
Phenotype and Reactivity of Lymphocytes Expanded from Benign Prostate Hyperplasic Tissues and Prostate Cancer
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Antibodies and Reagents
2.3. Expansion of Prostate Infiltrating Lymphocytes
2.4. Cytokine Production Assay and Intracellular Staining
2.5. PILs and Autologous/Heterologous Tissue Co-Cultures
2.6. Statistical Analysis
3. Results
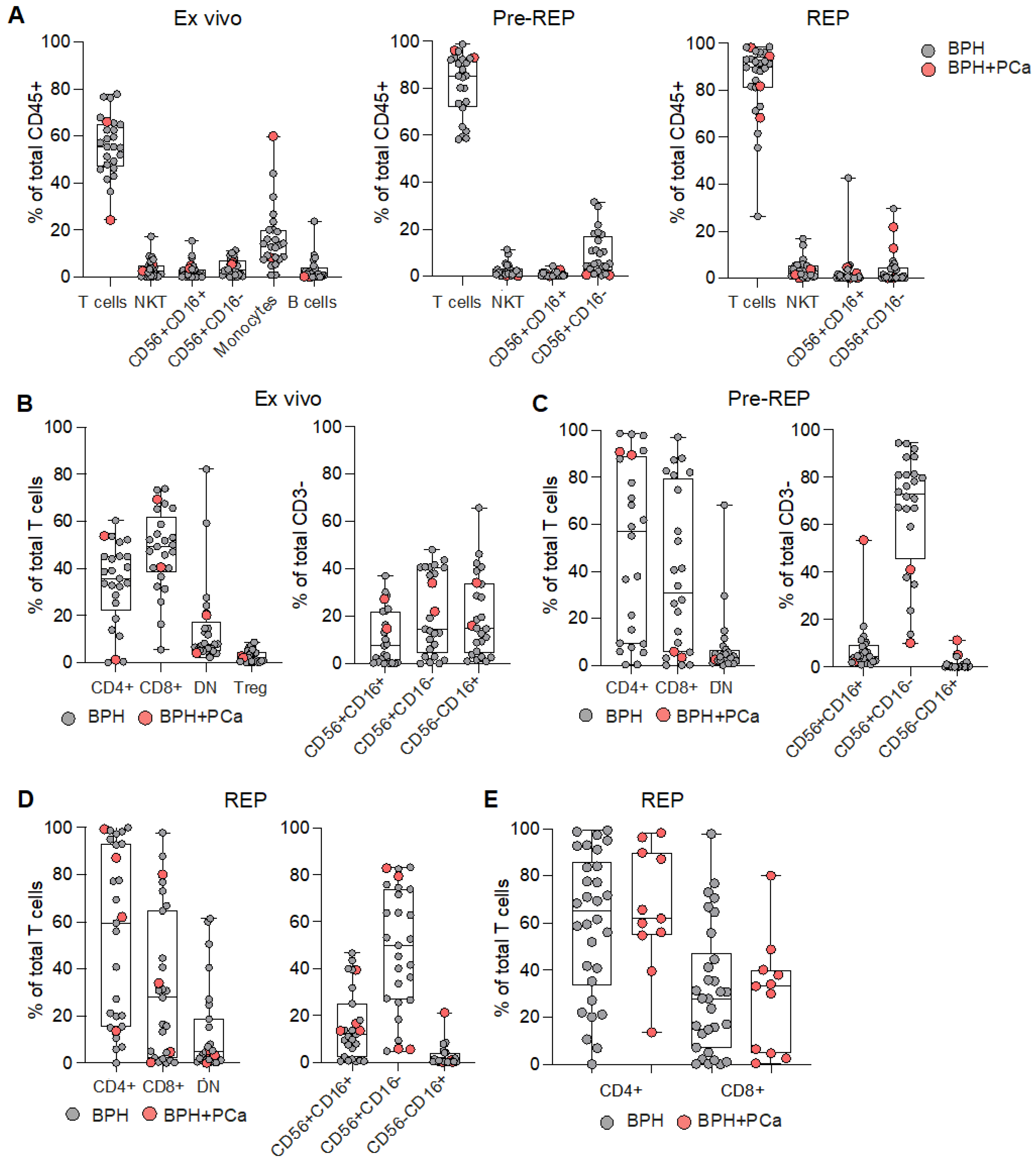
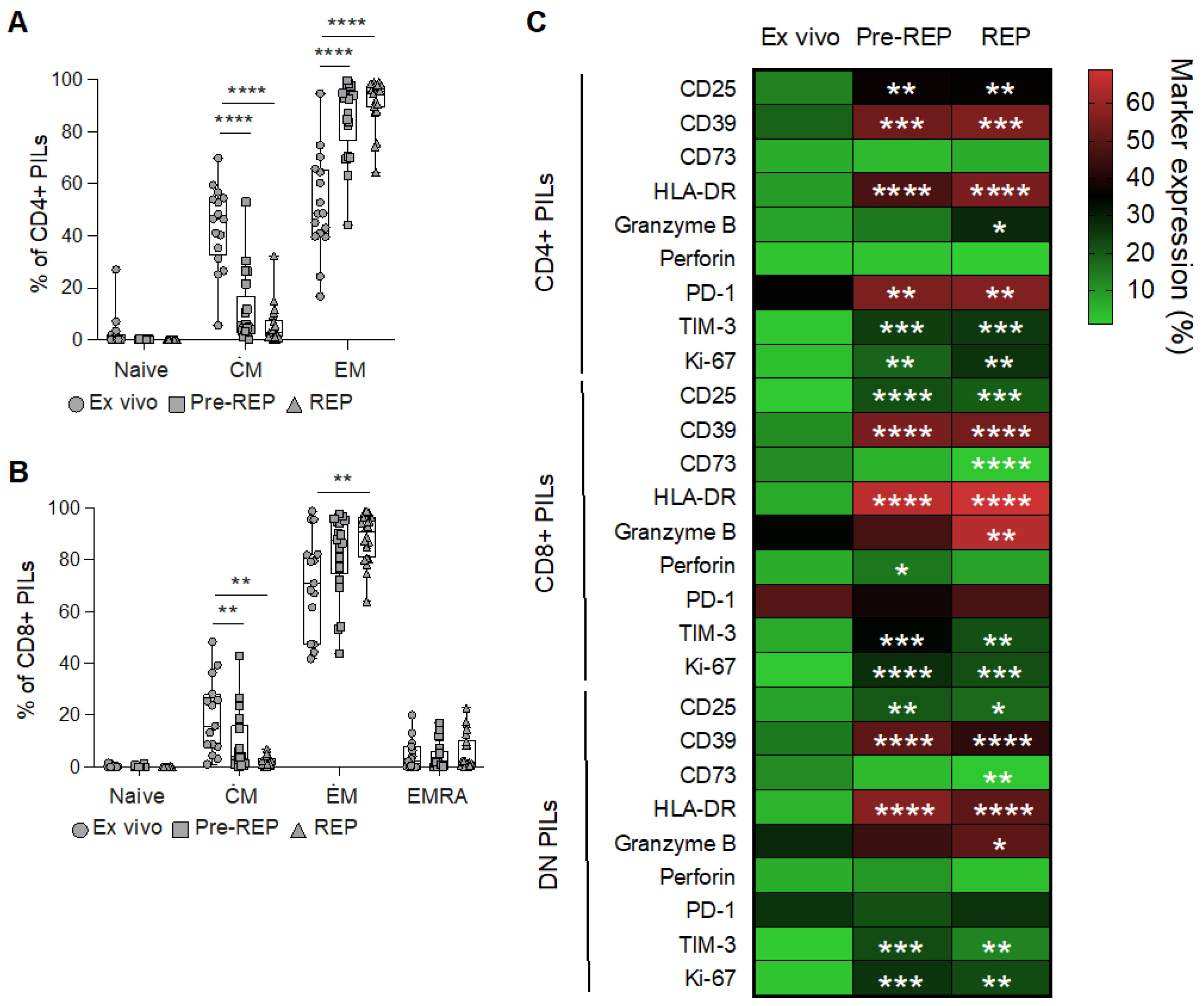
3.1. Phenotypic Characterization of Prostate Infiltrating Lymphocytes from BPH Tissue Samples
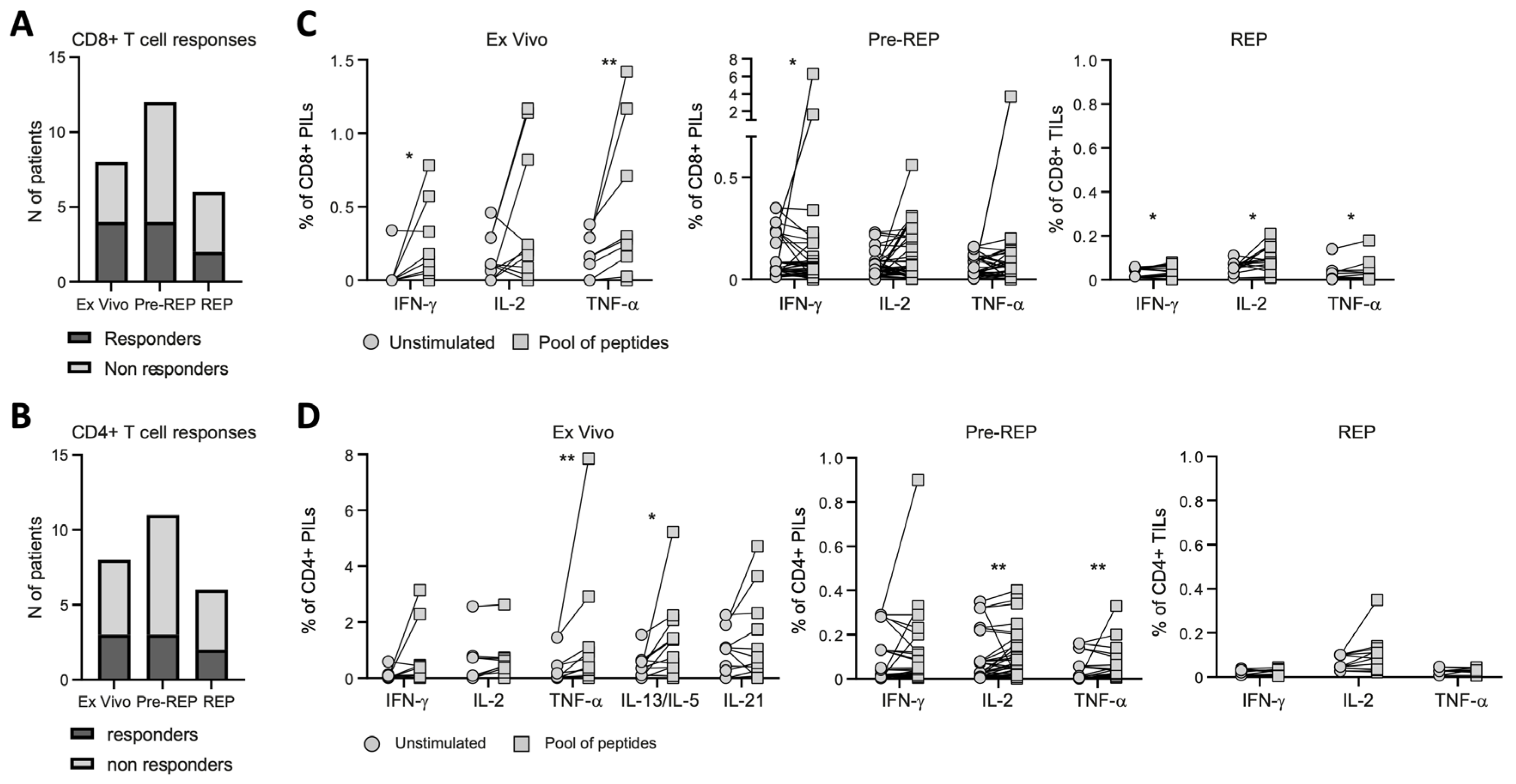
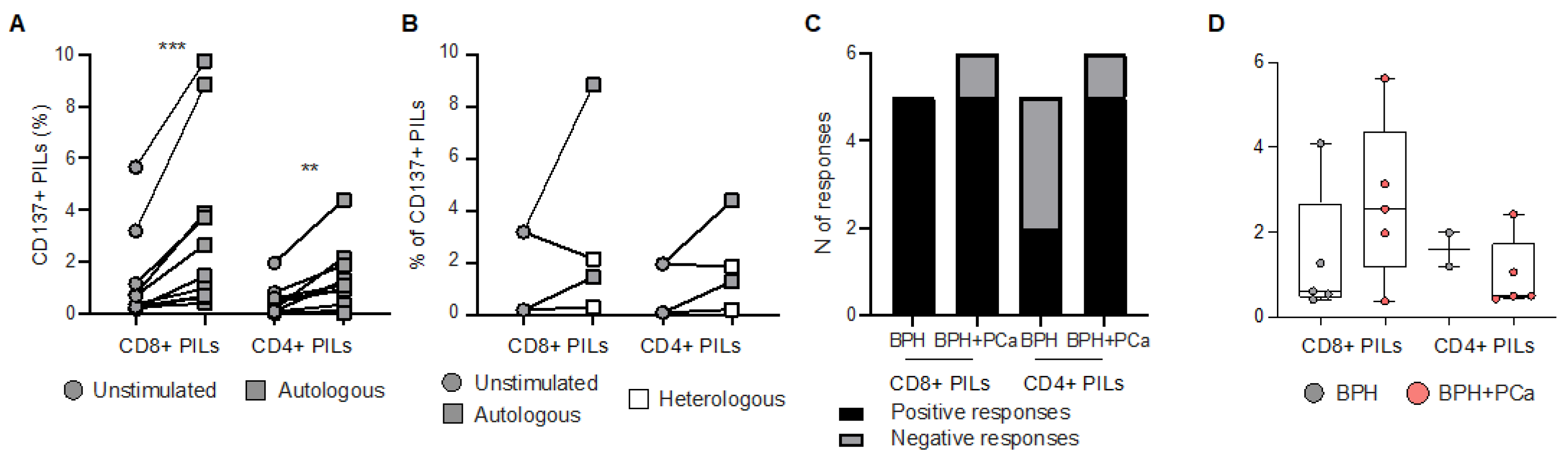
3.2. Reactivity of Prostate Infiltrating Lymphocytes from BPH Tissue Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orsted, D.D.; Bojesen, S.E. The link between benign prostatic hyperplasia and prostate cancer. Nat. Rev. Urol. 2013, 10, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Dunn, M.W.; Kazer, M.W. Prostate cancer overview. Semin. Oncol Nurs 2011, 27, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Ning, H.; Sun, Y.; Wu, H.; Lyu, J. Integrative exploration of the mutual gene signatures and immune microenvironment between benign prostate hyperplasia and castration-resistant prostate cancer. Aging Male 2023, 26, 2183947. [Google Scholar] [CrossRef] [PubMed]
- De Marzo, A.M.; Platz, E.A.; Sutcliffe, S.; Xu, J.; Gronberg, H.; Drake, C.G.; Nakai, Y.; Isaacs, W.B.; Nelson, W.G. Inflammation in prostate carcinogenesis. Nat. Rev. Cancer 2007, 7, 256–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitvogel, L.; Pietrocola, F.; Kroemer, G. Nutrition, inflammation and cancer. Nat. Immunol. 