
Melanin—The Éminence Grise of Melanoma and Parkinson’s Disease Development

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Origin of Melanocytes and Neurons of the Substantia Nigra
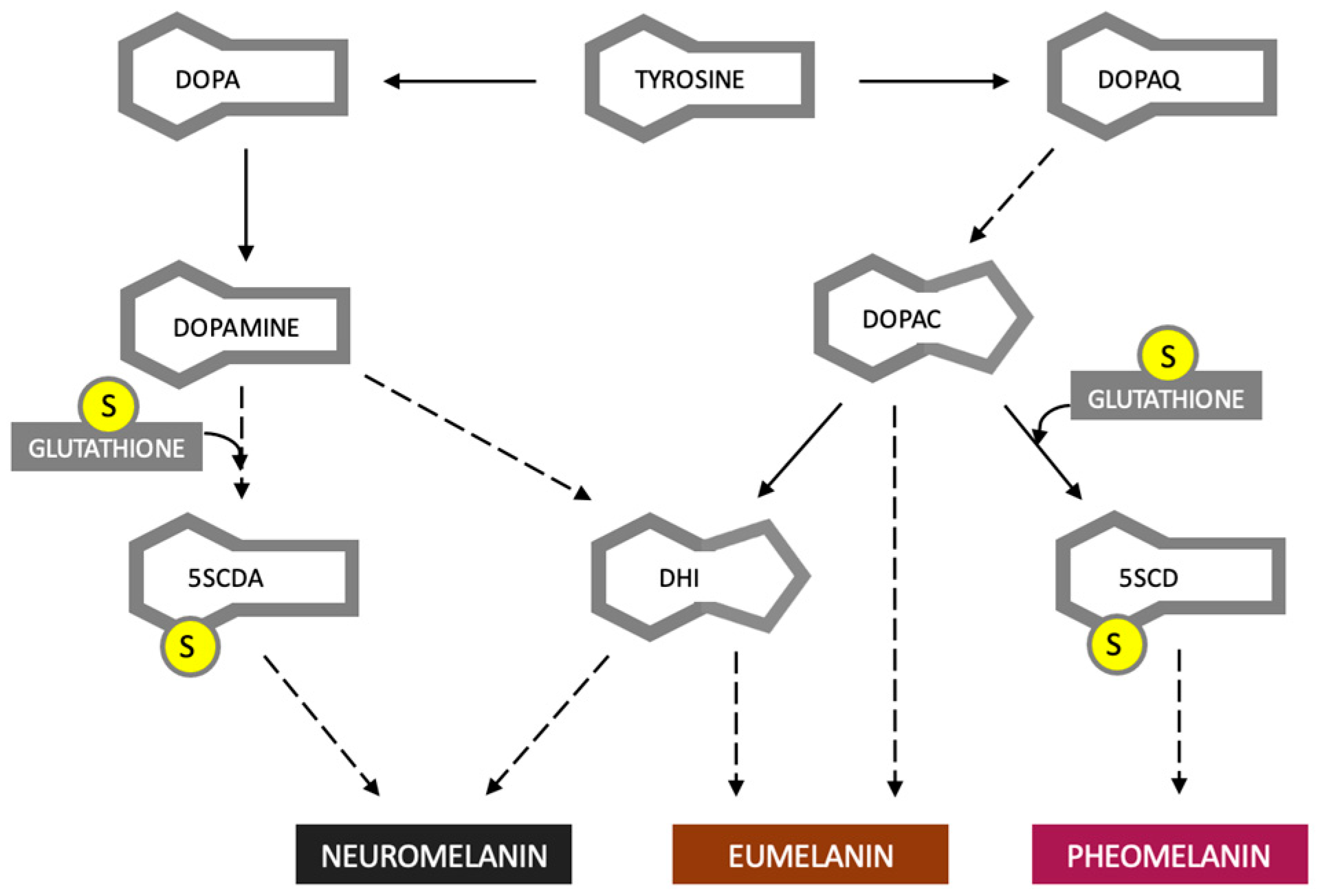
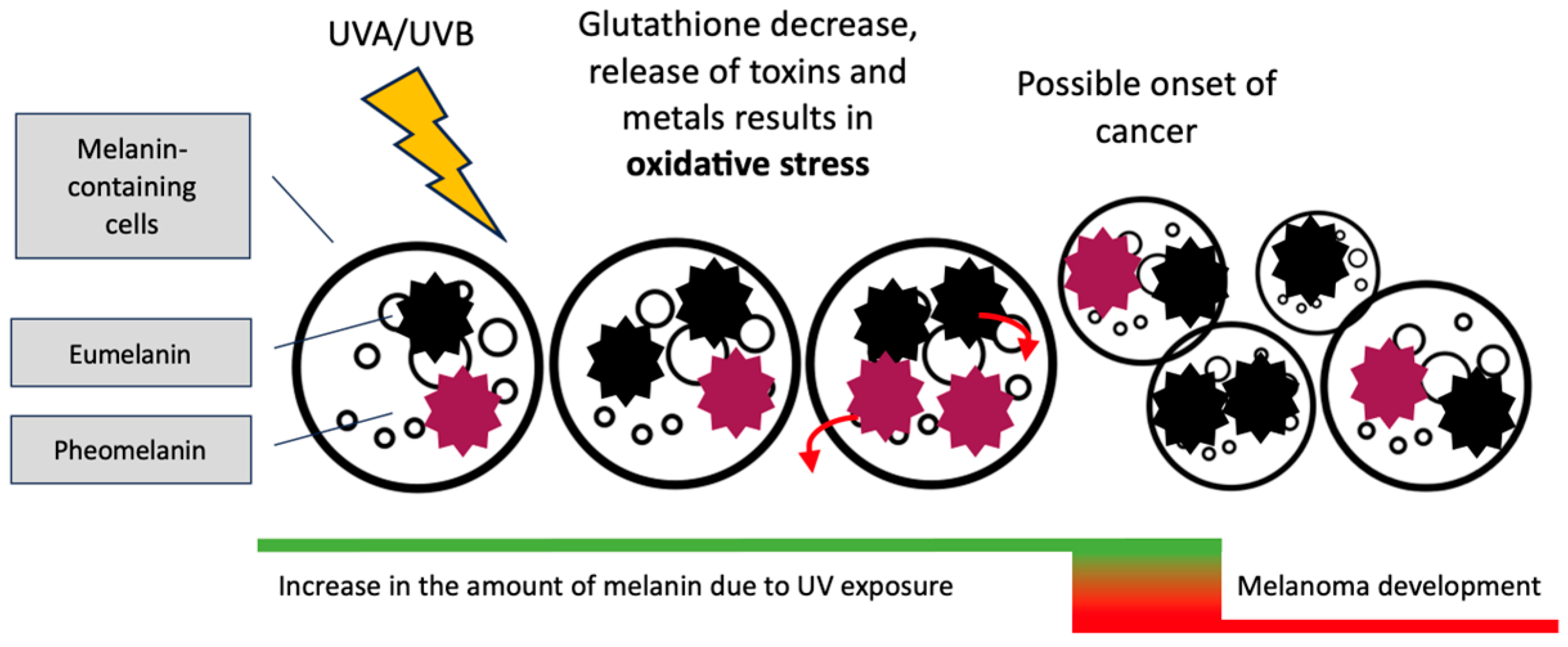
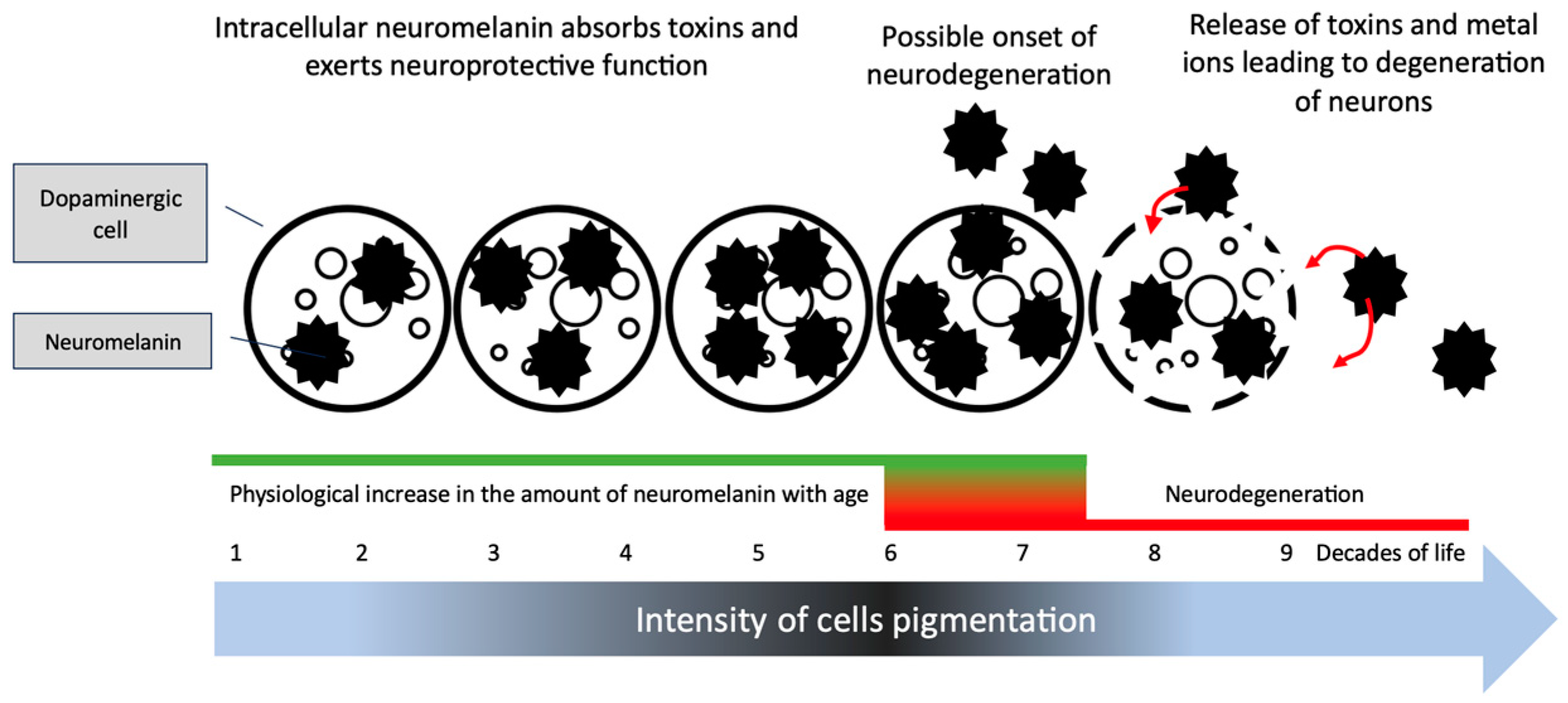
3. Eumelanin, Pheomelanin and Neuromelanin; Do Dyes Cause Trouble in the Cell?
4. Treatment of PD and Development of Melanoma
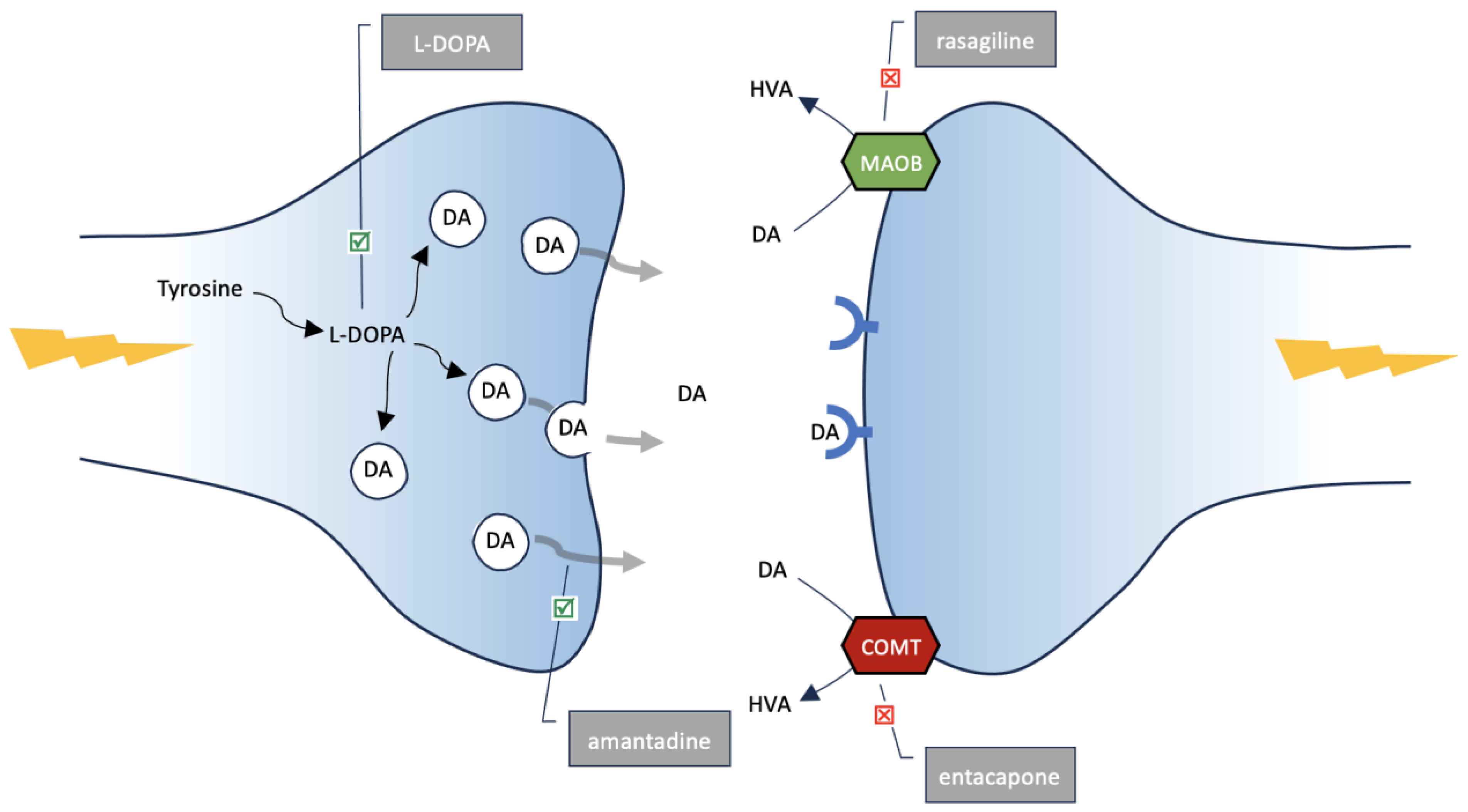
5. L-DOPA
6. Amantadine, MAO-B and COMT Inhibitors
7. Future and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, A. The biology of melanocyte and melanocyte stem cell. Acta Biochim. Biophys. Sin. 2014, 46, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Caini, S.; Gandini, S.; Sera, F.; Raimondi, S.; Fargnoli, M.C.; Boniol, M.; Armstrong, B.K. Meta-analysis of risk factors for cutaneous melanoma according to anatomical site and clinico-pathological variant. Eur. J. Cancer 2009, 45, 3054–3063. [Google Scholar] [CrossRef] [PubMed]
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, Risk Factors, Pathogenesis, Diagnosis and Classification. In Vivo 2014, 28, 1005–1011. Available online: http://iv.iiarjournals.org/content/28/6/1005.abstract (accessed on 14 November 2014). [PubMed]
- Dzwierzynski, W.W. Melanoma Risk Factors and Prevention. Clin. Plast. Surg. 2021, 48, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Tysnes, O.-B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural Transm. 2017, 124, 901–905. [Google Scholar] [CrossRef]
- Schapira, A.H.; Jenner, P. Etiology and pathogenesis of Parkinson’s disease. Mov. Disord. 2011, 26, 1049–1055. [Google Scholar] [CrossRef]
- Nagatsu, T.; Nakashima, A.; Watanabe, H.; Ito, S.; Wakamatsu, K.; Zucca, F.A.; Zecca, L.; Youdim, M.; Wulf, M.; Riederer, P.; et al. The role of tyrosine hydroxylase as a key player in neuromelanin synthesis and the association of neuromelanin with Parkinson’s disease. J. Neural Transm. 2023, 130, 611–625. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Chemistry of Mixed Melanogenesis—Pivotal Roles of Dopaquinone. Photochem. Photobiol. 2007, 84, 582–592. [Google Scholar] [CrossRef]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tymińska, A. Skin melanocytes: Biology and development. Postepy Dermatol Alergol. Adv. Dermatol. Allergol. Postępy Dermatol. I Alergol. 2013, 30, 30–41. [Google Scholar] [CrossRef]
