
Pseudogenes in Cancer: State of the Art

Abstract
:Simple Summary
Abstract
1. Introduction
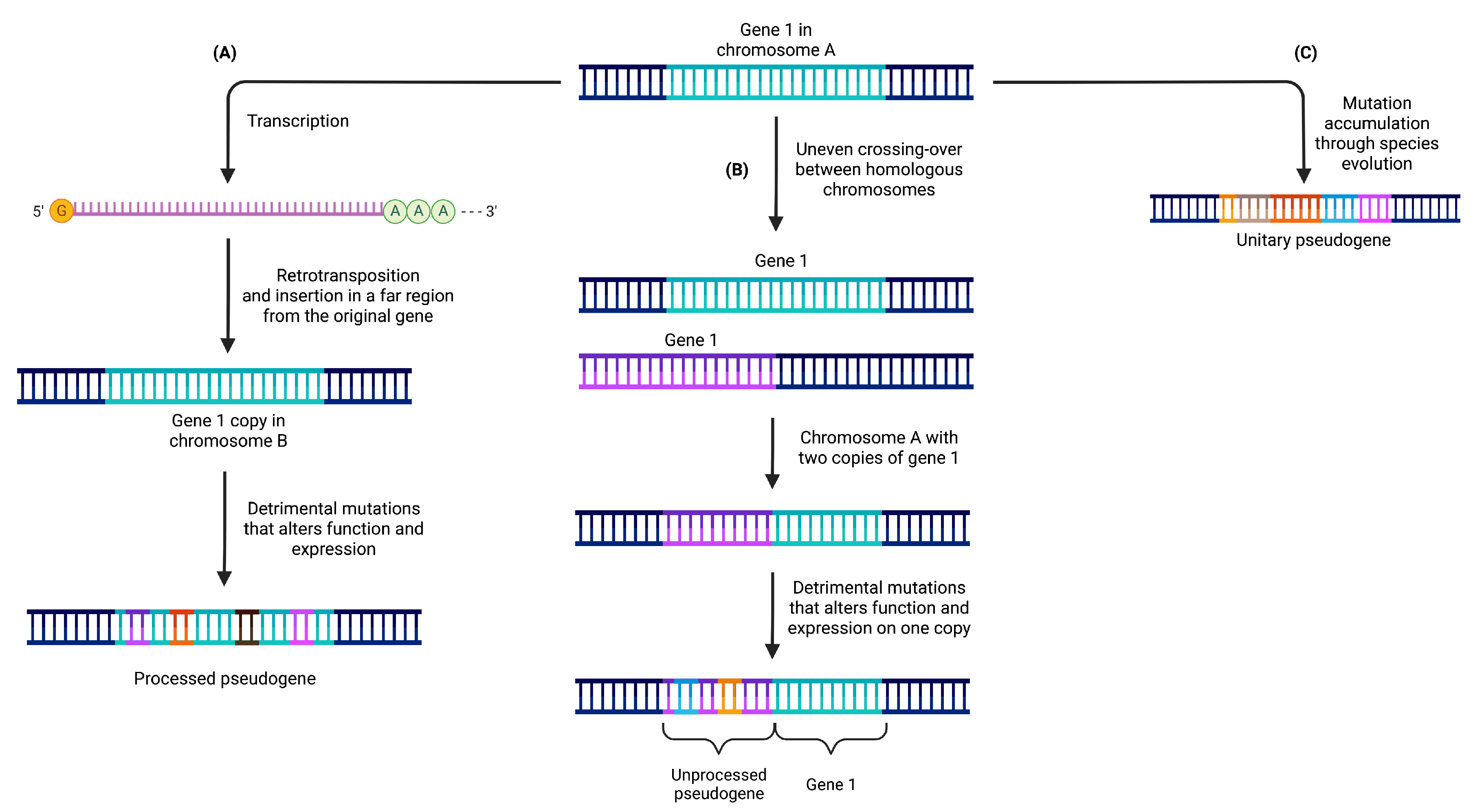
2. Classification of Pseudogenes
2.1. Processed Pseudogenes
2.2. Unprocessed Pseudogenes
2.3. Unitary Pseudogenes
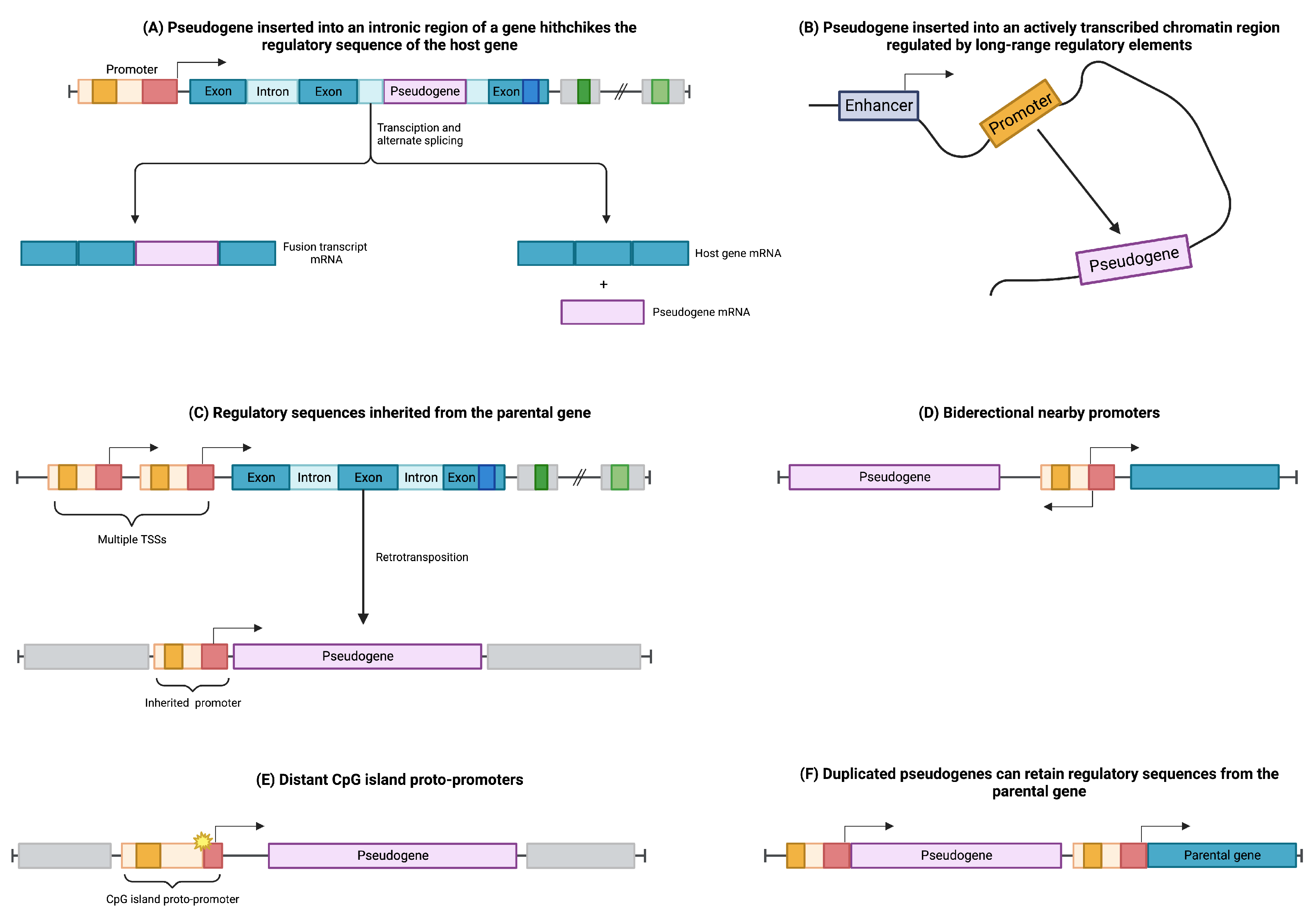
2.4. Regulation of Pseudogene Expression
3. Functional Role of Pseudogenes
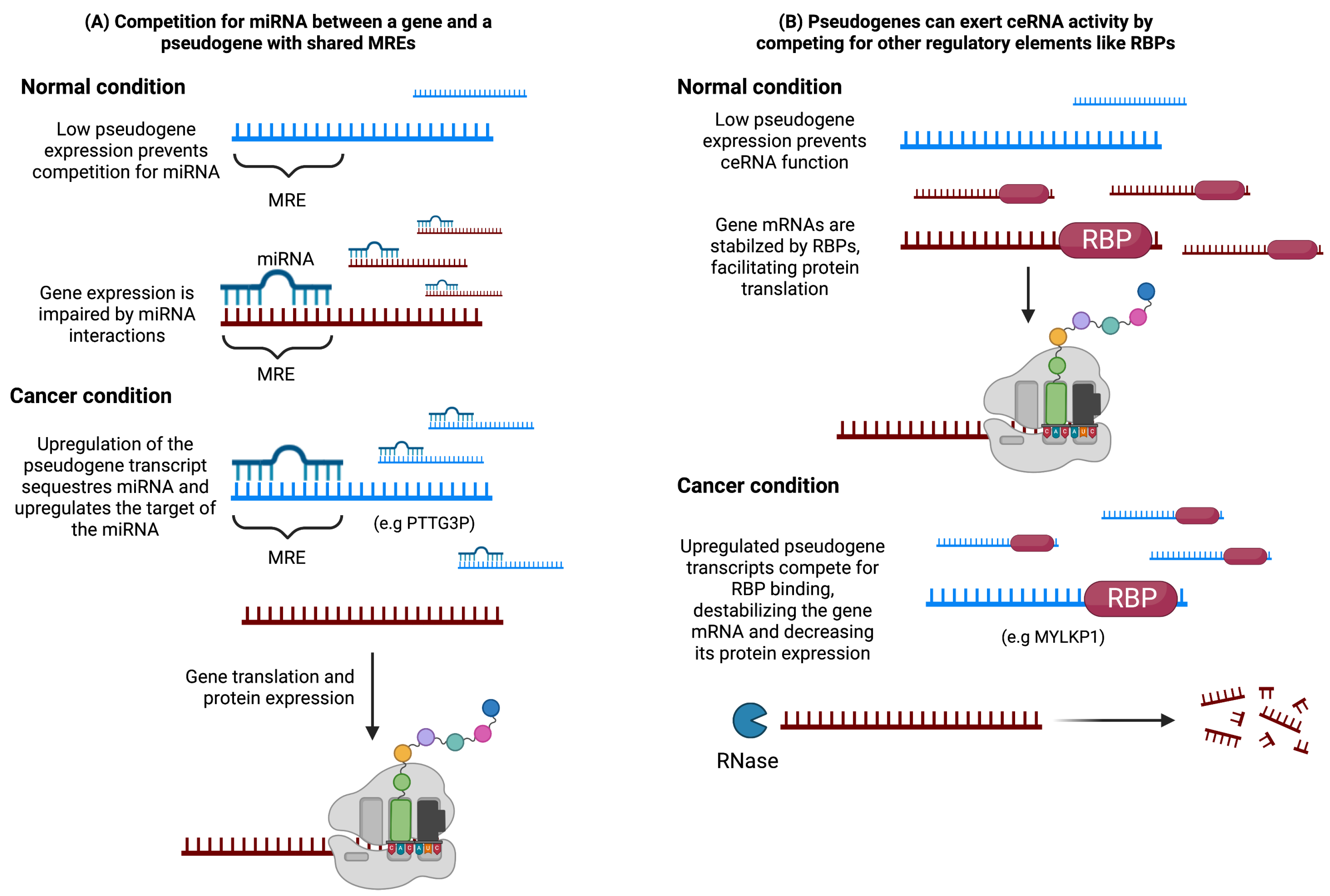
3.1. Pseudogenes as Endogenous Competitors
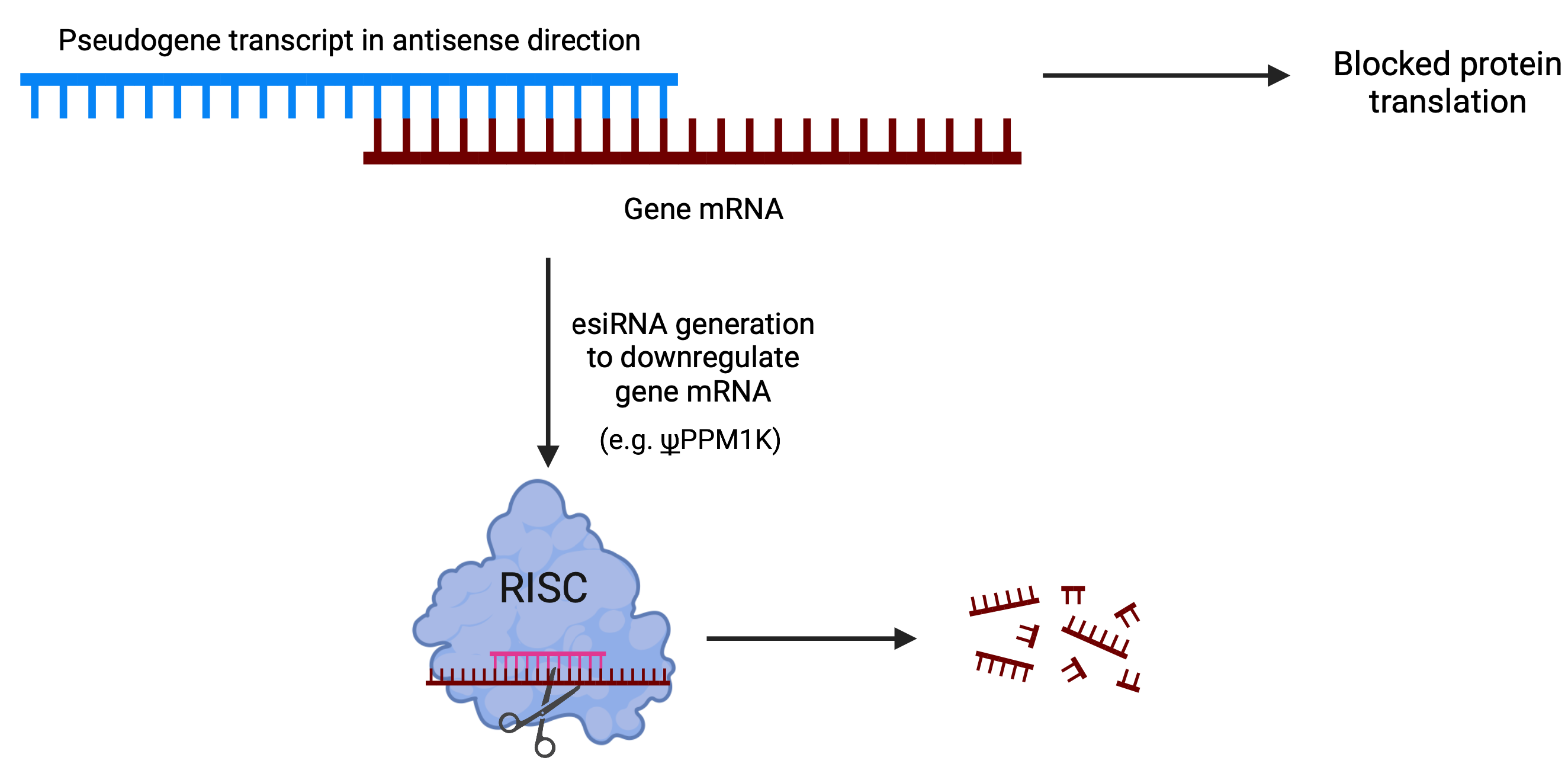
3.2. Pseudogene-Mediated Hybridization with Coding Genes
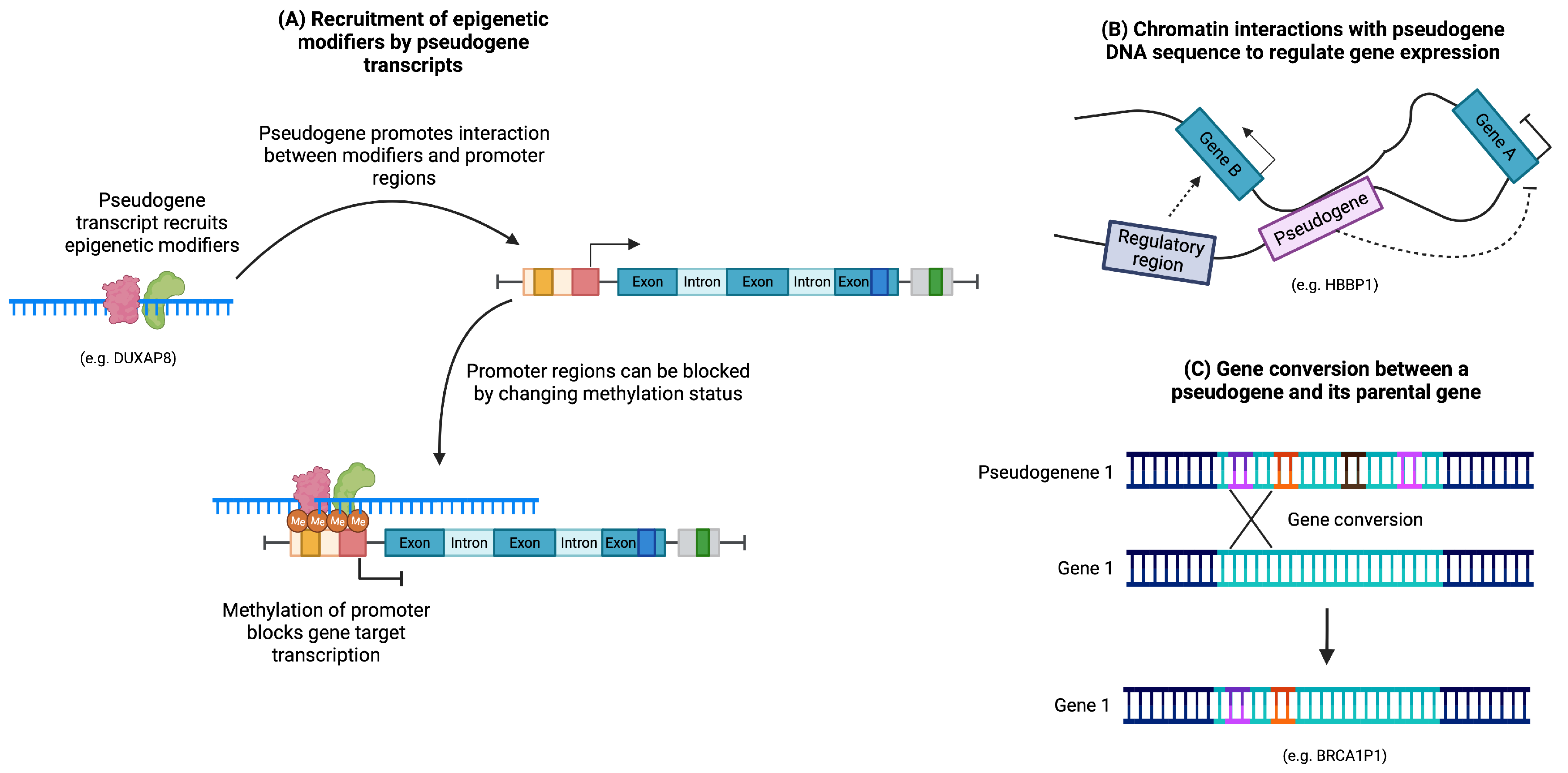
3.3. Regulation of DNA Structure
4. Pseudogenes as Functional Molecules in Cancer
4.1. Pseudogenes as ceRNAs in Cancer
4.2. Pseudogenes as Cancer Markers
4.3. Pseudogene Hybridization in Cancer
4.4. Pseudogenes Altering DNA Structure in Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pseudogene | Related Cancer | Action Mechanism | Ref |
|---|---|---|---|
| ADAM5 | Oropharynx squamous cell carcinoma | Upregulates ADAM9 by binding miR-122b-5p | [58] |
| ACTG1P25 | Breast cancer | Its upregulation promotes endocrine therapy resistance by competing with E2F1 for binding to PURA, thereby increasing E2F1 expression and activating cell cycle-related genes | [83] |
| AK4P1 | Pancreatic cancer | Upregulates genes related with cell proliferation through binding with tumor suppressive miR-375 | [84,85,86] |
| BRAFP1 | Thyroid tumors, among others | Elevate BRAF expression and MAPK activation through ceRNA mechanism | [87,88] |
| BRCA1P1 | Breast and ovarian cancer | A recombination between BRCA1P1 and BRCA1 can remove the promoter and initiation codon of BRCA1, thus blocking its tumor-suppressive functions | [89] |
| CYP2A7 | Lung cancer | Gene conversion with its parental gene (CYP2A6) produces a polymorphism with enhanced nicotine metabolism and associated with an increased cigarette consumption | [74,75] |
| CYP4Z2P | Breast cancer | The ceRNA network of CYP4Z1 and pseudogene CYP4Z2P inhibit apoptosis in cancer cells by sharing miRNA miR-125a-3p binding sites | [90] |
| DUXAP8 | Lung and pancreatic cance | Contributes to cancer progression by recruitment of epigenetic machinery to silence tumor suppressive genes | [80,81] |
| EBLN3P | Non-small cell lung cancer, osteosarcoma and colorectal cancer | Promotes cancer cell proliferation and epithelial-mesenchymal transition by sponging of different miRNAs | [91,92,93,94] |
| FTH1P3 | Non-small cell lung cancer | Recruits LSD1 to epigenetically downregulate TIMP3, promoting tumor malignancy | [95] |
| FLT1P1-S | Colorectal cancer | Positive regulator of VEGFR1 expression (opposite regulatory function from FLT1P1-AS) | [61] |
| OGFRP1 | Gastric cancer | Suppresses cell apoptosis by regulating the miR-149-5p/MAP3K3 axis. | [96] |
| LGMNP1 | Glioblastoma | Promotes aggresiveness of cancer cells by downregulating miR-495-3p, possibly through a RISC related mechanism | [97] |
| MSTO2P | Colorectal cancer | Promotes cancer cell proliferation by epigenetically downregulating CDKN1A through binding with EZH2 | [98] |
| MYLKP1 | Lung cancer | MYLKP1 is overexpressed in cancer cells and downregulates smMLCK, possibly by decresing its stability through competition for RBPs | [64,65] |
| OCT4-pg4 | HCC | Upregulates OCT4 by sequestering miR-145, promoting tumorigenicity | [99,100] |
| PCNAP1 | Breast cancer | Promotes invasion of cancer cells by binding with miR-340-5p, hence upregulating SOX4 | [101] |
| PDIA3P1 | Glioma | Sequestrates miR-124-3p to upregulate RELA expression, promoting glioma cells MES transition by activating the NF-B pathway | [102] |
| PPM1K | HCC | Produces esiRNAs that downregulates NEK8, inhibiting cell growth | [73] |
| PRELID1P6 | Glioma | Promotes cancer cell proliferation by upregulating hnRNPH1 and TRF2, which activates the Akt/mTOR pathway. It is downregulated by miR-1825. | [103] |
| PTENP1-AS | Melanoma | Recruits EZH2 and H3K27me3 to downregulate PTEN expression | [104] |
| PTTG3P | Oral and prostate cancer | Functions as ceRNA by binding with miR-142-5p (oral) and miR-146a-3p (prostate) | [59,60] |
| RPSAP52 | Breast cancer and sarcoma cell lines | Contributes to cancer progression by controlling the HMGA2/IGF2BP2/LIN28B axis and downregulating et-7 miRNAs | [105] |
| SALL4P5 | HCC | Demethylates SALL4 by interacting with DNMT1 | [82] |
| TCAM1P | Cervical cancer | Regulates cell cycle and promotes cancer cell proliferation, its expression is HVP-dependent and is regulated by HPV E6/E7 and EIF4A3 | [62] |
| TDGF1P3 | Colorectal cancer | Upregulates PKM2 by competing and binding with miR-338-3p | [106] |
