
Nasopharynx Battlefield: Cellular Immune Responses Mediated by Midkine in Nasopharyngeal Carcinoma and COVID-19

Abstract
:Simple Summary
Abstract
1. Introduction
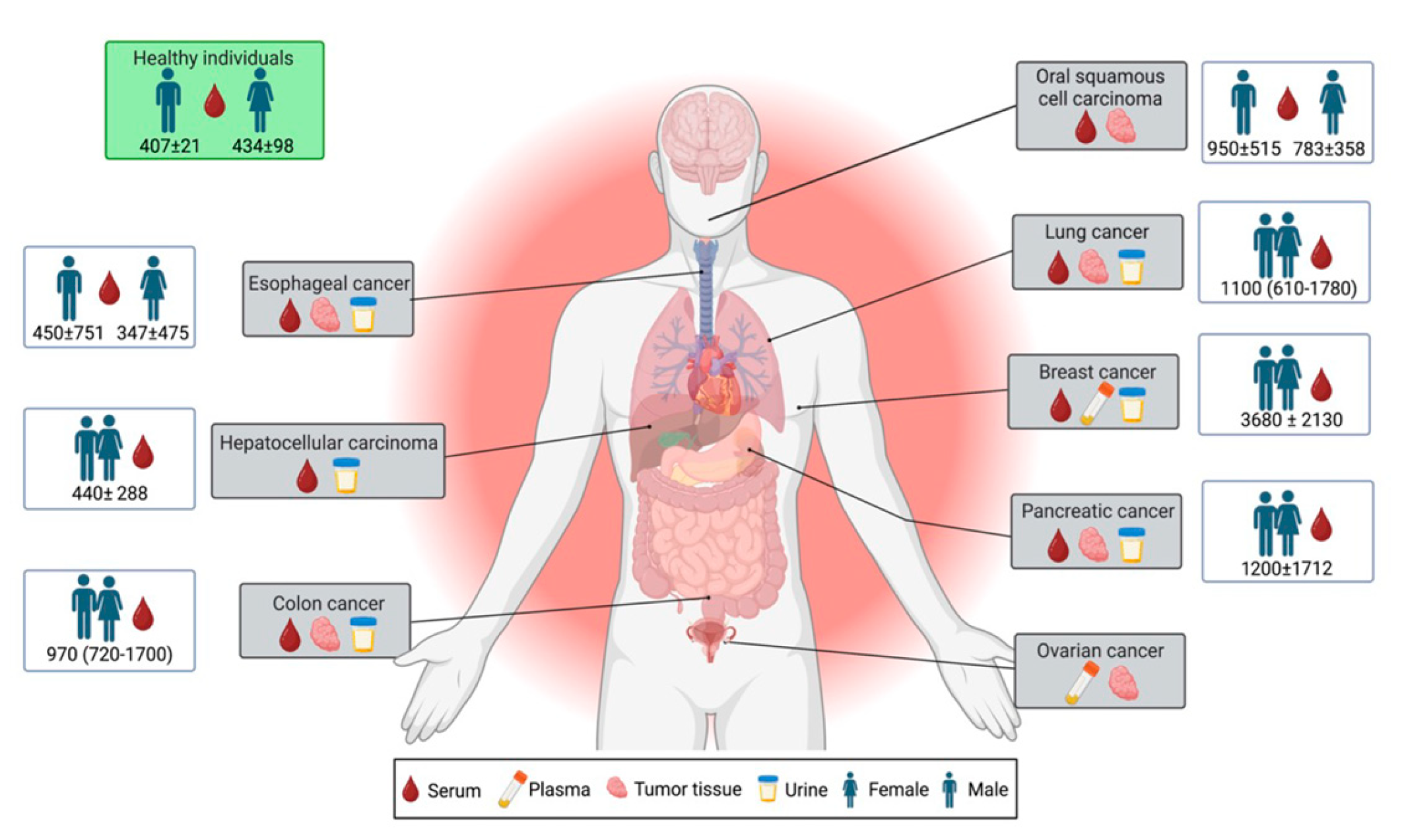
2. Midkine Physiological and Pathophysiological Properties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MK-Binding Receptors | Receptor Location | Downstream Signaling Network | Association with Biology/Responses | Ref. |
|---|---|---|---|---|
| Non-immune cells | ||||
| Protein tyrosine phosphatase ζ (PTPRZ1) | Embryonic stem cell | PI3K/AKT signaling pathway | Embryonic stem cell self-renewal | [19] |
| Notch2 | Keratinocytes | Notch2/Jak2/Stat3 signaling pathway | Cell plasticity and motility regulation | [26] |
| Syndecan-1 (SDC-1) | Neuronal cells | Neurite growth and neuronal cells migratory signaling | Early stage of neurogenesis | [20] |
| Syndecan-3 (SDC-3) | Development in post-natal period of neurogenesis | |||
| Syndecan-4 (SDC-4, ryudocan) | Peripheral nerve bundles | Neurite promoting signaling | Formation of neural network | [21] |
| ALK | Fibroblasts, cancer cells, endothelial cells | PI3K and MAPK signaling | Cell growth, survival and angiogenesis | [25] |
| α6β1-integrins | Neurons | Growth signaling | Neurite outgrowth of embryonic neurons | [27] |
| α4β1- integrins | Osteoblastic cells | Migratory signaling | Haptotactic migration of osteoblastic cells | |
| Neuroglycan C | Oligodendrocyte precursor-like cell | Cytoskeletal modification, process extension and cell spreading | Neural network development | [37] |
| Low-density lipoprotein receptor-related protein (LRP-1) | Synovial fibroblasts | IL-6, IL-8 and CCL2 signaling pathway | Immune cells accumulation | [23] |
| Fibroblast | Endocytosis of MK, nuclear targeting | Cell survival | [24] | |
| Nucleolin (NCL) | Fibroblast cytoplasm | Nuclear translocation of MK | ||
| Laminin binding protein precursor (LBP) | Rectal carcinoma cell | Nuclear translocation of MK | Tumor progression | [38] |
| Glypican-2 | Neuronal cell | Neurite growth and neuronal cells migratory signaling | Neuronal development at embryonic stage | [22] |
| TSPAN1 | Head and neck squamous cell carcinomas (HNSCC) cells | FAK phosphorylation and Stat1a pathway activation | Cell migration | [39] |
| NCAM | Embryonic cells | Unknown | Unknown | [40] |
| Brushin/megalin | ||||
| PG-M/versican | Embryonic cells | MK inhibition or promotion depending on the strength of binding | Unknown | [41] |
| LRP6 | Embryonic neurons | Unknown | Neurons survival | [42] |
| apoER2 | ||||
| Immune cells | ||||
| LRP-1 | Polymorphonuclear neutrophils (PMN) | PMN recruitment by β2 integrin-dependent axis | PMN extravasation, trafficking and adhesion to inflammatory sites | [43,44] |
| CD8+ T cells | Calcineurin/NFAT1 signaling | Naïve CD8+ T cells activation to produce CCL4, thus inducing microglia to produce CCL5 | [32] | |
| Macrophages and tubular epithelial cells | MCP-1 pathway | Macrophage infiltration | [45] | |
| Macrophage | Increase M2 phenotype marker, arginase-1 expression | Alter microglia/macrophages polarization status | [46] | |
| NCL | T cells | Formation of stable complex with NCL | Autocrine and paracrine regulate HIV infection | [47] |
| PTPRZ1 | B cells | MIF/CD74 and HGF/c-Met signaling | B cells survival | [36] |
3. MK Contributes to Carcinogenesis and Cancer Progression
3.1. Significance of MK in Tumor Microenvironment
3.2. Intercellular Communication of MK in NPC
3.3. Ligand–Receptor Pairs in Disease in NPC
4. Possible Contribution of MK to the Regulation of COVID-19
4.1. Intercellular Communication of MK in COVID-19
4.2. Ligand–Receptor Pairs in Disease in COVID-19
5. Impact of COVID-19 on Outcomes for Patients with NPC
5.1. Managing NPC and COVID-19: Challenges and Clinical Evidence for Standard Care Therapies
5.2. The Potential Influence of MK-Mediated Cellular Immunity on COVID-19-Contracted NPC Cancer Progression and Therapy Response
6. Implications for Therapies
Clinical Benefits of Therapies against MK Signaling
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. COVID-19 Weekly Epidemiological Update. 2023. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---27-april-2023 (accessed on 12 May 2023).
