
Pim-2 Kinase Regulates Energy Metabolism in Multiple Myeloma
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Information
2.2. Cell Culture
2.3. Metabolomic Analysis
2.4. Cell Viability Assay
2.5. Apoptosis Assay
2.6. 5-Ethynyl-2-Deoxyuridine (EdU) Assay
2.7. ATP Assay
2.8. Reactive Oxygen Species (ROS) Assay
2.9. Mitochondrial Mass Assay
2.10. Measurement of Mitochondrial Oxygen Consumption Rate (OCR)
2.11. Glycolysis Assay
2.12. SiRNA Transfection to Knock down Pim-2
2.13. Western Blotting Analysis
2.14. Statistical Analysis
3. Results
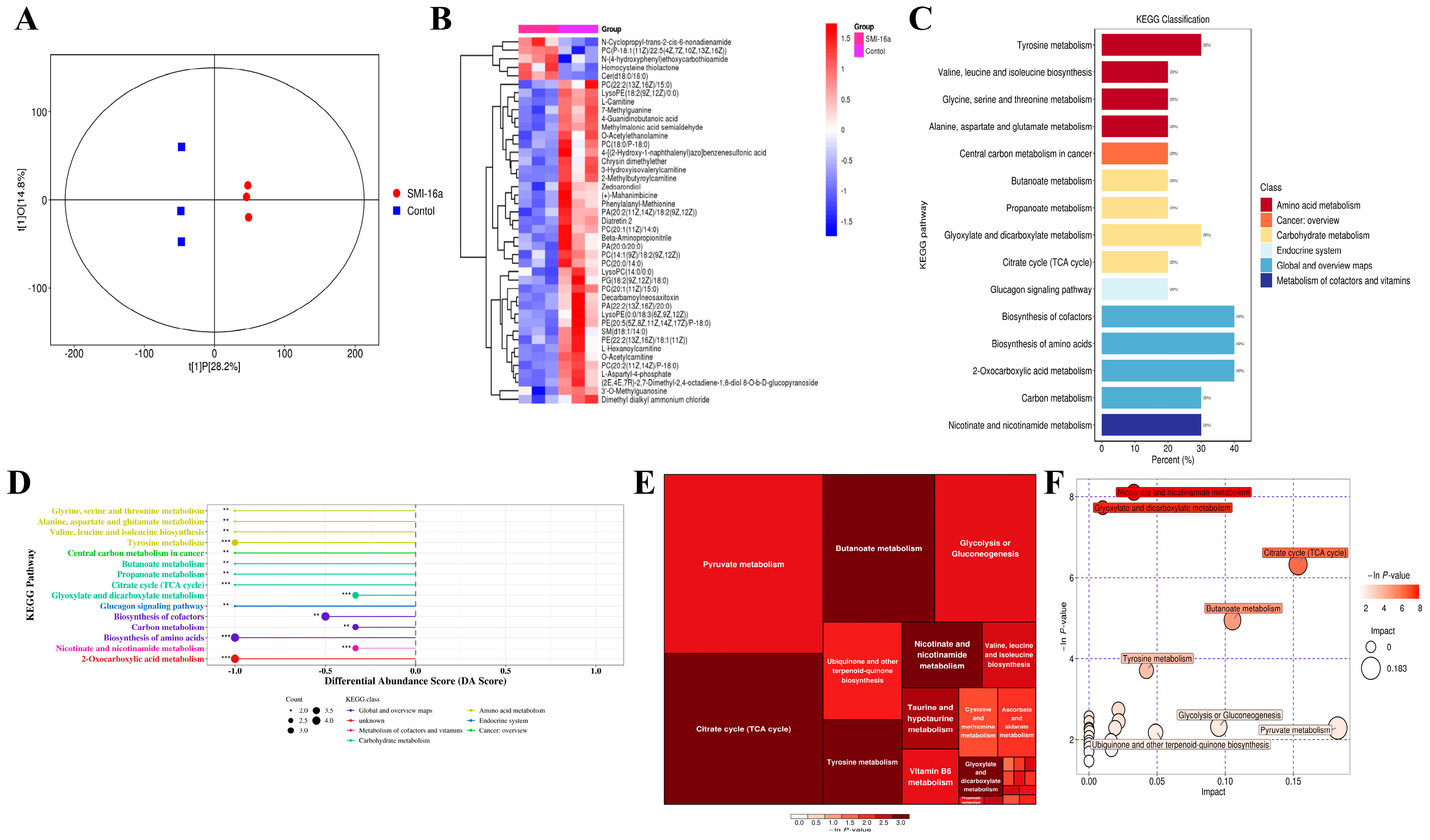
3.1. High Expression of Pim-2 Kinase Correlates with Poor Prognosis and Genes Involved in Energy Metabolism in MM Patients
3.2. Pim-2 Kinase Inhibitors Inhibit Glycometabolism in MM
3.3. Pim-2 Kinase Inhibitor Participates in the Regulation of MM Cell Proliferation and Apoptosis
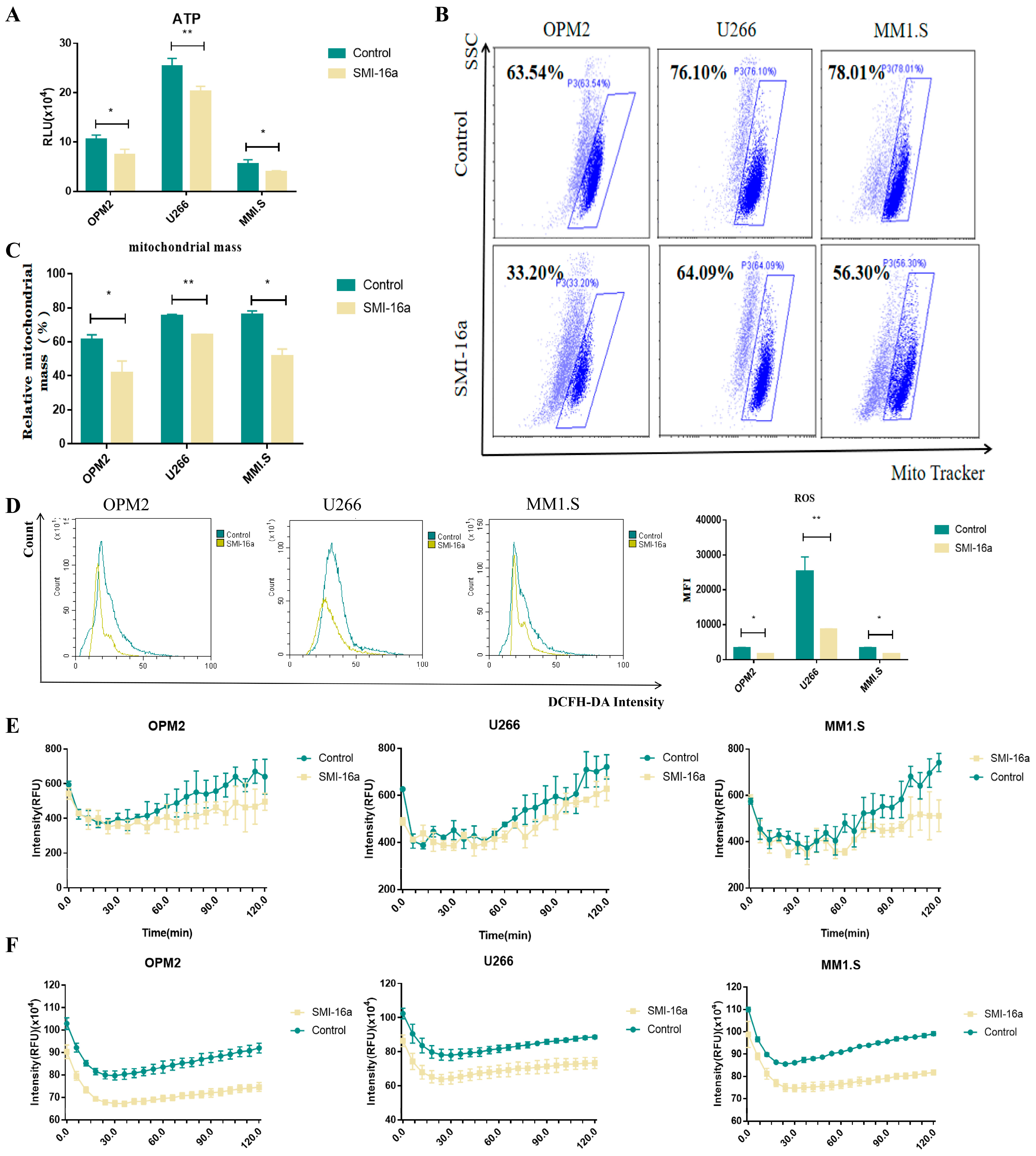
3.4. Inhibition of Pim-2 Reduces Cellular Glycometabolism and Energy Production
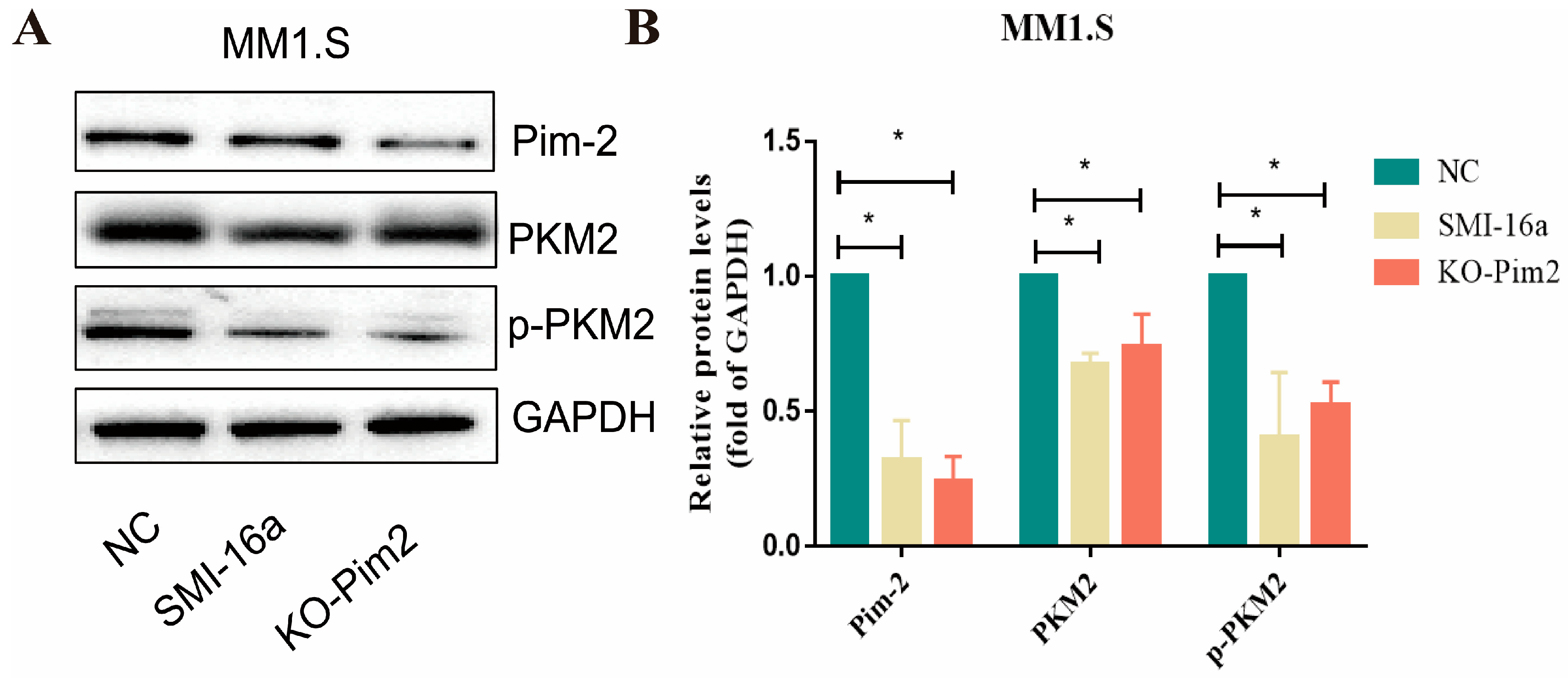
3.5. Pim-2 Negatively Regulates PKM2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Keane, N.A.; Reidy, M.; Natoni, A.; Raab, M.S.; O’Dwyer, M. Targeting the Pim kinases in multiple myeloma. Blood Cancer J. 2015, 5, e325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Han, M.; Ding, K.; Fu, R. The role of Pim kinase in immunomodulation. Am. J. Cancer Res. 2020, 10, 4085–4097. [Google Scholar] [PubMed]
- Szydlowski, M.; Garbicz, F.; Jablonska, E.; Gorniak, P.; Komar, D.; Pyrzynska, B.; Bojarczuk, K.; Prochorec-Sobieszek, M.; Szumera-Cieckiewicz, A.; Rymkiewicz, G.; et al. Inhibition of PIM Kinases in DLBCL Targets MYC Transcriptional Program and Augments the Efficacy of Anti-CD20 Antibodies. Cancer Res. 2021, 81, 6029–6043. [Google Scholar] [CrossRef]
