growth by inducing immunogenic cell death | [80] |
| HIF-1α downstream signaling (LOX) | LOX-blocking antibody | PDAC-bearing KPC mice | Gemcitabine | Increased leukocyte, macrophage, and neutrophil infiltration compared to the single treatment | Increased survival and decreased metastatic burden | [120] |
| Microenvironmental hypoxia | TH-302 | Patient-derived pancreatic xenografts subcutaneously implanted in the leg | Fractionated radiation therapy | NA | Decreased tumor growth in fast-growing tumors only | [114] |
| Microenvironmental hypoxia | TH-302 | AsPC1 cells orthotopically implanted in the pancreas of (Nu/Nu) mice | Single-dose radiation therapy | NA | Significantly more effective in delaying tumor growth than the single therapy | [115] |
| HIF-1α downstream signaling (CA9) | SLC-0111 | PK-8 or PK-1 cells subcutaneously implanted on the back of female NOD/SCID or NSG mice; Patient-derived pancreatic xenografts—subcutaneous implantation of tissue fragments into male C.B-17 SCID mice; PDAC-bearing KPCY mice | Gemcitabine | Decreased B220+ B cells with no impact on the number of CD3+ T cells in the combination treatment compared to single treatment | Decreased tumor growth and increased survival | [116] |
| HIF-1α | VHH212 nanobody | PANC-1 cells subcutaneously implanted in the flank of BALB/c nude mice | Gemcitabine | NA | Higher inhibition of tumor growth compared with gemcitabine alone | [117] |
| Microenvironmental hypoxia | Liposomal vinblastine-N-Oxide (CPD100Li) | PANC-1 cells subcutaneously implanted in the tail of female Nu/Nu mice | Gemcitabine | NA | Significant decrease in tumor growth compared to gemcitabine alone | [118] |
| Microenvironmental acidosis and hypoxia | Gold nanorods (GNRs) | KPC tumor cells subcutaneously implanted in male C57BL/6 mice; Luciferase-transfected KPC tumor cells orthotopically implanted in the pancreas of male C57BL/6 mice | Single-dose radiation therapy | NA | Significantly more effective in delaying tumor growth and decreasing tumor volume than the single therapy | [119] |
| HIF-2α | PT2399 | KPC cells subcutaneously implanted into the flank of syngeneic C57BL/6 female mice; KPC cells were orthotopically implanted into the tail of the pancreas of syngeneic C57BL/6 male mice | ICI | NA c | Combination treatment with anti-CTLA4 led to a significant decrease in tumor growth in the tested subcutaneous model compared to each drug alone; dual checkpoint blockade (anti-PD1 and anti-CTLA4) combination with PT2399 led to decreased tumor growth and enhanced survival in the orthotopic model (combined with anti-pd1) | [34] |
| Microenvironmental hypoxia | Oxygen microcapsules | KPC cells subcutaneously implanted into the flank of syngeneic C57BL/6 female mice; KPC cells were orthotopically implanted into the tail of the pancreas of syngeneic C57BL/6 male mice | ICI | Increased the infiltration of CD45+ immune cells and increased the proportion of M1 macrophages with no effect on MDSC infiltration compared to the single treatment | Combination treatment with anti-PD1 led to significant decreases in tumor growth compared to each drug alone | [108] |
6. Detection of Hypoxia in the TME of Patients with PDAC
6.1. Direct Oxygen Quantification
6.2. Immunohistochemistry (IHC)-Based Detection of Hypoxia-Related Markers
6.3. Imaging-Based Parameters as Hypoxia Biomarkers
6.4. Gene Signatures as Hypoxia Surrogates
| Signature and Cohort Characteristics | Survival Analysis a | Immune Analysis b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genes | Derivation | Scoring | Cohort | Groups (Patient Number) c | End Point | KM (p-Value) | Univariate Cox PH | Multivariate Cox PH | Method | High-Risk Group (Hypoxia-High) | Low-Risk Group (Hypoxia-Low) | Ref. |
| 30 d | Overlap between 200 genes of the hallmark HYPOXIA gene-set and microarray data of two pancreatic cancer cohorts (GSE15471 and GSE16515)—30 DEGs | Gene score: +1 if gene expression > median expression in entire cohort; −1 if < median expression in entire cohort. Hypoxia score is sum of 30 genes | PAAD TCGA | High (79) vs. Low (98) | OS PFS | 0.0062 0.0024 | NA | NA | NA | NA | NA | [141] |
| 8 (DDIT4, LDHA, MXI1, NDRG1, P4HA1, PGK1, SLC2A1, VEGFA) | Expression of 15 genes selected from 398 hypoxia genes collected from published prognostic or predictive signatures tested in 14 cancer cell lines exposed to 1% oxygen | Gene score: +1 if gene expression > median expression in entire cohort; −1 if <median expression in entire cohort. Hypoxia score is sum of 8 genes | PAAD TCGA | High (66) vs. Low (98) | OS DSS PFS | 0.0035 0.0047 0.01 | 1.9 (1.2–2.9) p = 0.004 2 (1.2–3.2) p = 0.005 1.7 (1.1–2.5) p = 0.011 | 1.7 (1.10–2.7) p = 0.016 1.6 (0.99–2.6) p = 0.056 1.5 (0.97–2.2) p = 0.067 | CIBERSORTx Immune score Cytolytic index 4-chemokine signature | M0 macrophages, low cytolytic index, low immune score and low chemokine score | CD8+ T cells, high cytolytic index, high immune score and high chemokine score | [23] |
| E-MTAB-6134 | High (136) vs. Low (173) | OS DFS | <0.0001 <0.0001 | 2.1 (1.6–2.8) p < 0.001 1.8 (1.3–2.3) p < 0.001 | 2.19 (1.60–3.0) p < 0.001 1.8 (1.39–2.5) p < 0.001 | |||||||
| 9 (ARNTL1, DCBLD2, DSG3, FAM83A, FOXM1, GZMK, IGF2BP2, SLC38A11, TPX2) | 15 overexpressed HIF-1 related genes in meta-PDAC cohort (GSE62452 and PAAD TCGA)—nine showed critical prognosis association using LASSO regression analysis | Multiplying expression of nine genes with their corresponding multivariable Cox regression coefficient—classification into high-, medium- and low-score based on cutoffs determined by X-tile 3.6.1 software | Meta-PDAC cohort | High (22) vs. Medium (73) vs. Low (110) | OS | 5.584 × 10−14 | 2.276 (1.741–2.975) p < 0.001 | 2.162 (1.632–2.865) p < 0.001 | Enrichment scores of 25 immune-related terms determined from previous studies in the meta-PDAC cohort only immunostaining for CD8+ T cells in 28 PDACs sorted into low- and high-HIF-1 scores based on median cutoff of HIF-1 scores determined using RT-qPCR | TIL, activated CD8+ T cells, cytolytic activity, activated B cell, immature B cell and Type 1 T-helper cells significantly more enriched in low-score group. High-HIF-1 score inversely correlated with CD8+ T cell density | [24] | |
| PDAC ICGC | High vs. Medium vs. Low | OS | 2.436 × 10−05 | NA | NA | |||||||
| GSE79668 | High-risk vs. Low-risk | OS | 1.246 × 10−04 | NA | NA | |||||||