| RP9P | Colorectal cancer | Promotes colorectal cancer progression through upregulation of FOXQ1 by competing for miR-133a-3p | [107] |
| UBE2CP3 | Gastric cancer | Promotes gastric cancer progression by upregulating ITGA2 through binding with miR-138-5p | [108] |
| Pseudogene | Related Cancer | Action Mechanism | Ref |
|---|---|---|---|
| ARHGAP27P1 | Gastric cancer | Exerts tumor-suppressive functions by interacting with JMJD3 and epigenetically activation of p15, p16 and p57 | [109] |
| FLT1P1-AS | Colorectal cancer | Downregulates VEGFR1 and VEGF-A by interacting with miR-520a and by blocking VEGFR1 translation | [61] |
| FOXO3P | Breast cancer | Suppresses tumor growth by binding with multiple miRNA, thus upregulating FOXO3 mRNA | [110] |
| Pseudogenes of FTH1 | Prostate cancer | The ceRNA networks formed by FTH1 and their pseudogenes exerts a tumor suppressive effect by bind with multiple miRNAs | [111] |
| GUSBP11 | Triple negative breast cancer | Inhibits cancer progression by upregulating SPNS2 through sequestering miR-579-3p | [112] |
| MT1JP | Triple negative breast cancer | Inhibits TNBC by regulating the miR-138/HIF-1 axis | [113] |
| PEBP1P2 | Clear cell renal cell carcinoma | Exerts tumor suppressive effects by recruiting the YBX1/ELAVL1 complex to stabilize PEBP1; additionally, can act as a ceRNA for KLF13 sponging several miRNAs | [114] |
| SNRPFP1 | HCC | Binds with miR-125-5p blocking its tumor suppressive action | [56] |
| TUSC2P | Esophageal squamous cell carcinoma | TUSC2P 3′UTR serves as a decoy to protect TUSC2 from binding with miR-17-5p, miR-520a-3p, miR-608, miR-661, thus upregulating its translation and inhibiting cancer cell survival and proliferation | [115,116] |
5. Co-Expression of Pseudogenes
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri-Fard, S.; Poulet, C.; Malaise, M.; Abak, A.; Mahmud Hussen, B.; Taheriazam, A.; Taheri, M.; Hallajnejad, M. The emerging role of non-coding RNAs in osteoarthritis. Front. Immunol. 2021, 12, 773171. [Google Scholar] [CrossRef] [PubMed]
- Ebert, M.S.; Sharp, P.A. Roles for microRNAs in conferring robustness to biological processes. Cell 2012, 149, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Pagni, S.; Mills, J.D.; Frankish, A.; Mudge, J.M.; Sisodiya, S.M. Non-coding regulatory elements: Potential roles in disease and the case of epilepsy. Neuropathol. Appl. Neurobiol. 2022, 48, e12775. [Google Scholar] [CrossRef] [PubMed]
- Shefler, A.; Patrick, M.T.; Wasikowski, R.; Chen, J.; Sarkar, M.K.; Gudjonsson, J.E.; Tsoi, L.C. Skin-expressing lncRNAs in inflammatory responses. Front. Genet. 2022, 13, 835740. [Google Scholar] [CrossRef]
- Lin, J.; Li, X.; Zhang, F.; Zhu, L.; Chen, Y. Transcriptome wide analysis of long non-coding RNA-associated ceRNA regulatory circuits in psoriasis. J. Cell. Mol. Med. 2021, 25, 6925–6935. [Google Scholar] [CrossRef]
- Perenthaler, E.; Yousefi, S.; Niggl, E.; Barakat, T.S. Beyond the exome: The non-coding genome and enhancers in neurodevelopmental disorders and malformations of cortical development. Front. Cell. Neurosci. 2019, 13, 352. [Google Scholar] [CrossRef] [Green Version]
- Jacq, C.; Miller, J.; Brownlee, G. A pseudogene structure in 5S DNA of Xenopus laevis. Cell 1977, 12, 109–120. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, D.; Yadava, A.; Srivastava, A.K. Molecular fossils “pseudogenes” as functional signature in biological system. Genes Genom. 2020, 42, 619–630. [Google Scholar] [CrossRef]
- Kalyana-Sundaram, S.; Kumar-Sinha, C.; Shankar, S.; Robinson, D.R.; Wu, Y.M.; Cao, X.; Asangani, I.A.; Kothari, V.; Prensner, J.R.; Lonigro, R.J.; et al. Expressed pseudogenes in the transcriptional landscape of human cancers. Cell 2012, 149, 1622–1634. [Google Scholar] [CrossRef] [Green Version]
- Schiff, C.; Milili, M.; Fougereau, M. Functional and pseudogenes are similarly organized and may equally contribute to the extensive antibody diversity of the IgVHII family. EMBO J. 1985, 4, 1225–1230. [Google Scholar] [CrossRef]
- Rothenfluh, H.S.; Blanden, R.V.; Steele, E.J. Evolution of V genes: DNA sequence structure of functional germline genes and pseudogenes. Immunogenetics 1995, 42, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Balakirev, E.S.; Chechetkin, V.; Lobzin, V.; Ayala, F.J. DNA polymorphism in the β-esterase gene cluster of Drosophila melanogaster. Genetics 2003, 164, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Sudbrak, R.; Reinhardt, R.; Hennig, S.; Lehrach, H.; Günther, E.; Walter, L. Comparative and evolutionary analysis of the rhesus macaque extended MHC class II region. Immunogenetics 2003, 54, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Svensson, Ö.; Arvestad, L.; Lagergren, J. Genome-wide survey for biologically functional pseudogenes. PLoS Comput. Biol. 2006, 2, e46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khachane, A.N.; Harrison, P.M. Assessing the genomic evidence for conserved transcribed pseudogenes under selection. BMC Genom. 2009, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Cheetham, S.W.; Faulkner, G.J.; Dinger, M.E. Overcoming challenges and dogmas to understand the functions of pseudogenes. Nat. Rev. Genet. 2020, 21, 191–201. [Google Scholar] [CrossRef]