- Sun, Y.; Luo, B.; Liu, Y.; Wu, Y.; Chen, Y. Immune damage mechanisms of COVID-19 and novel strategies in prevention and control of epidemic. Front. Immunol. 2023, 14, 1130398. [Google Scholar] [CrossRef] [PubMed]
- Montazersaheb, S.; Hosseiniyan Khatibi, S.M.; Hejazi, M.S.; Tarhriz, V.; Farjami, A.; Ghasemian Sorbeni, F.; Farahzadi, R.; Ghasemnejad, T. COVID-19 infection: An overview on cytokine storm and related interventions. Virol. J. 2022, 19, 92. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Ruan, F.; Huang, M.; Liang, L.; Huang, H.; Hong, Z.; Yu, J.; Kang, M.; Song, Y.; Xia, J.; et al. SARS-CoV-2 Viral Load in Upper Respiratory Specimens of Infected Patients. N. Engl. J. Med. 2020, 382, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.C.; Hung, E.C.; Woo, J.K.; Chan, P.K.; Leung, S.F.; Lai, F.P.; Cheng, A.S.; Yeung, S.W.; Chan, Y.W.; Tsui, T.K.; et al. Early detection of nasopharyngeal carcinoma by plasma Epstein-Barr virus DNA analysis in a surveillance program. Cancer 2013, 119, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Ka-Yue Chow, L.; Lai-Shun Chung, D.; Tao, L.; Chan, K.F.; Tung, S.Y.; Cheong Ngan, R.K.; Ng, W.T.; Wing-Mui Lee, A.; Yau, C.C.; Lai-Wan Kwong, D.; et al. Epigenomic landscape study reveals molecular subtypes and EBV-associated regulatory epigenome reprogramming in nasopharyngeal carcinoma. EBioMedicine 2022, 86, 104357. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, S.; Cassaniti, I.; Novazzi, F.; Fiorina, L.; Piralla, A.; Comolli, G.; Bruno, R.; Maserati, R.; Gulminetti, R.; Novati, S.; et al. EBV DNA increase in COVID-19 patients with impaired lymphocyte subpopulation count. Int. J. Infect. Dis. 2021, 104, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Guan, W.; Chen, R.; Wang, W.; Li, J.; Xu, K.; Li, C.; Ai, Q.; Lu, W.; Liang, H.; et al. Cancer patients in SARS-CoV-2 infection: A nationwide analysis in China. Lancet Oncol. 2020, 21, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Kadomatsu, K.; Tomomura, M.; Muramatsu, T. cDNA cloning and sequencing of a new gene intensely expressed in early differentiation stages of embryonal carcinoma cells and in mid-gestation period of mouse embryogenesis. Biochem. Biophys. Res. Commun. 1988, 151, 1312–1318. [Google Scholar] [CrossRef]
- Kadomatsu, K.; Huang, R.P.; Suganuma, T.; Murata, F.; Muramatsu, T. A retinoic acid responsive gene MK found in the teratocarcinoma system is expressed in spatially and temporally controlled manner during mouse embryogenesis. J. Cell. Biol. 1990, 110, 607–616. [Google Scholar] [CrossRef]
- Jones, D.R. Measuring midkine: The utility of midkine as a biomarker in cancer and other diseases. Br. J. Pharmacol. 2014, 171, 2925–2939. [Google Scholar] [CrossRef] [PubMed]
- Kemper, M.; Hentschel, W.; Grass, J.K.; Stuben, B.O.; Konczalla, L.; Rawnaq, T.; Ghadban, T.; Izbicki, J.R.; Reeh, M. Serum Midkine is a clinical significant biomarker for colorectal cancer and associated with poor survival. Cancer Med. 2020, 9, 2010–2018. [Google Scholar] [CrossRef] [PubMed]
- Shimada, H.; Nabeya, Y.; Okazumi, S.; Matsubara, H.; Kadomatsu, K.; Muramatsu, T.; Ikematsu, S.; Sakuma, S.; Ochiai, T. Increased serum midkine concentration as a possible tumor marker in patients with superficial esophageal cancer. Oncol. Rep. 2003, 10, 411–414. [Google Scholar] [CrossRef] [PubMed]
- El-Shayeb, A.F.; El-Habachi, N.M.; Mansour, A.R.; Zaghloul, M.S. Serum midkine is a more sensitive predictor for hepatocellular carcinoma than Dickkopf-1 and alpha-L-fucosidase in cirrhotic HCV patients. Medicine 2021, 100, e25112. [Google Scholar] [CrossRef]
- Ketenci, S.; Uygar Kalayci, M.; Dundar, B.; Duranay, R.; Sukru Aynacioglu, A. Elevated serum midkine levels in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infected patients. Int. Immunopharmacol. 2022, 110, 108939. [Google Scholar] [CrossRef] [PubMed]
- Yazihan, N.; Erol, S.A.; Akdas, S.; Goncu Ayhan, S.; Atalay, A.; Yildirim, M.; Biriken, D.; Akin, I.; Altiner, S.; Ceylan, M.N.; et al. Serum midkine level might be a diagnostic tool for COVID19 disease in pregnancy: From the disease severity, hospitalization and disease progression respects. Cytokine 2022, 149, 155751. [Google Scholar] [CrossRef]
- Obama, H.; Tsutsui, J.; Ozawa, M.; Yoshida, H.; Yoshida, Y.; Osame, M.; Muramatsu, T. Midkine (MK) expression in extraembryonic tissues, amniotic fluid, and cerebrospinal fluid during mouse embryogenesis. J. Biochem. 1995, 118, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Tan, Z.; Gu, B.; Wu, R.R.; Liu, Y.K.; Dai, L.C.; Zhang, M. Promotion of self-renewal of embryonic stem cells by midkine. Acta Pharmacol. Sin. 2010, 31, 629–637. [Google Scholar] [CrossRef]
- Nakanishi, T.; Kadomatsu, K.; Okamoto, T.; Ichihara-Tanaka, K.; Kojima, T.; Saito, H.; Tomoda, Y.; Muramatsu, T. Expression of syndecan-1 and -3 during embryogenesis of the central nervous system in relation to binding with midkine. J. Biochem. 1997, 121, 197–205. [Google Scholar]
- Kojima, T.; Katsumi, A.; Yamazaki, T.; Muramatsu, T.; Nagasaka, T.; Ohsumi, K.; Saito, H. Human ryudocan from endothelium-like cells binds basic fibroblast growth factor, midkine, and tissue factor pathway inhibitor. J. Biol. Chem. 1996, 271, 5914–5920. [Google Scholar] [CrossRef]
- Kurosawa, N.; Chen, G.Y.; Kadomatsu, K.; Ikematsu, S.; Sakuma, S.; Muramatsu, T. Glypican-2 binds to midkine: The role of glypican-2 in neuronal cell adhesion and neurite outgrowth. Glycoconj. J. 2001, 18, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Shindo, E.; Nanki, T.; Kusunoki, N.; Shikano, K.; Kawazoe, M.; Sato, H.; Kaneko, K.; Muraoka, S.; Kaburaki, M.; Akasaka, Y.; et al. The growth factor midkine may play a pathophysiological role in rheumatoid arthritis. Mod. Rheumatol. 2017, 27, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Muramatsu, T.; Hirai, M.; Inui, T.; Kimura, T.; Saito, H.; McCormick, L.M.; Bu, G.; Kadomatsu, K. Nuclear targeting by the growth factor midkine. Mol. Cell. Biol. 2002, 22, 6788–6796. [Google Scholar] [CrossRef]
- Stoica, G.E.; Kuo, A.; Powers, C.; Bowden, E.T.; Sale, E.B.; Riegel, A.T.; Wellstein, A. Midkine binds to anaplastic lymphoma kinase (ALK) and acts as a growth factor for different cell types. J. Biol. Chem. 2002, 277, 35990–35998. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hoque, M.O.; Wu, F.; Trink, B.; Sidransky, D.; Ratovitski, E.A. Midkine induces epithelial-mesenchymal transition through Notch2/Jak2-Stat3 signaling in human keratinocytes. Cell Cycle 2008, 7, 1613–1622. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, H.; Zou, P.; Suzuki, H.; Oda, Y.; Chen, G.Y.; Sakaguchi, N.; Sakuma, S.; Maeda, N.; Noda, M.; Takada, Y.; et al. alpha4beta1- and alpha6beta1-integrins are functional receptors for midkine, a heparin-binding growth factor. J. Cell Sci. 2004, 117, 5405–5415. [Google Scholar] [CrossRef] [PubMed]
