

Increased Expression Levels of Netrin-1 in Visceral Adipose Tissue during Obesity Favour Colon Cancer Cell Migration
 , , , , , , , , and
, , , , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Surgical Procedures, Sample Collection, and Analytical Measurements
2.3. Experimental Animals
2.4. Human Adipocyte and Colon Adenocarcinoma Cell Cultures
2.5. Analysis of Gene Expression Levels
2.6. Migration Assay
2.7. Data and Statistical Analysis
3. Results
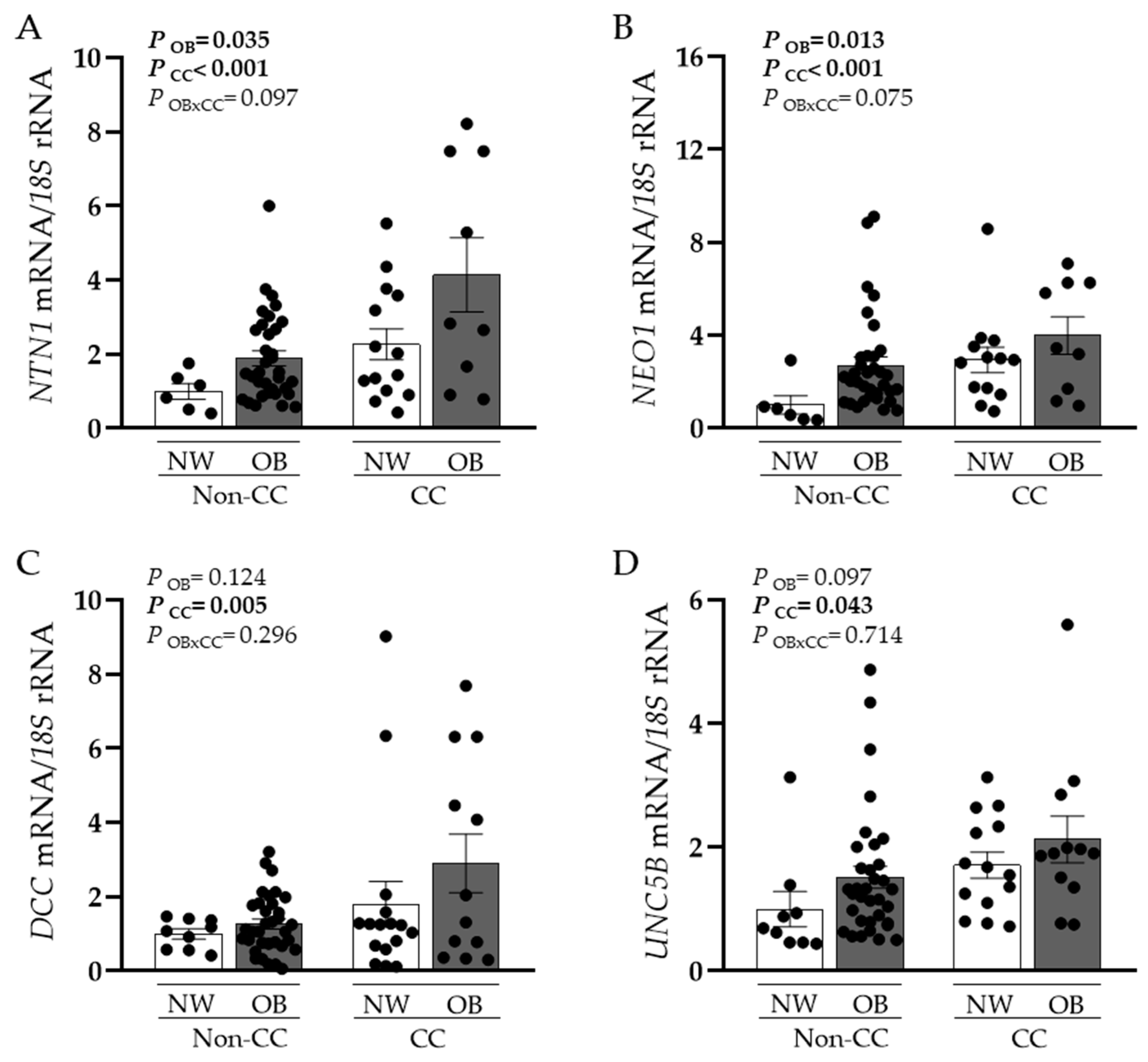
3.1. NTN1 and NEO1 Expression Levels Are Increased in VAT in Obesity and CC and Are Associated with Key Inflammatory Factors
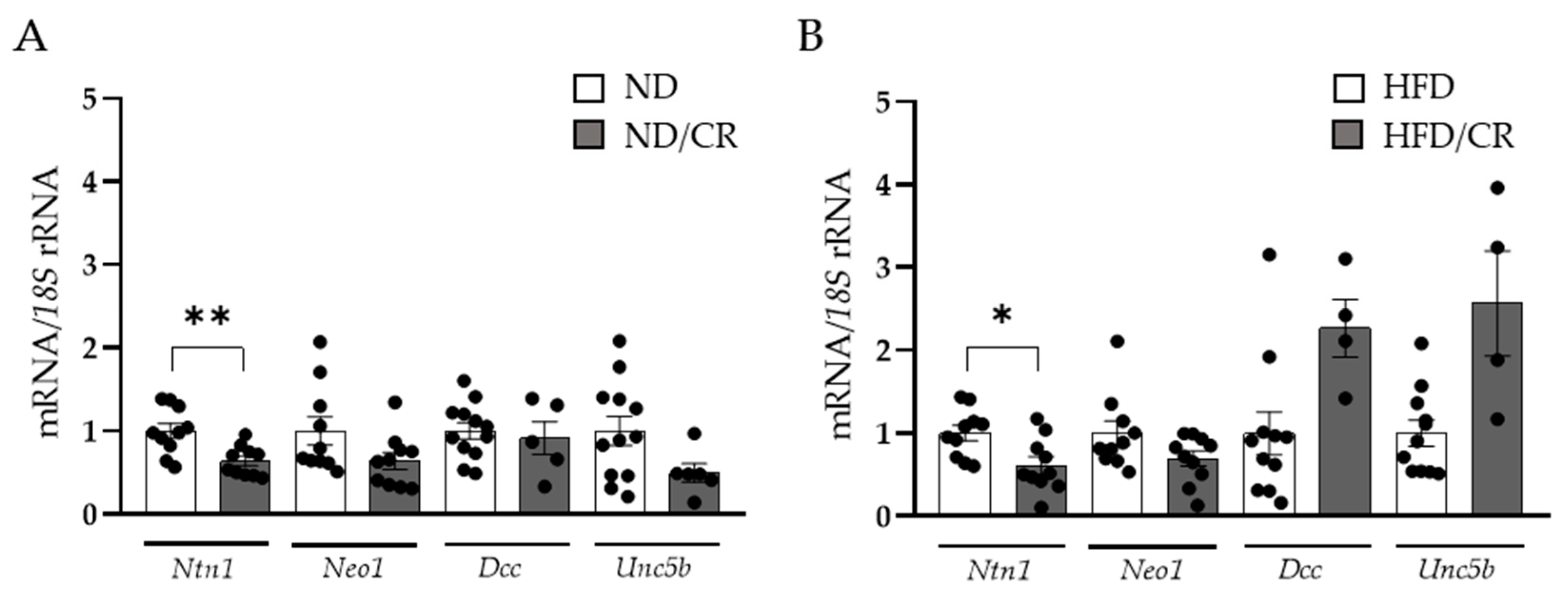
3.2. Decreased Ntn1 Expression Levels in the Colon of Rats after Caloric Restriction
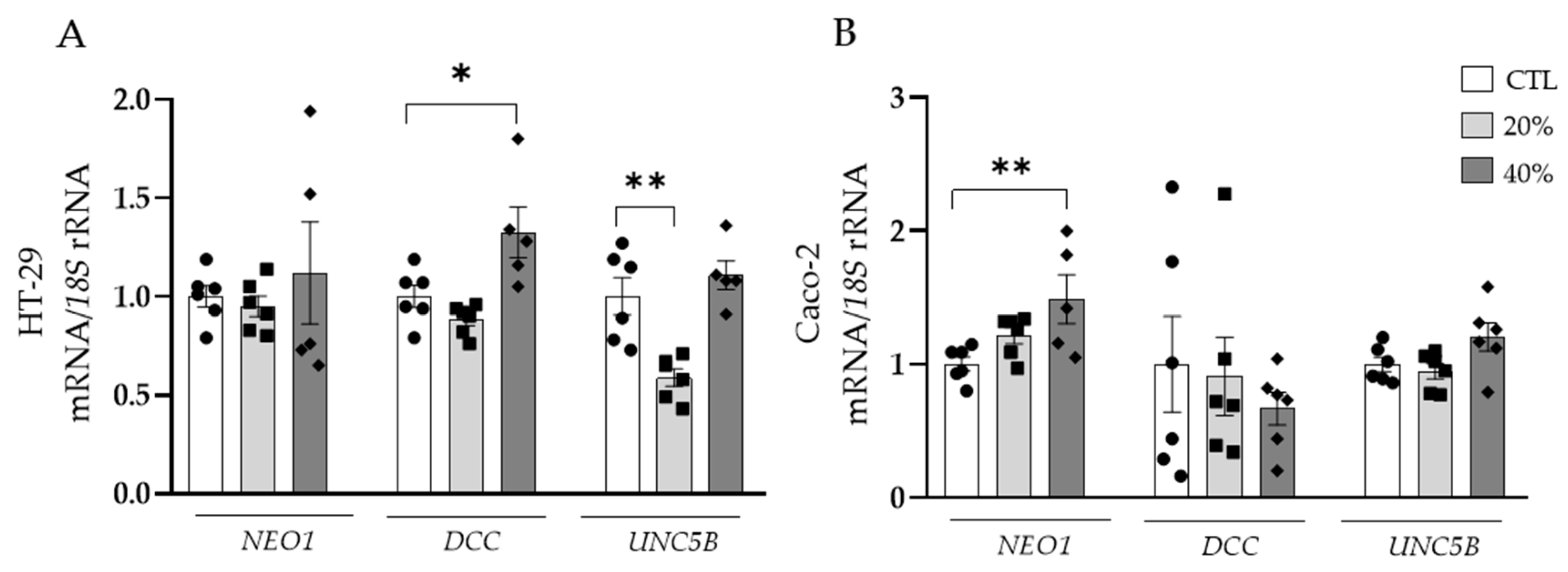
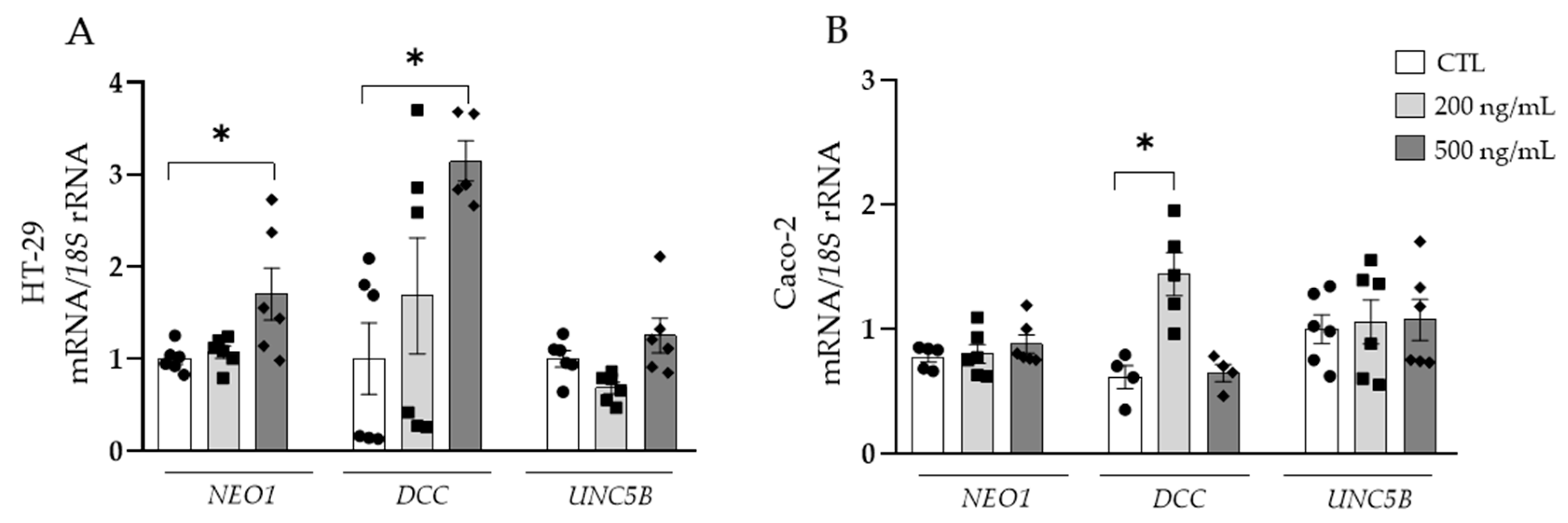
3.3. Adipocyte-Conditioned Media and NTN-1 Induce the Expression of NEO1 and DCC in Human Colorectal Cells
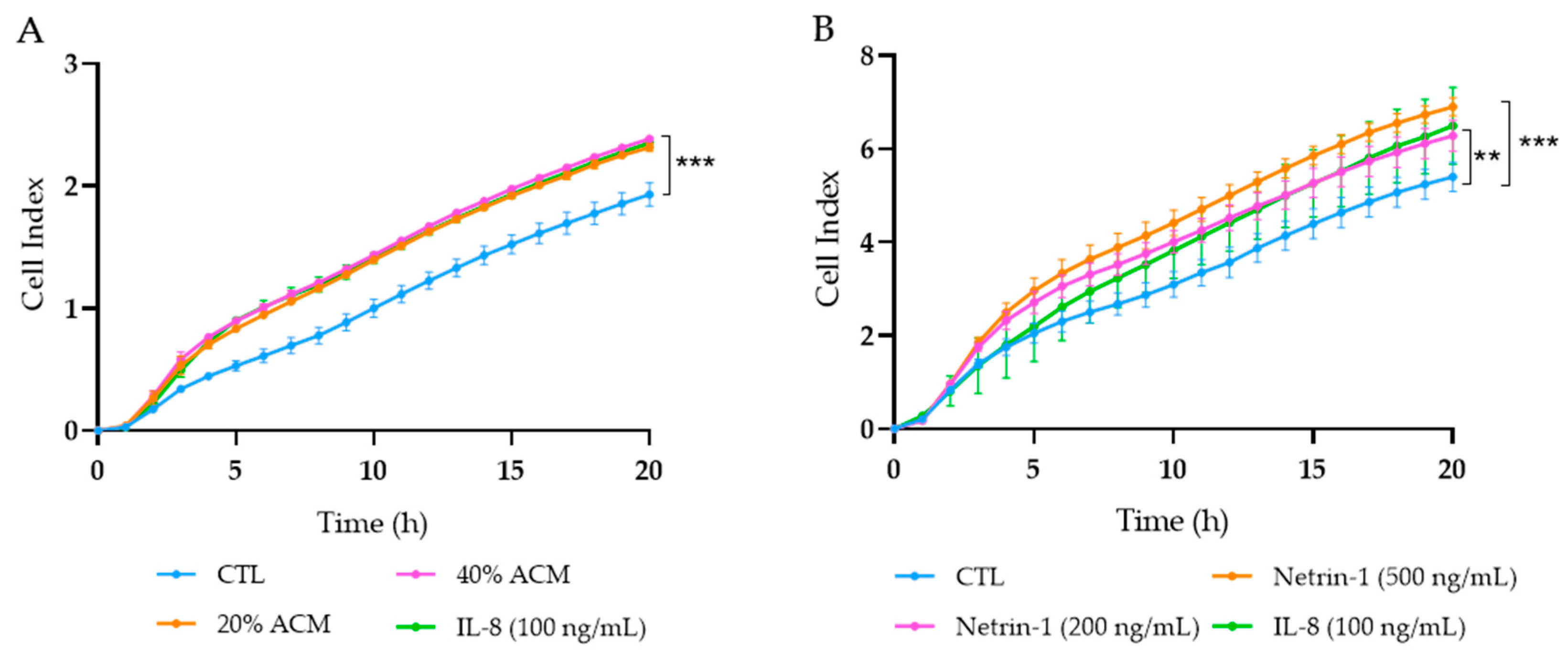
3.4. Effect of Adipocyte-Conditioned Media and NTN-1 on Caco-2 Cell Migration
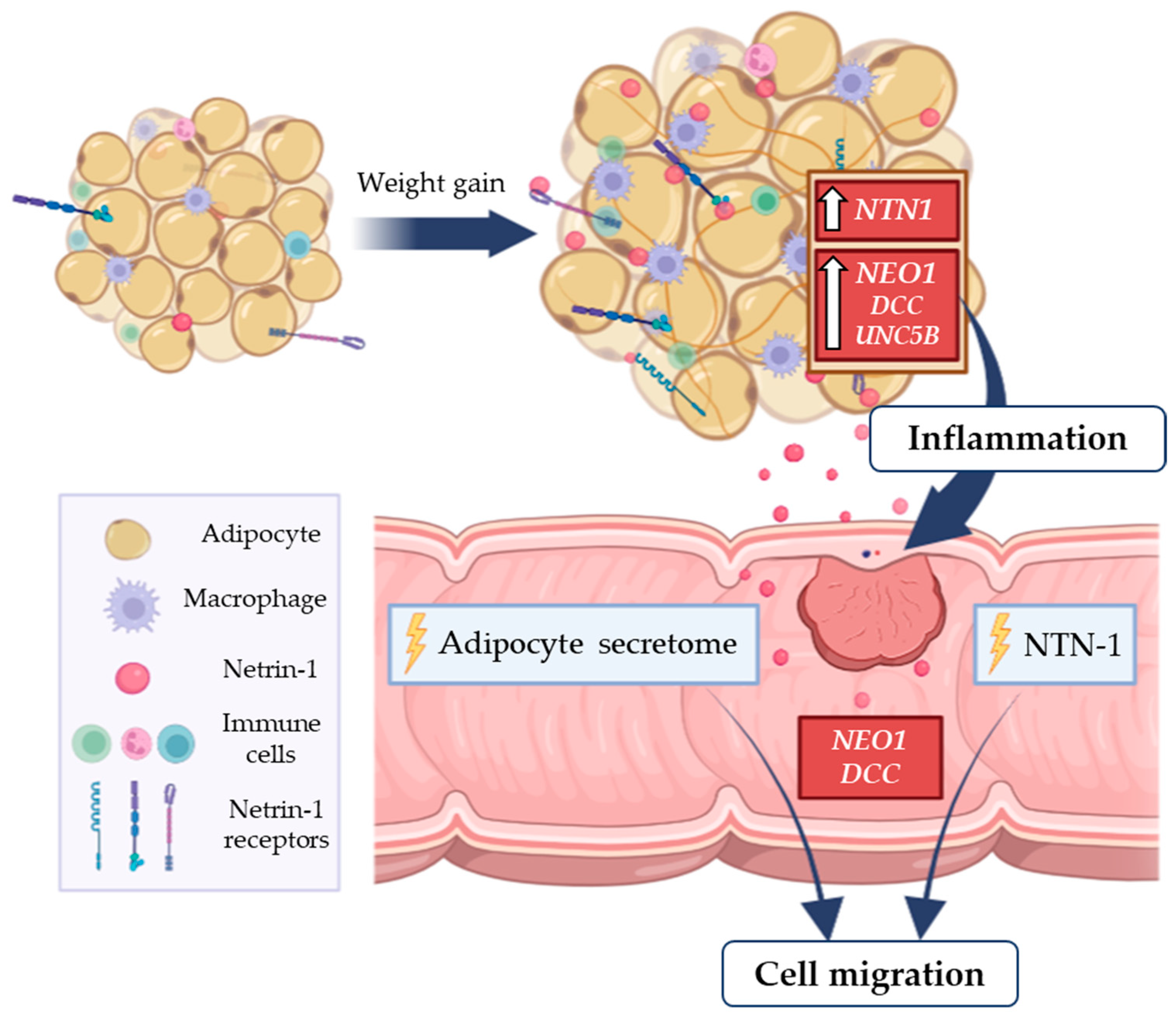
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Goding Sauer, A.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed]
- GBD 2015 Obesity Collaborators; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [PubMed]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body fatness and cancer--viewpoint of the IARC working group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef]
- Frühbeck, G.; Busetto, L.; Dicker, D.; Yumuk, V.; Goossens, G.H.; Hebebrand, J.; Halford, J.G.; Farpour-Lambert, N.J.; Blaak, E.E.; Woodward, E.; et al. The ABCD of Obesity: An EASO Position Statement on a Diagnostic Term with Clinical and Scientific Implications. Obes. Facts 2019, 12, 131–136. [Google Scholar] [CrossRef]
- Quail, D.F.; Dannenberg, A.J. The obese adipose tissue microenvironment in cancer development and progression. Nat. Rev. Endocrinol. 2019, 15, 139–154. [Google Scholar] [CrossRef]