- Geissler, R.; Simkin, A.; Floss, D.; Patel, R.; Fogarty, E.A.; Scheller, J.; Grimson, A. A widespread sequence-specific mRNA decay pathway mediated by hnRNPs A1 and A2/B1. Genes Dev. 2016, 30, 1070–1085. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Graber, T.; Jordan, L.; Cloutier, M.; Lewis, S.; Goulet, I.; Côté, J.; Holcik, M. hnRNP A1 regulates UV-induced NF-κB signalling through destabilization of cIAP1 mRNA. Cell Death Differ. 2009, 16, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Liu, S.; Gao, J.; Chen, C.; Zhang, X.; Yuan, H.; Chen, Z.; Yin, X.; Sun, C.; Mao, Y.; et al. Genome-wide analysis of pseudogenes reveals HBBP1’s human-specific essentiality in erythropoiesis and implication in β-thalassemia. Dev. Cell 2021, 56, 478–493. [Google Scholar] [CrossRef]
- Poliseno, L. Pseudogenes: Newly discovered players in human cancer. Sci. Signal. 2012, 5, re5. [Google Scholar] [CrossRef] [PubMed]
- Pink, R.C.; Wicks, K.; Caley, D.P.; Punch, E.K.; Jacobs, L.; Carter, D.R.F. Pseudogenes: Pseudo-functional or key regulators in health and disease? RNA 2011, 17, 792–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazazian, H.H. Processed pseudogene insertions in somatic cells. Mob. DNA 2014, 5, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troskie, R.L.; Faulkner, G.J.; Cheetham, S.W. Processed pseudogenes: A substrate for evolutionary innovation: Retrotransposition contributes to genome evolution by propagating pseudogene sequences with rich regulatory potential throughout the genome. BioEssays 2021, 43, 2100186. [Google Scholar] [CrossRef]
- Kanber, D.; Buiting, K.; Roos, C.; Gromoll, J.; Kaya, S.; Horsthemke, B.; Lohmann, D. The origin of the RB1 imprint. PLoS ONE 2013, 8, e81502. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Harrison, P.M.; Liu, Y.; Gerstein, M. Millions of years of evolution preserved: A comprehensive catalog of the processed pseudogenes in the human genome. Genome Res. 2003, 13, 2541–2558. [Google Scholar] [CrossRef]
- Ding, W.; Lin, L.; Chen, B.; Dai, J. L1 elements, processed pseudogenes and retrogenes in mammalian genomes. IUBMB Life 2006, 58, 677–685. [Google Scholar] [CrossRef]
- Wenke, T.; Döbel, T.; Sörensen, T.R.; Junghans, H.; Weisshaar, B.; Schmidt, T. Targeted identification of short interspersed nuclear element families shows their widespread existence and extreme heterogeneity in plant genomes. Plant Cell 2011, 23, 3117–3128. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Chen, Z.; Yu, J. Pseudogenes and their potential functions in hematopoiesis. Exp. Hematol. 2021, 103, 24–29. [Google Scholar] [CrossRef]
- Podlaha, O.; Zhang, J. Processed pseudogenes: The ‘fossilized footprints’ of past gene expression. Trends Genet. 2009, 25, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Mascagni, F.; Usai, G.; Cavallini, A.; Porceddu, A. Structural characterization and duplication modes of pseudogenes in plants. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Frankish, A.; Hunt, T.; Harrow, J.; Gerstein, M. Identification and analysis of unitary pseudogenes: Historic and contemporary gene losses in humans and other primates. Genome Biol. 2010, 11, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H. Conserved or lost: Molecular evolution of the key gene GULO in vertebrate vitamin C biosynthesis. Biochem. Genet. 2013, 51, 413–425. [Google Scholar] [CrossRef]
- Vinckenbosch, N.; Dupanloup, I.; Kaessmann, H. Evolutionary fate of retroposed gene copies in the human genome. Proc. Natl. Acad. Sci. USA 2006, 103, 3220–3225. [Google Scholar] [CrossRef]
- Kaessmann, H.; Vinckenbosch, N.; Long, M. RNA-based gene duplication: Mechanistic and evolutionary insights. Nat. Rev. Genet. 2009, 10, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Nakai, K. Retrotransposition as a source of new promoters. Mol. Biol. Evol. 2008, 25, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gerstein, M. Large-scale analysis of pseudogenes in the human genome. Curr. Opin. Genet. Dev. 2004, 14, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Song, R.; Regev, A.; Struhl, K. Many lncRNAs, 5′UTRs, and pseudogenes are translated and some are likely to express functional proteins. elife 2015, 4, e08890. [Google Scholar] [CrossRef]