- Zhang, X.; Song, M.; Kundu, J.K.; Lee, M.H.; Liu, Z.Z. PIM Kinase as an Executional Target in Cancer. J. Cancer Prev. 2018, 23, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xiu, J.; Ren, C.; Yu, Z. Protein kinase PIM2: A simple PIM family kinase with complex functions in cancer metabolism and therapeutics. J. Cancer 2021, 12, 2570–2581. [Google Scholar] [CrossRef]
- Cowan, A.J.; Green, D.J.; Kwok, M.; Lee, S.; Coffey, D.G.; Holmberg, L.A.; Tuazon, S.; Gopal, A.K.; Libby, E.N. Diagnosis and Management of Multiple Myeloma: A Review. JAMA 2022, 327, 464–477. [Google Scholar] [CrossRef]
- Kim, S.D.; Kim, H.N.; Lee, J.H.; Jin, W.J.; Hwang, S.J.; Kim, H.H.; Ha, H.; Lee, Z.H. Trapidil, a platelet-derived growth factor antagonist, inhibits osteoclastogenesis by down-regulating NFATc1 and suppresses bone loss in mice. Biochem. Pharmacol. 2013, 86, 782–790. [Google Scholar] [CrossRef]
- Paíno, T.; Garcia-Gomez, A.; González-Méndez, L.; San-Segundo, L.; Hernández-García, S.; López-Iglesias, A.-A.; Algarín, E.M.; Martín-Sánchez, M.; Corbacho, D.; Ortiz-de-Solorzano, C.; et al. The Novel Pan-PIM Kinase Inhibitor, PIM447, Displays Dual Antimyeloma and Bone-Protective Effects, and Potently Synergizes with Current Standards of Care. Clin. Cancer Res. 2017, 23, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Johrer, K.; Obkircher, M.; Neureiter, D.; Parteli, J.; Zelle-Rieser, C.; Maizner, E.; Kern, J.; Hermann, M.; Hamacher, F.; Merkel, O.; et al. Antimyeloma activity of the sesquiterpene lactone cnicin: Impact on Pim-2 kinase as a novel therapeutic target. J. Mol. Med. 2012, 90, 681–693. [Google Scholar] [CrossRef]
- Claudio, J.O.; Masih-Khan, E.; Tang, H.; Goncalves, J.; Voralia, M.; Li, Z.H.; Nadeem, V.; Cukerman, E.; Francisco-Pabalan, O.; Liew, C.C.; et al. A molecular compendium of genes expressed in multiple myeloma. Blood 2002, 100, 2175–2186. [Google Scholar] [CrossRef]