2017, 18, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Keil, K.P.; Vezina, C.M. DNA methylation as a dynamic regulator of development and disease processes: Spotlight on the prostate. Epigenomics 2015, 7, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Shoag, J.E.; Poliak, D.; Goueli, R.S.; Ravikumar, V.; Redmond, D.; Vosoughi, A.; Fontugne, J.; Pan, H.; Lee, D.; et al. Integrative multiplatform molecular profiling of benign prostatic hyperplasia identifies distinct subtypes. Nat. Commun. 2020, 11, 1987. [Google Scholar] [CrossRef] [Green Version]
- Kruslin, B.; Tomas, D.; Dzombeta, T.; Milkovic-Perisa, M.; Ulamec, M. Inflammation in Prostatic Hyperplasia and Carcinoma-Basic Scientific Approach. Front. Oncol. 2017, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Sun, R.; Peng, L.; Li, J.; Huang, Y.; Chen, Z.; Chen, B.; Li, J.; Ai, J.; Yang, L.; et al. Immune Cell Proinflammatory Microenvironment and Androgen-Related Metabolic Regulation During Benign Prostatic Hyperplasia in Aging. Front. Immunol. 2022, 13, 842008. [Google Scholar] [CrossRef]
- Radestad, E.; Egevad, L.; Jorns, C.; Mattsson, J.; Sundberg, B.; Nava, S.; Ericzon, B.G.; Henningsohn, L.; Levitsky, V.; Uhlin, M. Characterization of infiltrating lymphocytes in human benign and malignant prostate tissue. Oncotarget 2017, 8, 60257–60269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, M.R.; Al-Assiri, M.; Musalam, A.O. Phenotypic characterization of the infiltrating immune cells in normal prostate, benign nodular prostatic hyperplasia and prostatic adenocarcinoma. Exp. Mol. Pathol. 2009, 86, 108–113. [Google Scholar] [CrossRef] [PubMed]