- Mort, R.L.; Jackson, I.J.; Patton, E.E. The melanocyte lineage in development and disease. Development 2015, 142, 620–632. [Google Scholar] [CrossRef]
- Erickson, C.A.; Goins, T.L. Avian neural crest cells can migrate in the dorsolateral path only if they are specified as melanocytes. Development 1995, 121, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Paxinos, G.; Watson, C.; Halliday, G.M. The substantia nigra and ventral tegmental dopaminergic neurons from development to degeneration. J. Chem. Neuroanat. 2016, 76, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Marchand, R.; Poirier, L. Isthmic origin of neurons of the rat substantia nigra. Neuroscience 1983, 9, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Aubert, I.; Brana, C.; Pellevoisin, C.; Giros, B.; Caille, I.; Carles, D.; Vital, C.; Bloch, B. Molecular anatomy of the development of the human substantia nigra. J. Comp. Neurol. 1997, 379, 72–87. [Google Scholar] [CrossRef]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef]
- Nagatsu, T.; Nakashima, A.; Watanabe, H.; Ito, S.; Wakamatsu, K. Neuromelanin in Parkinson’s Disease: Tyrosine Hydroxylase and Tyrosinase. Int. J. Mol. Sci. 2022, 23, 4176. [Google Scholar] [CrossRef]
- Sansinenea, E.; Ortiz, A. Melanin: A Solution for Photoprotection of Bacillus thuringiensis Based Biopesticides. Biotechnol. Lett. 2015, 37, 483. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin Against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- Sample, A.; He, Y. Mechanisms and prevention of UV-induced melanoma. Photodermatol. Photoimmunol. Photomed. 2017, 34, 13–24. [Google Scholar] [CrossRef]
- Jin, S.G.; Padron, F.; Pfeifer, G.P. UVA Radiation, DNA Damage, and Melanoma. ACS Omega 2022, 7, 32936–32948. [Google Scholar] [CrossRef]
- Mostert, A.B. Melanin, the What, the Why and the How: An Introductory Review for Materials Scientists Interested in Flexible and Versatile Polymers. Polymers 2021, 13, 1670. [Google Scholar] [CrossRef] [PubMed]