- Pérez-Hernández, A.I.; Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Frühbeck, G. Mechanisms linking excess adiposity and carcinogenesis promotion. Front. Endocrinol. 2014, 5, 65. [Google Scholar]
- Xia, X.; Hu, Z.; Wang, S.; Yin, K. Netrin-1: An emerging player in inflammatory diseases. Cytokine Growth Factor Rev. 2022, 64, 46–56. [Google Scholar] [CrossRef]
- Ziegon, L.; Schlegel, M. Netrin-1: A modulator of macrophage driven acute and chronic inflammation. Int. J. Mol. Sci. 2021, 23, 275. [Google Scholar] [CrossRef]
- Mazelin, L.; Bernet, A.; Bonod-Bidaud, C.; Pays, L.; Arnaud, S.; Gespach, C.; Bredesen, D.E.; Scoazec, J.Y.; Mehlen, P. Netrin-1 controls colorectal tumorigenesis by regulating apoptosis. Nature 2004, 431, 80–84. [Google Scholar] [CrossRef]
- Ylivinkka, I.; Keski-Oja, J.; Hyytiäinen, M. Netrin-1: A regulator of cancer cell motility? Eur J Cell Biol 2016, 95, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, H. Netrin-1 and its receptors in tumorigenesis. Nat. Rev. Cancer 2004, 4, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Schlegel, M.; Brown, E.J.; Sansbury, B.E.; Weinstock, A.; Afonso, M.S.; Corr, E.M.; van Solingen, C.; Shanley, L.C.; Peled, D.; et al. Netrin-1 Alters Adipose Tissue Macrophage Fate and Function in Obesity. Immunometabolism 2019, 1, e190010. [Google Scholar] [CrossRef] [PubMed]
- Mentxaka, A.; Gómez-Ambrosi, J.; Ramírez, B.; Rodríguez, A.; Becerril, S.; Neira, G.; Valentí, V.; Moncada, R.; Silva, C.; Unamuno, X.; et al. Netrin-1 promotes visceral adipose tissue inflammation in obesity and is associated with insulin resistance. Nutrients 2022, 14, 4372. [Google Scholar] [CrossRef] [PubMed]
- Ramkhelawon, B.; Hennessy, E.J.; Ménager, M.; Ray, T.D.; Sheedy, F.J.; Hutchison, S.; Wanschel, A.; Oldebeken, S.; Geoffrion, M.; Spiro, W.; et al. Netrin-1 promotes adipose tissue macrophage retention and insulin resistance in obesity. Nat. Med. 2014, 20, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Fitamant, J.; Guenebeaud, C.; Coissieux, M.M.; Guix, C.; Treilleux, I.; Scoazec, J.Y.; Bachelot, T.; Bernet, A.; Mehlen, P. Netrin-1 expression confers a selective advantage for tumor cell survival in metastatic breast cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 4850–4855. [Google Scholar] [CrossRef]
