
The Role of CXC Chemokines in Cancer Progression


Abstract
:Simple Summary
Abstract
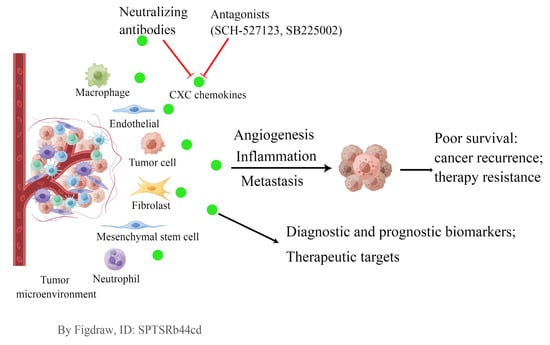
1. Introduction
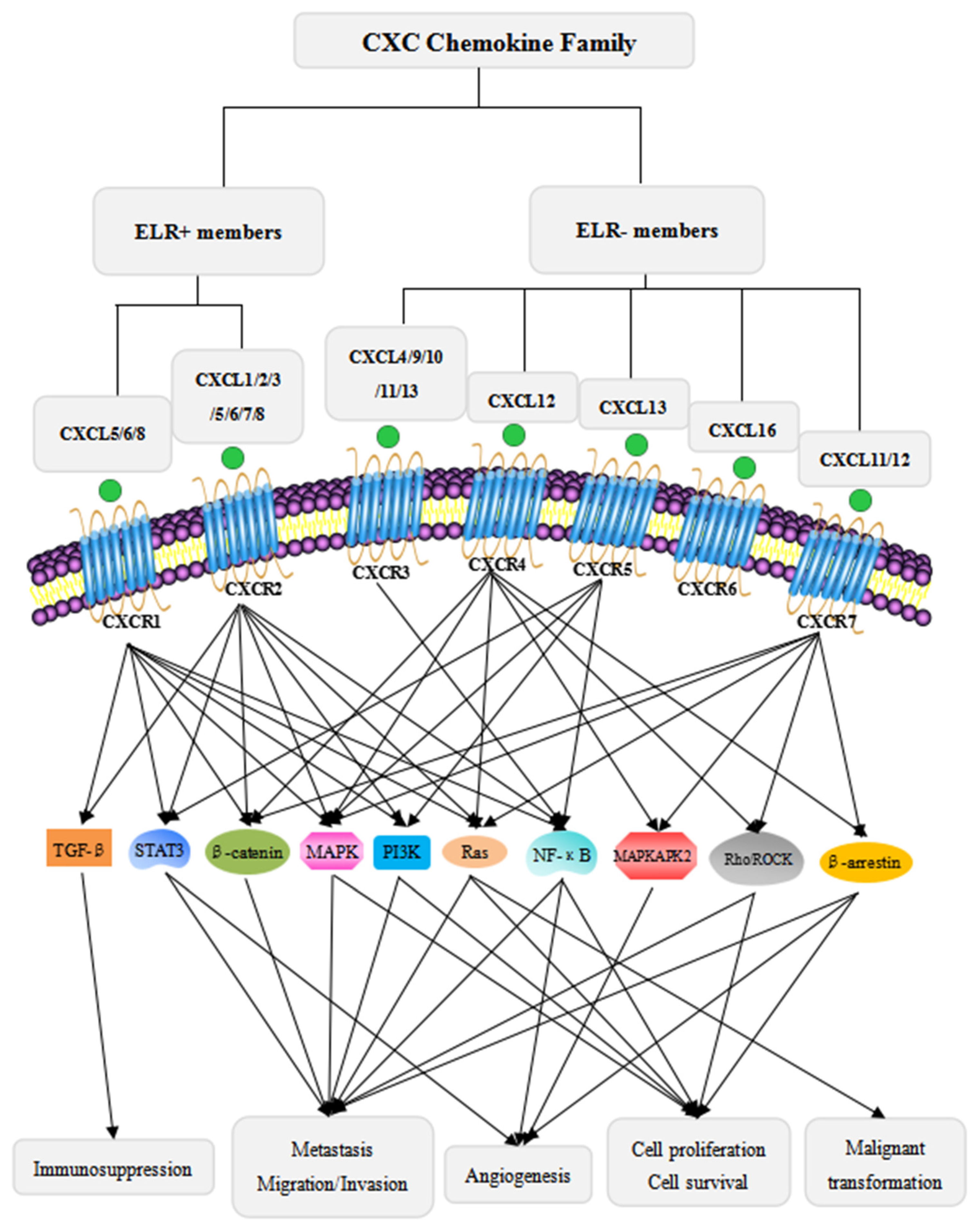
2. CXC Chemokine Family and the CXCL/CXCR Signaling Axes
3. CXC Chemokines and Tumor Angiogenesis
4. CXC Chemokines and Tumor-Associated Inflammation

{kind=link}
{kind=link}
| CXC Chemokine | Stimulus * | References | |||
|---|---|---|---|---|---|
| LPS | TNF | IL-1 | IFN-γ | ||
| CXCL1/GRO-α | + | + | + | - | [96] |
| CXCL2/GRO-β | + | + | + | ND | [49,97,98] |
| CXCL3/GRO-γ | + | + | + | ND | [84,99,100] |
| CXCL4/PF4 | + | ND | ND | ND | [101] |
| CXCL5/ENA-78 | + | + | + | - | [96] |
| CXCL6/GCP-2 | + | + | + | - | [102] |
| CXCL7/NAP-2 | + | + | + | + | [84,103] |
| CXCL8/IL-8 | + | + | + | - | [96,102] |
| CXCL9/MIG | + | - | - | + | [96,100] |
| CXCL10/IP-10 | + | + | + | + | [96] |
| CXCL11/I-TAC | + | + | + | + | [104,105] |
5. CXC Chemokines and Tumor Metastasis
6. Application of CXC Chemokines for Cancer Diagnosis, Prognosis, and Therapy
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Baird, A.-M.; Gray, S.G.; O’Byrne, K.J. Epigenetics Underpinning the Regulation of the CXC (ELR+) Chemokines in Non-Small Cell Lung Cancer. PLoS ONE 2011, 6, e14593. [Google Scholar] [CrossRef] [Green Version]
- Zlotnik, A.; Yoshie, O. Chemokines: A New Classification System and Their Role in Immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, N.; Zhang, T.-T.; Nakanishi, T. Involvement of CXCR4 in Normal and Abnormal Development. Cells 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Unver, N. Macrophage Chemoattractants Secreted by Cancer Cells: Sculptors of the Tumor Microenvironment and Another Crucial Piece of the Cancer Secretome as a Therapeutic Target. Cytokine Growth Factor Rev. 2019, 50, 13–18. [Google Scholar] [CrossRef]
- Gijsbers, K.; Gouwy, M.; Struyf, S.; Wuyts, A.; Proost, P.; Opdenakker, G.; Penninckx, F.; Ectors, N.; Geboes, K.; Van Damme, J. GCP-2/CXCL6 Synergizes with Other Endothelial Cell-Derived Chemokines in Neutrophil Mobilization and Is Associated with Angiogenesis in Gastrointestinal Tumors. Exp. Cell Res. 2005, 303, 331–342. [Google Scholar] [CrossRef]
- Zhou, Z.; Xia, G.; Xiang, Z.; Liu, M.; Wei, Z.; Yan, J.; Chen, W.; Zhu, J.; Awasthi, N.; Sun, X.; et al. A C-X-C Chemokine Receptor Type 2-Dominated Cross-Talk between Tumor Cells and Macrophages Drives Gastric Cancer Metastasis. Clin. Cancer Res. 2019, 25, 3317–3328. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liu, W.; Zheng, Y.; Wang, S.; Yang, B.; Li, M.; Song, J.; Zhang, F.; Zhang, X.; Wang, Q.; et al. CXCL1 Derived from Tumor-Associated Macrophages Promotes Breast Cancer Metastasis via Activating NF-ΚB/SOX4 Signaling. Cell Death Dis. 2018, 9, 880. [Google Scholar] [CrossRef]
- Sun, X.; He, X.; Zhang, Y.; Hosaka, K.; Andersson, P.; Wu, J.; Wu, J.; Jing, X.; Du, Q.; Hui, X.; et al. Inflammatory Cell-Derived CXCL3 Promotes Pancreatic Cancer Metastasis through a Novel Myofibroblast-Hijacked Cancer Escape Mechanism. Gut 2022, 71, 129–147. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Chen, X.; Li, Y.; Li, A.; Liu, D.; Li, F.; Luo, T. Functions of CXC Chemokines as Biomarkers and Potential Therapeutic Targets in the Hepatocellular Carcinoma Microenvironment. Transl. Cancer Res. 2021, 10, 2169–2187. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Li, G.; Wu, H.; Sun, K.; Chen, J.; Feng, Y.; Chen, C.; Cai, S.; Xu, J.; et al. CXCL1 from Tumor-Associated Lymphatic Endothelial Cells Drives Gastric Cancer Cell into Lymphatic System via Activating Integrin Β1/FAK/AKT Signaling. Cancer Lett. 2017, 385, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, C.; He, Y.; Wu, H.; Wang, Z.; Song, W.; Li, W.; He, W.; Cai, S.; Zhan, W. Lymphatic Endothelial Cell-Secreted CXCL1 Stimulates Lymphangiogenesis and Metastasis of Gastric Cancer. Int. J. Cancer 2012, 130, 787–797. [Google Scholar] [CrossRef]
- Wei, L.-Y.; Lee, J.-J.; Yeh, C.-Y.; Yang, C.-J.; Kok, S.-H.; Ko, J.-Y.; Tsai, F.-C.; Chia, J.-S. Reciprocal Activation of Cancer-Associated Fibroblasts and Oral Squamous Carcinoma Cells through CXCL1. Oral Oncol. 2019, 88, 115–123. [Google Scholar] [CrossRef]
- Park, G.-Y.; Pathak, H.B.; Godwin, A.K.; Kwon, Y. Epithelial-Stromal Communication via CXCL1-CXCR2 Interaction Stimulates Growth of Ovarian Cancer Cells through P38 Activation. Cell. Oncol. 2021, 44, 77–92. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Jiang, Q.; Deng, J.; Xu, F.; Chen, X.; Cheng, F.; Zhang, Y.; Yao, Y.; Xia, Z.; et al. Human Adipose-Derived Mesenchymal Stem Cell-Secreted CXCL1 and CXCL8 Facilitate Breast Tumor Growth By Promoting Angiogenesis. Stem Cells 2017, 35, 2060–2070. [Google Scholar] [CrossRef] [Green Version]
- Hariharan, N.; Ashcraft, K.A.; Svatek, R.S.; Livi, C.B.; Wilson, D.; Kaushik, D.; Leach, R.J.; Johnson-Pais, T.L. Adipose Tissue-Secreted Factors Alter Bladder Cancer Cell Migration. J. Obes. 2018, 2018, 9247864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Li, Y.; Liu, S.; Jiang, Y.; Ma, J.; Wan, L.; Li, Q.; Pang, T. CXCL8 Derived from Mesenchymal Stromal Cells Supports Survival and Proliferation of Acute Myeloid Leukemia Cells through the PI3K/AKT Pathway. FASEB J. 2019, 33, 4755–4764. [Google Scholar] [CrossRef]