- Mitsiadis, T.A.; Salmivirta, M.; Muramatsu, T.; Muramatsu, H.; Rauvala, H.; Lehtonen, E.; Jalkanen, M.; Thesleff, I. Expression of the heparin-binding cytokines, midkine (MK) and HB-GAM (pleiotrophin) is associated with epithelial-mesenchymal interactions during fetal development and organogenesis. Development 1995, 121, 37–51. [Google Scholar] [CrossRef]
- Ikutomo, M.; Sakakima, H.; Matsuda, F.; Yoshida, Y. Midkine-deficient mice delayed degeneration and regeneration after skeletal muscle injury. Acta Histochem. 2014, 116, 319–326. [Google Scholar] [CrossRef]
- Horiba, M.; Kadomatsu, K.; Yasui, K.; Lee, J.K.; Takenaka, H.; Sumida, A.; Kamiya, K.; Chen, S.; Sakuma, S.; Muramatsu, T.; et al. Midkine plays a protective role against cardiac ischemia/reperfusion injury through a reduction of apoptotic reaction. Circulation 2006, 114, 1713–1720. [Google Scholar] [CrossRef]
- Takenaka, H.; Horiba, M.; Ishiguro, H.; Sumida, A.; Hojo, M.; Usui, A.; Akita, T.; Sakuma, S.; Ueda, Y.; Kodama, I.; et al. Midkine prevents ventricular remodeling and improves long-term survival after myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H462–H469. [Google Scholar] [CrossRef]
- Guo, X.; Pan, Y.; Xiong, M.; Sanapala, S.; Anastasaki, C.; Cobb, O.; Dahiya, S.; Gutmann, D.H. Midkine activation of CD8(+) T cells establishes a neuron-immune-cancer axis responsible for low-grade glioma growth. Nat. Commun. 2020, 11, 2177. [Google Scholar] [CrossRef] [PubMed]
- Cerezo-Wallis, D.; Contreras-Alcalde, M.; Troule, K.; Catena, X.; Mucientes, C.; Calvo, T.G.; Canon, E.; Tejedo, C.; Pennacchi, P.C.; Hogan, S.; et al. Midkine rewires the melanoma microenvironment toward a tolerogenic and immune-resistant state. Nat. Med. 2020, 26, 1865–1877. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Maeda, K.; Sato, W.; Kosugi, T.; Sato, Y.; Kojima, H.; Kato, N.; Ishimoto, T.; Tsuboi, N.; Uchimura, K.; et al. Growth Factor Midkine Promotes T-Cell Activation through Nuclear Factor of Activated T Cells Signaling and Th1 Cell Differentiation in Lupus Nephritis. Am. J. Pathol. 2017, 187, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, Y.; Li, H.; Jin, S.; Kishida, S.; Kadomatsu, K.; Takeuchi, H.; Mizuno, T.; Suzumura, A. Midkine inhibits inducible regulatory T cell differentiation by suppressing the development of tolerogenic dendritic cells. J. Immunol. 2012, 188, 2602–2611. [Google Scholar] [CrossRef]
- Cohen, S.; Shoshana, O.Y.; Zelman-Toister, E.; Maharshak, N.; Binsky-Ehrenreich, I.; Gordin, M.; Hazan-Halevy, I.; Herishanu, Y.; Shvidel, L.; Haran, M.; et al. The cytokine midkine and its receptor RPTPzeta regulate B cell survival in a pathway induced by CD74. J. Immunol. 2012, 188, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Ichihara-Tanaka, K.; Oohira, A.; Rumsby, M.; Muramatsu, T. Neuroglycan C is a novel midkine receptor involved in process elongation of oligodendroglial precursor-like cells. J. Biol. Chem. 2006, 281, 30857–30864. [Google Scholar] [CrossRef]
- Salama, R.H.; Muramatsu, H.; Zou, K.; Inui, T.; Kimura, T.; Muramatsu, T. Midkine binds to 37-kDa laminin binding protein precursor, leading to nuclear transport of the complex. Exp. Cell Res. 2001, 270, 13–20. [Google Scholar] [CrossRef]
- Huang, Y.; Sook-Kim, M.; Ratovitski, E. Midkine promotes tetraspanin-integrin interaction and induces FAK-Stat1alpha pathway contributing to migration/invasiveness of human head and neck squamous cell carcinoma cells. Biochem. Biophys. Res. Commun. 2008, 377, 474–478. [Google Scholar] [CrossRef]
- Muramatsu, H.; Zou, K.; Sakaguchi, N.; Ikematsu, S.; Sakuma, S.; Muramatsu, T. LDL receptor-related protein as a component of the midkine receptor. Biochem. Biophys. Res. Commun. 2000, 270, 936–941. [Google Scholar] [CrossRef]
- Zou, K.; Muramatsu, H.; Ikematsu, S.; Sakuma, S.; Salama, R.H.; Shinomura, T.; Kimata, K.; Muramatsu, T. A heparin-binding growth factor, midkine, binds to a chondroitin sulfate proteoglycan, PG-M/versican. Eur. J. Biochem. 2000, 267, 4046–4053. [Google Scholar] [CrossRef]
- Sakaguchi, N.; Muramatsu, H.; Ichihara-Tanaka, K.; Maeda, N.; Noda, M.; Yamamoto, T.; Michikawa, M.; Ikematsu, S.; Sakuma, S.; Muramatsu, T. Receptor-type protein tyrosine phosphatase zeta as a component of the signaling receptor complex for midkine-dependent survival of embryonic neurons. Neurosci. Res. 2003, 45, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Weckbach, L.T.; Grabmaier, U.; Uhl, A.; Gess, S.; Boehm, F.; Zehrer, A.; Pick, R.; Salvermoser, M.; Czermak, T.; Pircher, J.; et al. Midkine drives cardiac inflammation by promoting neutrophil trafficking and NETosis in myocarditis. J. Exp. Med. 2019, 216, 350–368. [Google Scholar] [CrossRef] [PubMed]
- Weckbach, L.T.; Gola, A.; Winkelmann, M.; Jakob, S.M.; Groesser, L.; Borgolte, J.; Pogoda, F.; Pick, R.; Pruenster, M.; Muller-Hocker, J.; et al. The cytokine midkine supports neutrophil trafficking during acute inflammation by promoting adhesion via beta2 integrins (CD11/CD18). Blood 2014, 123, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, T.; Yuzawa, Y.; Sato, W.; Arata-Kawai, H.; Suzuki, N.; Kato, N.; Matsuo, S.; Kadomatsu, K. Midkine is involved in tubulointerstitial inflammation associated with diabetic nephropathy. Lab. Investig. 2007, 87, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Duan, C.; Pang, H.; Chen, Q.; Han, B.; Zha, C.; Dinislam, M.; Wu, P.; Li, Z.; Zhao, S.; et al. DNA damage repair alterations modulate M2 polarization of microglia to remodel the tumor microenvironment via the p53-mediated MDK expression in glioma. EBioMedicine 2019, 41, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Callebaut, C.; Nisole, S.; Briand, J.P.; Krust, B.; Hovanessian, A.G. Inhibition of HIV infection by the cytokine midkine. Virology 2001, 281, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Kadomatsu, K.; Hagihara, M.; Akhter, S.; Fan, Q.W.; Muramatsu, H.; Muramatsu, T. Midkine induces the transformation of NIH3T3 cells. Br. J. Cancer 1997, 75, 354–359. [Google Scholar] [CrossRef]
- Shin, D.H.; Jo, J.Y.; Kim, S.H.; Choi, M.; Han, C.; Choi, B.K.; Kim, S.S. Midkine Is a Potential Therapeutic Target of Tumorigenesis, Angiogenesis, and Metastasis in Non-Small Cell Lung Cancer. Cancers 2020, 12, 2402. [Google Scholar] [CrossRef]
- Masui, M.; Okui, T.; Shimo, T.; Takabatake, K.; Fukazawa, T.; Matsumoto, K.; Kurio, N.; Ibaragi, S.; Naomoto, Y.; Nagatsuka, H.; et al. Novel Midkine Inhibitor iMDK Inhibits Tumor Growth and Angiogenesis in Oral Squamous Cell Carcinoma. Anticancer Res. 2016, 36, 2775–2781. [Google Scholar]
- Xia, T.; Chen, D.; Liu, X.; Qi, H.; Wang, W.; Chen, H.; Ling, T.; Otkur, W.; Zhang, C.S.; Kim, J.; et al. Midkine noncanonically suppresses AMPK activation through disrupting the LKB1-STRAD-Mo25 complex. Cell Death Dis. 2022, 13, 414. [Google Scholar] [CrossRef]
- Kishida, S.; Mu, P.; Miyakawa, S.; Fujiwara, M.; Abe, T.; Sakamoto, K.; Onishi, A.; Nakamura, Y.; Kadomatsu, K. Midkine promotes neuroblastoma through Notch2 signaling. Cancer Res. 2013, 73, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, R.; Zhang, H.T.; Donnini, S.; Ziche, M.; Bicknell, R. An angiogenic role for the neurokines midkine and pleiotrophin in tumorigenesis. Cancer Res. 1997, 57, 1814–1819. [Google Scholar] [PubMed]