- Dadachova, E.; Casadevall, A. Ionizing radiation: How fungi cope, adapt, and exploit with the help of melanin. Curr. Opin. Microbiol. 2008, 11, 525–531. Available online: https://www.sciencedirect.com/science/article/pii/S1369527408001306 (accessed on 24 October 2008). [CrossRef] [PubMed]
- Caër, S.L. Water radiolysis: Influence of oxide surfaces on H2 production under ionizing radiation. Water 2011, 3, 235–253. [Google Scholar] [CrossRef]
- Hu, S.; Parmet, Y.; Allen, G.; Parker, D.F.; Ma, F.; Rouhani, P.; Kirsner, R.S. Disparity in melanoma: A trend analysis of melanoma incidence and stage at diagnosis among whites, Hispanics, and blacks in Florida. Arch. Dermatol. 2009, 145, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Yamashita, Y.; Umezawa, K.; Hirobe, T.; Ito, S.; Wakamatsu, K. The Pro-Oxidant Activity of Pheomelanin is Significantly Enhanced by UVA Irradiation: Benzothiazole Moieties Are More Reactive than Benzothiazine Moieties. Int. J. Mol. Sci. 2018, 19, 2889. [Google Scholar] [CrossRef] [PubMed]
- Lembo, S.; Di Caprio, R.; Micillo, R.; Balato, A.; Monfrecola, G.; Panzella, L.; Napolitano, A. Light-independent pro-inflammatory and pro-oxidant effects of purified human hair melanins on keratinocyte cell cultures. Exp. Dermatol. 2017, 26, 592–594. [Google Scholar] [CrossRef]
- Premi, S.; Wallisch, S.; Mano, C.M.; Weiner, A.B.; Bacchiocchi, A.; Wakamatsu, K.; Bechara, E.J.H.; Halaban, R.; Douki, T.; Brash, D.E. Chemiexcitation of melanin derivatives induces DNA photoproducts long after UV exposure. Science 2015, 347, 842–847. [Google Scholar] [CrossRef]
- Kim, E.; Panzella, L.; Napolitano, A.; Payne, G.F. Redox Activities of Melanins Investigated by Electrochemical Reverse Engineering: Implications for their Roles in Oxidative Stress. J. Investig. Dermatol. 2019, 140, 537–543. [Google Scholar] [CrossRef]
- Tobin, D.J. Human hair pigmentation—Biological aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef]
- Siwicka, Z.E.; Son, F.A.; Battistella, C.; Moore, M.H.; Korpanty, J.; McCallum, N.C.; Wang, Z.; Johnson, B.J.; Farha, O.K.; Gianneschi, N.C. Synthetic Porous Melanin. J. Am. Chem. Soc. 2021, 143, 3094–3103. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K.; Sarna, T. Photodegradation of Eumelanin and Pheomelanin and Its Pathophysiological Implications. Photochem. Photobiol. 2018, 94, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Premraj, L.; Khoo, T.K. Neuromelanin: Its role in the pathogenesis of idiopathic Parkinson’s disease and potential as a therapeutic target. Park. Relat. Disord. 2023, 112, 105448. [Google Scholar] [CrossRef] [PubMed]
