

Characterization of Estrogen Receptors in Pancreatic Adenocarcinoma with Tertiary Lymphoid Structures
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Data Source
2.2. Bioinformatic Analysis Methods
2.3. Patients and Samples
2.4. Immunohistochemistry (IHC)
2.5. Immune Cell Chemotaxis Assay
2.6. Statistical Methods and Software
3. Results
3.1. Characterization of Regulatory Factors of TLS Development in PAAD
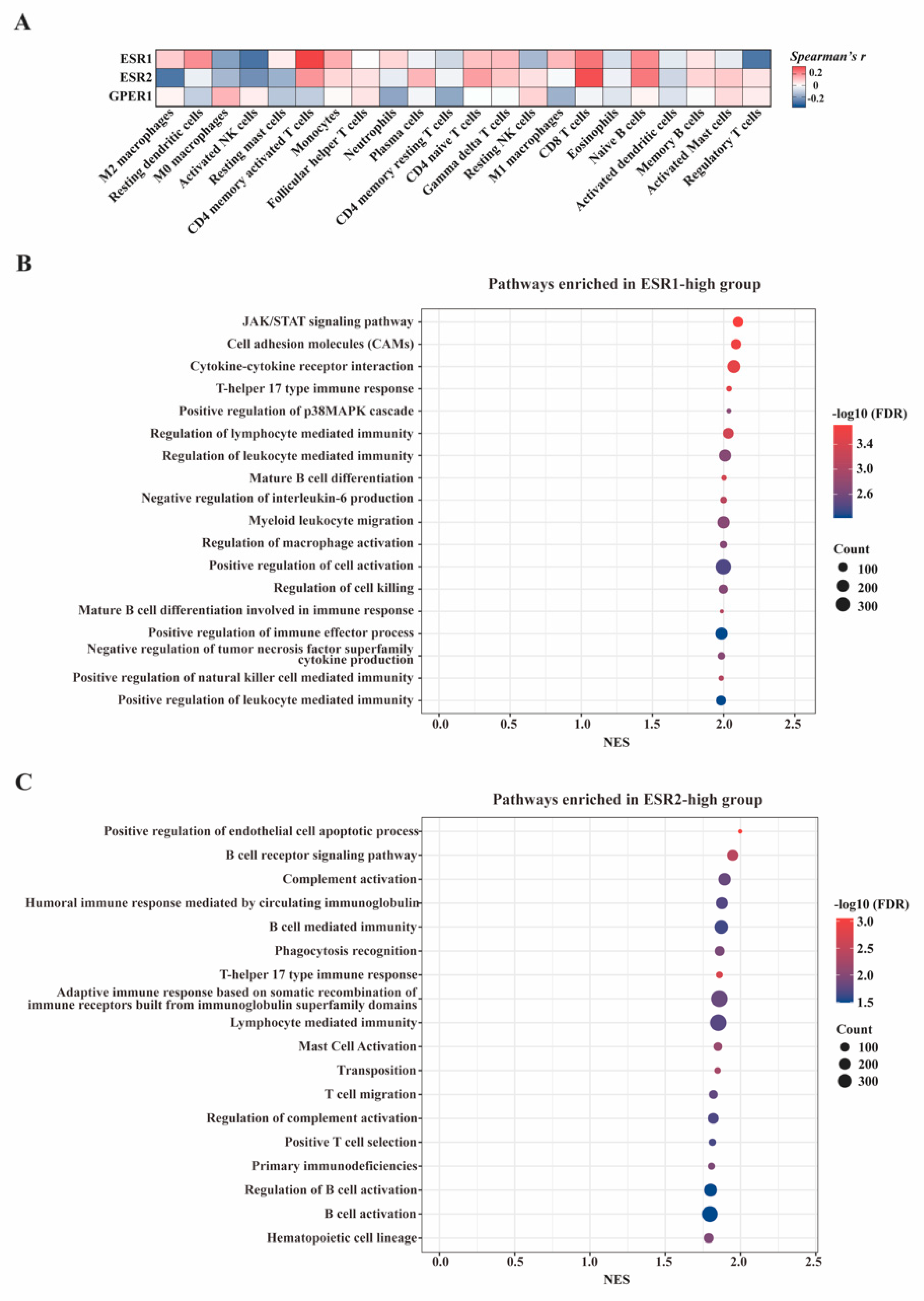
3.2. Correlation of Estrogen Receptors with TLS Development
3.3. Gender Bias in PAAD Prognosis and TLS Expression
3.4. Expression Patterns of ERs in PAAD
3.5. Association between Positive Ers Expression and TLS Presence in PAAD
3.6. In Vitro Verification of the Influence of Ers Expression on Immune Cell Chemotaxis
3.7. In-Silico Analyses of the Influence of Estrogen Receptors on Tumor Immune Microenvironment
4. Discussion
4.1. Prognostic Value of ERs in Non-Traditional Sex-Biased Cancer
4.2. Estrogen Signaling as a Target to Remodel PAAD Microenvironment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klein, A.P. Pancreatic cancer epidemiology: Understanding the role of lifestyle and inherited risk factors. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Clegg, D.; Hevener, A. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, H. Sex Hormone Receptor Signals in Human Malignancies. Int. J. Mol. Sci. 2019, 20, 2677. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Jin, J.; Qian, C.; Lou, J.; Lin, J.; Xu, A.; Xia, K.; Jin, L.; Liu, B.; Tao, H.; et al. Estrogen/ER in anti-tumor immunity regulation to tumor cell and tumor microenvironment. Cancer Cell Int. 2021, 21, 295. [Google Scholar] [CrossRef]
- Rothenberger, N.J.; Somasundaram, A.; Stabile, L. The Role of the Estrogen Pathway in the Tumor Microenvironment. Int. J. Mol. Sci. 2018, 19, 611. [Google Scholar] [CrossRef] [Green Version]
- Delaunay, F.; Pettersson, K.; Tujague, M.; Gustafsson, J.Å. Functional differences between the amino-terminal domains of estrogen receptors alpha and beta. Mol. Pharmacol. 2000, 58, 584–590. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, Z.; Jiang, G.; Zhou, Y.; Yang, X.; Huang, H.; Liu, H.; Du, J.; Wang, H. Epigenetic down regulation of G protein-coupled estrogen receptor (GPER) functions as a tumor suppressor in colorectal cancer. Mol. Cancer 2017, 16, 87. [Google Scholar] [CrossRef] [Green Version]
- Satake, M.; Sawai, H.; Go, V.L.W.; Satake, K.; Reber, H.A.; Hines, O.J.; Eibl, G. Estrogen receptors in pancreatic tumors. Pancreas 2006, 33, 119–127. [Google Scholar] [CrossRef]