- Han, P.; Liu, J.; Lei, Y.; Lin, Z.; Tian, D.; Yan, W. Netrin-1 promotes the collective cell migration of liver cancer cells in a 3D cell culture model. J. Physiol. Biochem. 2019, 75, 489–498. [Google Scholar] [CrossRef]
- Paradisi, A.; Maisse, C.; Coissieux, M.M.; Gadot, N.; Lépinasse, F.; Delloye-Bourgeois, C.; Delcros, J.G.; Svrcek, M.; Neufert, C.; Fléjou, J.F.; et al. Netrin-1 up-regulation in inflammatory bowel diseases is required for colorectal Cancer progression. Proc. Natl. Acad. Sci. USA 2009, 106, 17146–17151. [Google Scholar] [CrossRef]
- Li, B.; Shen, K.; Zhang, J.; Jiang, Y.; Yang, T.; Sun, X.; Ma, X.; Zhu, J. Serum netrin-1 as a biomarker for colorectal cancer detection. Cancer Biomark. 2020, 28, 391–396. [Google Scholar] [CrossRef]
- Kefeli, U.; Yildirim, M.E.; Aydin, D.; Madenci, O.C.; Yasar, N.; Sener, N.; Mert, A.G.; Yuksel, S.; Ercelep, O.B.; Korkmaz, T.; et al. Netrin-1 concentrations in patients with advanced gastric cancer and its relation with treatment. Biomarkers 2012, 17, 663–667. [Google Scholar] [CrossRef]
- Paradisi, A.; Maisse, C.; Bernet, A.; Coissieux, M.M.; Maccarrone, M.; Scoazec, J.Y.; Mehlen, P. NF-KappaB regulates netrin-1 expression and affects the conditional tumor suppressive activity of the netrin-1 receptors. Gastroenterology 2008, 135, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Paradisi, A.; Mehlen, P. Netrin-1, a missing link between chronic inflammation and tumor progression. Cell Cycle 2010, 9, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Castets, M.; Broutier, L.; Molin, Y.; Brevet, M.; Chazot, G.; Gadot, N.; Paquet, A.; Mazelin, L.; Jarrosson-Wuilleme, L.; Scoazec, J.-Y.; et al. DCC constrains tumour progression via its dependence receptor activity. Nature 2011, 482, 534–537. [Google Scholar] [CrossRef]