- De Nunzio, C.; Presicce, F.; Tubaro, A. Inflammatory mediators in the development and progression of benign prostatic hyperplasia. Nat. Rev. Urol. 2016, 13, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Naiyila, X.; Li, J.; Huang, Y.; Chen, B.; Zhu, M.; Li, J.; Chen, Z.; Yang, L.; Ai, J.; Wei, Q.; et al. A Novel Insight into the Immune-Related Interaction of Inflammatory Cytokines in Benign Prostatic Hyperplasia. J. Clin. Med. 2023, 12, 1821. [Google Scholar] [CrossRef]
- Yunger, S.; Bar El, A.; Zeltzer, L.A.; Fridman, E.; Raviv, G.; Laufer, M.; Schachter, J.; Markel, G.; Itzhaki, O.; Besser, M.J. Tumor-infiltrating lymphocytes from human prostate tumors reveal anti-tumor reactivity and potential for adoptive cell therapy. Oncoimmunology 2019, 8, e1672494. [Google Scholar] [CrossRef] [Green Version]
- Westdorp, H.; Skold, A.E.; Snijer, B.A.; Franik, S.; Mulder, S.F.; Major, P.P.; Foley, R.; Gerritsen, W.R.; de Vries, I.J. Immunotherapy for prostate cancer: Lessons from responses to tumor-associated antigens. Front. Immunol. 2014, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Nyberg, T.; Govindasami, K.; Leslie, G.; Dadaev, T.; Bancroft, E.; Ni Raghallaigh, H.; Brook, M.N.; Hussain, N.; Keating, D.; Lee, A.; et al. Homeobox B13 G84E Mutation and Prostate Cancer Risk. Eur. Urol. 2019, 75, 834–845. [Google Scholar] [CrossRef]