| 4 (ENO3, LDHA, PGK1, PGM1) | Network analysis of protein interactions of 200 genes of hallmark HYPOXIA gene-set—50 DEGs with highest interaction- 4 DEGs maintained association with survival following multivariate Cox regression analysis | Multiplying expression of nine genes with their corresponding multivariable Cox regression coefficient—classification into high- and low-hypoxia risk score based on the median risk score | PAAD TCGA | High-risk (88) vs. Low-risk (89) | OS | <0.001 | 1.986 (1.579–2.498) p < 0.001 | 1.878 (1.498–2.354) p < 0.001 | CIBERSORT Expression of genes unfavorably regulating immune-related processes. Expression of genes positively regulating T cells, DCs and MDSCs | Resting NK cells Higher expression of VEGFA, MICB and ICAM1. Higher expression of CXCL5 | CD8+ T cells, and naive B cells Higher expression of CCL21 and CCR7 | [142] |
| GSE78229 and GSE57495 | High-risk (58) vs. Low-risk (54) | OS | 0.024 | 1.410 (1.190–1.670) p < 0.001 | 1.622 (1.050–2.507) p = 0.029 | |||||||
| 8 (ANKZF1, CITED, ENO3, JMJD6, LDHA, NDST1, SIAH2, TES) | Correlation between 200 genes of hallmark HYPOXIA gene-set and RNA-seq data of PAAD TCGA cohort—108 DEGs were correlated—45 DEGs were associated with OS based on univariate Cox regression analysis—eight maintained association based on LASSO regression analysis | Multiplying expression of eight genes with their corresponding LASSO coefficient—classification into high- and low-hypoxia risk score based on the median risk score | PAAD TCGA | High-risk (81) vs. Low-risk (81) | OS | <0.0001 | 2.508 (1.575–3.992) p < 0.0001 | 2.503 (1.483–4.226) p < 0.0001 | CIBERSORT (applied only in TCGA cohort) Expression of immune checkpoint genes (applied only in TCGA cohort) | Neutrophils with higher expression of CD47 | Treg higher expression of BTLA, CTLA4, LAG3, TNFRSF4 and PDCD1 | [143] |
| GSE62452 | High-risk (33) vs. Low-risk (32) | OS | 0.00075 | NA | NA | |||||||
| 3 (ANXA2, LDHA, TES) | Overlap between 200 genes of hallmark HYPOXIA gene-set and RNA seq data of PAAD TCGA cohort—67 DEGs correlated with OS based on univariate Cox regression analysis—three maintained association with survival following multivariate Cox regression analysis | Multiplying expression of three genes with their corresponding multivariable Cox regression coefficient—classification into high- and low-hypoxia risk score based on the median risk score | PAAD TCGA | High-risk vs. Low-risk | OS | 0.00061 | 2.5746 (1.6083–4.122) p < 0.001 | NA | CIBERSORT | M0 macrophages, monocytes (ICGC and GSE57495) | CD8+ T cells (TCGA and ICGC), naïve B cells (TCGA and GSE57495) | [144] |
| PDAC ICGC | High-risk vs. Low-risk | OS | 0.004 | 3.0760 (1.7135–5.522) p < 0.001 | NA | |||||||
| GSE57495 | High-risk vs. Low-risk | OS | 0.031 | NA | NA | |||||||
7. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ducreux, M.; Cuhna, A.S.; Caramella, C.; Hollebecque, A.; Burtin, P.; Goéré, D.; Seufferlein, T.; Haustermans, K.; Van Laethem, J.L.; Conroy, T.; et al. Cancer of the pancreas: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26 (Suppl. S5), v56–v68. [Google Scholar] [CrossRef] [PubMed]
- Christenson, E.S.; Jaffee, E.; Azad, N.S. Current and emerging therapies for patients with advanced pancreatic ductal adenocarcinoma: A bright future. Lancet Oncol. 2020, 21, e135–e145. [Google Scholar] [CrossRef] [PubMed]
- Collaborators GPC. The global, regional, and national burden of pancreatic cancer and its attributable risk factors in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2019, 4, 934–947. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Lok, V.; Ngai, C.H.; Zhang, L.; Yuan, J.; Lao, X.Q.; Ng, K.; Chong, C.; Zheng, Z.-J.; Wong, M.C. Worldwide Burden of, Risk Factors for, and Trends in Pancreatic Cancer. Gastroenterology 2021, 160, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Bockorny, B.; Grossman, J.E.; Hidalgo, M. Facts and Hopes in Immunotherapy of Pancreatic Cancer. Clin. Cancer Res. 2022, 28, 4606–4617. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.-J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-Year Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2019, 381, 1535–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, L.; Rodríguez-Abreu, D.; Gadgeel, S.; Esteban, E.; Felip, E.; De Angelis, F.; Domine, M.; Clingan, P.; Hochmair, M.J.; Powell, S.F.; et al. Pembrolizumab plus Chemotherapy in Metastatic Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 378, 2078–2092. [Google Scholar] [CrossRef] [PubMed]
- Padrón, L.J.; Maurer, D.M.; O’Hara, M.H.; O’Reilly, E.M.; Wolff, R.A.; Wainberg, Z.A.; Ko, A.H.; Fisher, G.; Rahma, O.; Lyman, J.P.; et al. Sotigalimab and/or nivolumab with chemotherapy in first-line metastatic pancreatic cancer: Clinical and immunologic analyses from the randomized phase 2 PRINCE trial. Nat. Med. 2022, 28, 1167–1177. [Google Scholar] [CrossRef]
- Samstein, R.M.; Lee, C.-H.; Shoushtari, A.N.; Hellmann, M.D.; Shen, R.; Janjigian, Y.Y.; Barron, D.A.; Zehir, A.; Jordan, E.J.; Omuro, A.; et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat. Genet. 2019, 51, 202–206. [Google Scholar] [CrossRef]
- Hu, Z.I.; Shia, J.; Stadler, Z.K.; Varghese, A.M.; Capanu, M.; Salo-Mullen, E.; Lowery, M.A.; Diaz, L.A.; Mandelker, D.; Yu, K.H.; et al. Evaluating Mismatch Repair Deficiency in Pancreatic Adenocarcinoma: Challenges and Recommendations. Clin. Cancer Res. 2018, 24, 1326–1336. [Google Scholar] [CrossRef]
- Luchini, C.; Brosens, L.A.A.; Wood, L.D.; Chatterjee, D.; Shin, J.I.; Sciammarella, C.; Fiadone, G.; Malleo, G.; Salvia, R.; Kryklyva, V.; et al. Comprehensive characterisation of pancreatic ductal adenocarcinoma with microsatellite instability: Histology, molecular pathology and clinical implications. Gut 2021, 70, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Connor, A.A.; Gallinger, S. Pancreatic cancer evolution and heterogeneity: Integrating omics and clinical data. Nat. Rev. Cancer 2022, 22, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Philip, P.A.; Azar, I.; Xiu, J.; Hall, M.J.; Hendifar, A.E.; Lou, E.; Hwang, J.J.; Gong, J.; Feldman, R.; Ellis, M.; et al. Molecular Characterization of KRAS Wild-type Tumors in Patients with Pancreatic Adenocarcinoma. Clin. Cancer Res. 2022, 28, 2704–2714. [Google Scholar] [CrossRef] [PubMed]
- Connor, A.A.; Denroche, R.E.; Jang, G.H.; Timms, L.; Kalimuthu, S.N.; Selander, I.; McPherson, T.; Wilson, G.W.; Chan-Seng-Yue, M.; Borozan, I.; et al. Association of Distinct Mutational Signatures With Correlates of Increased Immune Activity in Pancreatic Ductal Adenocarcinoma. JAMA Oncol. 2017, 3, 774–783. [Google Scholar] [CrossRef]
- Fraune, C.; Burandt, E.; Simon, R.; Hube-Magg, C.; Makrypidi-Fraune, G.; Kluth, M.; Büscheck, F.; Höflmayer, D.; Blessin, N.C.; Mandelkow, T.; et al. MMR Deficiency is Homogeneous in Pancreatic Carcinoma and Associated with High Density of Cd8-Positive Lymphocytes. Ann. Surg. Oncol. 2020, 27, 3997–4006. [Google Scholar] [CrossRef] [Green Version]