- Frühbeck, G.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Becerril, S.; Salvador, J.; Colina, I.; Gómez-Ambrosi, J. Adiponectin-leptin ratio is a functional biomarker of adipose tissue inflammation. Nutrients 2019, 11, 454. [Google Scholar] [CrossRef]
- Gómez-Ambrosi, J.; Silva, C.; Catalán, V.; Rodríguez, A.; Galofré, J.C.; Escalada, J.; Valentí, V.; Rotellar, F.; Romero, S.; Ramírez, B.; et al. Clinical usefulness of a new equation for estimating body fat. Diabetes Care 2012, 35, 383–388. [Google Scholar] [CrossRef]
- Rodríguez, A.; Gomez-Ambrosi, J.; Catalan, V.; Rotellar, F.; Valentí, V.; Silva, C.; Mugueta, M.D.C.; Pulido, M.R.; Vázquez, R.; Salvador, J.; et al. The ghrelin O-acyltransferase–ghrelin system reduces TNF-α-induced apoptosis and autophagy in human visceral adipocytes. Diabetologia 2012, 55, 3038–3050. [Google Scholar] [CrossRef]
- Muruzábal, F.J.; Frühbeck, G.; Gómez-Ambrosi, J.; Archanco, M.; Burrell, M.A. Immunocytochemical detection of leptin in non-mammalian vertebrate stomach. Gen. Comp. Endocrinol. 2002, 128, 149–152. [Google Scholar] [CrossRef]
- Rodríguez, A.; Catalán, V.; Gómez-Ambrosi, J.; García-Navarro, S.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Salvador, J.; Burrell, M.A.; et al. Insulin- and leptin-mediated control of aquaglyceroporins in human adipocytes and hepatocytes is mediated via the PI3K/Akt/MTOR signaling cascade. J. Clin. Endocrinol. Metab. 2011, 96, E586–E597. [Google Scholar] [CrossRef]
- Frühbeck, G.; Mentxaka, A.; Ahechu, P.; Gómez-Ambrosi, J.; Ramírez, B.; Becerril, S.; Rodríguez, A.; Unamuno, X.; Cienfuegos, J.A.; Casado, M.; et al. The differential expression of the inflammasomes in adipose tissue and colon influences the development of colon cancer in a context of obesity by regulating intestinal inflammation. J. Inflamm. Res. 2021, 14, 6431–6446. [Google Scholar] [CrossRef]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Ortega, V.A.; Hernández-Lizoain, J.L.; Baixauli, J.; Becerril, S.; Rotellar, F.; Valentí, V.; et al. IL-32α-induced inflammation constitutes a link between obesity and colon cancer. Oncoimmunology 2017, 6, e1328338. [Google Scholar] [CrossRef]