- Lu, J.; Zavorotinskaya, T.; Dai, Y.; Niu, X.H.; Castillo, J.; Sim, J.; Yu, J.; Wang, Y.; Langowski, J.L.; Holash, J.; et al. Pim2 is required for maintaining multiple myeloma cell growth through modulating TSC2 phosphorylation. Blood 2013, 122, 1610–1620. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, H.; Yuan, X.; Wang, Y.; Li, L.; Wang, G.; Song, J.; Shao, Z.; Fu, R. Downregulation of Pim-2 induces cell cycle arrest in the G0/G1 phase via the p53-non-dependent p21 signaling pathway. Oncol. Lett. 2018, 15, 4079–4086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishita, D.; Katayama, R.; Sekimizu, K.; Tsuruo, T.; Fujita, N. Pim kinases promote cell cycle progression by phosphorylating and down-regulating p27Kip1 at the transcriptional and posttranscriptional levels. Cancer Res. 2008, 68, 5076–5085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, R.; Xie, Y.; Sun, X.; Zhang, M.; Zhou, J.; Liu, L.; Gao, J.; Xu, K. Identification of a Novel c-Myc Inhibitor 7594-0037 by Structure-Based Virtual Screening and Investigation of Its Anti-Cancer Effect on Multiple Myeloma. Drug Des. Dev. Ther. 2020, 14, 3983–3993. [Google Scholar] [CrossRef] [PubMed]
- Abe, M. Myeloma bone disease. Clin. Calcium 2014, 24, 1159–1168. [Google Scholar]
- Hiasa, M.; Teramachi, J.; Oda, A.; Amachi, R.; Harada, T.; Nakamura, S.; Miki, H.; Fujii, S.; Kagawa, K.; Watanabe, K.; et al. Pim-2 kinase is an important target of treatment for tumor progression and bone loss in myeloma. Leukemia 2015, 29, 207–217. [Google Scholar] [CrossRef]
- Luo, W.; Semenza, G.L. Emerging roles of PKM2 in cell metabolism and cancer progression. Trends Endocrinol. Metab. 2012, 23, 560–566. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Hu, H.; Chang, R.; Zhong, J.; Knabel, M.; O’Meally, R.; Cole, R.N.; Pandey, A.; Semenza, G.L. Pyruvate kinase M2 is a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell 2011, 145, 732–744. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, M.; Hennemann, H.; Xing, P.X.; Hoffmann, I.; Moroy, T. The oncogenic serine/threonine kinase Pim-1 phosphorylates and inhibits the activity of Cdc25C-associated kinase 1 (C-TAK1): A novel role for Pim-1 at the G2/M cell cycle checkpoint. J. Biol. Chem. 2004, 279, 48319–48328. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.S.; Balakrishnan, K.; Gandhi, V. Inflammation and survival pathways: Chronic lymphocytic leukemia as a model system. Biochem. Pharmacol. 2010, 80, 1936–1945. [Google Scholar] [CrossRef] [Green Version]
- Mondello, P.; Cuzzocrea, S.; Mian, M. Pim kinases in hematological malignancies: Where are we now and where are we going? J. Hematol. Oncol. 2014, 7, 95. [Google Scholar] [CrossRef]
- Yan, B.; Zemskova, M.; Holder, S.; Chin, V.; Kraft, A.; Koskinen, P.J.; Lilly, M. The PIM-2 kinase phosphorylates BAD on serine 112 and reverses BAD-induced cell death. J. Biol. Chem. 2003, 278, 45358–45367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, J.R.; Caserta, J.; Belko, K.; Howell, T.; Fetterly, G.; Baldino, C.; Lee, K.P. Novel inhibition of PIM2 kinase has significant anti-tumor efficacy in multiple myeloma. Leukemia 2017, 31, 1715–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnan, V.G.; Kumar, S.K. Inhibitors of the Cyclin-Dependent Kinase and PIM Kinase Pathways in the Treatment of Myeloma. Cancer J. 2016, 22, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Nakamura, S.; Oda, A.; Miki, H.; Tenshin, H.; Teramachi, J.; Hiasa, M.; Bat-Erdene, A.; Maeda, Y.; Oura, M.; et al. Unique anti-myeloma activity by thiazolidine-2,4-dione compounds with Pim inhibiting activity. Br. J. Haematol. 2018, 180, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koblish, H.; Li, Y.L.; Shin, N.; Hall, L.; Wang, Q.; Wang, K.; Covington, M.; Marando, C.; Bowman, K.; Boer, J.; et al. Preclinical characterization of INCB053914, a novel pan-PIM kinase inhibitor, alone and in combination with anticancer agents, in models of hematologic malignancies. PLoS ONE 2018, 13, e0199108. [Google Scholar] [CrossRef] [Green Version]