- Suzuki, I.K.; Gacquer, D.; Van Heurck, R.; Kumar, D.; Wojno, M.; Bilheu, A.; Herpoel, A.; Lambert, N.; Cheron, J.; Polleux, F.; et al. Human-specific NOTCH2NL genes expand cortical neurogenesis through Delta/Notch regulation. Cell 2018, 173, 1370–1384. [Google Scholar] [CrossRef] [Green Version]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejepinska, J.; Flemr, M.; Svoboda, P. The Canonical RNA Interference Pathway in Animals. In Regulatory RNAs: Basics, Methods and Applications; Mallick, B., Ghosh, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 111–149. [Google Scholar] [CrossRef]
- Tam, O.H.; Aravin, A.A.; Stein, P.; Girard, A.; Murchison, E.P.; Cheloufi, S.; Hodges, E.; Anger, M.; Sachidanandam, R.; Schultz, R.M.; et al. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 2008, 453, 534–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Totoki, Y.; Toyoda, A.; Kaneda, M.; Kuramochi-Miyagawa, S.; Obata, Y.; Chiba, H.; Kohara, Y.; Kono, T.; Nakano, T.; et al. Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes. Nature 2008, 453, 539–543. [Google Scholar] [CrossRef]
- Wen, Y.Z.; Zheng, L.L.; Liao, J.Y.; Wang, M.H.; Wei, Y.; Guo, X.M.; Qu, L.H.; Ayala, F.J.; Lun, Z.R. Pseudogene-derived small interference RNAs regulate gene expression in African Trypanosoma brucei. Proc. Natl. Acad. Sci. USA 2011, 108, 8345–8350. [Google Scholar] [CrossRef]
- Boeren, J.; Gribnau, J. Xist-mediated chromatin changes that establish silencing of an entire X chromosome in mammals. Curr. Opin. Cell Biol. 2021, 70, 44–50. [Google Scholar] [CrossRef]
- Fang, H.; Disteche, C.M.; Berletch, J.B. X inactivation and escape: Epigenetic and structural features. Front. Cell Dev. Biol. 2019, 7, 219. [Google Scholar] [CrossRef]
- Loda, A.; Heard, E. Xist RNA in action: Past, present, and future. PLoS Genet. 2019, 15, e1008333. [Google Scholar] [CrossRef] [Green Version]
- Elisaphenko, E.A.; Kolesnikov, N.N.; Shevchenko, A.I.; Rogozin, I.B.; Nesterova, T.B.; Brockdorff, N.; Zakian, S.M. A dual origin of the Xist gene from a protein-coding gene and a set of transposable elements. PLoS ONE 2008, 3, e2521. [Google Scholar] [CrossRef] [Green Version]
- Duret, L.; Chureau, C.; Samain, S.; Weissenbach, J.; Avner, P. The Xist RNA gene evolved in eutherians by pseudogenization of a protein-coding gene. Science 2006, 312, 1653–1655. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Keller, C.A.; Giardine, B.; Grevet, J.D.; Davies, J.O.; Hughes, J.R.; Kurita, R.; Nakamura, Y.; Hardison, R.C.; Blobel, G.A. Comparative analysis of three-dimensional chromosomal architecture identifies a novel fetal hemoglobin regulatory element. Genes Dev. 2017, 31, 1704–1713. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Madrazo, E.; Almagro, J.C.; Lara-Ochoa, F. Structural repertoire in VHpseudogenes of immunoglobulins: Comparison with human germline genes and human amino acid sequences. J. Mol. Biol. 1995, 246, 74–81. [Google Scholar] [CrossRef]
- Sisu, C. Pseudogenes as Biomarkers Biomarkers and Therapeutic Targets Therapeutic targets in Human Cancers. Methods Mol. Biol. 2021, 2324, 319–337. [Google Scholar]
- Stasiak, M.; Kolenda, T.; Kozłowska-Masłoń, J.; Sobocińska, J.; Poter, P.; Guglas, K.; Paszkowska, A.; Bliźniak, R.; Teresiak, A.; Kazimierczak, U.; et al. The world of pseudogenes: New diagnostic and therapeutic targets in cancers or still mystery molecules? Life 2021, 11, 1354. [Google Scholar] [CrossRef]
- Wang, N.; Guo, S.; Hao, F.; Zhang, Y.; Chen, Y.; Fei, X.; Wang, J. Pseudogene SNRPFP1 derived long non-coding RNA facilitates hepatocellular carcinoma progress in vitro by sponging tumor-suppressive miR-126-5p. Sci. Rep. 2022, 12, 21867. [Google Scholar] [CrossRef]
- Vo, D.T.; Karanam, N.K.; Ding, L.; Saha, D.; Yordy, J.S.; Giri, U.; Heymach, J.V.; Story, M.D. miR-125a-5p functions as tumor suppressor microRNA and is a marker of locoregional recurrence and poor prognosis in head and neck cancer. Neoplasia 2019, 21, 849–862. [Google Scholar] [CrossRef]
- Carron, J.; Torricelli, C.; Silva, J.K.; Liu, Y.; Pellegrino, R.; Lima, C.S.P.; Lourenço, G.J. Association of Inherited Copy Number Variation in ADAM3A and ADAM5 Pseudogenes with Oropharynx Cancer Risk and Outcome. Genes 2022, 13, 2408. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, X.; Su, K.; Zheng, Q.; Liu, P.; Xu, Z.; Zhang, Y. A novel mechanism of the lncRNA PTTG3P/miR-142-5p/JAG1 axis modulating tongue cancer cell phenotypes through the Notch1 signaling. Cells Dev. 2022, 169, 203762. [Google Scholar] [CrossRef]