- Chan, K.S.; Ho, B.C.S.; Shelat, V.G. A pilot study of estrogen receptor (ER) expression in pancreatic ductal adenocarcinoma (PDAC). Transl. Gastroenterol. Hepatol. 2021, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Younes, M.; Ly, C.J.; Singh, K.; Ertan, A.; Younes, P.S.; Bailey, J.M. Expression of estrogen receptor beta isoforms in pancreatic adenocarcinoma. Oncotarget 2018, 9, 37715–37720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunderson, A.J.; Rajamanickam, V.; Bui, C.; Bernard, B.; Pucilowska, J.; Ballesteros-Merino, C.; Schmidt, M.; McCarty, K.; Philips, M.; Piening, B.; et al. Germinal center reactions in tertiary lymphoid structures associate with neoantigen burden, humoral immunity and long-term survivorship in pancreatic cancer. Oncoimmunology 2021, 10, 1900635. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, T.N.; Thommen, D. Tertiary lymphoid structures in cancer. Science 2022, 375, eabf9419. [Google Scholar] [CrossRef] [PubMed]
- Sautès-Fridman, C.; Petitprez, F.; Calderaro, J.; Fridman, W.H. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Natale, C.A.; Li, J.; Pitarresi, J.R.; Norgard, R.J.; Dentchev, T.; Capell, B.C.; Seykora, J.T.; Stanger, B.Z.; Ridky, T.W. Pharmacologic Activation of the G Protein-Coupled Estrogen Receptor Inhibits Pancreatic Ductal Adenocarcinoma. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 868–880.e1. [Google Scholar] [CrossRef]
- Cortes, E.; Sarper, M.; Robinson, B.; Lachowski, D.; Chronopoulos, A.; Thorpe, S.D.; Lee, D.A.; Hernández, A.E.D.R. GPER is a mechanoregulator of pancreatic stellate cells and the tumor microenvironment. EMBO Rep. 2019, 20, e46556. [Google Scholar] [CrossRef] [PubMed]
- Cortes, E.; Lachowski, D.; Robinson, B.; Sarper, M.; Teppo, J.S.; Thorpe, S.D.; Lieberthal, T.J.; Iwamoto, K.; Lee, D.A.; Okada-Hatakeyama, M.; et al. Tamoxifen mechanically reprograms the tumor microenvironment via HIF-1A and reduces cancer cell survival. EMBO Rep. 2019, 20, e46557. [Google Scholar] [CrossRef]
- Cao, L.; Huang, C.; Cui Zhou, D.; Hu, Y.; Lih, T.M.; Savage, S.R.; Krug, K.; Clark, D.J.; Schnaubelt, M.; Chen, L.; et al. Proteogenomic characterization of pancreatic ductal adenocarcinoma. Cell 2021, 184, 5031–5052.e26. [Google Scholar] [CrossRef]
- Li, X.; Wan, Z.; Liu, X.; Ou, K.; Yang, L. A 12-chemokine gene signature is associated with the enhanced immunogram scores and is relevant for precision immunotherapy. Med. Oncol. 2022, 39, 43. [Google Scholar] [CrossRef]