- Adam, K.; Lambert, M.; Lestang, E.; Champenois, G.; Dusanter-Fourt, I.; Tamburini, J.; Bouscary, D.; Lacombe, C.; Zermati, Y.; Mayeux, P. Control of Pim2 kinase stability and expression in transformed human haematopoietic cells. Biosci. Rep. 2015, 35, e00274. [Google Scholar] [CrossRef] [Green Version]
- Cortes, J.; Tamura, K.; DeAngelo, D.J.; de Bono, J.; Lorente, D.; Minden, M.; Uy, G.L.; Kantarjian, H.; Chen, L.S.; Gandhi, V.; et al. Phase I studies of AZD1208, a proviral integration Moloney virus kinase inhibitor in solid and haematological cancers. Br. J. Cancer 2018, 118, 1425–1433. [Google Scholar] [CrossRef]
- He, X.; Du, S.; Lei, T.; Li, X.; Liu, Y.; Wang, H.; Tong, R.; Wang, Y. PKM2 in carcinogenesis and oncotherapy. Oncotarget 2017, 8, 110656. [Google Scholar] [CrossRef]
- Alquraishi, M.; Puckett, D.L.; Alani, D.S.; Humidat, A.S.; Frankel, V.D.; Donohoe, D.R.; Whelan, J.; Bettaieb, A. Pyruvate kinase M2: A simple molecule with complex functions. Free. Radic. Biol. Med. 2019, 143, 176–192. [Google Scholar] [CrossRef]
- Lu, C.; Qiao, P.; Sun, Y.; Ren, C.; Yu, Z. Positive regulation of PFKFB3 by PIM2 promotes glycolysis and paclitaxel resistance in breast cancer. Clin. Transl. Med. 2021, 11, e400. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhao, X.; Huang, L.; Zhang, T.; Yang, F.; Xie, L.; Song, S.; Miao, P.; Zhao, L.; Sun, X.; et al. Proviral insertion in murine lymphomas 2 (PIM2) oncogene phosphorylates pyruvate kinase M2 (PKM2) and promotes glycolysis in cancer cells. J. Biol. Chem. 2013, 288, 35406–35416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Ren, C.; Yang, T.; Qiao, P.; Wang, L.; Jiang, A.; Meng, Y.; Liu, Z.; Du, Y.; Yu, Z. Negative regulation of AMPKalpha1 by PIM2 promotes aerobic glycolysis and tumorigenesis in endometrial cancer. Oncogene 2019, 38, 