- Huang, H.; Li, J.; Lu, Y.; Min, L.; Li, D.; Dai, L. Role of midkine-progranulin interaction during angiogenesis of hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 8809–8820. [Google Scholar] [PubMed]
- Ratovitski, E.A.; Kotzbauer, P.T.; Milbrandt, J.; Lowenstein, C.J.; Burrow, C.R. Midkine induces tumor cell proliferation and binds to a high affinity signaling receptor associated with JAK tyrosine kinases. J. Biol. Chem. 1998, 273, 3654–3660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, L.; Xu, Y.; Xu, Y.; Zheng, M.; Zhang, P.; Wang, Q. Midkine promotes breast cancer cell proliferation and migration by upregulating NR3C1 expression and activating the NF-kappaB pathway. Mol. Biol. Rep. 2022, 49, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Ohuchida, T.; Okamoto, K.; Akahane, K.; Higure, A.; Todoroki, H.; Abe, Y.; Kikuchi, M.; Ikematsu, S.; Muramatsu, T.; Itoh, H. Midkine protects hepatocellular carcinoma cells against TRAIL-mediated apoptosis through down-regulation of caspase-3 activity. Cancer 2004, 100, 2430–2436. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Dong, Y.; Kong, X.; Beckett, L.A.; Gandour-Edwards, R.; Melamed, J. Midkine is a NF-kappaB-inducible gene that supports prostate cancer cell survival. BMC Med. Genom. 2008, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Kwiatkowski, D.J.; Henske, E.P. Midkine expression by stem-like tumor cells drives persistence to mTOR inhibition and an immune-suppressive microenvironment. Nat. Commun. 2022, 13, 5018. [Google Scholar] [CrossRef]
- Zhao, S.; Zhao, G.; Xie, H.; Huang, Y.; Hou, Y. A conjugate of an anti-midkine single-chain variable fragment to doxorubicin inhibits tumor growth. Braz. J. Med. Biol. Res. 2012, 45, 230–237. [Google Scholar] [CrossRef]
- Ding, L.; Wang, N.; Wang, Q.; Fan, X.; Xin, Y.; Wang, S. Midkine inhibition enhances anti-PD-1 immunotherapy in sorafenib-treated hepatocellular carcinoma via preventing immunosuppressive MDSCs infiltration. Cell Death Discov. 2023, 9, 92. [Google Scholar] [CrossRef]
- Shiratori, F.; Ito, M.; Yajima, S.; Suzuki, T.; Oshima, Y.; Nanami, T.; Funahashi, K.; Shimada, H. The effectiveness of serum midkine in detecting esophageal squamous cell carcinoma. Esophagus 2019, 16, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ota, K.; Fujimori, H.; Ueda, M.; Shiniriki, S.; Kudo, M.; Jono, H.; Fukuyoshi, Y.; Yamamoto, Y.; Sugiuchi, H.; Iwase, H.; et al. Midkine as a prognostic biomarker in oral squamous cell carcinoma. Br. J. Cancer 2008, 99, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Zekri, N.A.R.; El Kassas, M.; Salam, E.S.E.; Hassan, R.M.; Mohanad, M.; Gabr, R.M.; Lotfy, M.M.; Abdel-Zaher, R.A.T.; Bahnassy, A.A.; Ahmed, O.S. The possible role of Dickkopf-1, Golgi protein- 73 and Midkine as predictors of hepatocarcinogenesis: A review and an Egyptian study. Sci. Rep. 2020, 10, 5156. [Google Scholar] [CrossRef] [PubMed]
- Cetin Sorkun, H.; Akbulut, M.; Enli, Y.; Tepeli, E.; Ozkan, S.; Erdem, E. Quantitative comparison of immunohistochemical and PCR analysis of midkine expression in breast cancer types and serum midkine level. Turk. J. Med. Sci. 2016, 46, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Rawnaq, T.; Dietrich, L.; Wolters-Eisfeld, G.; Uzunoglu, F.G.; Vashist, Y.K.; Bachmann, K.; Simon, R.; Izbicki, J.R.; Bockhorn, M.; Gungor, C. The multifunctional growth factor midkine promotes proliferation and migration in pancreatic cancer. Mol. Cancer Res. 2014, 12, 670–680. [Google Scholar] [CrossRef]
- Ikematsu, S.; Yano, A.; Aridome, K.; Kikuchi, M.; Kumai, H.; Nagano, H.; Okamoto, K.; Oda, M.; Sakuma, S.; Aikou, T.; et al. Serum midkine levels are increased in patients with various types of carcinomas. Br. J. Cancer 2000, 83, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Stern, L.; Mueller, E.; Bellon, E.; Reeh, M.; Grotelueschen, R.; Guengoer, C.; Melling, N.; Goetz, M.; Perez, D.R.; Izbicki, J.R.; et al. Serum midkine as non-invasive biomarker for detection and prognosis of non-small cell lung cancer. Sci. Rep. 2021, 11, 14616. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Hu, C.; Yang, Z.; Zhang, X.; Zhao, L.; Xiong, J.; Ma, J.; Chen, L.; Qian, H.; Luo, X.; et al. Midkine promotes hepatocellular carcinoma metastasis by elevating anoikis resistance of circulating tumor cells. Oncotarget 2017, 8, 32523–32535. [Google Scholar] [CrossRef]
- Bie, C.; Chen, Y.; Tang, H.; Li, Q.; Zhong, L.; Peng, X.; Shi, Y.; Lin, J.; Lai, J.; Wu, S.; et al. Insulin-Like Growth Factor 1 Receptor Drives Hepatocellular Carcinoma Growth and Invasion by Activating Stat3-Midkine-Stat3 Loop. Dig. Dis. Sci. 2022, 67, 569–584. [Google Scholar] [CrossRef]
- Mirkin, B.L.; Clark, S.; Zheng, X.; Chu, F.; White, B.D.; Greene, M.; Rebbaa, A. Identification of midkine as a mediator for intercellular transfer of drug resistance. Oncogene 2005, 24, 4965–4974. [Google Scholar] [CrossRef]
- Erdogan, S.; Turkekul, K.; Dibirdik, I.; Doganlar, O.; Doganlar, Z.B.; Bilir, A.; Oktem, G. Midkine downregulation increases the efficacy of quercetin on prostate cancer stem cell survival and migration through PI3K/AKT and MAPK/ERK pathway. Biomed. Pharmacother. 2018, 107, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef] [PubMed]
- Hayward, S.; Gachehiladze, M.; Badr, N.; Andrijes, R.; Molostvov, G.; Paniushkina, L.; Sopikova, B.; Slobodova, Z.; Mgebrishvili, G.; Sharma, N.; et al. The CD151-midkine pathway regulates the immune microenvironment in inflammatory breast cancer. J. Pathol. 2020, 251, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Kam, N.W.; Laczka, O.; Li, X.; Wilkinson, J.; Hung, D.; Lai, S.P.H.; Wu, K.C.; Tsao, S.W.; Dai, W.; Che, C.M.; et al. ENOX2 inhibition enhances infiltration of effector memory T-cell and mediates response to chemotherapy in immune-quiescent nasopharyngeal carcinoma. J. Adv. Res. 2023. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Allam, M.; Hu, T.; Lee, J.; Aldrich, J.; Badve, S.S.; Gokmen-Polar, Y.; Bhave, M.; Ramalingam, S.S.; Schneider, F.; Coskun, A.F. Spatially variant immune infiltration scoring in human cancer tissues. NPJ Precis. Oncol. 2022, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Sun, Z.J. Turning cold tumors into hot tumors by improving T-cell infiltration. Theranostics 2021, 11, 5365–5386. [Google Scholar] [CrossRef] [PubMed]
- Vilain, R.E.; Menzies, A.M.; Wilmott, J.S.; Kakavand, H.; Madore, J.; Guminski, A.; Liniker, E.; Kong, B.Y.; Cooper, A.J.; Howle, J.R.; et al. Dynamic Changes in PD-L1 Expression and Immune Infiltrates Early During Treatment Predict Response to PD-1 Blockade in Melanoma. Clin. Cancer Res. 2017, 23, 5024–5033. [Google Scholar] [CrossRef]
- Lopez de Rodas, M.; Nagineni, V.; Ravi, A.; Datar, I.J.; Mino-Kenudson, M.; Corredor, G.; Barrera, C.; Behlman, L.; Rimm, D.L.; Herbst, R.S.; et al. Role of tumor infiltrating lymphocytes and spatial immune heterogeneity in sensitivity to PD-1 axis blockers in non-small cell lung cancer. J. Immunother. Cancer 2022, 10, e004440. [Google Scholar] [CrossRef]