- Middleton, L.W.; Shen, Z.; Varma, S.; Pollack, A.S.; Gong, X.; Zhu, S.; Zhu, C.; Foley, J.W.; Vennam, S.; Sweeney, R.T.; et al. Genomic analysis of benign prostatic hyperplasia implicates cellular re-landscaping in disease pathogenesis. JCI Insight 2019, 5, 5. [Google Scholar] [CrossRef]
- Bobisse, S.; Genolet, R.; Roberti, A.; Tanyi, J.L.; Racle, J.; Stevenson, B.J.; Iseli, C.; Michel, A.; Le Bitoux, M.A.; Guillaume, P.; et al. Sensitive and frequent identification of high avidity neo-epitope specific CD8(+) T cells in immunotherapy-naive ovarian cancer. Nat. Commun. 2018, 9, 1092. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, M.F.; Bobisse, S.; Costa-Nunes, C.; Cesson, V.; Jichlinski, P.; Speiser, D.E.; Harari, A.; Coukos, G.; Romero, P.; Nardelli-Haefliger, D.; et al. High-throughput monitoring of human tumor-specific T-cell responses with large peptide pools. Oncoimmunology 2015, 4, e1029702. [Google Scholar] [CrossRef] [Green Version]
- Parkhurst, M.; Gros, A.; Pasetto, A.; Prickett, T.; Crystal, J.S.; Robbins, P.; Rosenberg, S.A. Isolation of T-Cell Receptors Specifically Reactive with Mutated Tumor-Associated Antigens from Tumor-Infiltrating Lymphocytes Based on CD137 Expression. Clin. Cancer Res. 2017, 23, 2491–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Q.; Song, D.G.; Poussin, M.; Yamamoto, T.; Best, A.; Li, C.; Coukos, G.; Powell, D.J., Jr. CD137 accurately identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin. Cancer Res. 2014, 20, 44–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geginat, J.; Lanzavecchia, A.; Sallusto, F. Proliferation and differentiation potential of human CD8+ memory T-cell subsets in response to antigen or