- Golan, T.; O’Kane, G.M.; Denroche, R.E.; Raitses-Gurevich, M.; Grant, R.C.; Holter, S.; Wang, Y.; Zhang, A.; Jang, G.H.; Stossel, C.; et al. Genomic Features and Classification of Homologous Recombination Deficient Pancreatic Ductal Adenocarcinoma. Gastroenterology 2021, 160, 2119–2132. [Google Scholar] [CrossRef]
- Waddell, N.; Pajic, M.; Patch, A.-M.; Chang, D.K.; Kassahn, K.S.; Bailey, P.; Johns, A.L.; Miller, D.; Nones, K.; Quek, K.; et al. Whole genomes redefine the mutational landscape of pancreatic cancer. Nature 2015, 518, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Steele, C.D.; Abbasi, A.; Islam, S.M.A.; Bowes, A.L.; Khandekar, A.; Haase, K.; Hames-Fathi, S.; Ajayi, D.; Verfaillie, A.; Dhami, P.; et al. Signatures of copy number alterations in human cancer. Nature 2022, 606, 984–991. [Google Scholar] [CrossRef]
- Hassan Venkatesh, G.; Abou Khouzam, R.; Shaaban Moustafa Elsayed, W.; Ahmed Zeinelabdin, N.; Terry, S.; Chouaib, S. Hypoxia increases mutational load of breast cancer cells through frameshift mutations. Oncoimmunology 2020, 9, 1750750. [Google Scholar] [CrossRef] [Green Version]
- Hassan Venkatesh, G.; Abou Khouzam, R.; Shaaban Moustafa Elsayed, W.; Ahmed Zeinelabdin, N.; Terry, S.; Chouaib, S. Tumor hypoxia: An important regulator of tumor progression or a potential modulator of tumor immunogenicity? Oncoimmunology 2021, 10, 1974233. [Google Scholar] [CrossRef]
- Bhandari, V.; Hoey, C.; Liu, L.Y.; Lalonde, E.; Ray, J.; Livingstone, J.; Lesurf, R.; Shiah, Y.-J.; Vujcic, T.; Huang, X.; et al. Molecular landmarks of tumor hypoxia across cancer types. Nat. Genet. 2019, 51, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, V.; Li, C.H.; Bristow, R.G.; Boutros, P.C.; Aaltonen, L.A.; Abascal, F.; Abeshouse, A.; Aburatani, H.; Adams, D.J.; Agrawal, N.; et al. Divergent mutational processes distinguish hypoxic and normoxic tumours. Nat. Commun. 2020, 11, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou Khouzam, R.; Rao, S.P.; Venkatesh, G.H.; Zeinelabdin, N.A.; Buart, S.; Meylan, M.; Chouaib, S. An Eight-Gene Hypoxia Signature Predicts Survival in Pancreatic Cancer and Is Associated with an Immunosuppressed Tumor Microenvironment. Front. Immunol. 2021, 12, 680435. [Google Scholar] [CrossRef]
- Zhuang, H.; Wang, S.; Chen, B.; Zhang, Z.; Ma, Z.; Li, Z.; Liu, C.; Zhou, Z.; Gong, Y.; Huang, S.; et al. Prognostic Stratification Based on HIF-1 Signaling for Evaluating Hypoxic Status and Immune Infiltration in Pancreatic Ductal Adenocarcinomas. Front. Immunol. 2021, 12, 790661. [Google Scholar] [CrossRef]
- Erkan, M.; Kurtoglu, M.; Kleeff, J. The role of hypoxia in pancreatic cancer: A potential therapeutic target? Expert. Rev. Gastroenterol. Hepatol. 2016, 10, 301–316. [Google Scholar] [CrossRef]
- Tao, J.; Yang, G.; Zhou, W.; Qiu, J.; Chen, G.; Luo, W.; Zhao, F.; You, L.; Zheng, L.; Zhang, T.; et al. Targeting hypoxic tumor microenvironment in pancreatic cancer. J. Hematol. Oncol. 2021, 14, 14. [Google Scholar] [CrossRef]
- Samanta, D.; Semenza, G.L. Metabolic adaptation of cancer and immune cells mediated by hypoxia-inducible factors. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2018, 1870, 15–22. [Google Scholar] [CrossRef]
- Daniel, S.K.; Sullivan, K.M.; Labadie, K.P.; Pillarisetty, V.G. Hypoxia as a barrier to immunotherapy in pancreatic adenocarcinoma. Clin. Transl. Med. 2019, 8, 10. [Google Scholar] [CrossRef]
- Abou Khouzam, R.; Brodaczewska, K.; Filipiak, A.; Zeinelabdin, N.A.; Buart, S.; Szczylik, C.; Chouaib, S. Tumor Hypoxia Regulates Immune Escape/Invasion: Influence on Angiogenesis and Potential Impact of Hypoxic Biomarkers on Cancer Therapies. Front. Immunol. 2020, 11, 613114. [Google Scholar] [CrossRef]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1α and HIF2α: Sibling rivalry in hypoxic tumour growth and progression. Nat. Rev. Cancer 2011, 12, 9–22. [Google Scholar] [CrossRef]
- Toledo, R.A.; Jimenez, C.; Armaiz-Pena, G.; Arenillas, C.; Capdevila, J.; Dahia, P.L.M. Hypoxia-Inducible Factor 2 Alpha (HIF2α) Inhibitors: Targeting Genetically Driven Tumor Hypoxia. Endocr. Rev. 2022, bnac025. [Google Scholar] [CrossRef]
- Zhang, Q.; Lou, Y.; Zhang, J.; Fu, Q.; Wei, T.; Sun, X.; Liang, T. Hypoxia-inducible factor-2alpha promotes tumor progression and has crosstalk with Wnt/beta-catenin signaling in pancreatic cancer. Mol. Cancer 2017, 16, 119. [Google Scholar] [CrossRef] [Green Version]
- Criscimanna, A.; Duan, L.J.; Rhodes, J.A.; Fendrich, V.; Wickline, E.; Hartman, D.J.; Esni, F. PanIN-specific regulation of Wnt signaling by HIF2alpha during early pancreatic tumorigenesis. Cancer Res. 2013, 73, 4781–4790. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.J.G.; Huang, Y.; Fuentes, N.R.; Turner, M.C.; Monberg, M.E.; Lin, D.; Taniguchi, C.M. Stromal HIF2 Regulates Immune Suppression in the Pancreatic Cancer Microenvironment. Gastroenterology 2022, 162, 2018–2031. [Google Scholar] [CrossRef]
- Hasmim, M.; Janji, B.; Khaled, M.; Noman, M.Z.; Louache, F.; Bordereaux, D.; Abderamane, A.; Baud, V.; Mami-Chouaib, F.; Chouaib, S. Cutting Edge: NANOG Activates Autophagy under Hypoxic Stress by Binding to BNIP3L Promoter. J. Immunol. 2017, 198, 1423–1428. [Google Scholar] [CrossRef] [Green Version]
- Hasmim, M.; Noman, M.Z.; Messai, Y.; Bordereaux, D.; Gros, G.; Baud, V.; Chouaib, S. Cutting Edge: Hypoxia-Induced Nanog Favors the Intratumoral Infiltration of Regulatory T Cells and Macrophages via Direct Regulation of TGF-β1. J. Immunol. 2013, 191, 5802–5806. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, P.; Gilkes, D.M.; Takano, N.; Semenza, G.L. Hypoxia-inducible factor-dependent signaling between triple-negative breast cancer cells and mesenchymal stem cells promotes macrophage recruitment. Proc. Natl. Acad. Sci. USA 2014, 111, E2120–E2129. [Google Scholar] [CrossRef] [Green Version]
- Akalay, I.; Tan, T.Z.; Kumar, P.; Janji, B.; Mami-Chouaib, F.; Charpy, C.; Vielh, P.; Larsen, A.K.; Thiery, J.P.; Sabbah, M.; et al. Targeting WNT1-inducible signaling pathway protein 2 alters human breast cancer cell susceptibility to specific lysis through regulation of KLF-4 and miR-7 expression. Oncogene 2015, 34, 2261–2271. [Google Scholar] [CrossRef]
- Akalay, I.; Janji, B.; Hasmim, M.; Noman, M.Z.; André, F.; De Cremoux, P.; Bertheau, P.; Badoual, C.; Vielh, P.; Larsen, A.K.; et al. Epithelial-to-Mesenchymal Transition and Autophagy Induction in Breast Carcinoma Promote Escape from T-cell–Mediated Lysis. Cancer Res. 2013, 73, 2418–2427. [Google Scholar] [CrossRef] [Green Version]
- Noman, M.Z.; Janji, B.; Hu, S.; Wu, J.C.; Martelli, F.; Bronte, V.; Chouaib, S. Tumor-Promoting Effects of Myeloid-Derived Suppressor Cells Are Potentiated by Hypoxia-Induced Expression of miR-210. Cancer Res. 2015, 75, 3771–3787. [Google Scholar] [CrossRef]