- Tokunaga, R.; Nakagawa, S.; Sakamoto, Y.; Nakamura, K.; Naseem, M.; Izumi, D.; Kosumi, K.; Taki, K.; Higashi, T.; Miyata, T.; et al. 12-Chemokine signature, a predictor of tumor recurrence in colorectal cancer. Int. J. Cancer 2020, 147, 532–541. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Collins, J.R.; Alvord, W.G.; Roayaei, J.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. The DAVID Gene Functional Classification Tool: A novel biological module-centric algorithm to functionally analyze large gene lists. Genome Biol. 2007, 8, R183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steen, C.B.; Liu, C.L.; Alizadeh, A.A.; Newman, A.M. Profiling Cell Type Abundance and Expression in Bulk Tissues with CIBERSORTx. Methods Mol. Biol. 2020, 2117, 135–157. [Google Scholar] [PubMed]
- Liu, Y.; Xu, Z.; Zhang, Z.; Wen, G.; Sun, J.; Han, F. Efficacy and safety of TE/TEC/intensive paclitaxel neoadjuvant chemotherapy for the treatment of breast cancer. Oncol. Lett. 2019, 17, 907–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warfvinge, K.; Krause, D.N.; Maddahi, A.; Edvinsson, J.C.; Edvinsson, L.; Haanes, K.A. Estrogen receptors alpha, beta and GPER in the CNS and trigeminal system—Molecular and functional aspects. J. Headache Pain 2020, 21, 131. [Google Scholar] [CrossRef]
- Carmeci, C.; Thompson, D.A.; Ring, H.Z.; Francke, U.; Weigel, R. Identification of a gene (GPR30) with homology to the G-protein-coupled receptor superfamily associated with estrogen receptor expression in breast cancer. Genomics 1997, 45, 607–617. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Belinda, P.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, S.; Rizk, V.T.; Ma, Z.; Cheng, C.-H.; Berglund, A.E.; Coppola, D.; Khalil, F.; Mulé, J.J.; Soliman, H.H. Evaluation of invasive breast cancer samples using a 12-chemokine gene expression score: Correlation with clinical outcomes. Breast Cancer Res. 2017, 19, 71. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Ramos, C.M.; Quackenbush, J.; DeMeo, D.L. Genome-Wide Sex and Gender Differences in Cancer. Front. Oncol. 2020, 10, 597788. [Google Scholar] [CrossRef]
- Conforti, F.; Pala, L.; Bagnardi, V.; De Pas, T.; Martinetti, M.; Viale, G.; Gelber, R.D.; Goldhirsch, A. Cancer immunotherapy efficacy and patients’ sex: A systematic review and meta-analysis. Lancet Oncol. 2018, 19, 737–746. [Google Scholar] [CrossRef]
- Clocchiatti, A.; Cora, E.; Zhang, Y.; Dotto, G.P. Sexual dimorphism in cancer. Nat. Rev. Cancer 2016, 16, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Gong, X.; Yang, X.; Shang, X.; Du, Q.; Liao, Q.; Xie, R.; Chen, Y.; Xu, J. The roles of estrogen and estrogen receptors in gastrointestinal disease. Oncol. Lett. 2019, 18, 5673–5680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Liu, M.; Ding, Q.; Ji, X.; Hao, Y.; Wu, X.; Xiong, J. The direct effect of estrogen on cell viability and apoptosis in human gastric cancer cells. Mol. Cell. Biochem. 2014, 395, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, H.R.; Doucet, K.; Duke, V.; Morra, A.; Johnson, N. Estrogen prevents sustained COLO-205 human colon cancer cell growth by inducing apoptosis, decreasing c-myb protein, and decreasing transcription of the anti-apoptotic protein bcl-2. Tumour Biol. 2010, 31, 16–22. [Google Scholar] [CrossRef]