- Catalán, V.; Gómez-Ambrosi, J.; Rotellar, F.; Silva, C.; Rodríguez, A.; Salvador, J.; Gil, M.J.; Cienfuegos, J.A.; Frühbeck, G. Validation of endogenous control genes in human adipose tissue: Relevance to obesity and obesity-associated type 2 diabetes mellitus. Horm. Metab. Res. 2007, 39, 495–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, Y.; Manegold, P.C.; Hong, Y.K.; Zhang, W.; Pohl, A.; Lurje, G.; Winder, T.; Yang, D.; LaBonte, M.J.; Wilson, P.M.; et al. Interleukin-8 is associated with proliferation, migration, angiogenesis and chemosensitivity in vitro and in vivo in colon cancer cell line models. Int. J. Cancer 2011, 128, 2038–2049. [Google Scholar] [CrossRef] [PubMed]
- Kuai, W.X.; Wang, Q.; Yang, X.Z.; Zhao, Y.; Yu, R.; Tang, X.J. Interleukin-8 associates with adhesion, migration, invasion and chemosensitivity of human gastric cancer cells. World J. Gastroenterol. 2012, 18, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.; Kirstein, S. Real-time, label-free monitoring of cellular invasion and migration with the XCELLigence system. Nat. Methods 2009, 6, v–vi. [Google Scholar] [CrossRef]
- Ly, N.P.; Komatsuzaki, K.; Fraser, I.P.; Tseng, A.A.; Prodhan, P.; Moore, K.J.; Kinane, T.B. Netrin-1 inhibits leukocyte migration in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 14729–14734. [Google Scholar] [CrossRef]
- Martín, M.; Rodríguez, A.; Gómez-Ambrosi, J.; Ramírez, B.; Becerril, S.; Catalán, V.; López, M.; Diéguez, C.; Frühbeck, G.; Burrel, M.A. Caloric restriction prevents metabolic dysfunction and the changes in hypothalamic neuropeptides associated with obesity independently of dietary fat content in rats. Nutrients 2021, 13, 2128. [Google Scholar] [CrossRef]
- Nieman, K.M.; Romero, I.L.; Van Houten, B.; Lengyel, E. Adipose tissue and adipocytes support tumorigenesis and metastasis. Biochim. Biophys. Acta 2013, 1831, 1533–1541. [Google Scholar] [CrossRef]
- Rajesh, Y.; Sarkar, D. Association of adipose tissue and adipokines with development of obesity-induced liver cancer. Int. J. Mol. Sci. 2021, 22, 2163. [Google Scholar] [CrossRef]