- Noman, M.Z.; Desantis, G.; Janji, B.; Hasmim, M.; Karray, S.; Dessen, P.; Bronte, V.; Chouaib, S. PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J. Exp. Med. 2014, 211, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Chen, C.; Song, Y.; Cai, Q.; Li, J.; Tang, Y.; Han, X.; Qu, W.; Chen, A.; Wang, H.; et al. Hypoxia modifies the polarization of macrophages and their inflammatory microenvironment, and inhibits malignant behavior in cancer cells. Oncol. Lett. 2019, 18, 5871–5878. [Google Scholar] [CrossRef] [Green Version]
- Suthen, S.D.; Lim, C.J.; Nguyen, P.H.D.; Dutertre, C.; Lai, H.L.H.; Wasser, M.; Chua, C.; Lim, T.K.H.; Leow, W.Q.; Loh, T.J.; et al. Hypoxia-driven immunosuppression by Treg and type-2 conventional dendritic cells in HCC. Hepatology 2022, 76, 1329–1344. [Google Scholar] [CrossRef]
- Ziani, L.; Buart, S.; Chouaib, S.; Thiery, J. Hypoxia increases melanoma-associated fibroblasts immunosuppressive potential and inhibitory effect on T cell-mediated cytotoxicity. Oncoimmunology 2021, 10, 1950953. [Google Scholar] [CrossRef]
- Begg, K.; Tavassoli, M. Inside the hypoxic tumour: Reprogramming of the DDR and radioresistance. Cell Death Discov. 2020, 6, 1–15. [Google Scholar] [CrossRef]
- Kaplan, A.; Glazer, P.M. Impact of hypoxia on DNA repair and genome integrity. Mutagenesis 2020, 35, 61–68. [Google Scholar] [CrossRef]
- Pires, I.M.; Bencokova, Z.; Milani, M.; Folkes, L.K.; Li, J.-L.; Stratford, M.R.; Harris, A.L.; Hammond, E.M. Effects of Acute versus Chronic Hypoxia on DNA Damage Responses and Genomic Instability. Cancer Res. 2010, 70, 925–935. [Google Scholar] [CrossRef] [Green Version]
- Bader, S.B.; Ma, T.S.; Simpson, C.J.; Liang, J.; Maezono, S.E.B.; Olcina, M.M.; Buffa, F.M.; Hammond, E.M. Replication catastrophe induced by cyclic hypoxia leads to increased APOBEC3B activity. Nucleic Acids Res. 2021, 49, 7492–7506. [Google Scholar] [CrossRef]
- Wörmann, S.M.; Zhang, A.; Thege, F.I.; Cowan, R.W.; Rupani, D.N.; Wang, R.; Manning, S.L.; Gates, C.; Wu, W.; Levin-Klein, R.; et al. APOBEC3A drives deaminase domain-independent chromosomal instability to promote pancreatic cancer metastasis. Nat. Cancer 2021, 2, 1338–1356. [Google Scholar] [CrossRef]
- Connor, A.A.; Denroche, R.E.; Jang, G.H.; Lemire, M.; Zhang, A.; Chan-Seng-Yue, M.; Wilson, G.; Grant, R.C.; Merico, D.; Lungu, I.; et al. Integration of Genomic and Transcriptional Features in Pancreatic Cancer Reveals Increased Cell Cycle Progression in Metastases. Cancer Cell 2019, 35, 267–282.e7. [Google Scholar] [CrossRef]
- Bailey, P.; Chang, D.K.; Forget, M.-A.; Lucas, F.A.S.; Alvarez, H.A.; Haymaker, C.; Chattopadhyay, C.; Kim, S.-H.; Ekmekcioglu, S.; Grimm, E.A.; et al. Exploiting the neoantigen landscape for immunotherapy of pancreatic ductal adenocarcinoma. Sci. Rep. 2016, 6, 35848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germano, G.; Lamba, S.; Rospo, G.; Barault, L.; Magrì, A.; Maione, F.; Russo, M.; Crisafulli, G.; Bartolini, A.; Lerda, G.; et al. Inactivation of DNA repair triggers neoantigen generation and impairs tumour growth. Nature 2017, 552, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Rospo, G.; Lorenzato, A.; Amirouchene-Angelozzi, N.; Magrì, A.; Cancelliere, C.; Corti, G.; Negrino, C.; Amodio, V.; Montone, M.; Bartolini, A.; et al. Evolving neoantigen profiles in colorectal cancers with DNA repair defects. Genome Med. 2019, 11, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalmers, Z.R.; Connelly, C.F.; Fabrizio, D.; Gay, L.; Ali, S.M.; Ennis, R.; Schrock, A.; Campbell, B.; Shlien, A.; Chmielecki, J.; et al. Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden. Genome Med. 2017, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balachandran, V.P.; Łuksza, M.; Zhao, J.N.; Makarov, V.; Moral, J.A.; Remark, R.; Herbst, B.; Askan, G.; Bhanot, U.; Sendabaoglu, Y.; et al. Identification of unique neoantigen qualities in long-term survivors of pancreatic cancer. Nature 2017, 551, 512–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łuksza, M.; Sethna, Z.M.; Rojas, L.A.; Lihm, J.; Bravi, B.; Elhanati, Y.; Soares, K.; Amisaki, M.; Dobrin, A.; Hoyos, D.; et al. Neoantigen quality predicts immunoediting in survivors of pancreatic cancer. Nature 2022, 606, 389–395. [Google Scholar] [CrossRef]
- Knudsen, E.S.; Vail, P.; Balaji, U.; Ngo, H.; Botros, I.W.; Makarov, V.; Riaz, N.; Balachandran, V.; Leach, S.; Thompson, D.M.; et al. Stratification of Pancreatic Ductal Adenocarcinoma: Combinatorial Genetic, Stromal, and Immunologic Markers. Clin. Cancer Res. 2017, 23, 4429–4440. [Google Scholar] [CrossRef] [Green Version]
- Romero, J.M.; Grünwald, B.; Jang, G.-H.; Bavi, P.P.; Jhaveri, A.; Masoomian, M.; Fischer, S.E.; Zhang, A.; Denroche, R.E.; Lungu, I.M.; et al. A Four-Chemokine Signature Is Associated with a T-cell–Inflamed Phenotype in Primary and Metastatic Pancreatic Cancer. Clin. Cancer Res. 2020, 26, 1997–2010. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Gillen, S.; Schuster, T.; Büschenfelde, C.M.Z.; Friess, H.; Kleeff, J. Preoperative/Neoadjuvant Therapy in Pancreatic Cancer: A Systematic Review and Meta-analysis of Response and Resection Percentages. PLoS Med. 2010, 7, e1000267. [Google Scholar] [CrossRef]
- Conroy, T.; Hammel, P.; Hebbar, M.; Ben Abdelghani, M.; Wei, A.C.; Raoul, J.-L.; Choné, L.; Francois, E.; Artru, P.; Biagi, J.J.; et al. FOLFIRINOX or Gemcitabine as Adjuvant Therapy for Pancreatic Cancer. N. Engl. J. Med. 2018, 379, 2395–2406. [Google Scholar] [CrossRef]
- Von Hoff, D.D.; Ervin, T.; Arena, F.P.; Chiorean, E.G.; Infante, J.; Moore, M.; Renschler, M.F. Increased survival in pancreatic cancer with nab-paclitaxel plus gemcitabine. N. Engl. J. Med. 2013, 369, 1691–1703. [Google Scholar] [CrossRef] [Green Version]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouché, O.; Guimbaud, R.; Bécouarn, Y.; Adenis, A.; Raoul, J.-L.; Gourgou-Bourgade, S.; De La Fouchardière, C.; et al. FOLFIRINOX versus Gemcitabine for Metastatic Pancreatic Cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Wang-Gillam, A.; Hubner, R.A.; Siveke, J.T.; Von Hoff, D.D.; Belanger, B.; de Jong, F.A.; Mirakhur, B.; Chen, L.-T. NAPOLI-1 phase 3 study of liposomal irinotecan in metastatic pancreatic cancer: Final overall survival analysis and characteristics of long-term survivors. Eur. J. Cancer 2019, 108, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Kamisawa, T.; Wood, L.D.; Itoi, T.; Takaori, K. Pancreatic cancer. Lancet 2016, 388, 73–85. [Google Scholar] [CrossRef]
- Pereira, S.P.; Oldfield, L.; Ney, A.; Hart, P.A.; Keane, M.G.; Pandol, S.J.; Costello, E. Early detection of pancreatic cancer. Lancet Gastroenterol. Hepatol. 2020, 5, 698–710. [Google Scholar] [CrossRef]
- Yamasaki, A.; Yanai, K.; Onishi, H. Hypoxia and pancreatic ductal adenocarcinoma. Cancer Lett. 2020, 484, 9–15. [Google Scholar] [CrossRef]