homeostatic cytokines. Blood 2003, 101, 4260–4266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebelt, K.; Babaryka, G.; Frankenberger, B.; Stief, C.G.; Eisenmenger, W.; Kirchner, T.; Schendel, D.J.; Noessner, E. Prostate cancer lesions are surrounded by FOXP3+, PD-1+ and B7-H1+ lymphocyte clusters. Eur. J. Cancer 2009, 45, 1664–1672. [Google Scholar] [CrossRef]
- He, J.; Yi, M.; Tan, L.; Huang, J.; Huang, L. The immune checkpoint regulator PD-L1 expression are associated with clinical progression in prostate cancer. World J. Surg. Oncol. 2021, 19, 215. [Google Scholar] [CrossRef] [PubMed]
- Sarma, A.V.; Wei, J.T. Clinical practice. Benign prostatic hyperplasia and lower urinary tract symptoms. N. Engl. J. Med. 2012, 367, 248–257. [Google Scholar] [CrossRef]
- Bechis, S.K.; Otsetov, A.G.; Ge, R.; Olumi, A.F. Personalized medicine for the management of benign prostatic hyperplasia. J. Urol. 2014, 192, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Isaacsson Velho, P.; Antonarakis, E.S. PD-1/PD-L1 pathway inhibitors in advanced prostate cancer. Expert Rev. Clin. Pharmacol. 2018, 11, 475–486. [Google Scholar] [CrossRef]
- Vitkin, N.; Nersesian, S.; Siemens, D.R.; Koti, M. The Tumor Immune Contexture of Prostate Cancer. Front. Immunol. 2019, 10, 603. [Google Scholar] [CrossRef] [Green Version]
- Jafari, S.; Molavi, O.; Kahroba, H.; Hejazi, M.S.; Maleki-Dizaji, N.; Barghi, S.; Kiaie, S.H.; Jadidi-Niaragh, F. Clinical application of immune checkpoints in targeted immunotherapy of prostate cancer. Cell. Mol. Life Sci. 2020, 77, 3693–3710. [Google Scholar] [CrossRef]