- Awaji, M.; Futakuchi, M.; Heavican, T.; Iqbal, J.; Singh, R.K. Cancer-Associated Fibroblasts Enhance Survival and Progression of the Aggressive Pancreatic Tumor Via FGF-2 and CXCL8. Cancer Microenviron. 2019, 12, 37–46. [Google Scholar] [CrossRef]
- Taub, D.D.; Oppenheim, J.J. Chemokines, Inflammation and the Immune System. Ther. Immunol. 1994, 1, 229–246. [Google Scholar]
- Dhawan, P.; Richmond, A. Role of CXCL1 in Tumorigenesis of Melanoma. J. Leukoc. Biol. 2002, 72, 9–18. [Google Scholar] [CrossRef]
- Lee, N.-H.; Nikfarjam, M.; He, H. Functions of the CXC Ligand Family in the Pancreatic Tumor Microenvironment. Pancreatology 2018, 18, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Polverini, P.J.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Kasper, J.; Dzuiba, J.; Van Damme, J.; Walz, A.; Marriott, D. The Functional Role of the ELR Motif in CXC Chemokine-Mediated Angiogenesis. J. Biol. Chem. 1995, 270, 27348–27357. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liang, M.; Lin, Y.; Lv, J.; Chen, M.; Zhou, P.; Fu, F.; Wang, C. Transcriptional Expressions of CXCL9/10/12/13 as Prognosis Factors in Breast Cancer. J. Oncol. 2020, 2020, 4270957. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, R.; Jin, R.; Huang, Z. The Role of CXCL Chemokine Family in the Development and Progression of Gastric Cancer. Int. J. Clin. Exp. Pathol. 2020, 13, 484–492. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Huynh, C.; Dingemanse, J.; Meyer Zu Schwabedissen, H.E.; Sidharta, P.N. Relevance of the CXCR4/CXCR7-CXCL12 Axis and Its Effect in Pathophysiological Conditions. Pharmacol. Res. 2020, 161, 105092. [Google Scholar] [CrossRef]
- Zhang, Z.; Tan, X.; Luo, J.; Cui, B.; Lei, S.; Si, Z.; Shen, L.; Yao, H. GNA13 Promotes Tumor Growth and Angiogenesis by Upregulating CXC Chemokines via the NF-ΚB Signaling Pathway in Colorectal Cancer Cells. Cancer Med. 2018, 7, 5611–5620. [Google Scholar] [CrossRef]
- Yasumoto, K.; Koizumi, K.; Kawashima, A.; Saitoh, Y.; Arita, Y.; Shinohara, K.; Minami, T.; Nakayama, T.; Sakurai, H.; Takahashi, Y.; et al. Role of the CXCL12/CXCR4 Axis in Peritoneal Carcinomatosis of Gastric Cancer. Cancer Res. 2006, 66, 2181–2187. [Google Scholar] [CrossRef] [Green Version]
- Verbeke, H.; Geboes, K.; Van Damme, J.; Struyf, S. The Role of CXC Chemokines in the Transition of Chronic Inflammation to Esophageal and Gastric Cancer. Biochim. Biophys. Acta 2012, 1825, 117–129. [Google Scholar] [CrossRef]
- Chao, C.-C.; Lee, W.-F.; Wang, S.-W.; Chen, P.-C.; Yamamoto, A.; Chang, T.-M.; Weng, S.-L.; Liu, J.-F. CXC Chemokine Ligand-13 Promotes Metastasis via CXCR5-Dependent Signaling Pathway in Non-Small Cell Lung Cancer. J. Cell. Mol. Med. 2021, 25, 9128–9140. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, B.; Li, R.; Chen, J.; Xu, G.; Zhu, Y.; Li, J.; Liang, Q.; Hua, Q.; Wang, L.; et al. KIAA1199 Drives Immune Suppression to Promote Colorectal Cancer Liver Metastasis by Modulating Neutrophil Infiltration. Hepatology 2022, 76, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, A.; Tian, Y.; Wu, J.D.; Liu, Y.; Li, T.; Chen, Y.; Han, X.; Wu, K. The CXCL8-CXCR1/2 Pathways in Cancer. Cytokine Growth Factor Rev. 2016, 31, 61–71. [Google Scholar] [CrossRef]
- Wang, L.; Shi, L.; Gu, J.; Zhan, C.; Xi, J.; Ding, J.; Ge, D. CXCL5 Regulation of Proliferation and Migration in Human Non-Small Cell Lung Cancer Cells. J. Physiol. Biochem. 2018, 74, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.; Sarkissyan, M.; Wu, Y.; Vadgama, J.V. GROα Overexpression Drives Cell Migration and Invasion in Triple Negative Breast Cancer Cells. Oncol. Rep. 2017, 38, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Zhao, Y.; Xu, J. Activated CXCL5-CXCR2 Axis Promotes the Migration, Invasion and EMT of Papillary Thyroid Carcinoma Cells via Modulation of β-Catenin Pathway. Biochimie 2018, 148, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.-W.; Xia, G.-K.; Wu, Y.; Chen, W.; Xiang, Z.; Schwarz, R.E.; Brekken, R.A.; Awasthi, N.; He, Y.-L.; Zhang, C.-H. CXCL1 Promotes Tumor Growth through VEGF Pathway Activation and Is Associated with Inferior Survival in Gastric Cancer. Cancer Lett. 2015, 359, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Alonso-Valenteen, F.; Wang, Z.; Deng, N.; Lee, T.-Y.; Gao, B.; Zhang, Y.; Xu, Y.; Zhang, X.; Billet, S.; et al. A Chemokine Regulatory Loop Induces Cholesterol Synthesis in Lung-Colonizing Triple-Negative Breast Cancer Cells to Fuel Metastatic Growth. Mol. Ther. 2022, 30, 672–687. [Google Scholar] [CrossRef]
- Suarez-Lopez, L.; Kong, Y.W.; Sriram, G.; Patterson, J.C.; Rosenberg, S.; Morandell, S.; Haigis, K.M.; Yaffe, M.B. MAPKAP Kinase-2 Drives Expression of Angiogenic Factors by Tumor-Associated Macrophages in a Model of Inflammation-Induced Colon Cancer. Front. Immunol. 2020, 11, 607891. [Google Scholar] [CrossRef]
- Lin, S.-C.; Gou, G.-H.; Hsia, C.-W.; Ho, C.-W.; Huang, K.-L.; Wu, Y.-F.; Lee, S.-Y.; Chen, Y.-H. Simulated Microgravity Disrupts Cytoskeleton Organization and Increases Apoptosis of Rat Neural Crest Stem Cells Via Upregulating CXCR4 Expression and RhoA-ROCK1-P38 MAPK-P53 Signaling. Stem Cells Dev. 2016, 25, 1172–1193. [Google Scholar] [CrossRef]
- Daniel, S.K.; Seo, Y.D.; Pillarisetty, V.G. The CXCL12-CXCR4/CXCR7 Axis as a Mechanism of Immune Resistance in Gastrointestinal Malignancies. Semin. Cancer Biol. 2020, 65, 176–188. [Google Scholar] [CrossRef]