- Pouysségur, J.; Dayan, F.; Mazure, N.M. Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature 2006, 441, 437–443. [Google Scholar] [CrossRef]
- Shah, V.; Sheppard, B.; Sears, R.; Alani, A.W. Hypoxia: Friend or Foe for drug delivery in Pancreatic Cancer. Cancer Lett. 2020, 492, 63–70. [Google Scholar] [CrossRef]
- Geismann, C.; Arlt, A. Coming in the Air: Hypoxia Meets Epigenetics in Pancreatic Cancer. Cells 2020, 9, 2353. [Google Scholar] [CrossRef]
- McKeown, S.R. Defining normoxia, physoxia and hypoxia in tumours-implications for treatment response. Br. J. Radiol. 2014, 87, 20130676. [Google Scholar] [CrossRef] [Green Version]
- Vaupel, P.; Höckel, M.; Mayer, A. Detection and Characterization of Tumor Hypoxia Using pO2 Histography. Antioxid. Redox Signal 2007, 9, 1221–1236. [Google Scholar] [CrossRef]
- Bachem, M.G.; Schünemann, M.; Ramadani, M.; Siech, M.; Beger, H.; Buck, A.; Zhou, S.; Schmid-Kotsas, A.; Adler, G. Pancreatic carcinoma cells induce fibrosis by stimulating proliferation and matrix synthesis of stellate cells. Gastroenterology 2005, 128, 907–921. [Google Scholar] [CrossRef]
- Ye, L.-Y.; Zhang, Q.; Bai, X.-L.; Pankaj, P.; Hu, Q.-D.; Liang, T.-B. Hypoxia-inducible factor 1α expression and its clinical significance in pancreatic cancer: A meta-analysis. Pancreatology 2014, 14, 391–397. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, J.; Chen, J.; Wang, Y.; Zhou, S.; Huang, L.; Bai, Y.; Peng, C.; Shen, B.; Chen, H.; et al. RER1 enhances carcinogenesis and stemness of pancreatic cancer under hypoxic environment. J. Exp. Clin. Cancer Res. 2019, 38, 15. [Google Scholar] [CrossRef] [Green Version]
- Hermann, P.C.; Huber, S.L.; Herrler, T.; Aicher, A.; Ellwart, J.W.; Guba, M.; Bruns, C.J.; Heeschen, C. Distinct Populations of Cancer Stem Cells Determine Tumor Growth and Metastatic Activity in Human Pancreatic Cancer. Cell Stem Cell 2007, 1, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Li, J.; Sun, L.; Qin, T.; Xiao, Y.; Chen, K.; Qian, W.; Duan, W.; Lei, J.; Ma, J.; et al. Hypoxia-driven paracrine osteopontin/integrin αvβ3 signaling promotes pancreatic cancer cell epithelial–mesenchymal transition and cancer stem cell-like properties by modulating forkhead box protein M1. Mol. Oncol. 2019, 13, 228–245. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, R.K.; Abbruzzese, J.; Dragovich, T.; Kirkpatrick, L.; Guillen, J.M.; Baker, A.F.; Pestano, L.A.; Green, S.; Von Hoff, D.D. A randomized phase II study of PX-12, an inhibitor of thioredoxin in patients with advanced cancer of the pancreas following progression after a gemcitabine-containing combination. Cancer Chemother. Pharmacol. 2010, 67, 503–509. [Google Scholar] [CrossRef]
- Davidson, A.; Veillard, A.-S.; Tognela, A.; Chan, M.; Hughes, B.; Boyer, M.; Briscoe, K.; Begbie, S.; Abdi, E.; Crombie, C.; et al. A phase III randomized trial of adding topical nitroglycerin to first-line chemotherapy for advanced nonsmall-cell lung cancer: The Australasian lung cancer trials group NITRO trial. Ann. Oncol. 2015, 26, 2280–2286. [Google Scholar] [CrossRef]
- Zhao, T.; Ren, H.; Jia, L.; Chen, J.; Xin, W.; Yan, F.; Li, J.; Wang, X.; Gao, S.; Qian, D.; et al. Inhibition of HIF-1α by PX-478 enhances the anti-tumor effect of gemcitabine by inducing immunogenic cell death in pancreatic ductal adenocarcinoma. Oncotarget 2015, 6, 2250–2262. [Google Scholar] [CrossRef]
- Schneider, M.A.; Linecker, M.; Fritsch, R.; Muehlematter, U.J.; Stocker, D.; Pestalozzi, B.; Samaras, P.; Jetter, A.; Kron, P.; Petrowsky, H.; et al. Phase Ib dose-escalation study of the hypoxia-modifier Myo-inositol trispyrophosphate in patients with hepatopancreatobiliary tumors. Nat. Commun. 2021, 12, 3807. [Google Scholar] [CrossRef]
- Martí-Díaz, R.; Montenegro, M.; Cabezas-Herrera, J.; Goding, C.; Rodríguez-López, J.; Sánchez-Del-Campo, L. Acriflavine, a Potent Inhibitor of HIF-1α, Disturbs Glucose Metabolism and Suppresses ATF4-Protective Pathways in Melanoma under Non-Hypoxic Conditions. Cancers 2020, 13, 102. [Google Scholar] [CrossRef]
- Borad, M.J.; Reddy, S.G.; Bahary, N.; Uronis, H.E.; Sigal, D.; Cohn, A.L.; Schelman, W.R.; Stephenson, J., Jr.; Chiorean, E.G.; Rosen, P.J.; et al. Randomized Phase II Trial of Gemcitabine Plus TH-302 Versus Gemcitabine in Patients With Advanced Pancreatic Cancer. J. Clin. Oncol. 2015, 33, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Limani, P.; Linecker, M.; Kron, P.; Samaras, P.; Pestalozzi, B.; Stupp, R.; Jetter, A.; Dutkowski, P.; Müllhaupt, B.; Schlegel, A.; et al. Development of OXY111A, a novel hypoxia-modifier as a potential antitumor agent in patients with hepato-pancreato-biliary neoplasms—Protocol of a first Ib/IIa clinical trial. BMC Cancer 2016, 16, 812. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, K.S.; Kim, G.P.; Foster, N.R.; Wang-Gillam, A.; Erlichman, C.; McWilliams, R.R. Phase II trial of gemcitabine and tanespimycin (17AAG) in metastatic pancreatic cancer: A Mayo Clinic Phase II Consortium study. Investig. New Drugs 2015, 33, 963–968. [Google Scholar] [CrossRef] [Green Version]
- Renouf, D.J.; Hedley, D.; Krzyzanowska, M.K.; Schmuck, M.; Wang, L.; Moore, M.J. A phase II study of the HSP90 inhibitor AUY922 in chemotherapy refractory advanced pancreatic cancer. Cancer Chemother. Pharmacol. 2016, 78, 541–545. [Google Scholar] [CrossRef]
- Grande, E.; Rodriguez-Antona, C.; López, C.; Alonso-Gordoa, T.; Benavent, M.; Capdevila, J.; García-Carbonero, R. Sunitinib and Evofosfamide (TH-302) in Systemic Treatment-Naive Patients with Grade 1/2 Metastatic Pancreatic Neuroendocrine Tumors: The GETNE-1408 Trial. Oncologist 2021, 26, 941–949. [Google Scholar] [CrossRef]
- Hegde, A.; Jayaprakash, P.; Couillault, C.A.; Piha-Paul, S.; Karp, D.; Rodon, J.; Pant, S.; Fu, S.; Dumbrava, E.E.; Yap, T.A.; et al. A Phase I Dose-Escalation Study to Evaluate the Safety and Tolerability of Evofosfamide in Combination with Ipilimumab in Advanced Solid Malignancies. Clin. Cancer Res. 2021, 27, 3050–3060. [Google Scholar] [CrossRef]
- Phillips, R.M. Targeting the hypoxic fraction of tumours using hypoxia-activated prodrugs. Cancer Chemother. Pharmacol. 2016, 77, 441–457. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, L.; Li, X.-F. Targeting Hypoxia: Hypoxia-Activated Prodrugs in Cancer Therapy. Front. Oncol. 2021, 11, 700407. [Google Scholar] [CrossRef]
- Melisi, D.; Oh, D.Y.; Hollebecque, A.; Calvo, E.; Varghese, A.; Borazanci, E.; Garcia-Carbonero, R. Safety and activity of the TGFbeta receptor I kinase inhibitor galunisertib plus the anti-PD-L1 antibody durvalumab in metastatic pancreatic cancer. J. Immunother. Cancer 2021, 9, e002068. [Google Scholar] [CrossRef]