- Zhang, X.; Cui, P.; Ding, B.; Guo, Y.; Han, K.; Li, J.; Chen, H.; Zhang, W. Netrin-1 elicits metastatic potential of non-small cell lung carcinoma cell by enhancing cell invasion, migration and vasculogenic mimicry via EMT induction. Cancer Gene Ther. 2018, 25, 18–26. [Google Scholar] [CrossRef]
- Chen, H.; Chen, Q.; Luo, Q. Expression of netrin-1 by hypoxia contributes to the invasion and migration of prostate carcinoma cells by regulating YAP activity. Exp. Cell Res. 2016, 349, 302–309. [Google Scholar] [CrossRef]
- Kefeli, U.; Ucuncu Kefeli, A.; Cabuk, D.; Isik, U.; Sonkaya, A.; Acikgoz, O.; Ozden, E.; Uygun, K. Netrin-1 in cancer: Potential biomarker and therapeutic target? Tumor Biol. 2017, 39, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frühbeck, G. Obesity: Aquaporin enters the picture. Nature 2005, 438, 436–437. [Google Scholar] [CrossRef] [PubMed]
- Catalan, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Fernández-Real, J.M.; Salvador, J.; et al. Increased Levels of Calprotectin in Obesity Are Related to Macrophage Content: Impact on Inflammation and Effect of Weight Loss. Mol. Med. 2011, 17, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Ringel, A.E.; Drijvers, J.M.; Baker, G.J.; Catozzi, A.; García-Cañaveras, J.C.; Gassaway, B.M.; Miller, B.C.; Juneja, V.R.; Nguyen, T.H.; Joshi, S.; et al. Obesity shapes metabolism in the tumor microenvironment to suppress anti-tumor immunity. Cell 2020, 183, 1848–1866. [Google Scholar] [CrossRef]
- Meynet, O.; Ricci, J.E. Caloric restriction and cancer: Molecular mechanisms and clinical implications. Trends Mol. Med. 2014, 20, 419–427. [Google Scholar] [CrossRef]
- Olivo-Marston, S.E.; Hursting, S.D.; Perkins, S.N.; Schetter, A.; Khan, M.; Croce, C.; Harris, C.C.; Lavigne, J. Effects of calorie restriction and diet-induced obesity on murine colon carcinogenesis, growth and inflammatory factors, and microRNA expression. PLoS ONE 2014, 9, e94765. [Google Scholar] [CrossRef]