- Miller, B.C.; Sen, D.R.; Al Abosy, R.; Bi, K.; Virkud, Y.V.; LaFleur, M.W.; Yates, K.B.; Lako, A.; Felt, K.; Naik, G.S.; et al. Subsets of exhausted CD8(+) T cells differentially mediate tumor control and respond to checkpoint blockade. Nat. Immunol. 2019, 20, 326–336. [Google Scholar] [CrossRef]
- Siddiqui, I.; Schaeuble, K.; Chennupati, V.; Fuertes Marraco, S.A.; Calderon-Copete, S.; Pais Ferreira, D.; Carmona, S.J.; Scarpellino, L.; Gfeller, D.; Pradervand, S.; et al. Intratumoral Tcf1(+)PD-1(+)CD8(+) T Cells with Stem-like Properties Promote Tumor Control in Response to Vaccination and Checkpoint Blockade Immunotherapy. Immunity 2019, 50, 195–211. [Google Scholar] [CrossRef]
- Vinogradov, S.; Warren, G.; Wei, X. Macrophages associated with tumors as potential targets and therapeutic intermediates. Nanomedicine 2014, 9, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, R.; Su, X.; Pan, Y.; Han, X.; Shao, C.; Shi, Y. Harnessing tumor-associated macrophages as aids for cancer immunotherapy. Mol. Cancer 2019, 18, 177. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Zhao, Y.L.; Peng, L.S.; Chen, N.; Chen, W.; Lv, Y.P.; Mao, F.Y.; Zhang, J.Y.; Cheng, P.; Teng, Y.S.; et al. Tumour-activated neutrophils in gastric cancer foster immune suppression and disease progression through GM-CSF-PD-L1 pathway. Gut 2017, 66, 1900–1911. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, J.; Shaul, M.E.; Mishalian, I.; Hovav, A.H.; Levy, L.; Zolotriov, L.; Granot, Z.; Fridlender, Z.G. Tumor-associated neutrophils induce apoptosis of non-activated CD8 T-cells in a TNFalpha and NO-dependent mechanism, promoting a tumor-supportive environment. Oncoimmunology 2017, 6, e1356965. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Schrum, A.G.; Cho, H.I.; Celis, E.; Gabrilovich, D.I. Mechanism of T cell tolerance induced by myeloid-derived suppressor cells. J. Immunol. 2010, 184, 3106–3116. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.Y.; Ma, G.; Weber, K.J.; Ozao-Choy, J.; Wang, G.; Yin, B.; Divino, C.M.; Chen, S.H. Immune stimulatory receptor CD40 is required for T-cell suppression and T regulatory cell activation mediated by myeloid-derived suppressor cells in cancer. Cancer Res. 2010, 70, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression–implications for anticancer therapy. Nat. Rev. Clin. Oncol. 2019, 16, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Thornton, A.M.; Shevach, E.M. CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J. Exp. Med. 1998, 188, 287–296. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Yang, L.; Liu, G.; Li, Y.; Pan, Y. The emergence of tumor-infiltrating lymphocytes in nasopharyngeal carcinoma: Predictive value and immunotherapy implications. Genes Dis. 2022, 9, 1208–1219. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Chen, L.; Mao, Y.P.; Li, Y.Q.; Jiang, W.; Xu, S.Y.; Zhang, Y.; Chen, Y.P.; Li, X.M.; He, Q.M.; et al. Prognostic value of immune score in nasopharyngeal carcinoma using digital pathology. J. Immunother. Cancer 2020, 8, e000334. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, G.; Zhang, C.; Liao, X.; Xie, J.; Liang, T.; Liao, W.; Song, L.; Zhang, X. Prognostic significance of tumor-infiltrating lymphocytes and macrophages in nasopharyngeal carcinoma: A systematic review and meta-analysis. Eur. Arch. Otorhinolaryngol. 2022, 279, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Ooft, M.L.; van Ipenburg, J.A.; Braunius, W.W.; Zuur, C.I.; Koljenovic, S.; Willems, S.M. Prognostic role of tumor infiltrating lymphocytes in EBV positive and EBV negative nasopharyngeal carcinoma. Oral Oncol. 2017, 71, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L. Prognostic significance of tumor-associated macrophages in patients with nasopharyngeal carcinoma: A meta-analysis. Medicine 2020, 99, e21999. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.T.; Ye, S.B.; Liu, Y.N.; He, J.; Chen, Q.Y.; Mai, H.Q.; Zhang, C.X.; Cui, J.; Zhang, X.S.; Busson, P.; et al. LMP1-mediated glycolysis induces myeloid-derived suppressor cell expansion in nasopharyngeal carcinoma. PLoS Pathog. 2017, 13, e1006503. [Google Scholar] [CrossRef] [PubMed]
- Yarza, R.; Bover, M.; Agullo-Ortuno, M.T.; Iglesias-Docampo, L.C. Current approach and novel perspectives in nasopharyngeal carcinoma: The role of targeting proteasome dysregulation as a molecular landmark in nasopharyngeal cancer. J. Exp. Clin. Cancer Res. 2021, 40, 202. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.A.; Vickers, M.A.; Barker, R.N. Regulatory T cells secreting IL-10 dominate the immune response to EBV latent membrane protein 1. J. Immunol. 2003, 170, 6183–6189. [Google Scholar] [CrossRef]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein-Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160270. [Google Scholar] [CrossRef]
- Wang, J.; Huang, H.; Lu, J.; Bi, P.; Wang, F.; Liu, X.; Zhang, B.; Luo, Y.; Li, X. Tumor cells induced-M2 macrophage favors accumulation of Treg in nasopharyngeal carcinoma. Int. J. Clin. Exp. Pathol. 2017, 10, 8389–8401. [Google Scholar]
- Ooft, M.L.; van Ipenburg, J.A.; Sanders, M.E.; Kranendonk, M.; Hofland, I.; de Bree, R.; Koljenovic, S.; Willems, S.M. Prognostic role of tumour-associated macrophages and regulatory T cells in EBV-positive and EBV-negative nasopharyngeal carcinoma. J. Clin. Pathol. 2018, 71, 267–274. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Q.H.; Wang, F.; Tan, J.J.; Deng, Y.Q.; Peng, X.H.; Liu, X.; Zhang, B.; Xu, X.; Li, X.P. Exosomal miR-9 inhibits angiogenesis by targeting MDK and regulating PDK/AKT pathway in nasopharyngeal carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 147. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Mao, X.; Zhang, S.; Xie, H.; Yan, B.; Wang, B.; Sun, J.; Wei, L. HPV + HNSCC-derived exosomal miR-9 induces macrophage M1 polarization and increases tumor radiosensitivity. Cancer Lett. 2020, 478, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Agaoglu, F.Y.; Dizdar, Y.; Dogan, O.; Alatli, C.; Ayan, I.; Savci, N.; Tas, S.; Dalay, N.; Altun, M. P53 overexpression in nasopharyngeal carcinoma. In Vivo 2004, 18, 555–560. [Google Scholar] [PubMed]
- Fang, W.; Zhang, J.; Hong, S.; Zhan, J.; Chen, N.; Qin, T.; Tang, Y.; Zhang, Y.; Kang, S.; Zhou, T.; et al. EBV-driven LMP1 and IFN-gamma up-regulate PD-L1 in nasopharyngeal carcinoma: Implications for oncotargeted therapy. Oncotarget 2014, 5, 12189–12202. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Liu, Q.; Li, R.; Chen, S.; Tan, J.; Li, L.; Dong, X.; Huang, C.; Wen, T.; Liu, J. Targeting MDK Abrogates IFN-gamma-Elicited Metastasis inCancers of Various Origins. Front. Oncol. 2022, 12, 885656. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Xie, L.; Ge, J.; Li, H.; Zhong, S.; Liu, X. Integrating single-cell RNA-seq and spatial transcriptomics reveals MDK-NCL dependent immunosuppressive environment in endometrial carcinoma. Front. Immunol. 2023, 14, 1145300. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Q.; Yang, S.; Lu, H.S. Expression of midkine and vascular endothelial growth factor in gastric cancer and the association of high levels with poor prognosis and survival. Mol. Med. Rep. 2012, 5, 415–419. [Google Scholar] [CrossRef]