- Jin, S.; Sun, Y.; Liang, X.; Gu, X.; Ning, J.; Xu, Y.; Chen, S.; Pan, L. Emerging new therapeutic antibody derivatives for cancer treatment. Signal Transduct. Target. Ther. 2022, 7, 39. [Google Scholar] [CrossRef]
- Fylaktakidou, K.C.; Lehn, J.-M.; Greferath, R.; Nicolau, C. Inositol tripyrophosphate: A new membrane permeant allosteric effector of haemoglobin. Bioorg. Med. Chem. Lett. 2005, 15, 1605–1608. [Google Scholar] [CrossRef]
- Duarte, C.D.; Greferath, R.; Nicolau, C.; Lehn, J.-M. myo-Inositol Trispyrophosphate: A Novel Allosteric Effector of Hemoglobin with High Permeation Selectivity across the Red Blood Cell Plasma Membrane. Chembiochem 2010, 11, 2543–2548. [Google Scholar] [CrossRef]
- Limani, P.; Linecker, M.; Kachaylo, E.; Tschuor, C.; Kron, P.; Schlegel, A.; Ungethuem, U.; Jang, J.H.; Georgiopoulou, S.; Nicolau, C.; et al. Antihypoxic Potentiation of Standard Therapy for Experimental Colorectal Liver Metastasis through Myo-Inositol Trispyrophosphate. Clin. Cancer Res. 2016, 22, 5887–5897. [Google Scholar] [CrossRef] [Green Version]
- Kieda, C.; El Hafny-Rahbi, B.; Collet, G.; Lamerant-Fayel, N.; Grillon, C.; Guichard, A.; Dulak, J.; Jozkowicz, A.; Kotlinowski, J.; Fylaktakidou, K.C.; et al. Stable tumor vessel normalization with pO2 increase and endothelial PTEN activation by inositol trispyrophosphate brings novel tumor treatment. J. Mol. Med. 2013, 91, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Raykov, Z.; Grekova, S.P.; Bour, G.; Lehn, J.M.; Giese, N.; Nicolau, C.; Aprahamian, M. Myo-inositol trispyrophosphate-mediated hypoxia reversion controls pancreatic cancer in rodents and enhances gemcitabine efficacy. Int. J. Cancer 2013, 134, 2572–2582. [Google Scholar] [CrossRef]
- Aprahamian, M.; Bour, G.; Akladios, C.Y.; Fylaktakidou, K.; Greferath, R.; Soler, L.; Nicolau, C. Myo-InositolTrisPyroPhosphate treatment leads to HIF-1α suppression and eradication of early hepatoma tumors in rats. Chembiochem. 2011, 12, 777–783. [Google Scholar] [CrossRef]
- Ignat, M.; Akladios, C.Y.; Lindner, V.; Khetchoumian, K.; Teletin, M.; Muttter, D.; Aprahamian, P.M.; Marescaux, J. Development of a methodology for in vivo follow-up of hepatocellular carcinoma in hepatocyte specific Trim24-null mice treated with myo-inositol trispyrophosphate. J. Exp. Clin. Cancer Res. 2016, 35, 155. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.-B.; Cao-Pham, T.-T.; Jordan, B.F.; Deschoemaeker, S.; Heyerick, A.; Gallez, B. Impact of myo -inositol trispyrophosphate (ITPP) on tumour oxygenation and response to irradiation in rodent tumour models. J. Cell. Mol. Med. 2019, 23, 1908–1916. [Google Scholar] [CrossRef]
- Derbal-Wolfrom, L.; Pencreach, E.; Saandi, T.; Aprahamian, M.; Martin, E.; Greferath, R.; Tufa, E.; Choquet, P.; Lehn, J.-M.; Nicolau, C.; et al. Increasing the oxygen load by treatment with myo-inositol trispyrophosphate reduces growth of colon cancer and modulates the intestine homeobox gene Cdx2. Oncogene 2012, 32, 4313–4318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grgic, I.; Tschanz, F.; Borgeaud, N.; Gupta, A.; Clavien, P.-A.; Guckenberger, M.; Graf, R.; Pruschy, M. Tumor Oxygenation by Myo-Inositol Trispyrophosphate Enhances Radiation Response. Int. J. Radiat. Oncol. 2021, 110, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- Chida, K.; Kawazoe, A.; Suzuki, T.; Kawazu, M.; Ueno, T.; Takenouchi, K.; Yoshino, T. Transcriptomic Profiling of MSI-H/dMMR Gastrointestinal Tumors to Identify Determinants of Responsiveness to Anti-PD-1 Therapy. Clin. Cancer Res. 2022, 28, 2110–2117. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, P.; Ai, M.; Liu, A.; Budhani, P.; Bartkowiak, T.; Sheng, J.; Ager, C.; Nicholas, C.; Jaiswal, A.R.; Sun, Y.; et al. Targeted hypoxia reduction restores T cell infiltration and sensitizes prostate cancer to immunotherapy. J. Clin. Investig. 2018, 128, 5137–5149. [Google Scholar] [CrossRef] [PubMed]
- Chafe, S.C.; McDonald, P.C.; Saberi, S.; Nemirovsky, O.; Venkateswaran, G.; Burugu, S.; Gao, D.; Delaidelli, A.; Kyle, A.H.; Baker, J.H.E.; et al. Targeting Hypoxia-Induced Carbonic Anhydrase IX Enhances Immune-Checkpoint Blockade Locally and Systemically. Cancer Immunol. Res. 2019, 7, 1064–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatfield, S.M.; Kjaergaard, J.; Lukashev, D.; Schreiber, T.H.; Belikoff, B.; Abbott, R.; Sethumadhavan, S.; Philbrook, P.; Ko, K.; Cannici, R.; et al. Immunological mechanisms of the antitumor effects of supplemental oxygenation. Sci. Transl. Med. 2015, 7, 277ra30. [Google Scholar] [CrossRef] [Green Version]
- Scharping, N.E.; Menk, A.V.; Whetstone, R.D.; Zeng, X.; Delgoffe, G.M. Efficacy of PD-1 Blockade Is Potentiated by Metformin-Induced Reduction of Tumor Hypoxia. Cancer Immunol. Res. 2017, 5, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, X.; Chen, L.; Wang, J.; Zhang, J.; Tang, J.; Ji, Y.; Song, J.; Wang, L.; Zhao, Y.; et al. Oxygen microcapsules improve immune checkpoint blockade by ameliorating hypoxia condition in pancreatic ductal adenocarcinoma. Bioact. Mater. 2023, 20, 259–270. [Google Scholar] [CrossRef]
- Lequeux, A.; Noman, M.Z.; Xiao, M.; Van Moer, K.; Hasmim, M.; Benoit, A.; Bosseler, M.; Viry, E.; Arakelian, T.; Berchem, G.; et al. Targeting HIF-1 alpha transcriptional activity drives cytotoxic immune effector cells into melanoma and improves combination immunotherapy. Oncogene 2021, 40, 4725–4735. [Google Scholar] [CrossRef]
- Liu, X.; Ye, N.; Liu, S.; Guan, J.; Deng, Q.; Zhang, Z.; Xiao, C.; Ding, Z.; Zhang, B.; Chen, X.; et al. Hyperbaric Oxygen Boosts PD-1 Antibody Delivery and T Cell Infiltration for Augmented Immune Responses Against Solid Tumors. Adv. Sci. 2021, 8, e2100233. [Google Scholar] [CrossRef]
- Bailey, C.M.; Liu, Y.; Liu, M.; Du, X.; Devenport, M.; Zheng, P.; Wang, Y. Targeting HIF-1α abrogates PD-L1–mediated immune evasion in tumor microenvironment but promotes tolerance in normal tissues. J. Clin. Investig. 2022, 132, e150846. [Google Scholar] [CrossRef] [PubMed]
- Salman, S.; Meyers, D.J.; Wicks, E.E.; Lee, S.N.; Datan, E.; Thomas, A.M.; Anders, N.M.; Hwang, Y.; Lyu, Y.; Yang, Y.; et al. HIF inhibitor 32-134D eradicates murine hepatocellular carcinoma in combination with anti-PD1 therapy. J. Clin. Investig. 2022, 132, e156774. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.L.; Bankson, J.A.; Lemos, R., Jr.; Lai, S.Y.; Thittai, A.K.; He, Y.; Hostetter, G.; Demeure, M.J.; Von Hoff, D.D.; Powis, G. Radiosensitization and Stromal Imaging Response Correlates for the HIF-1 Inhibitor PX-478 Given with or without Chemotherapy in Pancreatic Cancer. Mol. Cancer Ther. 2010, 9, 2057–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohse, I.; Rasowski, J.; Cao, P.; Pintilie, M.; Do, T.; Tsao, M.-S.; Hill, R.P.; Hedley, D.W. Targeting hypoxic microenvironment of pancreatic xenografts with the hypoxia-activated prodrug TH-302. Oncotarget 2016, 7, 33571–33580. [Google Scholar] [CrossRef] [Green Version]