- Zuarth Gonzalez, J.; Shahbazi Nia, S.; Ortiz, Y.T.; McMahon, L.R.; German, N.; Wilkerson, J.L. Dementia, Dopamine and Medicinal Chemistry Drug Design. West Tex. J. Med. 2023, 1, 1–23. [Google Scholar]
- Fedorow, H.; Tribl, F.; Halliday, G.; Gerlach, M.; Riederer, P.; Double, K. Neuromelanin in human dopamine neurons: Comparison with peripheral melanins and relevance to Parkinson’s disease. Prog. Neurobiol. 2005, 75, 109–124. [Google Scholar] [CrossRef]
- Drukarch, B.; van Muiswinkel, F.L. Drug treatment of Parkinson’s disease: Time for phase II. Biochem. Pharmacol. 2000, 59, 1023–1031. Available online: https://www.sciencedirect.com/science/article/pii/S0006295299003408 (accessed on 8 March 2000). [CrossRef] [PubMed]
- Foley, J.M.; Banter, D. On the Nature of Pigment Granules in the Cells of the Locus Coeruleus and Substantia Nigra. J. Neuropathol. Exp. Neurol. 1958, 17, 586–598. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Giménez, J.; Cuadros, T.; Bové, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepúlveda, M.; Peñuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 973. [Google Scholar] [CrossRef]
- Zecca, L.; Fariello, R.; Riederer, P.; Sulzer, D.; Gatti, A.; Tampellini, D. The absolute concentration of nigral neuromelanin, assayed by a new sensitive method, increases throughout the life and is dramatically decreased in Parkinson’s disease. FEBS Lett. 2002, 510, 216–220. [Google Scholar] [CrossRef]
- Biondetti, E.; Gaurav, R.; Yahia-Cherif, L.; Mangone, G.; Pyatigorskaya, N.; Valabrègue, R.; Ewenczyk, C.; Hutchison, M.; François, C.; Arnulf, I.; et al. Spatiotemporal changes in substantia nigra neuromelanin content in Parkinson’s disease. Brain 2020, 143, 2757–2770. [Google Scholar] [CrossRef]
- Zucca, F.A.; Basso, E.; Cupaioli, F.A.; Ferrari, E.; Sulzer, D.; Casella, L.; Zecca, L. Neuromelanin of the Human Substantia Nigra: An Update. Neurotox. Res. 2013, 25, 13–23. [Google Scholar] [CrossRef]