- Steinbach, G.; Kumar, S.P.; Reddy, B.S.; Lipkin, M.; Holt, P.R. Effects of caloric restriction and dietary fat on epithelial cell proliferation in rat colon. Cancer Res. 1993, 53, 2745–2749. [Google Scholar]
- Grandin, M.; Meier, M.; Delcros, J.G.; Nikodemus, D.; Reuten, R.; Patel, T.R.; Goldschneider, D.; Orriss, G.; Krahn, N.; Boussouar, A.; et al. Structural decoding of the netrin-1/UNC5 interaction and its therapeutical implications in cancers. Cancer Cell 2016, 29, 173–185. [Google Scholar] [CrossRef]
- Rodrigues, S.; De Wever, O.; Bruyneel, E.; Rooney, R.J.; Gespach, C. Opposing roles of netrin-1 and the dependence receptor DCC in cancer cell invasion, tumor growth and metastasis. Oncogene 2007, 26, 5615–5625. [Google Scholar] [CrossRef]
- Mehlen, P.; Llambi, F. Role of netrin-1 and netrin-1 dependence receptors in colorectal cancers. Br. J. Cancer 2005, 93, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Boyer, N.P.; Gupton, S.L. Revisiting netrin-1: One who guides (axons). Front. Cell. Neurosci. 2018, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Untiveros, G.; Raskind, A.; Linares, L.; Dotti, A.; Strizzi, L. Netrin-1 stimulates migration of neogenin expressing aggressive melanoma cells. Int. J. Mol. Sci. 2022, 23, 12751. [Google Scholar] [CrossRef]
- Shin, S.K.; Nagasaka, T.; Jung, B.H.; Matsubara, N.; Kim, W.H.; Carethers, J.M.; Boland, C.R.; Goel, A. Epigenetic and genetic alterations in netrin-1 receptors UNC5C and DCC in human colon cancer. Gastroenterology 2007, 133, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Li, D.Y.; Luo, H.R.; Guan, K.L.; Ye, K. Netrin-1 exerts oncogenic activities through enhancing yes-associated protein stability. Proc. Natl. Acad. Sci. USA 2015, 112, 7255–7260. [Google Scholar] [CrossRef] [PubMed]
- First in Human Evaluation of Safety, Pharmacokinetics, and Clinical Activity of a Monoclonal Antibody Targeting Netrin-1 in Patients with Advanced/Metastatic Solid Tumors (NP137). Available online: https://clinicaltrials.gov/ct2/show/NCT02977195 (accessed on 12 December 2022).