- Huang, S.; Liao, Q.; Li, W.; Deng, G.; Jia, M.; Fang, Q.; Ji, H.; Meng, M. The lncRNA PTTG3P promotes the progression of CRPC via upregulating PTTG1. Bull. Cancer 2021, 108, 359–368. [Google Scholar] [CrossRef]
- Ye, X.; Fan, F.; Bhattacharya, R.; Bellister, S.; Boulbes, D.R.; Wang, R.; Xia, L.; Ivan, C.; Zheng, X.; Calin, G.A.; et al. VEGFR-1 Pseudogene Expression and Regulatory Function in Human Colorectal Cancer CellsVEGFR1 Pseudogene Expression and Function in Colorectal Cancer Cells. Mol. Cancer Res. 2015, 13, 1274–1282. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Ren, C.; Yang, L.; Zhang, Z.; Gong, M.; Chen, K. Identification and validation of the high expression of pseudogene TCAM1P in cervical cancer via integrated bioinformatics analysis. Cancer Cell Int. 2022, 22, 17. [Google Scholar] [CrossRef]
- Lazar, V.; Garcia, J.G. A single human myosin light chain kinase gene (MLCK; MYLK) transcribes multiple nonmuscle isoforms. Genomics 1999, 57, 256–267. [Google Scholar] [CrossRef]
- Han, Y.J.; Ma, S.F.; Yourek, G.; Park, Y.D.; Garcia, J.G. A transcribed pseudogene of MYLK promotes cell proliferation. FASEB J. 2011, 25, 2305. [Google Scholar] [CrossRef]
- Hu, X.; Yang, L.; Mo, Y.Y. Role of pseudogenes in tumorigenesis. Cancers 2018, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.; Xiao, C.; Yan, C.; Chen, D.; Huang, Q.; Fan, Y.; Li, Z.; Xu, H. Knockdown of pseudogene derived from lncRNA DUXAP10 inhibits cell proliferation, migration, invasion, and promotes apoptosis in pancreatic cancer. J. Cell. Biochem. 2018, 119, 3671–3682. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, X.; Wei, C.; Nie, F.; Huang, M.; Sun, M. Over-expression of oncigenic pesudogene DUXAP10 promotes cell proliferation and invasion by regulating LATS1 and β-catenin in gastric cancer. J. Exp. Clin. Cancer Res. 2018, 37, 1–12. [Google Scholar] [CrossRef]
- Lian, Y.; Xu, Y.; Xiao, C.; Xia, R.; Gong, H.; Yang, P.; Chen, T.; Wu, D.; Cai, Z.; Zhang, J.; et al. The pseudogene derived from long non-coding RNA DUXAP10 promotes colorectal cancer cell growth through epigenetically silencing of p21 and PTEN. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Moreau-Aubry, A.; Le Guiner, S.; Labarrière, N.; Gesnel, M.C.; Jotereau, F.; Breathnach, R. A processed pseudogene codes for a new antigen recognized by a CD8+ T cell clone on melanoma. J. Exp. Med. 2000, 191, 1617–1624. [Google Scholar] [CrossRef]
- Hendrickson, R.C.; Cicinnati, V.R.; Albers, A.; Dworacki, G.; Gambotto, A.; Pagliano, O.; Tüting, T.; Mayordomo, J.I.; Visus, C.; Appella, E.; et al. Identification of a 17β-hydroxysteroid dehydrogenase type 12 pseudogene as the source of a highly restricted BALB/c Meth A tumor rejection peptide. Cancer Immunol. Immunother. 2010, 59, 113–124. [Google Scholar] [CrossRef]
- Poliseno, L.; Marranci, A.; Pandolfi, P.P. Pseudogenes in human cancer. Front. Med. 2015, 2, 68. [Google Scholar] [CrossRef] [Green Version]
- Xiang, R.; Ma, L.; Yang, M.; Zheng, Z.; Chen, X.; Jia, F.; Xie, F.; Zhou, Y.; Li, F.; Wu, K.; et al. Increased expression of peptides from non-coding genes in cancer proteomics datasets suggests potential tumor neoantigens. Commun. Biol. 2021, 4, 496. [Google Scholar] [CrossRef]
- Chan, W.L.; Yuo, C.Y.; Yang, W.K.; Hung, S.Y.; Chang, Y.S.; Chiu, C.C.; Yeh, K.T.; Huang, H.D.; Chang, J.G. Transcribed pseudogene ψPPM1K generates endogenous siRNA to suppress oncogenic cell growth in hepatocellular carcinoma. Nucleic Acids Res. 2013, 41, 3734–3747. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Pitarque, M.; Ingelman-Sundberg, M. 3′-UTR polymorphism in the human CYP2A6 gene affects mRNA stability and enzyme expression. Biochem. Biophys. Res. Commun. 2006, 340, 491–497. [Google Scholar] [CrossRef]
- Gambier, N.; Batt, A.; Marie, B.; Pfister, M.; Siest, G.; Visvikis-Siest, S. Association of CYP2A6* 1B genetic variant with the amount of smoking in French adults from the Stanislas cohort. Pharmacogenomics J. 2005, 5, 271–275. [Google Scholar] [CrossRef]
- Yang, S.J.; Weng, J.L.; Wei, B.; Du, X.K. Long non-coding RNA DUXAP8 regulates the cell proliferation and invasion of non-small-cell lung cancer. Open Life Sci. 2019, 14, 201–207. [Google Scholar] [CrossRef]
- Li, L.; Hao, S.; Ni, M.; Jin, S.; Tian, Y. DUXAP8 promotes the proliferation and migration of ovarian cancer cells via down-regulating microRNA-29a-3p expression. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 1837–1844. [Google Scholar]