- D’Amato, R.J.; Alexander, G.M.; Schwartzman, R.J.; Kitt, C.A.; Price, D.L.; Snyder, S.H. Evidence for neuromelanin involvement in MPTP-induced neurotoxicity. Nature 1987, 327, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Korzhevskii, D.E.; Kirik, O.V.; Guselnikova, V.V.; Tsyba, D.L.; Fedorova, E.A.; Grigorev, I.P. Changes in cytoplasmic and extracellular neuromelanin in human substantia nigra with normal aging. Eur. J. Histochem. 2021, 65 (Suppl. S1), 3283. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Rey, N. Animal Models of Parkinson’s Disease. In Challenges in Parkinson′s Disease; Dorszewska, J., Kozubski, W., Eds.; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef]
- Nakamura, K.; Wang, W.; Kang, U.J. The role of glutathione in dopaminergic neuronal survival. J. Neurochem. 2002, 69, 1850–1858. [Google Scholar] [CrossRef] [PubMed]
- Sian, J.; Dexter, D.T.; Lees, A.J.; Daniel, S.; Jenner, P.; Marsden, C.D. Glutathione-related enzymes in brain in Parkinson’s disease. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1994, 36, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Thouvenot, É.; Camu, W. Vitamine D et neurologie. La Presse Médicale 2013, 42, 1398–1404. [Google Scholar] [CrossRef]
- Behl, T.; Arora, A.; Singla, R.K.; Sehgal, A.; Makeen, H.A.; Albratty, M.; Meraya, A.M.; Najmi, A.; Bungau, S.G. Understanding the role of “sunshine vitamin D” in Parkinson’s disease: A review. Front. Pharmacol. 2022, 13, 993033. [Google Scholar] [CrossRef]
- Paolino, G.; Moliterni, E.; Didona, D.; Garelli, V.; Corsetti, P.; Lopez, T.; Richetta, A.G.; Cantisani, C.; Bottoni, U.; Calvieri, S. Clinicopathological features, vitamin D serological levels and prognosis in cutaneous melanoma of shield-sites: An update. Med. Oncol. 2014, 32, 451. [Google Scholar] [CrossRef]
- Holick, M.F. Sunlight, UV Radiation, Vitamin D, and Skin Cancer: How Much Sunlight Do We Need? In Sunlight, Vitamin D and Skin Cancer; Reichrath, J., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 19–36. [Google Scholar] [CrossRef]
- Salemi, M.; Mogavero, M.P.; Lanza, G.; Mongioì, L.M.; Calogero, A.E.; Ferri, R. Examples of Inverse Comorbidity between Cancer and Neurodegenerative Diseases: A Possible Role for Noncoding RNA. Cells 2022, 11, 1930. [Google Scholar] [CrossRef]