- Lounsbury, N. Advances in CXCR7 Modulators. Pharmaceuticals 2020, 13, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Molino Del Barrio, I.; Wilkins, G.C.; Meeson, A.; Ali, S.; Kirby, J.A. Breast Cancer: An Examination of the Potential of ACKR3 to Modify the Response of CXCR4 to CXCL12. Int. J. Mol. Sci. 2018, 19, 3592. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, E.L.; Lee, W.; Lu, J.; Lowy, A.M.; Kim, J. Chemokine CXCL12 Activates Dual CXCR4 and CXCR7-Mediated Signaling Pathways in Pancreatic Cancer Cells. J. Transl. Med. 2012, 10, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, R.; Blando, J.M.; Perez, C.J.; Abba, M.C.; Benavides, F.; Kazanietz, M.G. Protein Kinase C Epsilon Cooperates with PTEN Loss for Prostate Tumorigenesis through the CXCL13-CXCR5 Pathway. Cell Rep. 2017, 19, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Mi, Y.; Guan, B.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Cai, S.; Xu, Y.; Li, X.; et al. Tumor-Derived Exosomal MiR-934 Induces Macrophage M2 Polarization to Promote Liver Metastasis of Colorectal Cancer. J. Hematol. Oncol. 2020, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-S.; Chin, C.-C.; Chen, C.-N.; Kuo, Y.-H.; Chen, T.-C.; Yu, H.-R.; Tung, S.-Y.; Shen, C.-H.; Hsieh, Y.-Y.; Guo, S.-E.; et al. Stromal Cell-Derived Factor-1/CXC Receptor 4 and Β1 Integrin Interaction Regulates Urokinase-Type Plasminogen Activator Expression in Human Colorectal Cancer Cells. J. Cell. Physiol. 2012, 227, 1114–1122. [Google Scholar] [CrossRef]
- Mo, M.; Tong, S.; Li, T.; Zu, X.; Hu, X. Serum CXCL13 Level Is Associated with Tumor Progression and Unfavorable Prognosis in Penile Cancer. OncoTargets Ther. 2020, 13, 8757–8769. [Google Scholar] [CrossRef]
- Lee, C.-H.; Syu, S.-H.; Liu, K.-J.; Chu, P.-Y.; Yang, W.-C.; Lin, P.; Shieh, W.-Y. Interleukin-1 Beta Transactivates Epidermal Growth Factor Receptor via the CXCL1-CXCR2 Axis in Oral Cancer. Oncotarget 2015, 6, 38866–38880. [Google Scholar] [CrossRef] [Green Version]
- Acharyya, S.; Oskarsson, T.; Vanharanta, S.; Malladi, S.; Kim, J.; Morris, P.G.; Manova-Todorova, K.; Leversha, M.; Hogg, N.; Seshan, V.E.; et al. A CXCL1 Paracrine Network Links Cancer Chemoresistance and Metastasis. Cell 2012, 150, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Põld, M.; Zhu, L.X.; Sharma, S.; Burdick, M.D.; Lin, Y.; Lee, P.P.N.; Põld, A.; Luo, J.; Krysan, K.; Dohadwala, M.; et al. Cyclooxygenase-2-Dependent Expression of Angiogenic CXC Chemokines ENA-78/CXC Ligand (CXCL) 5 and Interleukin-8/CXCL8 in Human Non-Small Cell Lung Cancer. Cancer Res. 2004, 64, 1853–1860. [Google Scholar] [CrossRef] [Green Version]
- Verbeke, H.; Struyf, S.; Berghmans, N.; Van Coillie, E.; Opdenakker, G.; Uyttenhove, C.; Van Snick, J.; Van Damme, J. Isotypic Neutralizing Antibodies against Mouse GCP-2/CXCL6 Inhibit Melanoma Growth and Metastasis. Cancer Lett. 2011, 302, 54–62. [Google Scholar] [CrossRef]
- Catusse, J.; Struyf, S.; Wuyts, A.; Weyler, M.; Loos, T.; Gijsbers, K.; Gouwy, M.; Proost, P.; Van Damme, J. Rabbit Neutrophil Chemotactic Protein (NCP) Activates Both CXCR1 and CXCR2 and Is the Functional Homologue for Human CXCL6. Biochem. Pharmacol. 2004, 68, 1947–1955. [Google Scholar] [CrossRef]
- Arenberg, D.A.; Kunkel, S.L.; Polverini, P.J.; Morris, S.B.; Burdick, M.D.; Glass, M.C.; Taub, D.T.; Iannettoni, M.D.; Whyte, R.I.; Strieter, R.M. Interferon-Gamma-Inducible Protein 10 (IP-10) Is an Angiostatic Factor That Inhibits Human Non-Small Cell Lung Cancer (NSCLC) Tumorigenesis and Spontaneous Metastases. J. Exp. Med. 1996, 184, 981–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaks, A. Role of CXC Group Chemokines in Lung Cancer Development and Progression. J. Thorac. Dis. 2017, 9, S164–S171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, N.; Wildbaum, G. The Role of Chemokines in Shaping the Balance Between CD4(+) T Cell Subsets and Its Therapeutic Implications in Autoimmune and Cancer Diseases. Front. Immunol. 2015, 6, 609. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wu, K.; Tao, K.; Lu, X.; Ma, J.; Mao, Z.; Li, H.; Shi, L.; Li, J.; Niu, Y.; et al. Vasoactive Intestinal Peptide Induces CD14+ HLA-DR-/Low Myeloid-Derived Suppressor Cells in Gastric Cancer. Mol. Med. Rep. 2015, 12, 760–768. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, C.; Kumar, R.; Ender, J.; Godavarthy, P.S.; Hartmann, M.; Hey, J.; Breuer, K.; Weissenberger, E.S.; Minciacchi, V.R.; Karantanou, C.; et al. The Age of the Bone Marrow Microenvironment Influences B-Cell Acute Lymphoblastic Leukemia Progression via CXCR5-CXCL13. Blood 2021, 138, 1870–1884. [Google Scholar] [CrossRef]
- Chuntharapai, A.; Lee, J.; Hébert, C.A.; Kim, K.J. Monoclonal Antibodies Detect Different Distribution Patterns of IL-8 Receptor A and IL-8 Receptor B on Human Peripheral Blood Leukocytes. J. Immunol. 1994, 153, 5682–5688. [Google Scholar]
- Murdoch, C.; Muthana, M.; Coffelt, S.B.; Lewis, C.E. The Role of Myeloid Cells in the Promotion of Tumour Angiogenesis. Nat. Rev. Cancer 2008, 8, 618–631. [Google Scholar] [CrossRef]