- Ma, H.w.; Xie, M.; Sun, M.; Chen, T.y.; Jin, R.r.; Ma, T.s.; Chen, Q.n.; Zhang, E.b.; He, X.z.; De, W.; et al. The pseudogene derived long noncoding RNA DUXAP8 promotes gastric cancer cell proliferation and migration via epigenetically silencing PLEKHO1 expression. Oncotarget 2017, 8, 52211. [Google Scholar] [CrossRef] [Green Version]
- Pang, R.; Yang, S. lncRNA DUXAP8 inhibits papillary thyroid carcinoma cell apoptosis via sponging the miR-20b-5p/SOS1 axis. Oncol. Rep. 2021, 45, 1–10. [Google Scholar] [CrossRef]
- Sun, M.; Nie, F.q.; Zang, C.; Wang, Y.; Hou, J.; Wei, C.; Li, W.; He, X.; Lu, K.h. The pseudogene DUXAP8 promotes non-small-cell lung cancer cell proliferation and invasion by epigenetically silencing EGR1 and RHOB. Mol. Ther. 2017, 25, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Lian, Y.; Yang, J.; Lian, Y.; Xiao, C.; Hu, X.; Xu, H. DUXAP8, a pseudogene derived lncRNA, promotes growth of pancreatic carcinoma cells by epigenetically silencing CDKN1A and KLF2. Cancer Commun. 2018, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.; Liu, Y.V.; Gao, C.; Bassal, M.A.; Jones, A.I.; Yang, J.; Chen, Z.; Li, Y.; Yang, H.; Chen, L.; et al. Pseudogene-mediated DNA demethylation leads to oncogene activation. Sci. Adv. 2021, 7, eabg1695. [Google Scholar] [CrossRef]
- Yu, S.; Wang, Y.; Gong, X.; Fan, Z.; Wang, Z.; Liang, Z.; Wu, R.; Cao, B.; Wang, N.; Bi, C.; et al. LncRNA AGPG confers endocrine resistance in breast cancer by promoting E2F1 activity. Cancer Res. 2023; Online ahead of print. [Google Scholar]
- Xu, W.; Lou, W.; Mei, L. A key regulatory loop AK4P1/miR-375/SP1 in pancreatic adenocarcinoma. Epigenetics 2023, 18, 2148433. [Google Scholar] [CrossRef]
- Li, L.; Deng, T.; Zhang, Q.; Yang, Y.; Liu, Y.; Yuan, L.; Xie, M. AK4P1 is a cancer-promoting pseudogene in pancreatic adenocarcinoma cells whose transcripts can be transmitted by exosomes. Oncol. Lett. 2022, 23, 1–9. [Google Scholar] [CrossRef]
- Jia, L.; Zhang, Y.; Pu, F.; Yang, C.; Yang, S.; Yu, J.; Xu, Z.; Yang, H.; Zhou, Y.; Zhu, S. Pseudogene AK4P1 promotes pancreatic ductal adenocarcinoma progression through relieving miR-375-mediated YAP1 degradation. Aging 2022, 14, 1983. [Google Scholar] [CrossRef]
- Zou, M.; Baitei, E.Y.; Alzahrani, A.S.; Al-Mohanna, F.; Farid, N.R.; Meyer, B.; Shi, Y. Oncogenic activation of MAP kinase by BRAF pseudogene in thyroid tumors. Neoplasia 2009, 11, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Karreth, F.A.; Reschke, M.; Ruocco, A.; Ng, C.; Chapuy, B.; Léopold, V.; Sjoberg, M.; Keane, T.M.; Verma, A.; Ala, U.; et al. The BRAF pseudogene functions as a competitive endogenous RNA and induces lymphoma in vivo. Cell 2015, 161, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Puget, N.; Gad, S.; Perrin-Vidoz, L.; Sinilnikova, O.M.; Stoppa-Lyonnet, D.; Lenoir, G.M.; Mazoyer, S. Distinct BRCA1 rearrangements involving the BRCA1 pseudogene suggest the existence of a recombination hot spot. Am. J. Hum. Genet. 2002, 70, 858–865. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zheng, L.; Xin, Y.; Tan, Z.; Zhang, Y.; Meng, X.; Wang, Z.; Xi, T. The competing endogenous RNA network of CYP 4Z1 and pseudogene CYP 4Z2P exerts an anti-apoptotic function in breast cancer. FEBS Lett. 2017, 591, 991–1000. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Huang, H.; Guo, Z.; Huang, H.; Niu, Z.; Ji, Y.; Zhang, Y.; Bian, H.; Hu, W. Heavy Ion-Responsive lncRNA EBLN3P Functions in the Radiosensitization of Non-Small Cell Lung Cancer Cells Mediated by TNPO1. Cancers 2023, 15, 511. [Google Scholar] [CrossRef]
- Dai, S.; Li, N.; Zhou, M.; Yuan, Y.; Yue, D.; Li, T.; Zhang, X. LncRNA EBLN3P promotes the progression of osteosarcoma through modifying the miR-224-5p/Rab10 signaling axis. Sci. Rep. 2021, 11, 1992. [Google Scholar] [CrossRef]
- Xu, X.h.; Song, W.; Li, J.h.; Huang, Z.q.; Liu, Y.f.; Bao, Q.; Shen, Z.w. Long non-coding RNA EBLN3P regulates UHMK1 expression by sponging miR-323a-3p and promotes colorectal cancer progression. Front. Med. 2021, 8, 651600. [Google Scholar] [CrossRef]
- Sun, M.X.; An, H.Y.; Sun, Y.B.; Sun, Y.b.; Bai, B. LncRNA EBLN3P attributes methotrexate resistance in osteosarcoma cells through miR-200a-3p/O-GlcNAc transferase pathway. J. Orthop. Surg. Res. 2022, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Chen, W.; Li, W.; Ding, Y.; Tu, P. E2F1-induced ferritin heavy chain 1 pseudogene 3 (FTH1P3) accelerates non-small cell lung cancer gefitinib resistance. Biochem. Biophys. Res. Commun. 2020, 530, 624–631. [Google Scholar] [CrossRef]
- Qin, H.; Li, X.; Zhang, W.; Ding, Z. LncRNA OGFRP1 promotes cell proliferation and suppresses cell radiosensitivity in gastric cancer by targeting the miR-149-5p/MAP3K3 axis. J. Mol. Histol. 2022, 53, 257–271. [Google Scholar] [CrossRef]