- Murray, M.J. The role of netrins and their receptors in epithelial mesenchymal plasticity during development. Cells Tissues Organs 2017, 203, 71–81. [Google Scholar] [CrossRef]
- Sturm, A.; Baumgart, D.C.; d’Heureuse, J.H.; Hotz, A.; Wiedenmann, B.; Dignass, A.U. CXCL8 modulates human intestinal epithelial cells through a CXCR1 dependent pathway. Cytokine 2005, 29, 42–48. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-CC | CC | ||||||
|---|---|---|---|---|---|---|---|
| NW | OB | NW | OB | P OB | P CC | P OB × CC | |
| n | 9 | 37 | 16 | 12 | |||
| Age (years) | 44 ± 6.4 | 46 ± 2.2 | 60 ± 2.6 | 64 ± 3.1 | 0.410 | <0.001 | 0.652 |
| Weight (kg) | 59.5 ± 2.8 | 115 ± 3.8 *** | 64.5± 2.2 ††† | 80.3 ± 2.9 **,†††,# | <0.001 | 0.010 | <0.001 |
| BMI (kg/m2) | 20.8 ± 0.5 | 40.9 ± 1.3 *** | 21.8 ± 0.4 ††† | 29.5 ± 0.7 ***,†††,### | <0.001 | 0.001 | <0.001 |
| Estimated BF (%) | 22.4 ± 2.1 | 50.0 ± 1.3 *** | 28.3 ± 1.6 ††† | 35.2 ± 1.7 **,††† | <0.001 | 0.076 | <0.001 |
| Glucose (mg/dL) | 88 ± 4.6 | 119.9 ± 8.4 | 130.2 ± 14.9 | 127.5 ± 8.8 | 0.341 | 0.217 | 0.105 |
| Triglycerides (mg/dL) | 71 ± 11.1 | 131.9 ± 9.7 | 94.2 ± 21.8 | 182 ± 19 | 0.005 | 0.382 | 0.569 |
| Leucocytes (×106) | 6.4 ± 0.5 | 7.3 ± 0.4 | 6.8 ± 1 | 7.7 ± 0.8 | 0.542 | 0.524 | 0.507 |
| S100A8 (ng/mL) | 357.4 ± 93.6 | 552.9 ± 69 | 233 ± 34.8 | 387.9 ± 57.4 | <0.001 | 0.591 | 0.372 |
| Lactoferrin (ng/mL) | 37.9 ± 5.7 | 71.3 ± 7.7 | 50.8 ± 7.2 | 55.8 ± 7.6 | 0.037 | 0.981 | 0.197 |
| CCL5 (ng/mL) | 5.1 ± 0.7 | 9.3 ± 0.7 ** | 6.6 ± 0.7 † | 5.4 ± 0.7 †† | 0.048 | 0.043 | 0.002 |
| mRNA NTN1 | mRNA NEO1 | mRNA DCC | mRNA UNC5B | |||||
|---|---|---|---|---|---|---|---|---|
| r | P | r | P | r | P | r | P | |
| mRNA NTN1 | - | - | 0.69 | <0.001 | 0.32 | 0.011 | 0.37 | 0.004 |
| mRNA NEO1 | 0.69 | <0.001 | - | - | 0.37 | 0.003 | 0.49 | <0.001 |
| mRNA DCC | 0.32 | 0.011 | 0.37 | 0.003 | - | - | 0.47 | <0.001 |
| mRNA UNC5B | 0.37 | 0.004 | 0.49 | <0.001 | 0.47 | 0.000 | - | - |
| mRNA ASC | 0.40 | <0.001 | 0.57 | <0.001 | 0.13 | 0.307 | 0.41 | <0.001 |
| mRNA IL1B | 0.03 | 0.811 | 0.21 | 0.128 | 0.20 | 0.146 | −0.04 | 0.756 |
| mRNA IL18 | 0.43 | <0.001 | 0.54 | <0.001 | 0.16 | 0.225 | 0.43 | <0.001 |
| mRNA NLRP6 | −0.07 | 0.59 | −0.02 | 0.872 | 0.07 | 0.610 | 0.06 | 0.656 |
| Non CC | CC | ||||||
|---|---|---|---|---|---|---|---|
| NW | OB | NW | OB | P OB | P CC | P OB × CC | |
| ASC | 1.00 ± 0.23 | 1.46 ± 0.10 ** | 1.62 ± 0.19 ** | 1.61 ± 0.29 * | 0.003 | 0.001 | 0.010 |
| IL1B | 1.00 ± 0.31 | 5.71 ± 0.26 | 5.34 ± 2.04 | 8.49 ± 2.61 | 0.016 | 0.022 | 0.439 |
| IL18 | 1.00 ± 0.28 | 0.95 ± 0.12 | 1.41 ± 0.23 | 1.14 ± 0.36 | 0.026 | 0.004 | 0.024 |
| NLRP6 | 1.00 ± 0.23 | 6.94 ± 1.36 | 6.00 ± 1.63 | 9.61 ± 2.94 | 0.006 | 0.004 | 0.137 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mentxaka, A.; Gómez-Ambrosi, J.; Neira, G.; Ramírez, B.; Becerril, S.; Rodríguez, A.; Valentí, V.; Moncada, R.; Baixauli, J.; Burrell, M.A.; et al. Increased Expression Levels of Netrin-1 in Visceral Adipose Tissue during Obesity Favour Colon Cancer Cell Migration. Cancers 2023, 15, 1038. https://doi.org/10.3390/cancers15041038
Mentxaka A, Gómez-Ambrosi J, Neira G, Ramírez B, Becerril S, Rodríguez A, Valentí V, Moncada R, Baixauli J, Burrell MA, et al. Increased Expression Levels of Netrin-1 in Visceral Adipose Tissue during Obesity Favour Colon Cancer Cell Migration. Cancers. 2023; 15(4):1038. https://doi.org/10.3390/cancers15041038
Chicago/Turabian StyleMentxaka, Amaia, Javier Gómez-Ambrosi, Gabriela Neira, Beatriz Ramírez, Sara Becerril, Amaia Rodríguez, Víctor Valentí, Rafael Moncada, Jorge Baixauli, María A. Burrell, and et al. 2023. "Increased Expression Levels of Netrin-1 in Visceral Adipose Tissue during Obesity Favour Colon Cancer Cell Migration" Cancers 15, no. 4: 1038. https://doi.org/10.3390/cancers15041038