- Dalvin, L.A.; Damento, G.M.; Yawn, B.P.; Abbott, B.A.; Hodge, D.O.; Pulido, J.S. Parkinson Disease and Melanoma: Confirming and Reexamining an Association. Mayo Clin. Proc. 2017, 92, 1070–1079. [Google Scholar] [CrossRef]
- Vermeij, J.D.; Winogrodzka, A.; Trip, J.; Weber, W.E.J. Parkinson’s disease, levodopa-use and the risk of melanoma. Park. Relat. Disord. 2009, 15, 551–553. Available online: https://www.sciencedirect.com/science/article/pii/S1353802009001151 (accessed on 5 June 2009). [CrossRef]
- Krasowska, D.; Gerkowicz, A.; Mlak, R.; Leziak, M.; Małecka-Massalska, T.; Krasowska, D. Risk of Nonmelanoma Skin Cancers and Parkinson’s Disease—Meta-Analysis and Systematic Review. Cancers 2021, 13, 587. [Google Scholar] [CrossRef]
- Dean, D.N.; Lee, J.C. Defining an amyloid link Between Parkinson’s disease and melanoma. Proc. Natl. Acad. Sci. USA 2020, 117, 22671–22673. [Google Scholar] [CrossRef] [PubMed]
- Marmol, S.; Feldman, M.; Singer, C.; Margolesky, J. Amantadine Revisited: A Contender for Initial Treatment in Parkinson’s Disease? CNS Drugs 2021, 35, 1141–1152. [Google Scholar] [CrossRef]
- Haasio, K. Toxicology and Safety of Comt Inhibitors. In International Review of Neurobiology; Nissinen, E., Ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 163–189. Available online: https://www.sciencedirect.com/science/article/pii/B9780123813268000077 (accessed on 21 November 2010).
- Weinreb, O.; Amit, T.; Bar-Am, O.; Youdim, M.B. Rasagiline: A novel anti-Parkinsonian monoamine oxidase-B inhibitor with neuroprotective activity. Prog. Neurobiol. 2010, 92, 330–344. [Google Scholar] [CrossRef]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jung, H.; Kim, K.; Lim, K.-M.; Kim, J.-Y.; Jho, E.-H.; Oh, E.-S. D-tyrosine negatively regulates melanin synthesis by competitively inhibiting tyrosinase activity. Pigment Cell Melanoma Res. 2017, 31, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.L.; Haupt, H.M.; Stern, J.B.; Multhaupt, H.A.B. Tyrosinase expression in malignant melanoma, desmoplastic melanoma, and peripheral nerve tumors: An immunohistochemical study. Arch. Pathol. Lab. Med. 2002, 126, 816–822. [Google Scholar] [CrossRef]