- Singh, S.; Nannuru, K.C.; Sadanandam, A.; Varney, M.L.; Singh, R.K. CXCR1 and CXCR2 Enhances Human Melanoma Tumourigenesis, Growth and Invasion. Br. J. Cancer 2009, 100, 1638–1646. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Sadanandam, A.; Varney, M.L.; Nannuru, K.C.; Singh, R.K. Small Interfering RNA-Mediated CXCR1 or CXCR2 Knock-down Inhibits Melanoma Tumor Growth and Invasion. Int. J. Cancer 2010, 126, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Mo, F.; Li, Q.; Han, X.; Shi, H.; Chen, S.; Wei, Y.; Wei, X. Targeting CXCR2 Inhibits the Progression of Lung Cancer and Promotes Therapeutic Effect of Cisplatin. Mol. Cancer 2021, 20, 62. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Zheng, J.; Hou, K.; Wang, J.; Chen, X.; Lu, X.; Bo, J.; Xu, C.; Shen, K.; Wang, J. Role of Chemokine Receptor CXCR7 in Bladder Cancer Progression. Biochem. Pharmacol. 2012, 84, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Keane, M.P.; Belperio, J.A.; Xue, Y.Y.; Burdick, M.D.; Strieter, R.M. Depletion of CXCR2 Inhibits Tumor Growth and Angiogenesis in a Murine Model of Lung Cancer. J. Immunol. 2004, 172, 2853–2860. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Raimondo, M.; Woodward, T.A.; Wallace, M.B.; Gill, K.R.; Tong, Z.; Burdick, M.D.; Yang, Z.; Strieter, R.M.; Hoffman, R.M.; et al. CXC-Chemokine/CXCR2 Biological Axis Promotes Angiogenesis in Vitro and in Vivo in Pancreatic Cancer. Int. J. Cancer 2009, 125, 1027–1037. [Google Scholar] [CrossRef]
- Takikawa, T.; Hamada, S.; Matsumoto, R.; Tanaka, Y.; Kataoka, F.; Sasaki, A.; Masamune, A. Senescent Human Pancreatic Stellate Cells Secrete CXCR2 Agonist CXCLs to Promote Proliferation and Migration of Human Pancreatic Cancer AsPC-1 and MIAPaCa-2 Cell Lines. Int. J. Mol. Sci. 2022, 23, 9275. [Google Scholar] [CrossRef]
- Mian, B.M.; Dinney, C.P.N.; Bermejo, C.E.; Sweeney, P.; Tellez, C.; Yang, X.D.; Gudas, J.M.; McConkey, D.J.; Bar-Eli, M. Fully Human Anti-Interleukin 8 Antibody Inhibits Tumor Growth in Orthotopic Bladder Cancer Xenografts via down-Regulation of Matrix Metalloproteases and Nuclear Factor-KappaB. Clin. Cancer Res. 2003, 9, 3167–3175. [Google Scholar]
- Huang, S.; Mills, L.; Mian, B.; Tellez, C.; McCarty, M.; Yang, X.-D.; Gudas, J.M.; Bar-Eli, M. Fully Humanized Neutralizing Antibodies to Interleukin-8 (ABX-IL8) Inhibit Angiogenesis, Tumor Growth, and Metastasis of Human Melanoma. Am. J. Pathol. 2002, 161, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Bachmeier, B.E.; Mohrenz, I.V.; Mirisola, V.; Schleicher, E.; Romeo, F.; Höhneke, C.; Jochum, M.; Nerlich, A.G.; Pfeffer, U. Curcumin Downregulates the Inflammatory Cytokines CXCL1 and -2 in Breast Cancer Cells via NFkappaB. Carcinogenesis 2008, 29, 779–789. [Google Scholar] [CrossRef]
- Yang, C.; Yu, H.; Chen, R.; Tao, K.; Jian, L.; Peng, M.; Li, X.; Liu, M.; Liu, S. CXCL1 Stimulates Migration and Invasion in ER-negative Breast Cancer Cells via Activation of the ERK/MMP2/9 Signaling Axis. Int. J. Oncol. 2019, 55, 684–696. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.K.; Chai, X.; Ghosh, S.; Ray, D.; Wang, M.; Rasheed, S.A.K.; Casey, P.J. Gα-13 Induces CXC Motif Chemokine Ligand 5 Expression in Prostate Cancer Cells by Transactivating NF-ΚB. J. Biol. Chem. 2019, 294, 18192–18206. [Google Scholar] [CrossRef] [PubMed]
- Varney, M.L.; Singh, S.; Li, A.; Mayer-Ezell, R.; Bond, R.; Singh, R.K. Small Molecule Antagonists for CXCR2 and CXCR1 Inhibit Human Colon Cancer Liver Metastases. Cancer Lett. 2011, 300, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Sadanandam, A.; Nannuru, K.C.; Varney, M.L.; Mayer-Ezell, R.; Bond, R.; Singh, R.K. Small-Molecule Antagonists for CXCR2 and CXCR1 Inhibit Human Melanoma Growth by Decreasing Tumor Cell Proliferation, Survival, and Angiogenesis. Clin. Cancer Res. 2009, 15, 2380–2386. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Labonte, M.J.; Zhang, W.; Bohanes, P.O.; Gerger, A.; Yang, D.; Benhaim, L.; Paez, D.; Rosenberg, D.O.; Nagulapalli Venkata, K.C.; et al. The CXCR2 Antagonist, SCH-527123, Shows Antitumor Activity and Sensitizes Cells to Oxaliplatin in Preclinical Colon Cancer Models. Mol. Cancer Ther. 2012, 11, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Walser, T.C.; Rifat, S.; Ma, X.; Kundu, N.; Ward, C.; Goloubeva, O.; Johnson, M.G.; Medina, J.C.; Collins, T.L.; Fulton, A.M. Antagonism of CXCR3 Inhibits Lung Metastasis in a Murine Model of Metastatic Breast Cancer. Cancer Res. 2006, 66, 7701–7707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, J.M.; Summers, B.C.; Wang, Y.; Melikian, A.; Berahovich, R.; Miao, Z.; Penfold, M.E.T.; Sunshine, M.J.; Littman, D.R.; Kuo, C.J.; et al. A Novel Chemokine Receptor for SDF-1 and I-TAC Involved in Cell Survival, Cell Adhesion, and Tumor Development. J. Exp. Med. 2006, 203, 2201–2213. [Google Scholar] [CrossRef]
- Yoon, Y.; Liang, Z.; Zhang, X.; Choe, M.; Zhu, A.; Cho, H.T.; Shin, D.M.; Goodman, M.M.; Chen, Z.; Shim, H. CXC Chemokine Receptor-4 Antagonist Blocks Both Growth of Primary Tumor and Metastasis of Head and Neck Cancer in Xenograft Mouse Models. Cancer Res. 2007, 67, 7518–7524. [Google Scholar] [CrossRef] [Green Version]
- Rubin, J.B.; Kung, A.L.; Klein, R.S.; Chan, J.A.; Sun, Y.; Schmidt, K.; Kieran, M.W.; Luster, A.D.; Segal, R.A. A Small-Molecule Antagonist of CXCR4 Inhibits Intracranial Growth of Primary Brain Tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 13513–13518. [Google Scholar] [CrossRef] [Green Version]
- Tamamura, H.; Hori, A.; Kanzaki, N.; Hiramatsu, K.; Mizumoto, M.; Nakashima, H.; Yamamoto, N.; Otaka, A.; Fujii, N. T140 Analogs as CXCR4 Antagonists Identified as Anti-Metastatic Agents in the Treatment of Breast Cancer. FEBS Lett. 2003, 550, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Wente, M.N.; Keane, M.P.; Burdick, M.D.; Friess, H.; Büchler, M.W.; Ceyhan, G.O.; Reber, H.A.; Strieter, R.M.; Hines, O.J. Blockade of the Chemokine Receptor CXCR2 Inhibits Pancreatic Cancer Cell-Induced Angiogenesis. Cancer Lett. 2006, 241, 221–227. [Google Scholar] [CrossRef]