6537–6549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Yu, H.L.; Wang, F.J.; Han, Y.L.; Yang, W.L. Pim-2 Modulates Aerobic Glycolysis and Energy Production during the Development of Colorectal Tumors. Int. J. Med. Sci. 2015, 12, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Zong, W.X.; Rabinowitz, J.D.; White, E. Mitochondria and Cancer. Mol. Cell 2016, 61, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yu, G.; Chu, H.; Wang, X.; Xiong, L.; Cai, G.; Liu, R.; Gao, H.; Tao, B.; Li, W.; et al. Macrophage-Associated PGK1 Phosphorylation Promotes Aerobic Glycolysis and Tumorigenesis. Mol. Cell 2018, 71, 201–215.e207. [Google Scholar] [CrossRef] [Green Version]
- Maiso, P.; Huynh, D.; Moschetta, M.; Sacco, A.; Aljawai, Y.; Mishima, Y.; Asara, J.M.; Roccaro, A.M.; Kimmelman, A.C.; Ghobrial, I.M. Metabolic signature identifies novel targets for drug resistance in multiple myeloma. Cancer Res. 2015, 75, 2071–2082. [Google Scholar] [CrossRef] [Green Version]
- Zaal, E.A.; Wu, W.; Jansen, G.; Zweegman, S.; Cloos, J.; Berkers, C.R. Bortezomib resistance in multiple myeloma is associated with increased serine synthesis. Cancer Metab. 2017, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Abdollahi, P.; Vandsemb, E.N.; Elsaadi, S.; Rost, L.M.; Yang, R.; Hjort, M.A.; Andreassen, T.; Misund, K.; Slordahl, T.S.; Ro, T.B.; et al. Phosphatase of regenerating liver-3 regulates cancer cell metabolism in multiple myeloma. FASEB J. 2021, 35, e21344. [Google Scholar] [CrossRef]
- Cheng, Y.; Sun, F.; Thornton, K.; Jing, X.; Dong, J.; Yun, G.; Pisano, M.; Zhan, F.; Kim, S.H.; Katzenellenbogen, J.A.; et al. FOXM1 regulates glycolysis and energy production in multiple myeloma. Oncogene 2022, 41, 3899–3911. [Google Scholar] [CrossRef]