- Hajj, C.; Russell, J.; Hart, C.P.; Goodman, K.A.; Lowery, M.; Haimovitz-Friedman, A.; Deasy, J.; Humm, J.L. A Combination of Radiation and the Hypoxia-Activated Prodrug Evofosfamide (TH-302) is Efficacious against a Human Orthotopic Pancreatic Tumor Model. Transl. Oncol. 2017, 10, 760–765. [Google Scholar] [CrossRef]
- McDonald, P.C.; Chafe, S.C.; Brown, W.S.; Saberi, S.; Swayampakula, M.; Venkateswaran, G.; Nemirovsky, O.; Gillespie, J.A.; Karasinska, J.M.; Kalloger, S.E.; et al. Regulation of pH by Carbonic Anhydrase 9 Mediates Survival of Pancreatic Cancer Cells With Activated KRAS in Response to Hypoxia. Gastroenterology 2019, 157, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.A.G.; Hu, M.; Ren, H.; Wang, J.; Cheng, X.; Li, R.; Yuan, B.; Balan, Y.; Bai, Z.; Huang, H. VHH212 nanobody targeting the hypoxia-inducible factor 1α suppresses angiogenesis and potentiates gemcitabine therapy in pancreatic cancer in vivo. Cancer Biol. Med. 2021, 18, 772–787. [Google Scholar] [CrossRef]
- Shah, V.M.; Dorrell, C.; Al-Fatease, A.; Allen-Petersen, B.L.; Woo, Y.; Bortnyak, Y.; Gheewala, R.; Sheppard, B.C.; Sears, R.C.; Alani, A.W. Microfluidics Formulated Liposomes of Hypoxia Activated Prodrug for Treatment of Pancreatic Cancer. Pharmaceutics 2022, 14, 713. [Google Scholar] [CrossRef]
- Rauta, P.R.; Mackeyev, Y.; Sanders, K.; Kim, J.B.; Gonzalez, V.V.; Zahra, Y.; Shohayeb, M.A.; Abousaida, B.; Vijay, G.V.; Tezcan, O.; et al. Pancreatic tumor microenvironmental acidosis and hypoxia transform gold nanorods into cell-penetrant particles for potent radiosensitization. Sci. Adv. 2022, 8, eabm9729. [Google Scholar] [CrossRef]
- Miller, B.W.; Morton, J.P.; Pinese, M.; Saturno, G.; Jamieson, N.B.; McGhee, E.; Timpson, P.; Leach, J.; McGarry, L.; Shanks, E.; et al. Targeting the LOX / hypoxia axis reverses many of the features that make pancreatic cancer deadly: Inhibition of LOX abrogates metastasis and enhances drug efficacy. EMBO Mol. Med. 2015, 7, 1063–1076. [Google Scholar] [CrossRef]
- Koong, A.C.; Mehta, V.K.; Le, Q.T.; Fisher, G.A.; Terris, D.J.; Brown, J.M.; Bastidas, A.J.; Vierra, M. Pancreatic tumors show high levels of hypoxia. Int. J. Radiat. Oncol. Biol. Phys. 2000, 48, 919–922. [Google Scholar] [CrossRef]
- Vaupel, P.; Flood, A.B.; Swartz, H.M. Oxygenation Status of Malignant Tumors vs. Normal Tissues: Critical Evaluation and Updated Data Source Based on Direct Measurements with pO2 Microsensors. Appl. Magn. Reson. 2021, 52, 1451–1479. [Google Scholar] [CrossRef]
- Gallez, B. The Role of Imaging Biomarkers to Guide Pharmacological Interventions Targeting Tumor Hypoxia. Front. Pharmacol. 2022, 13, 853568. [Google Scholar] [CrossRef]
- Griffiths, J.R.; Robinson, S.P. The OxyLite: A fibre-optic oxygen sensor. Br. J. Radiol. 1999, 72, 627–630. [Google Scholar] [CrossRef]
- Flood, A.B.; Schaner, P.E.; Vaupel, P.; Williams, B.B.; Gallez, B.; Chen, E.Y.; Ali, A.; Liu, T.; Lawson, V.H.; Schreiber, W.; et al. Clinical and Statistical Considerations when Assessing Oxygen Levels in Tumors: Illustrative Results from Clinical EPR Oximetry Studies. Adv. Exp. Med. Biol. 2020, 1232, 155–168. [Google Scholar] [CrossRef]
- Zoa, A.; Yang, Y.; Wang, W.; Yang, J.; Wang, J.; Wang, H.; Dong, M.; Tian, Y. High expression of hypoxia-inducible factor 1-alpha predicts poor prognosis in pancreatic ductal adenocarcinoma: A meta-analysis and database validation protocol. Transl. Cancer Res. 2021, 11, 3080–3091. [Google Scholar] [CrossRef]
- Dhani, N.C.; Serra, S.; Pintilie, M.; Schwock, J.; Xu, J.; Gallinger, S.; Hill, R.P.; Hedley, D. Analysis of the intra- and intertumoral heterogeneity of hypoxia in pancreatic cancer patients receiving the nitroimidazole tracer pimonidazole. Br. J. Cancer 2015, 113, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, I.; Bilkey, J.; McKee, T.D.; Serra, S.; Pintilie, M.; Do, T.; Xu, J.; Tsao, M.-S.; Gallinger, S.; Hill, R.P.; et al. Digital quantitative tissue image analysis of hypoxia in resected pancreatic ductal adenocarcinomas. Front. Oncol. 2022, 12, 926497. [Google Scholar] [CrossRef]
- O’Connor, J.P.B.; Robinson, S.P.; Waterton, J.C. Imaging tumour hypoxia with oxygen-enhanced MRI and BOLD MRI. Br. J. Radiol. 2019, 92, 20180642. [Google Scholar] [CrossRef]
- Akisik, M.F.; Sandrasegaran, K.; Bu, G.; Lin, C.; Hutchins, G.D.; Chiorean, E.G. Pancreatic Cancer: Utility of Dynamic Contrast-enhanced MR Imaging in Assessment of Antiangiogenic Therapy. Radiology 2010, 256, 441–449. [Google Scholar] [CrossRef]
- Klaassen, R.; Gurney-Champion, O.J.; Wilmink, J.W.; Besselink, M.G.; Engelbrecht, M.R.; Stoker, J.; Nederveen, A.J.; van Laarhoven, H.W. Repeatability and correlations of dynamic contrast enhanced and T2* MRI in patients with advanced pancreatic ductal adenocarcinoma. Magn. Reson. Imaging 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, R.; Steins, A.; Gurney-Champion, O.J.; Bijlsma, M.F.; Van Tienhoven, G.; Engelbrecht, M.R.W.; Van Eijck, C.H.J.; Suker, M.; Wilmink, J.W.; Besselink, M.G.; et al. Pathological validation and prognostic potential of quantitative MRI in the characterization of pancreas cancer: Preliminary experience. Mol. Oncol. 2020, 14, 2176–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, B.F.; Gallez, B. Surrogate MR markers of response to chemo- or radiotherapy in association with co-treatments: A retrospective analysis of multi-modal studies. Contrast Media Mol. Imaging 2010, 5, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Segard, T.; Robins, P.D.; Yusoff, I.F.; Ee, H.; Morandeau, L.; Campbell, E.M.; Francis, R.J. Detection of Hypoxia With 18F-Fluoromisonidazole (18F-FMISO) PET/CT in Suspected or Proven Pancreatic Cancer. Clin. Nucl. Med. 2013, 38, 1–6. [Google Scholar] [CrossRef]
- Yamane, T.; Aikawa, M.; Yasuda, M.; Fukushima, K.; Seto, A.; Okamoto, K.; Koyama, I.; Kuji, I. [18F]FMISO PET/CT as a preoperative prognostic factor in patients with pancreatic cancer. EJNMMI Res. 2019, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Klaassen, R.; Bennink, R.J.; van Tienhoven, G.; Bijlsma, M.F.; Besselink, M.G.; Henegouwen, M.I.V.B.; Wilmink, J.W.; Nederveen, A.J.; Windhorst, A.D.; Hulshof, M.C.; et al. Feasibility and repeatability of PET with the hypoxia tracer [18F]HX4 in oesophageal and pancreatic cancer. Radiother. Oncol. 2015, 116, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Metran-Nascente, C.; Yeung, I.; Vines, D.C.; Metser, U.; Dhani, N.C.; Green, D.; Milosevic, M.; Jaffray, D.; Hedley, D.W. Measurement of Tumor Hypoxia in Patients with Advanced Pancreatic Cancer Based on 18F-Fluoroazomyin Arabinoside Uptake. J. Nucl. Med. 2016, 57, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Elamir, A.M.; Stanescu, T.; Shessel, A.; Tadic, T.; Yeung, I.; Letourneau, D.; Kim, J.; Lukovic, J.; Dawson, L.A.; Wong, R.; et al. Simulated dose painting of hypoxic sub-volumes in pancreatic cancer stereotactic body radiotherapy. Phys. Med. Biol. 2021, 66, 185008. [Google Scholar] [CrossRef]