- Arenberg, D.A.; Keane, M.P.; DiGiovine, B.; Kunkel, S.L.; Morris, S.B.; Xue, Y.Y.; Burdick, M.D.; Glass, M.C.; Iannettoni, M.D.; Strieter, R.M. Epithelial-Neutrophil Activating Peptide (ENA-78) Is an Important Angiogenic Factor in Non-Small Cell Lung Cancer. J. Clin. Investig. 1998, 102, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Carretero-Ortega, J.; Walsh, C.T.; Hernández-García, R.; Reyes-Cruz, G.; Brown, J.H.; Vázquez-Prado, J. Phosphatidylinositol 3,4,5-Triphosphate-Dependent Rac Exchanger 1 (P-Rex-1), a Guanine Nucleotide Exchange Factor for Rac, Mediates Angiogenic Responses to Stromal Cell-Derived Factor-1/Chemokine Stromal Cell Derived Factor-1 (SDF-1/CXCL-12) Linked to Rac Activation, Endothelial Cell Migration, and in Vitro Angiogenesis. Mol. Pharmacol. 2010, 77, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Singh, S.; Varney, M.L.; Kindle, S.; Singh, R.K. Modulation of CXCL-8 Expression in Human Melanoma Cells Regulates Tumor Growth, Angiogenesis, Invasion, and Metastasis. Cancer Med. 2012, 1, 306–317. [Google Scholar] [CrossRef]
- Keshamouni, V.G.; Arenberg, D.A.; Reddy, R.C.; Newstead, M.J.; Anthwal, S.; Standiford, T.J. PPAR-Gamma Activation Inhibits Angiogenesis by Blocking ELR+CXC Chemokine Production in Non-Small Cell Lung Cancer. Neoplasia 2005, 7, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Kamohara, H.; Takahashi, M.; Ishiko, T.; Ogawa, M.; Baba, H. Induction of Interleukin-8 (CXCL-8) by Tumor Necrosis Factor-Alpha and Leukemia Inhibitory Factor in Pancreatic Carcinoma Cells: Impact of CXCL-8 as an Autocrine Growth Factor. Int. J. Oncol. 2007, 31, 627–632. [Google Scholar] [PubMed]
- Wu, K.; Yu, S.; Liu, Q.; Bai, X.; Zheng, X.; Wu, K. The Clinical Significance of CXCL5 in Non-Small Cell Lung Cancer. OncoTargets Ther. 2017, 10, 5561–5573. [Google Scholar] [CrossRef] [Green Version]
- Sparmann, A.; Bar-Sagi, D. Ras-Induced Interleukin-8 Expression Plays a Critical Role in Tumor Growth and Angiogenesis. Cancer Cell 2004, 6, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Huang, H.; Wang, Z.; Zhang, G. The Inflammatory CXC Chemokines, GROαhigh, IP-10low, and MIGlow, in Tumor Microenvironment Can Be Used as New Indicators for Non-Small Cell Lung Cancer Progression. Immunol. Investig. 2017, 46, 361–374. [Google Scholar] [CrossRef]
- Shalapour, S.; Karin, M. Immunity, Inflammation, and Cancer: An Eternal Fight between Good and Evil. J. Clin. Investig. 2015, 125, 3347–3355. [Google Scholar] [CrossRef] [Green Version]
- Vandercappellen, J.; Van Damme, J.; Struyf, S. The Role of CXC Chemokines and Their Receptors in Cancer. Cancer Lett. 2008, 267, 226–244. [Google Scholar] [CrossRef]
- Moore, B.B.; Arenberg, D.A.; Strieter, R.M. The Role of CXC Chemokines in the Regulation of Angiogenesis in Association with Lung Cancer. Trends Cardiovasc. Med. 1998, 8, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Tazzyman, S.; Barry, S.T.; Ashton, S.; Wood, P.; Blakey, D.; Lewis, C.E.; Murdoch, C. Inhibition of Neutrophil Infiltration into A549 Lung Tumors in Vitro and in Vivo Using a CXCR2-Specific Antagonist Is Associated with Reduced Tumor Growth. Int. J. Cancer 2011, 129, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, E.; Ko, E.; Ham, M.; Lee, H.M.; Kim, E.-S.; Koh, M.; Lim, H.K.; Jung, J.; Park, S.Y.; et al. Tumor-Associated Macrophages Secrete CCL2 and Induce the Invasive Phenotype of Human Breast Epithelial Cells through Upregulation of ERO1-α and MMP-9. Cancer Lett. 2018, 437, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Greten, T.F. Does CSF1R Blockade Turn into Friendly Fire? Cancer Cell 2017, 32, 546–547. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.B.; Hajal, C.; Benjamin, D.C.; Yu, C.; Azizgolshani, H.; Hynes, R.O.; Kamm, R.D. Inflamed Neutrophils Sequestered at Entrapped Tumor Cells via Chemotactic Confinement Promote Tumor Cell Extravasation. Proc. Natl. Acad. Sci. USA 2018, 115, 7022–7027. [Google Scholar] [CrossRef] [Green Version]
- Arenberg, D.A.; Polverini, P.J.; Kunkel, S.L.; Shanafelt, A.; Hesselgesser, J.; Horuk, R.; Strieter, R.M. The Role of CXC Chemokines in the Regulation of Angiogenesis in Non-Small Cell Lung Cancer. J. Leukoc. Biol. 1997, 62, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Carmody, R.J.; Ruan, Q.; Palmer, S.; Hilliard, B.; Chen, Y.H. Negative Regulation of Toll-like Receptor Signaling by NF-KappaB P50 Ubiquitination Blockade. Science 2007, 317, 675–678. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.J.; Lu, D.; Sparer, T.E.; Masi, T.; Goff, M.R.; Karlstad, M.D.; Collier, J.J. NF-ΚB and STAT1 Control CXCL1 and CXCL2 Gene Transcription. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E131–E149. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Kim, J.-K.; Jeon, H.-Y.; Ham, S.W.; Kim, H. CD133 Regulates IL-1β Signaling and Neutrophil Recruitment in Glioblastoma. Mol. Cells 2017, 40, 515–522. [Google Scholar] [CrossRef]