- Wu, X.; Guo, J.; Chen, Y.; Liu, X.; Yang, G.; Wu, Y.; Tian, Y.; Liu, N.; Yang, L.; Wei, S.; et al. The 60-kDa heat shock protein regulates energy rearrangement and protein synthesis to promote proliferation of multiple myeloma cells. Br. J. Haematol. 2020, 190, 741–752. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | MM (n = 23) | Control (n = 30) | p Value |
|---|---|---|---|
| Age | 64.69 ± 8.63 | 47.57 ± 13.62 | 0.1079 |
| Female gender (%) | 13 (56.52) | 4 (57.14) | 0.9778 |
| Hemoglobin | 68 ± 17.22 | 89.26 ± 30.99 | 0.7342 |
| Serum calcium | 2.26 ± 0.16 | 2.38 ± 0.36 | 0.3823 |
| Creatinine | 59.43 ± 24.90 | 214.78 ± 230.08 | <0.0001 |
| Albumin | 37.17 ± 2.40 | 32.91 ± 5.80 | 0.0934 |
| Lactate dehydrogenase | 187.33 ± 54.42 | 223.11 ± 142.89 | 0.5566 |
| Label | High Pim-2 Expression Group (Mean ± Std) | Low Pim-2 Expression Group (Mean ± Std) | p Value |
|---|---|---|---|
| PDHA1 | 1 (3.61 ± 0.39) | 0 (3.45 ± 0.39) | <0.001 |
| DLAT | 1 (3.49 ± 0.55) | 0 (3.33 ± 0.66) | <0.001 |
| CS | 1 (4.42 ± 0.36) | 0 (4.44 ± 0.37) | 0.87 |
| OGDH | 1 (4.55 ± 0.61) | 0 (4.38 ± 0.73) | <0.001 |
| DLST | 1 (3.95 ± 0.46) | 0 (4.08 ± 0.48) | <0.001 |
| DLD | 1 (3.17 ± 0.45) | 0 (3.06 ± 0.48) | <0.001 |
| IDH2 | 1 (6.49 ± 0.71) | 0 (6.32 ± 0.84) | <0.05 |
| SDHB | 1 (4.51 ± 0.35) | 0 (4.35 ± 0.34) | <0.001 |
| FH | 1 (4.94 ± 0.67) | 0 (4.59 ± 0.68) | <0.001 |
| MDH2 | 1 (5.12 ± 0.40) | 0 (5.02 ± 0.45) | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Guo, Y.; Liu, X.; Cao, P.; Liu, H.; Dong, X.; Ding, K.; Fu, R. Pim-2 Kinase Regulates Energy Metabolism in Multiple Myeloma. Cancers 2023, 15, 67. https://doi.org/10.3390/cancers15010067
Liu Z, Guo Y, Liu X, Cao P, Liu H, Dong X, Ding K, Fu R. Pim-2 Kinase Regulates Energy Metabolism in Multiple Myeloma. Cancers. 2023; 15(1):67. https://doi.org/10.3390/cancers15010067
Chicago/Turabian StyleLiu, Zhaoyun, Yixuan Guo, Xiaohan Liu, Panpan Cao, Hui Liu, Xifeng Dong, Kai Ding, and Rong Fu. 2023. "Pim-2 Kinase Regulates Energy Metabolism in Multiple Myeloma" Cancers 15, no. 1: 67. https://doi.org/10.3390/cancers15010067