- Al-Khyatt, W.; Tufarelli, C.; Khan, R.; Iftikhar, S.Y. Selective oestrogen receptor antagonists inhibit oesophageal cancer cell proliferation in vitro. BMC Cancer 2018, 18, 121. [Google Scholar] [CrossRef] [Green Version]
- Yue, W.; Yager, J.D.; Wang, J.-P.; Jupe, E.R.; Santen, R.J. Estrogen receptor-dependent and independent mechanisms of breast cancer carcinogenesis. Steroids 2013, 78, 161–170. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef]
- Andersson, G.; Borgquist, S.; Jirström, K. Hormonal factors and pancreatic cancer risk in women: The Malmö Diet and Cancer Study. Int. J. Cancer 2018, 143, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Celik, L.; Lund, J.D.D.; Schiøtt, B. Conformational dynamics of the estrogen receptor alpha: Molecular dynamics simulations of the influence of binding site structure on protein dynamics. Biochemistry 2007, 46, 1743–1758. [Google Scholar] [CrossRef]
- Allec, S.I.; Sun, Y.; Sun, J.; Chang CE, A.; Wong, B.M. Heterogeneous CPU+GPU-Enabled Simulations for DFTB Molecular Dynamics of Large Chemical and Biological Systems. J. Chem. Theory Comput. 2019, 15, 2807–2815. [Google Scholar] [CrossRef]
- Chakraborty, B.; Byemerwa, J.; Krebs, T.; Lim, F.; Chang, C.-Y.; McDonnell, D.P. Estrogen receptor signaling in the immune system. Endocr. Rev. 2022, 44, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.; Jeganathan, V.; Chinnasamy, P.; Grimaldi, C.; Diamond, B. Differential roles of estrogen receptors α and β in control of B-cell maturation and selection. Mol. Med. 2011, 17, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, L.; Liu, X.; Ma, C.; Zhang, J.; Jiao, Y.; You, L.; Chen, Z.J.; Zhao, Y. Estrogen promotes B cell activation in vitro through down-regulating CD80 molecule expression. Gynecol. Endocrinol. 2011, 27, 593–596. [Google Scholar] [CrossRef]
- Mohammad, I.; Starskaia, I.; Nagy, T.; Guo, J.; Yatkin, E.; Väänänen, K.; Watford, W.T.; Chen, Z. Estrogen receptor α contributes to T cell-mediated autoimmune inflammation by promoting T cell activation and proliferation. Sci. Signal. 2018, 11, eaap9415. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | ERα Expression | ERβ Expression | GPER Expression | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ERα(+) | ERα(−) | Chi-Square Test | ERβ(+) | ERβ(−) | Chi-Square Test | GPER(+) | GPER(−) | Chi-Square Test | |
| Total | 72(41.4%) | 102(58.6%) | 127(73.0%) | 47(27.0%) | 134(77.0%) | 40(23.0%) | |||
| Gender | 0.475 | 0.821 | 0.954 | ||||||