- Luo, M.S.; Huang, G.J.; Liu, B.X. Immune infiltration in nasopharyngeal carcinoma based on gene expression. Medicine 2019, 98, e17311. [Google Scholar] [CrossRef]
- Lucas, M.; Karrer, U.; Lucas, A.; Klenerman, P. Viral escape mechanisms--escapology taught by viruses. Int. J. Exp. Pathol. 2001, 82, 269–286. [Google Scholar] [CrossRef]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Taefehshokr, N.; Taefehshokr, S.; Hemmat, N.; Heit, B. Covid-19: Perspectives on Innate Immune Evasion. Front. Immunol. 2020, 11, 580641. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Liu, W.; Li, X.; Zhao, P.; Shereen, M.A.; Zhu, C.; Huang, S.; Liu, S.; Yu, X.; Yue, M.; et al. HIF-1alpha promotes SARS-CoV-2 infection and aggravates inflammatory responses to COVID-19. Signal Transduct. Target. Ther. 2021, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, P.R.; Mucenski, M.L.; Le Cras, T.D.; Nichols, W.C.; Whitsett, J.A. Midkine is regulated by hypoxia and causes pulmonary vascular remodeling. J. Biol. Chem. 2004, 279, 37124–37132. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pan, Y.; Fanelli, V.; Wu, S.; Luo, A.A.; Islam, D.; Han, B.; Mao, P.; Ghazarian, M.; Zeng, W.; et al. Mechanical Stress and the Induction of Lung Fibrosis via the Midkine Signaling Pathway. Am. J. Respir. Crit. Care Med. 2015, 192, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Misa, K.; Tanino, Y.; Wang, X.; Nikaido, T.; Kikuchi, M.; Sato, Y.; Togawa, R.; Tanino, M.; Tanaka, S.; Kadomatsu, K.; et al. Involvement of midkine in the development of pulmonary fibrosis. Physiol. Rep. 2017, 5, e13383. [Google Scholar] [CrossRef]
- Pan, Y.; Jiang, X.; Yang, L.; Chen, L.; Zeng, X.; Liu, G.; Tang, Y.; Qian, C.; Wang, X.; Cheng, F.; et al. SARS-CoV-2-specific immune response in COVID-19 convalescent individuals. Signal Transduct. Target. Ther. 2021, 6, 256. [Google Scholar] [CrossRef]
- Gubernatorova, E.O.; Gorshkova, E.A.; Polinova, A.I.; Drutskaya, M.S. IL-6: Relevance for immunopathology of SARS-CoV-2. Cytokine Growth Factor Rev. 2020, 53, 13–24. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045. [Google Scholar] [CrossRef]
- Schultheiss, C.; Willscher, E.; Paschold, L.; Gottschick, C.; Klee, B.; Henkes, S.S.; Bosurgi, L.; Dutzmann, J.; Sedding, D.; Frese, T.; et al. The IL-1beta, IL-6, and TNF cytokine triad is associated with post-acute sequelae of COVID-19. Cell Rep. Med. 2022, 3, 100663. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Bost, P.; Giladi, A.; Liu, Y.; Bendjelal, Y.; Xu, G.; David, E.; Blecher-Gonen, R.; Cohen, M.; Medaglia, C.; Li, H.; et al. Host-Viral Infection Maps Reveal Signatures of Severe COVID-19 Patients. Cell 2020, 181, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetite, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef] [PubMed]
- Leppkes, M.; Knopf, J.; Naschberger, E.; Lindemann, A.; Singh, J.; Herrmann, I.; Sturzl, M.; Staats, L.; Mahajan, A.; Schauer, C.; et al. Vascular occlusion by neutrophil extracellular traps in COVID-19. EBioMedicine 2020, 58, 102925. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Ren, X.; Li, Y.; Li, H. Midkine promotes kidney injury in diabetic kidney disease by increasing neutrophil extracellular traps formation. Ann. Transl. Med. 2022, 10, 693. [Google Scholar] [CrossRef] [PubMed]
- Narita, H.; Chen, S.; Komori, K.; Kadomatsu, K. Midkine is expressed by infiltrating macrophages in in-stent restenosis in hypercholesterolemic rabbits. J. Vasc. Surg. 2008, 47, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Lian, Q.; Zhang, K.; Zhang, Z.; Duan, F.; Guo, L.; Luo, W.; Mok, B.W.; Thakur, A.; Ke, X.; Motallebnejad, P.; et al. Differential effects of macrophage subtypes on SARS-CoV-2 infection in a human pluripotent stem cell-derived model. Nat. Commun. 2022, 13, 2028. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L.; et al. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- De Zuani, M.; Laznickova, P.; Tomaskova, V.; Dvoncova, M.; Forte, G.; Stokin, G.B.; Sramek, V.; Helan, M.; Fric, J. High CD4-to-CD8 ratio identifies an at-risk population susceptible to lethal COVID-19. Scand. J. Immunol. 2022, 95, e13125. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lubke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Schmitz, K.S.; Raadsen, M.P.; Grifoni, A.; Okba, N.M.A.; Endeman, H.; van den Akker, J.P.C.; Molenkamp, R.; Koopmans, M.P.G.; van Gorp, E.C.M.; et al. Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Sci. Immunol. 2020, 5, eabd2071. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168. [Google Scholar] [CrossRef]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C.; et al. Broad and strong memory CD4(+) and CD8(+) T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Lai, X.; Sun, J.; Chen, Z.; Zhang, Z.; Dai, J.; Liu, D.; Li, Y.; Li, F.; Wang, Y.; et al. Mapping and role of T cell response in SARS-CoV-2-infected mice. J. Exp. Med. 2021, 218, e20202187. [Google Scholar] [CrossRef] [PubMed]
- McMahan, K.; Yu, J.; Mercado, N.B.; Loos, C.; Tostanoski, L.H.; Chandrashekar, A.; Liu, J.; Peter, L.; Atyeo, C.; Zhu, A.; et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021, 590, 630–634. [Google Scholar] [CrossRef]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients With Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Zhang, M.; Yang, C.X.; Zhang, N.; Wang, X.C.; Yang, X.P.; Dong, X.Q.; Zheng, Y.T. Elevated exhaustion levels and reduced functional diversity of T cells in peripheral blood may predict severe progression in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 541–543. [Google Scholar] [CrossRef]
- Xu, G.; Qi, F.; Li, H.; Yang, Q.; Wang, H.; Wang, X.; Liu, X.; Zhao, J.; Liao, X.; Liu, Y.; et al. The differential immune responses to COVID-19 in peripheral and lung revealed by single-cell RNA sequencing. Cell Discov. 2020, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; John Wherry, E. T cell responses in patients with COVID-19. Nat. Rev. Immunol. 2020, 20, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Kilian, C.; Turner, J.E.; Bosurgi, L.; Roedl, K.; Bartsch, P.; Gnirck, A.C.; Cortesi, F.; Schultheiss, C.; Hellmig, M.; et al. Clonal expansion and activation of tissue-resident memory-like Th17 cells expressing GM-CSF in the lungs of severe COVID-19 patients. Sci. Immunol. 2021, 6, eabf6692. [Google Scholar] [CrossRef]
- Vanderbeke, L.; Van Mol, P.; Van Herck, Y.; De Smet, F.; Humblet-Baron, S.; Martinod, K.; Antoranz, A.; Arijs, I.; Boeckx, B.; Bosisio, F.M.; et al. Monocyte-driven atypical cytokine storm and aberrant neutrophil activation as key mediators of COVID-19 disease severity. Nat. Commun. 2021, 12, 4117. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.; Liu, J.; Zhang, Z.; Wan, X.; Huang, B.; Chen, Y.; Zhang, Y. COVID-19: Immunopathogenesis and Immunotherapeutics. Signal Transduct. Target. Ther. 2020, 5, 128. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]