- Liao, K.; Qian, Z.; Zhang, S.; Chen, B.; Li, Z.; Huang, R.; Cheng, L.; Wang, T.; Yang, R.; Lan, J.; et al. The LGMN pseudogene promotes tumor progression by acting as a miR-495-3p sponge in glioblastoma. Cancer Lett. 2020, 490, 111–123. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, X. LncRNA MSTO2P promotes colorectal cancer progression through epigenetically silencing CDKN1A mediated by EZH2. World J. Surg. Oncol. 2022, 20, 1–9. [Google Scholar] [CrossRef]
- Scarola, M.; Comisso, E.; Rosso, M.; Del Sal, G.; Schneider, C.; Schoeftner, S.; Benetti, R. FUS-dependent loading of SUV39H1 to OCT4 pseudogene-lncRNA programs a silencing complex with OCT4 promoter specificity. Commun. Biol. 2020, 3, 632. [Google Scholar] [CrossRef]
- Hawkins, P.G.; Morris, K.V. Transcriptional regulation of Oct4 by a long non-coding RNA antisense to Oct4-pseudogene 5. Transcription 2010, 1, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; He, Y.; Shao, Y.; Chen, Q.; Liu, H. lncRNA PCNAP1 predicts poor prognosis in breast cancer and promotes cancer metastasis via miR-340-5p-dependent upregulation of SOX4. Oncol. Rep. 2020, 44, 1511–1523. [Google Scholar] [CrossRef]
- Wang, S.; Qi, Y.; Gao, X.; Qiu, W.; Liu, Q.; Guo, X.; Qian, M.; Chen, Z.; Zhang, Z.; Wang, H.; et al. Hypoxia-induced lncRNA PDIA3P1 promotes mesenchymal transition via sponging of miR-124-3p in glioma. Cell Death Dis. 2020, 11, 168. [Google Scholar] [CrossRef] [Green Version]
- Xi, S.; Cai, H.; Lu, J.; Zhang, Y.; Yu, Y.; Chen, F.; Huang, Q.; Wang, F.; Chen, Z. The pseudogene PRELID1P6 promotes glioma progression via the hnHNPH1-Akt/mTOR axis. Oncogene 2021, 40, 4453–4467. [Google Scholar] [CrossRef]
- Vidarsdottir, L.; Azimi, A.; Das, I.; Sigvaldadottir, I.; Suryo Rahmanto, A.; Petri, A.; Kauppinen, S.; Ingvar, C.; Jönsson, G.; Olsson, H.; et al. PTENP1-AS contributes to BRAF inhibitor resistance and is associated with adverse clinical outcome in stage III melanoma. Sci. Rep. 2021, 11, 11023. [Google Scholar] [CrossRef]
- Oliveira-Mateos, C.; Sánchez-Castillo, A.; Soler, M.; Obiols-Guardia, A.; Piñeyro, D.; Boque-Sastre, R.; Calleja-Cervantes, M.E.; Castro de Moura, M.; Martínez-Cardús, A.; Rubio, T.; et al. The transcribed pseudogene RPSAP52 enhances the oncofetal HMGA2-IGF2BP2-RAS axis through LIN28B-dependent and independent let-7 inhibition. Nat. Commun. 2019, 10, 3979. [Google Scholar] [CrossRef] [Green Version]
- Ni, Q.; An, M.; Luo, S.; Li, X.; He, G.; Shen, M.; Xu, L.; Huang, J.; Yan, M.; Fan, Y.; et al. Pseudogene TDGF1P3 regulates the proliferation and metastasis of colorectal cancer cells via the miR-338-3p–PKM2 axis. Biochem. Biophys. Res. Commun. 2023, 638, 7–13. [Google Scholar] [CrossRef]
- Jin, Z.; Liu, B.; Lin, B.; Yang, R.; Wu, C.; Xue, W.; Zou, X.; Qian, J. The novel lncRNA RP9P promotes colorectal cancer progression by modulating miR-133a-3p/FOXQ1 axis. Front. Oncol. 2022, 12, 843064. [Google Scholar] [CrossRef]
- Li, D.; She, J.; Hu, X.; Zhang, M.; Sun, R.; Qin, S. The ELF3-regulated lncRNA UBE2CP3 is over-stabilized by RNA–RNA interactions and drives gastric cancer metastasis via miR-138-5p/ITGA2 axis. Oncogene 2021, 40, 5403–5415. [Google Scholar] [CrossRef]
- Zhang, G.; Xu, Y.; Zou, C.; Tang, Y.; Lu, J.; Gong, Z.; Ma, G.; Zhang, W.; Jiang, P. Long noncoding RNA ARHGAP27P1 inhibits gastric cancer cell proliferation and cell cycle progression through epigenetically regulating p15 and p16. Aging 2019, 11, 9090. [Google Scholar] [CrossRef]
- Yang, W.; Du, W.; Li, X.; Yee, A.; Yang, B. Foxo3 activity promoted by non-coding effects of circular RNA and Foxo3 pseudogene in the inhibition of tumor growth and angiogenesis. Oncogene 2016, 35, 3919–3931. [Google Scholar] [CrossRef]
- Chan, J.J.; Kwok, Z.H.; Chew, X.H.; Zhang, B.; Liu, C.; Soong, T.W.; Yang, H.; Tay, Y. A FTH1 gene: Pseudogene: MicroRNA network regulates tumorigenesis in prostate cancer. Nucleic Acids Res. 2018, 46, 1998–2011. [Google Scholar] [CrossRef]
- Wu, G.; Sun, P.; Qin, C. GUSBP11 inhibited the progression of triple negative breast cancer via targeting the miR-579-3p/SPNS2 Axis. Cell J. (Yakhteh) 2022, 24, 230–238. [Google Scholar]