| Male | 37(21.3%) | 58(33.3%) | 70(40.2%) | 25(14.4%) | 73(42.0%) | 22(12.6%) | |||
| Female | 35(20.1) | 44(25.3%) | 57(32.8%) | 22(12.6%) | 61(35.1%) | 18(10.3%) | |||
| Age | 0.413 | 0.628 | 0.453 | ||||||
| ≤60 | 35(20.1%) | 56(32.2%) | 65(37.4%) | 26(14.9%) | 68(39.1%) | 23(13.2%) | |||
| >60 | 37(21.3%) | 46(26.4%) | 62(35.6%) | 21(12.1%) | 66(37.9%) | 17(9.8%) | |||
| TNM Stage | 0.153 | 0.017 | 0.006 | ||||||
| I/II | 50(28.7%) | 60(34.5%) | 87(50.0%) | 23(13.2%) | 92(52.9%) | 18(10.3%) | |||
| III/IV | 22(12.6%) | 42(24.1%) | 40(23.0%) | 24(13.8%) | 42(24.1%) | 22(12.6%) | |||
| T | 0.470 | 0.002 | 0.024 | ||||||
| T1/T2 | 49(28.2%) | 64(36.8%) | 91(52.3%) | 22(12.6%) | 93(53.4%) | 20(11.5%) | |||
| T3/T4 | 23(13.2%) | 38(21.8%) | 36(20.7%) | 25(14.4%) | 41(23.6%) | 20(11.5%) | |||
| N | 0.733 | 0.245 | 0.826 | ||||||
| N0 | 40(23.0%) | 54(31.0%) | 72(41.4%) | 22(12.6%) | 73(42.0%) | 21(12.1%) | |||
| N1/N2 | 32 | 48(27.6%) | 55(31.6%) | 25(14.4%) | 61(35.1%) | 19(10.9%) | |||
| M | 0.216 | 0.938 | 0.090 | ||||||
| M0 | 67(38.5%) | 89(51.1%) | 114(65.5%) | 42(24.1%) | 123(70.7%) | 33(19.0%) | |||
| M1 | 5(2.9%) | 13(7.5%) | 13(7.5%) | 5(2.9%) | 11(6.3%) | 7(4.0%) | |||
| Grade | 0.011 | 0.023 | 0.200 | ||||||
| G1/2 | 23(13.2%) | 16(9.2%) | 34(19.5%) | 5(2.9%) | 33(19.0%) | 6(3.4%) | |||
| G3 | 49(28.2%) | 86(49.4%) | 93(53.4%) | 42(24.1%) | 101(58.0%) | 34(19.5%) | |||
| Nerve Invasion | 0.975 | 0.453 | 0.439 | ||||||
| No | 10(5.7%) | 14(8.0%) | 16(9.2%) | 8(4.6%) | 17(9.8%) | 7(4.0%) | |||
| Yes | 62(35.6%) | 88(50.6%) | 111(63.8%) | 39(22.4%) | 117(67.2%) | 33(19.0%) | |||
| Vascular Invasion | 0.256 | 0.221 | 0.107 | ||||||
| No | 57(32.8%) | 73(42.0%) | 98(56.3%) | 32(18.4%) | 104(59.8%) | 26(14.9%) | |||
| Yes | 15(8.6%) | 29(16.7%) | 29(16.7%) | 15(8.6%) | 30(17.2%) | 14(8.0%) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Liu, Y.; Lin, X.; Wang, R.; Dai, Z.; Chen, Y.; Ma, M.; Tasiheng, Y.; Yan, Y.; Wang, X.; et al. Characterization of Estrogen Receptors in Pancreatic Adenocarcinoma with Tertiary Lymphoid Structures. Cancers 2023, 15, 828. https://doi.org/10.3390/cancers15030828
Zou X, Liu Y, Lin X, Wang R, Dai Z, Chen Y, Ma M, Tasiheng Y, Yan Y, Wang X, et al. Characterization of Estrogen Receptors in Pancreatic Adenocarcinoma with Tertiary Lymphoid Structures. Cancers. 2023; 15(3):828. https://doi.org/10.3390/cancers15030828
Chicago/Turabian StyleZou, Xuan, Yu Liu, Xuan Lin, Ruijie Wang, Zhengjie Dai, Yusheng Chen, Mingjian Ma, Yesiboli Tasiheng, Yu Yan, Xu Wang, and et al. 2023. "Characterization of Estrogen Receptors in Pancreatic Adenocarcinoma with Tertiary Lymphoid Structures" Cancers 15, no. 3: 828. https://doi.org/10.3390/cancers15030828