- Du, R.H.; Liang, L.R.; Yang, C.Q.; Wang, W.; Cao, T.Z.; Li, M.; Guo, G.Y.; Du, J.; Zheng, C.L.; Zhu, Q.; et al. Predictors of mortality for patients with COVID-19 pneumonia caused by SARS-CoV-2: A prospective cohort study. Eur. Respir. J. 2020, 55. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Sato, W.; Kadomatsu, K.; Yuzawa, Y.; Muramatsu, H.; Hotta, N.; Matsuo, S.; Muramatsu, T. Midkine is involved in neutrophil infiltration into the tubulointerstitium in ischemic renal injury. J. Immunol. 2001, 167, 3463–3469. [Google Scholar] [CrossRef]
- Zhai, M.; Zhang, S. A Nasopharyngeal Carcinoma Patient With COVID-19 Infection After Immunotherapy: A Case Report and Literature Review. In Vivo 2020, 34, 3753–3756. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Yi, F.; Ren, X.; Li, Z.; Wang, D.; Wang, N. Case Report: A Nasopharyngeal Cancer Patient Got COVID-19 During Radiochemotherapy in Wuhan. Front. Oncol. 2020, 10, 1755. [Google Scholar] [CrossRef] [PubMed]
- Kadriyan, H.; Kurniawan, Y.; Indriyani, S.A.K.; Pieter, N.L.A.; Gaffar, M.; Punagi, A.Q. COVID-19 infection in a teenage patient with nasopharyngeal cancer. In Advances in Health Sciences Research; Atlantis Press International B.V.: Dordrecht, The Netherlands, 2022; pp. 78–83. [Google Scholar]
- Venkatraman, P.; Sahay, J.J.; Maidili, T.; Rajan, R.; Pooja, S. Breakthrough of COVID-19 using radiotherapy treatment modalities. Radiother. Oncol. 2020, 148, 225–226. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.J.; Liu, Y.L.; Wen, K.; Kurts, C.; Wu, H.; Mei, Q.; Li, J. Potentially improved response of COVID-19 vaccinated nasopharyngeal cancer patients to combination therapy with anti-PD-1 blockade and chemotherapy. Ann. Oncol. 2023, 34, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.; Cazier, J.B.; Angelis, V.; Arnold, R.; Bisht, V.; Campton, N.A.; Chackathayil, J.; Cheng, V.W.; Curley, H.M.; Fittall, M.W.; et al. COVID-19 mortality in patients with cancer on chemotherapy or other anticancer treatments: A prospective cohort study. Lancet 2020, 395, 1919–1926. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, C.; Cai, X.; Xie, Z.; Zhou, L.; Cheng, B.; Zhong, R.; Xiong, S.; Li, J.; Chen, Z.; et al. The association between CD8+ tumor-infiltrating lymphocytes and the clinical outcome of cancer immunotherapy: A systematic review and meta-analysis. EClinicalMedicine 2021, 41, 101134. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Perica, K.; Klebanoff, C.A.; Wolchok, J.D. Clinical implications of T cell exhaustion for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2022, 19, 775–790. [Google Scholar] [CrossRef]
- Urra, J.M.; Cabrera, C.M.; Porras, L.; Rodenas, I. Selective CD8 cell reduction by SARS-CoV-2 is associated with a worse prognosis and systemic inflammation in COVID-19 patients. Clin. Immunol. 2020, 217, 108486. [Google Scholar] [CrossRef]
- Hu, R.; Yan, Y.; Li, Q.; Lin, Y.; Jin, W.; Li, H.; Lu, Y.; Pang, T. Increased drug efflux along with midkine gene high expression in childhood B-lineage acute lymphoblastic leukemia cells. Int. J. Hematol. 2010, 92, 105–110. [Google Scholar] [CrossRef]
- Gungor, C.; Zander, H.; Effenberger, K.E.; Vashist, Y.K.; Kalinina, T.; Izbicki, J.R.; Yekebas, E.; Bockhorn, M. Notch signaling activated by replication stress-induced expression of midkine drives epithelial-mesenchymal transition and chemoresistance in pancreatic cancer. Cancer Res. 2011, 71, 5009–5019. [Google Scholar] [CrossRef]
- Garassino, M.C.; Whisenant, J.G.; Huang, L.C.; Trama, A.; Torri, V.; Agustoni, F.; Baena, J.; Banna, G.; Berardi, R.; Bettini, A.C.; et al. COVID-19 in patients with thoracic malignancies (TERAVOLT): First results of an international, registry-based, cohort study. Lancet Oncol. 2020, 21, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Giannakoulis, V.G.; Papoutsi, E.; Siempos, I.I. Effect of Cancer on Clinical Outcomes of Patients With COVID-19: A Meta-Analysis of Patient Data. JCO Glob. Oncol. 2020, 6, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Kuderer, N.M.; Choueiri, T.K.; Shah, D.P.; Shyr, Y.; Rubinstein, S.M.; Rivera, D.R.; Shete, S.; Hsu, C.Y.; Desai, A.; de Lima Lopes, G., Jr.; et al. Clinical impact of COVID-19 on patients with cancer (CCC19): A cohort study. Lancet 2020, 395, 1907–1918. [Google Scholar] [CrossRef] [PubMed]
- Pinato, D.J.; Lee, A.J.X.; Biello, F.; Segui, E.; Aguilar-Company, J.; Carbo, A.; Bruna, R.; Bower, M.; Rizzo, G.; Benafif, S.; et al. Presenting Features and Early Mortality from SARS-CoV-2 Infection in Cancer Patients during the Initial Stage of the COVID-19 Pandemic in Europe. Cancers 2020, 12, 1841. [Google Scholar] [CrossRef] [PubMed]
- Rivera, D.R.; Peters, S.; Panagiotou, O.A.; Shah, D.P.; Kuderer, N.M.; Hsu, C.Y.; Rubinstein, S.M.; Lee, B.J.; Choueiri, T.K.; de Lima Lopes, G., Jr.; et al. Utilization of COVID-19 Treatments and Clinical Outcomes among Patients with Cancer: A COVID-19 and Cancer Consortium (CCC19) Cohort Study. Cancer Discov. 2020, 10, 1514–1527. [Google Scholar] [CrossRef]
- Saini, K.S.; Tagliamento, M.; Lambertini, M.; McNally, R.; Romano, M.; Leone, M.; Curigliano, G.; de Azambuja, E. Mortality in patients with cancer and coronavirus disease 2019: A systematic review and pooled analysis of 52 studies. Eur. J. Cancer 2020, 139, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Westblade, L.F.; Brar, G.; Pinheiro, L.C.; Paidoussis, D.; Rajan, M.; Martin, P.; Goyal, P.; Sepulveda, J.L.; Zhang, L.; George, G.; et al. SARS-CoV-2 Viral Load Predicts Mortality in Patients with and without Cancer Who Are Hospitalized with COVID-19. Cancer Cell 2020, 38, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.; Vitetta, L. Adjunctive Treatments for the Prevention of Chemotherapy- and Radiotherapy-Induced Mucositis. Integr. Cancer Ther. 2018, 17, 1027–1047. [Google Scholar] [CrossRef]
- Muramatsu, T. Midkine: A promising molecule for drug development to treat diseases of the central nervous system. Curr. Pharm. Des. 2011, 17, 410–423. [Google Scholar] [CrossRef]
- Takei, Y.; Kadomatsu, K.; Goto, T.; Muramatsu, T. Combinational antitumor effect of siRNA against midkine and paclitaxel on growth of human prostate cancer xenografts. Cancer 2006, 107, 864–873. [Google Scholar] [CrossRef]
- Tian, W.; Shen, J.; Chen, W. Suppression of midkine gene promotes the antitumoral effect of cisplatin on human gastric cancer cell line AGS in vitro and in vivo via the modulation of Notch signaling pathway. Oncol. Rep. 2017, 38, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Fan, Y.; Chen, B.; Hu, Y.; Gao, Y.; Wei, D. Suppression of metastasis of human pancreatic cancer cells to the liver by small interfering RNA-mediated targeting of the midkine gene. Oncol. Lett. 2013, 6, 1338–1342. [Google Scholar] [CrossRef]