- Abou Khouzam, R.; Goutham, H.V.; Zaarour, R.F.; Chamseddine, A.N.; Francis, A.; Buart, S.; Chouaib, S. Integrating tumor hypoxic stress in novel and more adaptable strategies for cancer immunotherapy. Semin. Cancer Biol. 2020, 65, 140–154. [Google Scholar] [CrossRef]
- Abou Khouzam, R.; Zaarour, R.F.; Brodaczewska, K.; Azakir, B.; Venkatesh, G.H.; Thiery, J.; Chouaib, S. The Effect of Hypoxia and Hypoxia-Associated Pathways in the Regulation of Antitumor Response: Friends or Foes? Front. Immunol. 2022, 13, 828875. [Google Scholar] [CrossRef]
- Li, H.; Peng, C.; Zhu, C.; Nie, S.; Qian, X.; Shi, Z.; Shen, S. Hypoxia promotes the metastasis of pancreatic cancer through regulating NOX4/KDM5A-mediated histone methylation modification changes in a HIF1A-independent manner. Clin. Epigenet. 2021, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; He, X.; Cheng, X.; Cao, G.; Chen, B.; Chen, S.; Xiong, M. A 4-gene-based hypoxia signature is associated with tumor immune microenvironment and predicts the prognosis of pancreatic cancer patients. World J. Surg. Oncol. 2021, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zheng, J.; Mou, W.; Lu, G.; Chen, S.; Du, J.; Zheng, Y.; Chen, S.; Shen, B.; Li, J.; et al. Development and validation of a hypoxia-stemness-based prognostic signature in pancreatic adenocarcinoma. Front. Pharmacol. 2022, 13, 939542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-J.; Shao, C.; Yin, Y.-X.; Sun, Q.; Li, Y.-N.; Zha, Y.-W.; Li, M.-Y.; Hu, B.-L. Hypoxia-Related Signature Is a Prognostic Biomarker of Pancreatic Cancer. Dis. Markers 2022, 2022, 6449997. [Google Scholar] [CrossRef]
- Tan, Z.; Lei, Y.; Xu, J.; Shi, S.; Hua, J.; Zhang, B.; Meng, Q.; Liu, J.; Zhang, Y.; Wei, M.; et al. The value of a metabolic reprogramming-related gene signature for pancreatic adenocarcinoma prognosis prediction. Aging 2020, 12, 24228–24241. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, Q.; Peng, W.-J.; Hu, J.-F.; Wang, Z.-Y.; Liu, H.; Huang, L.-N. Testin is a tumor suppressor in non-small cell lung cancer. Oncol. Rep. 2017, 37, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Chen, Y.B.; Zhao, J.J.; Zhang, X.F.; Zhu, G.C.; Weng, D.S.; Xia, J.C. TES functions as a Mena-dependent tumor suppressor in gastric cancer carcinogenesis and metastasis. Cancer Commun. 2019, 39, 3. [Google Scholar] [CrossRef]


| Target | Compound | Reference |
|---|---|---|
| Hypoxia | Inositol-trispyrophosphate (ITPP) | NCT02528526 [81,84] |
| Hypoxia-inducible factors (HIF) | XL888 | NCT03095781 |
| Tanespimycin | NCT00577889 [85] | |
| AUY922 | NCT01484860 [86] | |
| Acriflavine | [82] | |
| Hypoxia-activated prodrugs (HAP) | Evofosfamide | NCT02402062 [87], NCT00743379, NCT02047500 [83], NCT01381822, NCT03098160 [88] |
| Mitomycin | [89] | |
| Tirapazamine | [90] | |
| Apaziquone | [90] | |
| Metabolism | BPM31510 | NCT02650804 |
| NIR178 | NCT03207867 | |
| CPI444 | NCT03454451 | |
| Zoledronic acid | NCT00892242 | |
| Epacadostat | NCT03006302 | |
| Immunity | BMS813160 | NCT03184870, NCT03767582 |
| BL8040 | NCT02907099, NCT02826486 | |
| Olaptesed | NCT03168139 | |
| Plerixafor | NCT03168139 | |
| Chemokines/cytokines | Galunisertib | NCT02734160 [91] |
| Vactosertib | NCT02154646 | |
| AP21009 | NCT00844064 | |
| M7824 fusion protein | [92] | |
| NIS793 mAB | [28] | |
| Tocilizumab | NCT02767557 | |
| Siltuximab | NCT00841191 |
| PET Radiotracer | Patients with Pancreatic Cancer | Hypoxia-Related Parameter(s) | Other Parameters | Hypoxic Fraction | Other Associations | Ref. |
|---|---|---|---|---|---|---|
| 18F-FMISO | Seven patients with PDAC | SUVmax and TBR (considering background uptake in skeletal muscle) | 18F-FDG PET/CT imaging to demarcate the tumor zone. CT or MRI in non-distinguishable tumors to identify a suitable ROI | 28% increased uptake values | - No association between 18F-FMISO SUVmax or TBR with tumor size, histological type or metabolic activity. | [134] |
| 25 patients with PDAC | Peak tumor–blood ratio: SUVpeak of 18F-FMISO in the tumor divided by SUVpeak of the aorta | Tumor ROIs were manually registered by two nuclear medicine physicians IHC of HIF-1α in 22 patients | 36% visually positive for 18F-FMISO | - Patients with high peak tumor–blood ratio experienced worse RFS and OS. - Ratio not associated with HIF-1α. | [135] | |
| 18F-HX4 | 13 patients with PDAC | TBRmax: SUVmax in the VOI divided by average SUV in the aorta. VOI and HV: all voxels in the tumor VOI with a TBR > 1 | - | High repeatability of the amount and location of elevated 18F-HX4 uptake | - TBRmax values were more stable compared to the SUVmax and they varied by 16%. | [136] |
| 18F-FAZA | 20 patients with locally advanced or metastatic PDAC (four cases with both primary tumor and liver metastasis were evaluated) | HF: percentage of voxels with SUVs more than three standard deviations from the mean SUV of skeletal muscle, as obtained from two-hour static scans | Tumor perfusion: based on tracer kinetics by acquiring dynamic scans minutes after the injection of 18F-FAZA and applying a two-compartment model, including blood and extravascular space, under the assumption that pancreatic tumors have low perfusion (flow limited). | Heterogeneity in the HF ranged from values less than 5% to those greater than 50% | - No correlation with tumor volume or perfusion. - Similar 18F-FAZA SUV reported in primary and metastatic tumors. - A trend of higher HF in primary tumors in patients with metastasis than those who are metastasis-free. | [137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Khouzam, R.; Lehn, J.-M.; Mayr, H.; Clavien, P.-A.; Wallace, M.B.; Ducreux, M.; Limani, P.; Chouaib, S. Hypoxia, a Targetable Culprit to Counter Pancreatic Cancer Resistance to Therapy. Cancers 2023, 15, 1235. https://doi.org/10.3390/cancers15041235
Abou Khouzam R, Lehn J-M, Mayr H, Clavien P-A, Wallace MB, Ducreux M, Limani P, Chouaib S. Hypoxia, a Targetable Culprit to Counter Pancreatic Cancer Resistance to Therapy. Cancers. 2023; 15(4):1235. https://doi.org/10.3390/cancers15041235
Chicago/Turabian StyleAbou Khouzam, Raefa, Jean-Marie Lehn, Hemma Mayr, Pierre-Alain Clavien, Michael Bradley Wallace, Michel Ducreux, Perparim Limani, and Salem Chouaib. 2023. "Hypoxia, a Targetable Culprit to Counter Pancreatic Cancer Resistance to Therapy" Cancers 15, no. 4: 1235. https://doi.org/10.3390/cancers15041235