- Blass, E.; Ott, P.A. Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat. Rev. Clin. Oncol. 2021, 18, 215–229. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BPH | BPH+PCa | |
|---|---|---|
| Number of patients | 44 | 12 |
| Age, yr., median (IQR) | 70 (66–77) | 79 (70.75–82.75) |
| Concomitant chronic prostatitis | 20 | 2 |
| Gleason score | N/A | |
| 6 | 4 | |
| 7–8 | 3 | |
| ≥9 | 5 | |
| Grade | N/A | |
| 1–2 | 5 | |
| 3–4 | 2 | |
| 5 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, R.; Lozano, L.E.; Anastasio, A.; Lofek, S.; Mastelic-Gavillet, B.; Navarro Rodrigo, B.; Nguyen, S.; Dartiguenave, F.; Rodrigues-Dias, S.-C.; Cesson, V.; et al. Phenotype and Reactivity of Lymphocytes Expanded from Benign Prostate Hyperplasic Tissues and Prostate Cancer. Cancers 2023, 15, 3114. https://doi.org/10.3390/cancers15123114
Ahmed R, Lozano LE, Anastasio A, Lofek S, Mastelic-Gavillet B, Navarro Rodrigo B, Nguyen S, Dartiguenave F, Rodrigues-Dias S-C, Cesson V, et al. Phenotype and Reactivity of Lymphocytes Expanded from Benign Prostate Hyperplasic Tissues and Prostate Cancer. Cancers. 2023; 15(12):3114. https://doi.org/10.3390/cancers15123114
Chicago/Turabian StyleAhmed, Ritaparna, Leyder Elena Lozano, Amandine Anastasio, Sebastien Lofek, Beatris Mastelic-Gavillet, Blanca Navarro Rodrigo, Sylvain Nguyen, Florence Dartiguenave, Sonia-Cristina Rodrigues-Dias, Valérie Cesson, and et al. 2023. "Phenotype and Reactivity of Lymphocytes Expanded from Benign Prostate Hyperplasic Tissues and Prostate Cancer" Cancers 15, no. 12: 3114. https://doi.org/10.3390/cancers15123114