- Takei, Y.; Kadomatsu, K.; Matsuo, S.; Itoh, H.; Nakazawa, K.; Kubota, S.; Muramatsu, T. Antisense oligodeoxynucleotide targeted to Midkine, a heparin-binding growth factor, suppresses tumorigenicity of mouse rectal carcinoma cells. Cancer Res. 2001, 61, 8486–8491. [Google Scholar]
- Takei, Y.; Kadomatsu, K.; Itoh, H.; Sato, W.; Nakazawa, K.; Kubota, S.; Muramatsu, T. 5’-,3’-inverted thymidine-modified antisense oligodeoxynucleotide targeting midkine. Its design and application for cancer therapy. J. Biol. Chem. 2002, 277, 23800–23806. [Google Scholar] [CrossRef] [PubMed]
- Kerzerho, J.; Schneider, A.; Favry, E.; Castelli, F.A.; Maillere, B. The signal peptide of the tumor-shared antigen midkine hosts CD4+ T cell epitopes. J. Biol. Chem. 2013, 288, 13370–13377. [Google Scholar] [CrossRef]
- Kerzerho, J.; Adotevi, O.; Castelli, F.A.; Dosset, M.; Bernardeau, K.; Szely, N.; Lang, F.; Tartour, E.; Maillere, B. The angiogenic growth factor and biomarker midkine is a tumor-shared antigen. J. Immunol. 2010, 185, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Terao, S.; Shirakawa, T.; Kubo, S.; Bishunu, A.; Lee, S.J.; Goda, K.; Tsukuda, M.; Hamada, K.; Tagawa, M.; Takenaka, A.; et al. Midkine promoter-based conditionally replicative adenovirus for targeting midkine-expressing human bladder cancer model. Urology 2007, 70, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Reynolds, P.N.; Yamamoto, M.; Wang, M.; Takayama, K.; Matsubara, S.; Muramatsu, T.; Curiel, D.T. A midkine promoter-based conditionally replicative adenovirus for treatment of pediatric solid tumors and bone marrow tumor purging. Cancer Res. 2001, 61, 7882–7888. [Google Scholar]
- Toyoda, E.; Doi, R.; Kami, K.; Mori, T.; Ito, D.; Koizumi, M.; Kida, A.; Nagai, K.; Ito, T.; Masui, T.; et al. Midkine promoter-based conditionally replicative adenovirus therapy for midkine-expressing human pancreatic cancer. J. Exp. Clin. Cancer Res. 2008, 27, 30. [Google Scholar] [CrossRef]
- Kohno, S.; Nakagawa, K.; Hamada, K.; Harada, H.; Yamasaki, K.; Hashimoto, K.; Tagawa, M.; Nagato, S.; Furukawa, K.; Ohnishi, T. Midkine promoter-based conditionally replicative adenovirus for malignant glioma therapy. Oncol. Rep. 2004, 12, 73–78. [Google Scholar] [CrossRef]
- Uehara, N.; Otsuki, N.; Kubo, M.; Kitamoto, J.; Kojima, Y.; Teshima, M.; Shinomiya, H.; Shirakawa, T.; Nibu, K.I. Oncolytic effect of Midkine promoter-based conditionally replicating adenoviruses expressing EGFR siRNA in head and neck squamous cancer cell line T891. Cancer Rep. 2020, 3, e1231. [Google Scholar] [CrossRef]
- Roquefort Therapeutics PLC. Available online: https://www.roquefortplc.com/portfolio (accessed on 9 June 2023).





| Tumor Biological Roles | Indications | Signaling and Mechanisms | Responses Involved | Ref. |
|---|---|---|---|---|
| Immune suppression | Melanoma | NF-κB signaling activation, immunomodulator secretion | Educating macrophages towards immune tolerant states. Inducing resistance towards immune checkpoint blockade (ICB) through CD8+ T cells dysfunction | [33] |
| Angiomyolipoma/Lymphangioleiomyomatosis (AML/LAM) | mTORC1 pathway | Immunosuppression and drug resistance | [59] | |
| Glioma | p53 from DNA damage repair alteration promote MK activation and overexpression | Facilitating glioma cells and microglia communication, lead to M2 polarization of microglia | [46] | |
| Tumor growth, progression | HCC | PI3K/AKT/NF-κB/TrkB signaling | Intensifying anoikis resistance, supporting recurrence and metastasis | [69] |
| HCC | STAT3 pathway activation via stimulating JAK1/2 phosphorylation | Tumor growth and invasion | [70] | |
| HCC, kidney and breast cancers | Disruption of LKB1-STRAD-Mo25 complex, leads to AMPK suppression | Cancer cell proliferation | [51] | |
| Neuroblastoma | Notch2-Hes-1 signaling | Tumorigenesis | [52] | |
| HNSCC | Tyrosine phosphorylation of FAK, paxillin and Stat1α pathway activation | Tumor cell migration and invasion | [39] | |
| NSCLC | NF-κB-Hes-1 signaling | Hypoxia-induced progression and metastasis | [49] | |
| Angiogenesis | NSCLC | VEGF-induced angiogenesis | Endothelial cells migration and neovascularization | |
| Breast cancer | Endothelial proliferation | Increasing vascular density | [53] | |
| HCC | Interaction with progranulin | Endothelial cells proliferation, migration, and tubulogenesis | [54] | |
| Epithelial-mesenchymal transition (EMT) | NSCLC | Notch2-NF-κB-Hes-1 signaling | Upregulation of EMT markers | [49] |
| Mitogenesis | Kidney cancer | JAK/STAT pathway | Autocrine tumor cells proliferation | [55] |
| Breast cancer | NF-κB-NR3C1 pathway | Tumor cells proliferation | [56] | |
| Anti-apoptosis | Prostate cancer | Extracellular signal-regulated kinase 1/2 and p38 mitogen-activated protein kinase pathway | Suppression of TNF-α-induced apoptosis | [58] |
| HCC | TRAIL/ActD-mediated pathway | Protect cells from caspase-3-mediated apoptosis | [57] | |
| Drug resistance | Neuroblastoma and osteosarcoma | AKT pathway | Cytoprotection of neighboring drug-sensitive cells, contribute to chemotherapy resistance | [71] |
| Prostate cancer | PI3K/AKT and MAPK/ERK pathway | Reduce quercetin efficacy on prostate cancer stem cell proliferation | [72] |
| Patent Number | Disease/Process | Treatment | Company, Publish Date |
|---|---|---|---|
| WO2018016674A1 | Brain cancer | An MK inhibitor to overcome resistance to temozolomide and/or radiation therapy | Samsung Electronic Co. Ltd., Suwon, South Korea, 2018 |
| WO2016058047A1 | Cancer, inflammatory and auto-immune diseases | Humanized antibody, IP14, to block MK | Cellmid Limited, Sydney, Australia, 2016 |
| US8080649B2 | Cancer, autoimmune disease, postoperative adhesion, and endometriosis | Aptamer possessing inhibitory activity against MK | Ribomic Inc, Tokyo, Japan, 2011 |
| US20110159022A1 | Cancers | Peptide derived from MK with CD4 T/CD8 T epitope restricted by HLA molecules, as anticancer vaccine or monitoring of cellular response against MK during cancer or anticancer treatment | Commissariat a lEnergie Atomique et aux Energies Alternatives, Paris, France, 2011 |
| US9658233B1 | Tumor growth diagnosis | A kit and method to determine MK and pleiotrophin level | Dept of Health and Human Services, Washington, DC, USA, 2017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kam, N.-W.; Lau, C.-Y.; Che, C.-M.; Lee, V.H.-F. Nasopharynx Battlefield: Cellular Immune Responses Mediated by Midkine in Nasopharyngeal Carcinoma and COVID-19. Cancers 2023, 15, 4850. https://doi.org/10.3390/cancers15194850
Kam N-W, Lau C-Y, Che C-M, Lee VH-F. Nasopharynx Battlefield: Cellular Immune Responses Mediated by Midkine in Nasopharyngeal Carcinoma and COVID-19. Cancers. 2023; 15(19):4850. https://doi.org/10.3390/cancers15194850
Chicago/Turabian StyleKam, Ngar-Woon, Cho-Yiu Lau, Chi-Ming Che, and Victor Ho-Fun Lee. 2023. "Nasopharynx Battlefield: Cellular Immune Responses Mediated by Midkine in Nasopharyngeal Carcinoma and COVID-19" Cancers 15, no. 19: 4850. https://doi.org/10.3390/cancers15194850