- Bandow, K.; Kusuyama, J.; Shamoto, M.; Kakimoto, K.; Ohnishi, T.; Matsuguchi, T. LPS-Induced Chemokine Expression in Both MyD88-Dependent and -Independent Manners Is Regulated by Cot/Tpl2-ERK Axis in Macrophages. FEBS Lett. 2012, 586, 1540–1546. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, Z.; Zhu, K.; Bao, R.; Meng, Y.; Bian, J.; Wan, X.; Yang, T. Effects of CXCL4/CXCR3 on the Lipopolysaccharide-Induced Injury in Human Umbilical Vein Endothelial Cells. J. Cell. Physiol. 2019, 234, 22378–22385. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, A.; Struyf, S.; Gijsbers, K.; Schutyser, E.; Put, W.; Conings, R.; Lenaerts, J.-P.; Geboes, K.; Opdenakker, G.; Menten, P.; et al. The CXC Chemokine GCP-2/CXCL6 Is Predominantly Induced in Mesenchymal Cells by Interleukin-1β and Is Down-Regulated by Interferon-γ: Comparison with Interleukin-8/CXCL8. Lab. Investig. 2003, 83, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-S.; Liao, K.-W.; Chen, M.-F.; Huang, Y.-C.; Chu, R.-M.; Chi, K.-H. Canine CXCL7 and Its Functional Expression in Dendritic Cells Undergoing Maturation. Vet. Immunol. Immunopathol. 2010, 135, 128–136. [Google Scholar] [CrossRef]
- Furuya, M.; Suyama, T.; Usui, H.; Kasuya, Y.; Nishiyama, M.; Tanaka, N.; Ishiwata, I.; Nagai, Y.; Shozu, M.; Kimura, S. Up-Regulation of CXC Chemokines and Their Receptors: Implications for Proinflammatory Microenvironments of Ovarian Carcinomas and Endometriosis. Hum. Pathol. 2007, 38, 1676–1687. [Google Scholar] [CrossRef] [PubMed]
- Widney, D.P.; Xia, Y.R.; Lusis, A.J.; Smith, J.B. The Murine Chemokine CXCL11 (IFN-Inducible T Cell Alpha Chemoattractant) Is an IFN-Gamma- and Lipopolysaccharide-Inducible Glucocorticoid-Attenuated Response Gene Expressed in Lung and Other Tissues during Endotoxemia. J. Immunol. 2000, 164, 6322–6331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.F.; Huang, Y.; Han, Y.Y.; Lin, L.Y.; Sun, W.H.; Rabson, A.B.; Wang, Y.; Shi, Y.F. TNFα-Activated Mesenchymal Stromal Cells Promote Breast Cancer Metastasis by Recruiting CXCR2+ Neutrophils. Oncogene 2017, 36, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Danø, K.; Behrendt, N.; Høyer-Hansen, G.; Johnsen, M.; Lund, L.R.; Ploug, M.; Rømer, J. Plasminogen Activation and Cancer. Thromb. Haemost. 2005, 93, 676–681. [Google Scholar] [CrossRef]
- Nv, S.; Tp, F.; We, G. EpCAM Modulates NF-ΚB Signaling and Interleukin-8 Expression in Breast Cancer. Mol. Cancer Res. 2013, 11, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wang, N.; Wang, S.; Yang, B.; Situ, H.; Zhong, L.; Lin, Y.; Wang, Z. XIAOPI Formula Inhibits the Pre-Metastatic Niche Formation in Breast Cancer via Suppressing TAMs/CXCL1 Signaling. Cell Commun. Signal. 2020, 18, 48. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Park, K.H.; Bang, S.; Kim, M.H.; Lee, J.-E.; Gang, J.; Koh, S.S.; Song, S.Y. CXCL5 Overexpression Is Associated with Late Stage Gastric Cancer. J. Cancer Res. Clin. Oncol. 2007, 133, 835–840. [Google Scholar] [CrossRef]
- Rajkumar, T.; Vijayalakshmi, N.; Gopal, G.; Sabitha, K.; Shirley, S.; Raja, U.M.; Ramakrishnan, S.A. Identification and Validation of Genes Involved in Gastric Tumorigenesis. Cancer Cell Int. 2010, 10, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervi, D.; Yip, T.-T.; Bhattacharya, N.; Podust, V.N.; Peterson, J.; Abou-Slaybi, A.; Naumov, G.N.; Bender, E.; Almog, N.; Italiano, J.E.; et al. Platelet-Associated PF-4 as a Biomarker of Early Tumor Growth. Blood 2008, 111, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Qiao, Y.; Liu, W.; Wang, W.; Shen, H.; Lu, Y.; Hao, G.; Zheng, J.; Tian, Y. CXCL5 Is a Potential Diagnostic and Prognostic Marker for Bladder Cancer Patients. Tumour Biol. 2016, 37, 4569–4577. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Matsui, Y.; Kobayashi, T.; Saito, R.; Hatahira, S.; Kawakami, K.; Nakamura, E.; Nishiyama, H.; Ogawa, O. Urine CXCL1 as a Biomarker for Tumor Detection and Outcome Prediction in Bladder Cancer. Cancer Biomark. 2015, 15, 357–364. [Google Scholar] [CrossRef]
- Wilhelm, A.; Lemmenmeier, I.; Lalos, A.; Posabella, A.; Kancherla, V.; Piscuoglio, S.; Delko, T.; von Flüe, M.; Glatz, K.; Droeser, R.A. The Prognostic Significance of CXCR4 and SDF-1 in Differentiated Thyroid Cancer Depends on CD8+ Density. BMC Endocr. Disord. 2022, 22, 292. [Google Scholar] [CrossRef]
- Unver, N. Identification of the Dominant Angiogenic CXCL Class Chemokines Associated with Non-Small Cell Lung Cancer via Bioinformatics Tools. Med. Oncol. 2021, 38, 68. [Google Scholar] [CrossRef]
- Spaks, A.; Jaunalksne, I.; Spaka, I.; Chudasama, D.; Pirtnieks, A.; Krievins, D. Diagnostic Value of Circulating CXC Chemokines in Non-Small Cell Lung Cancer. Anticancer Res. 2015, 35, 6979–6983. [Google Scholar]
- Mlynska, A.; Salciuniene, G.; Zilionyte, K.; Garberyte, S.; Strioga, M.; Intaite, B.; Barakauskiene, A.; Lazzari, G.; Dobrovolskiene, N.; Krasko, J.A.; et al. Chemokine Profiling in Serum from Patients with Ovarian Cancer Reveals Candidate Biomarkers for Recurrence and Immune Infiltration. Oncol. Rep. 2019, 41, 1238–1252. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Hong, Z.; Mao, Y.; Di, W. Correlations of AKIP1, CXCL1 and CXCL2 Expressions with Clinicopathological Features and Survival Profiles in Cervical Cancer Patients. Transl. Cancer Res. 2020, 9, 726–734. [Google Scholar] [CrossRef]
- Wei, Y.; Lin, C.; Li, H.; Xu, Z.; Wang, J.; Li, R.; Liu, H.; Zhang, H.; He, H.; Xu, J. CXCL13 Expression Is Prognostic and Predictive for Postoperative Adjuvant Chemotherapy Benefit in Patients with Gastric Cancer. Cancer Immunol. Immunother. 2018, 67, 261–269. [Google Scholar] [CrossRef]