- Inzelberg, R.; Israeli-Korn, S.D. The particular relationship between Parkinson’s disease and malignancy: A focus on skin cancers. J. Neural Transm. 2009, 116, 1503–1507. [Google Scholar] [CrossRef]
- Haraszti, S.; Ezaldein, H.; Delost, G.R. Eruptive melanocytic nevi in a patient with Parkinson disease treated by carbidopa-levodopa. JAAD Case Rep. 2018, 5, 21–23. [Google Scholar] [CrossRef]
- Porcar Saura, S.; Guillén Climent, S.; Pinazo Canales, M.I.; Monteagudo, C. Melanocytic Hyperactivation Simulating an Acral Lentiginous Melanoma in a Patient With Parkinson Disease Treated by Levodopa. Am. J. Dermatopathol. 2021, 43, 238–241. Available online: https://journals.lww.com/amjdermatopathology/fulltext/2021/03000/melanocytic_hyperactivation_simulating_an_acral.18.aspx (accessed on 22 September 2020). [CrossRef]
- Bougea, A.; Spantideas, N.; Katoulis, A.; Stefanis, L. Levodopa-induced skin disorders in patients with Parkinson disease: A systematic literature review approach. Acta Neurol. Belg. 2019, 119, 325–336. [Google Scholar] [CrossRef]
- Olsen, J.H.; Tangerud, K.; Wermuth, L.; Frederiksen, K.; Friis, S. Treatment with levodopa and risk for malignant melanoma. Mov. Disord. 2007, 22, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Kannengiesser, C.; Dalle, S.; Leccia, M.-T.; Avril, M.F.; Bonadona, V.; Chompret, A.; Lasset, C.; Leroux, D.; Thomas, L.; Lesueur, F.; et al. New founder germline mutations ofCDKN2A in melanoma-prone families and multiple primary melanoma development in a patient receiving levodopa treatment. Genes Chromosom. Cancer 2007, 46, 751–760. [Google Scholar] [CrossRef]
- Zanetti, R.; Rosso, S. Levodopa and the risk of melanoma. Lancet 2007, 369, 257–258. Available online: https://www.sciencedirect.com/science/article/pii/S0140673607601251 (accessed on 27 January 2007). [CrossRef] [PubMed]
- Horecka, A.; Szpetnar, M.; Hordyjewska, A.; Babula, D.; Gołąb, P.; Kurzepa, J. Actylise treatment does not influence nitric oxide metabolites serum level. Pharmacol. Rep. 2016, 68, 598–600. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Li, Q.; Cheng, B.; Jing, L. Primary culture of human face skin melanocytes for the study of hyperpigmentation. Cytotechnology 2013, 66, 891–898. [Google Scholar] [CrossRef]