- Wang, G.; Dong, Y.; Liu, H.; Ji, N.; Cao, J.; Liu, A.; Tang, X.; Ren, Y. Long noncoding RNA (lncRNA) metallothionein 1 J, pseudogene (MT1JP) is downregulated in triple-negative breast cancer and upregulates microRNA-138 (miR-138) to downregulate hypoxia-inducible factor-1α (HIF-1α). Bioengineered 2022, 13, 13718–13727. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yin, H.; Chen, Y.; Pan, C.; Hang, H.; Lu, Y.; Ma, W.; Li, X.; Gan, W.; Guo, H.; et al. Low expression of PEBP1P2 promotes metastasis of clear cell renal cell carcinoma by post-transcriptional regulation of PEBP1 and KLF13 mRNA. Exp. Hematol. Oncol. 2022, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rutnam, Z.J.; Du, W.W.; Yang, W.; Yang, X.; Yang, B.B. The pseudogene TUSC2P promotes TUSC2 function by binding multiple microRNAs. Nat. Commun. 2014, 5, 2914. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Gong, R.; He, B.; Chen, F.; Hu, Z. TUSC2P suppresses the tumor function of esophageal squamous cell carcinoma by regulating TUSC2 expression and correlates with disease prognosis. BMC Cancer 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Yang, Y.; Han, L.; Yuan, Y.; Li, J.; Hei, N.; Liang, H. Gene co-expression network analysis reveals common system-level properties of prognostic genes across cancer types. Nat. Commun. 2014, 5, 3231. [Google Scholar] [CrossRef] [Green Version]
- Dorantes-Gilardi, R.; García-Cortés, D.; Hernández-Lemus, E.; Espinal-Enríquez, J. Multilayer approach reveals organizational principles disrupted in breast cancer co-expression networks. Appl. Netw. Sci. 2020, 5, 1–23. [Google Scholar] [CrossRef]
- Dorantes-Gilardi, R.; García-Cortés, D.; Hernández-Lemus, E.; Espinal-Enríquez, J. k-core genes underpin structural features of breast cancer. Sci. Rep. 2021, 11, 16284. [Google Scholar] [CrossRef]
- Drago-García, D.; Espinal-Enríquez, J.; Hernández-Lemus, E. Network analysis of EMT and MET micro-RNA regulation in breast cancer. Sci. Rep. 2017, 7, 13534. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Fuentes, J.M.; Hernández-Lemus, E.; Espinal-Enríquez, J. Oncogenic Role of miR-217 During Clear Cell Renal Carcinoma Progression. Front. Oncol. 2022, 12, 934711. [Google Scholar] [CrossRef]
- Alcalá-Corona, S.A.; de Anda-Jáuregui, G.; Espinal-Enríquez, J.; Hernández-Lemus, E. Network modularity in breast cancer molecular subtypes. Front. Physiol. 2017, 8, 915. [Google Scholar] [CrossRef] [Green Version]
- Alcalá-Corona, S.A.; Espinal-Enríquez, J.; de Anda-Jáuregui, G.; Hernández-Lemus, E. The hierarchical modular structure of HER2+ breast cancer network. Front. Physiol. 2018, 9, 1423. [Google Scholar] [CrossRef] [Green Version]
- Alcalá-Corona, S.A.; Sandoval-Motta, S.; Espinal-Enriquez, J.; Hernández-Lemus, E. Modularity in biological networks. Front. Genet. 2021, 12, 701331. [Google Scholar] [CrossRef] [PubMed]
- Carron, J.; Della Coletta, R.; Lourenço, G.J. Pseudogene transcripts in head and neck cancer: Literature review and in silico analysis. Genes 2021, 12, 1254. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.; Xie, D.; Hu, J.; Wu, T.; Cao, K.; Luo, X. Identification of Candidate lncRNA and Pseudogene Biomarkers Associated with Carbon-Nanotube-Induced Malignant Transformation of Lung Cells and Prediction of Potential Preventive Drugs. Int. J. Environ. Res. Public Health 2022, 19, 2936. [Google Scholar] [CrossRef]
- Lin, Q.Y.; Qi, Q.L.; Hou, S.; Chen, Z.; Zhang, L.; Zhao, H.G.; Lin, C.H. LncRNA PVT1 acts as a tumor promoter in thyroid cancer and promotes tumor progression by mediating miR-423-5p-PAK3. Cancer Manag. Res. 2020, 12, 13403. [Google Scholar] [CrossRef]
- Espinal-Enríquez, J.; Fresno, C.; Anda-Jáuregui, G.; Hernández-Lemus, E. RNA-Seq based genome-wide analysis reveals loss of inter-chromosomal regulation in breast cancer. Sci. Rep. 2017, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- García-Cortés, D.; de Anda-Jáuregui, G.; Fresno, C.; Hernández-Lemus, E.; Espinal-Enríquez, J. Gene co-expression is distance-dependent in breast cancer. Front. Oncol. 2020, 10, 1232. [Google Scholar] [CrossRef]
- Zamora-Fuentes, J.M.; Hernández-Lemus, E.; Espinal-Enríquez, J. Gene expression and co-expression networks are strongly altered through stages in clear cell renal carcinoma. Front. Genet. 2020, 11, 578679. [Google Scholar] [CrossRef]
- Andonegui-Elguera, S.D.; Zamora-Fuentes, J.M.; Espinal-Enríquez, J.; Hernández-Lemus, E. Loss of long distance co-expression in lung cancer. Front. Genet. 2021, 12, 625741. [Google Scholar] [CrossRef]
- Nakaramura-Garcia, A.K.; Espinal-Enriquez, J. The network structure of hematopoietic cancers. bioRxiv 2022. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura-García, A.K.; Espinal-Enríquez, J. Pseudogenes in Cancer: State of the Art. Cancers 2023, 15, 4024. https://doi.org/10.3390/cancers15164024
Nakamura-García AK, Espinal-Enríquez J. Pseudogenes in Cancer: State of the Art. Cancers. 2023; 15(16):4024. https://doi.org/10.3390/cancers15164024
Chicago/Turabian StyleNakamura-García, Arturo Kenzuke, and Jesús Espinal-Enríquez. 2023. "Pseudogenes in Cancer: State of the Art" Cancers 15, no. 16: 4024. https://doi.org/10.3390/cancers15164024