- Takahashi, Y.; Akishima-Fukasawa, Y.; Kobayashi, N.; Sano, T.; Kosuge, T.; Nimura, Y.; Kanai, Y.; Hiraoka, N. Prognostic Value of Tumor Architecture, Tumor-Associated Vascular Characteristics, and Expression of Angiogenic Molecules in Pancreatic Endocrine Tumors. Clin. Cancer Res. 2007, 13, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufies, M.; Giuliano, S.; Viotti, J.; Borchiellini, D.; Cooley, L.S.; Ambrosetti, D.; Guyot, M.; Ndiaye, P.D.; Parola, J.; Claren, A.; et al. CXCL7 Is a Predictive Marker of Sunitinib Efficacy in Clear Cell Renal Cell Carcinomas. Br. J. Cancer 2017, 117, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.-Y.W.; Wu, P.-Y.; Wu, T.-J.; Hou, H.-A.; Chou, W.-C.; Teng, C.-L.J.; Lin, C.-R.; Chen, J.-M.M.; Lin, T.-Y.; Su, H.-C.; et al. Aurora A and NF-ΚB Survival Pathway Drive Chemoresistance in Acute Myeloid Leukemia via the TRAF-Interacting Protein TIFA. Cancer Res. 2017, 77, 494–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahham, D.; Weiss, I.D.; Abraham, M.; Beider, K.; Hanna, W.; Shlomai, Z.; Eizenberg, O.; Zamir, G.; Izhar, U.; Shapira, O.M.; et al. In Vitro and in Vivo Therapeutic Efficacy of CXCR4 Antagonist BKT140 against Human Non-Small Cell Lung Cancer. J. Thorac. Cardiovasc. Surg. 2012, 144, 1167–1175.e1. [Google Scholar] [CrossRef] [Green Version]
- Rial, N.S.; Lazennec, G.; Prasad, A.R.; Krouse, R.S.; Lance, P.; Gerner, E.W. Regulation of Deoxycholate Induction of CXCL8 by the Adenomatous Polyposis Coli Gene in Colorectal Cancer. Int. J. Cancer 2009, 124, 2270–2280. [Google Scholar] [CrossRef]
- Bellocq, A.; Antoine, M.; Flahault, A.; Philippe, C.; Crestani, B.; Bernaudin, J.F.; Mayaud, C.; Milleron, B.; Baud, L.; Cadranel, J. Neutrophil Alveolitis in Bronchioloalveolar Carcinoma: Induction by Tumor-Derived Interleukin-8 and Relation to Clinical Outcome. Am. J. Pathol. 1998, 152, 83–92. [Google Scholar]
- Haraguchi, M.; Komuta, K.; Akashi, A.; Matsuzaki, S.; Furui, J.; Kanematsu, T. Elevated IL-8 Levels in the Drainage Vein of Resectable Dukes’ C Colorectal Cancer Indicate High Risk for Developing Hepatic Metastasis. Oncol. Rep. 2002, 9, 159–165. [Google Scholar] [CrossRef]
- Kantola, T.; Klintrup, K.; Väyrynen, J.P.; Vornanen, J.; Bloigu, R.; Karhu, T.; Herzig, K.-H.; Näpänkangas, J.; Mäkelä, J.; Karttunen, T.J.; et al. Stage-Dependent Alterations of the Serum Cytokine Pattern in Colorectal Carcinoma. Br. J. Cancer 2012, 107, 1729–1736. [Google Scholar] [CrossRef] [Green Version]
- Kuniyasu, H.; Yasui, W.; Shinohara, H.; Yano, S.; Ellis, L.M.; Wilson, M.R.; Bucana, C.D.; Rikita, T.; Tahara, E.; Fidler, I.J. Induction of Angiogenesis by Hyperplastic Colonic Mucosa Adjacent to Colon Cancer. Am. J. Pathol. 2000, 157, 1523–1535. [Google Scholar] [CrossRef] [Green Version]
- McCarron, S.L.; Edwards, S.; Evans, P.R.; Gibbs, R.; Dearnaley, D.P.; Dowe, A.; Southgate, C.; Easton, D.F.; Eeles, R.A.; Howell, W.M. Influence of Cytokine Gene Polymorphisms on the Development of Prostate Cancer. Cancer Res. 2002, 62, 3369–3372. [Google Scholar]
- Barash, U.; Zohar, Y.; Wildbaum, G.; Beider, K.; Nagler, A.; Karin, N.; Ilan, N.; Vlodavsky, I. Heparanase Enhances Myeloma Progression via CXCL10 Downregulation. Leukemia 2014, 28, 2178–2187. [Google Scholar] [CrossRef] [PubMed]
- Struyf, S.; Burdick, M.D.; Peeters, E.; Van den Broeck, K.; Dillen, C.; Proost, P.; Van Damme, J.; Strieter, R.M. Platelet Factor-4 Variant Chemokine CXCL4L1 Inhibits Melanoma and Lung Carcinoma Growth and Metastasis by Preventing Angiogenesis. Cancer Res. 2007, 67, 5940–5948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Zhang, R.; Gu, F.; Pei, Y.; Sun, L.; Huang, Y.; Niu, G.; Li, J. Plasma CXCL3 Levels Are Associated with Tumor Progression and an Unfavorable Colorectal Cancer Prognosis. J. Immunol. Res. 2022, 2022, 1336509. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-J.; Noh, S.; Jeung, H.-C.; Jung, M.; Kim, T.S.; Noh, S.H.; Roh, J.K.; Chung, H.C.; Rha, S.Y. Chemokine Growth-Regulated Oncogene 1 as a Putative Biomarker for Gastric Cancer Progression. Cancer Sci. 2010, 101, 2200–2206. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kodama, T.; Kita, M.; Imanishi, J.; Kashima, K.; Graham, D.Y. Relation between Cytokines and Helicobacter Pylori in Gastric Cancer. Helicobacter 2001, 6, 116–124. [Google Scholar] [CrossRef]
- O’Riordan, J.M.; Abdel-latif, M.M.; Ravi, N.; McNamara, D.; Byrne, P.J.; McDonald, G.S.A.; Keeling, P.W.N.; Kelleher, D.; Reynolds, J.V. Proinflammatory Cytokine and Nuclear Factor Kappa-B Expression along the Inflammation-Metaplasia-Dysplasia-Adenocarcinoma Sequence in the Esophagus. Am. J. Gastroenterol. 2005, 100, 1257–1264. [Google Scholar] [CrossRef]
- Li, B.; Su, H.; Cao, J.; Zhang, L. CXCL13 Rather than IL-31 Is a Potential Indicator in Patients with Hepatocellular Carcinoma. Cytokine 2017, 89, 91–97. [Google Scholar] [CrossRef]
- Zhu, Y.M.; Bagstaff, S.M.; Woll, P.J. Production and Upregulation of Granulocyte Chemotactic Protein-2/CXCL6 by IL-1beta and Hypoxia in Small Cell Lung Cancer. Br. J. Cancer 2006, 94, 1936–1941. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhang, X.; Han, D.; Cao, J.; Tian, J. Tumour-Associated Macrophages Mediate the Invasion and Metastasis of Bladder Cancer Cells through CXCL8. PeerJ 2020, 8, e8721. [Google Scholar] [CrossRef]