- Peeters, M.; Page, G.; Maloteaux, J.-M.; Hermans, E. Hypersensitivity of dopamine transmission in the rat striatum after treatment with the NMDA receptor antagonist amantadine. Brain Res. 2002, 949, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Krasowska, D.; Gerkowicz, A.; Wróblewska-Łuczka, P.; Grabarska, A.; Załuska-Ogryzek, K.; Krasowska, D.; Łuszczki, J.J. Anticancer Activity of Amantadine and Evaluation of Its Interactions with Selected Cytostatics in Relation to Human Melanoma Cells. Int. J. Mol. Sci. 2022, 23, 7653. [Google Scholar] [CrossRef] [PubMed]
- Johannes, C.B.; Saltus, C.W.; Kaye, J.A.; Calingaert, B.; Kaplan, S.; Gordon, M.F.; Andrews, E.B. The risk of melanoma with rasagiline compared with other antiparkinsonian medications: A retrospective cohort study in the United States medicare database. Pharmacoepidemiol. Drug Saf. 2022, 31, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Meier-Davis, S.R.; Dines, K.; Arjmand, F.M.; Hamlin, R.; Huang, B.; Wen, J.; Christianson, C.; Shudo, J.; Nagata, T. Comparison of oral and transdermal administration of rasagiline mesylate on human melanoma tumor growthin vivo. Cutan. Ocul. Toxicol. 2012, 31, 312–317. [Google Scholar] [CrossRef]
- Magina, S.; Vieira-Coelho, M.A.; Serrão, M.P.; Kosmus, C.; Moura, E.; Moura, D. Ultraviolet B radiation differentially modifies catechol-O-methyltransferase activity in keratinocytes and melanoma cells. Photodermatol. Photoimmunol. Photomed. 2012, 28, 137–141. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krasowska, D.; Małek, A.; Kurzepa, J.; Kapka-Skrzypczak, L.; Krasowska, D.; Kurzepa, J. Melanin—The Éminence Grise of Melanoma and Parkinson’s Disease Development. Cancers 2023, 15, 5541. https://doi.org/10.3390/cancers15235541
Krasowska D, Małek A, Kurzepa J, Kapka-Skrzypczak L, Krasowska D, Kurzepa J. Melanin—The Éminence Grise of Melanoma and Parkinson’s Disease Development. Cancers. 2023; 15(23):5541. https://doi.org/10.3390/cancers15235541
Chicago/Turabian StyleKrasowska, Danuta, Agata Małek, Joanna Kurzepa, Lucyna Kapka-Skrzypczak, Dorota Krasowska, and Jacek Kurzepa. 2023. "Melanin—The Éminence Grise of Melanoma and Parkinson’s Disease Development" Cancers 15, no. 23: 5541. https://doi.org/10.3390/cancers15235541