- Nazari, A.; Khorramdelazad, H.; Hassanshahi, G. Biological/Pathological Functions of the CXCL12/CXCR4/CXCR7 Axes in the Pathogenesis of Bladder Cancer. Int. J. Clin. Oncol. 2017, 22, 991–1000. [Google Scholar] [CrossRef]
- Varney, M.L.; Johansson, S.L.; Singh, R.K. Distinct Expression of CXCL8 and Its Receptors CXCR1 and CXCR2 and Their Association with Vessel Density and Aggressiveness in Malignant Melanoma. Am. J. Clin. Pathol. 2006, 125, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Zhang, Z.; Mantellini, M.G.; Karl, E.; Zeitlin, B.; Verhaegen, M.; Soengas, M.S.; Lingen, M.; Strieter, R.M.; Nunez, G.; et al. Bcl-2 Orchestrates a Cross-Talk between Endothelial and Tumor Cells That Promotes Tumor Growth. Cancer Res. 2007, 67, 9685–9693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, H.; Patel, V.; Wang, H.; Edmunds, R.K.; Gutkind, J.S.; Yeudall, W.A. Down-Regulation of CXCL5 Inhibits Squamous Carcinogenesis. Cancer Res. 2006, 66, 4279–4284. [Google Scholar] [CrossRef] [Green Version]
- Liotti, F.; Collina, F.; Pone, E.; La Sala, L.; Franco, R.; Prevete, N.; Melillo, R.M. Interleukin-8, but Not the Related Chemokine CXCL1, Sustains an Autocrine Circuit Necessary for the Properties and Functions of Thyroid Cancer Stem Cells. Stem Cells Dayt. Ohio 2017, 35, 135–146. [Google Scholar] [CrossRef]
- Qin, X.-J.; Lin, X.; Xue, G.; Fan, H.-L.; Wang, H.-Y.; Wu, J.-F.; Pei, D. CXCL10 Is a Potential Biomarker and Associated with Immune Infiltration in Human Papillary Thyroid Cancer. Biosci. Rep. 2021, 41, BSR20203459. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]

| CXC Chemokine | Alternative Name | Receptor | Effects | References |
|---|---|---|---|---|
| CXCL1 | Melanoma growth -stimulating activity (MGSA)-α/Growth-related oncogene (GRO)-α | CXCR2 |
| [8,13,20,31,48] |
| CXCL2 | MGSA β/GRO-β/ Macrophage inflammatory protein-2 (MIP-2) | CXCR2 |
| [20,21,22,49] |
| CXCL3 | MGSA γ/GRO-γ | CXCR2 |
| [9,20,21,22,31] |
| CXCL4 | Platelet factor 4 (PF4) | CXCR3 |
| [21,22,27] |
| CXCL5 | Epithelial-derived neutrophil-activating factor-78 (ENA-78) | CXCR1 CXCR2 |
| [20,33,35,50] |
| CXCL6 | Granulocyte chemotactic protein-2 (GCP-2) | CXCR1 CXCR2 |
| [20,51,52] |
| CXCL7 | Neutrophil-activating polypeptide-2 (NAP-2) | CXCR2 |
| [20,21,22] |
| CXCL8 | Interleukin-8 (IL-8) | CXCR1 CXCR2 |
| [20,32,50] |
| CXCL9 | Monokine induced by interferon-γ (MIG) | CXCR3 |
| [21,22,53,54] |
| CXCL10 | Interferon-inducible protein 10 (IP-10) | CXCR3 |
| [21,22,53,54,55] |
| CXCL11 | Interferon-inducible T cell α chemoattractant (I-TAC) | CXCR3 |
| [21,22,41,54] |
| CXCL12 | Stromal cell-derived factor-1 (SDF-1) | CXCR4 CXCR7 |
| [28,38,40,41] |
| CXCL13 | B-lymphocyte chemoattractant (BLC) | CXCR3 CXCR5 |
| [47,56,57] |
| CXC Chemokines | Signaling Pathways | Tumor Effects | References |
|---|---|---|---|
| CXCL1 |
|
| [8,11,12,14,20,34,36,69,70] |
| CXCL5 |
|
| [33,35,71] |
| CXCL8 | PI3K/AKT and ERK/MAPK signaling pathways |
| [17,32,60] |
| CXCL1/3 | TGF-β signaling pathway |
| [31] |
| CXCL1/5 | CXCR2/STAT3 pathway |
| [7] |
| CXCL1/2/4 | NF-κB pathway |
| [27] |
| CXCL1/2/8 | NF-κB pathway |
| [37] |
| CXCL12 |
|
| [38,39,40,42,43,46] |
| CXCL11/12 | β-arrestin signaling |
| [41] |
| CXCL13 | NF-κB pathway, PI3K/AKT pathway, ERK/MAPK pathway, STAT3 pathway |
| [30,44,45,47] |
| Cancer Type | CXC Chemokine | Effects | References |
|---|---|---|---|
| Lung cancer | CXCL1/2/5/6/7 CXCL4/8/9/10//11(early-stage) | Tumor promotion | [81,116,117,138] |
| Gastric cancer (GC) | CXCL1/13 CXCL5/8 (late-stage) | Tumor promotion. CXCL13 could be used to stratify GC patients in the T2–4 stage, and low expression of it predicts better therapy response | [110,120,134,135] |
| Colorectal cancer (CRC) | CXCL1/2/3/5/8/11 (up) CXCL12/14 (down) | Tumor promotion | [133] |
| Esophageal adenocarcinoma | CXCL8 (late-stage) | Tumor promotion | [136] |
| Oral squamous carcinoma | CXCL1 | Tumor promotion | [13] |
| Pancreatic carcinoma | CXCL3/8/12 | Tumor promotion | [9,85,121] |
| Hepatocellular carcinoma | CXCL1/3/5 CXCL13 (late-stage) | Tumor promotion | [10,137] |
| Triple-negative breast cancer (TNBC) | CXCL1/2/8 | Tumor promotion | [37] |
| Cervical cancer | CXCL1/2 | Tumor promotion | [119] |
| Ovarian cancer | CXCL1/9 | Tumor promotion | [118] |
| Prostate cancer | CXCL8/13 | Tumor promotion | [32,44] |
| Bladder cancer | CXCL1/5/8/12 | Tumor promotion | [113,114,139,140] |
| Clear cell renal cell carcinoma | CXCL7 (down) | Tumor promotion | [138] |
| Melanoma | CXCL1/8 | Tumor promotion | [20,141] |
| Head and neck squamous cell carcinoma (HNSCC) | CXCL5/8 | Tumor promotion | [142,143] |
| Thyroid cancer | CXCL8/12 CXCL10 (down) | Tumor promotion | [115,144,145] |
| Liposarcoma, Osteosarcoma | CXCL4 (early-stage) | Tumor promotion | [112] |
| B-cell acute lymphoblastic leukemia | CXCL13 | Tumor promotion | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.; Yang, W.; Sun, A.; Wei, Z.; Lin, Q. The Role of CXC Chemokines in Cancer Progression. Cancers 2023, 15, 167. https://doi.org/10.3390/cancers15010167
Wu T, Yang W, Sun A, Wei Z, Lin Q. The Role of CXC Chemokines in Cancer Progression. Cancers. 2023; 15(1):167. https://doi.org/10.3390/cancers15010167
Chicago/Turabian StyleWu, Tiantian, Wannian Yang, Aiqin Sun, Zhixiao Wei, and Qiong Lin. 2023. "The Role of CXC Chemokines in Cancer Progression" Cancers 15, no. 1: 167. https://doi.org/10.3390/cancers15010167