
Blood Plasma Metabolome Profiling at Different Stages of Renal Cell Carcinoma

 , ,
, ,  ,
,  ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Subject Collection
2.2. Study Design
2.3. Sample Preparation
2.4. Metabolite Profiling
2.5. Mass Spectrum Processing
2.6. Statistical Analysis
2.7. Metabolite Annotation
2.8. Pathway Analysis
3. Results
3.1. Mass Spectrometry Analysis
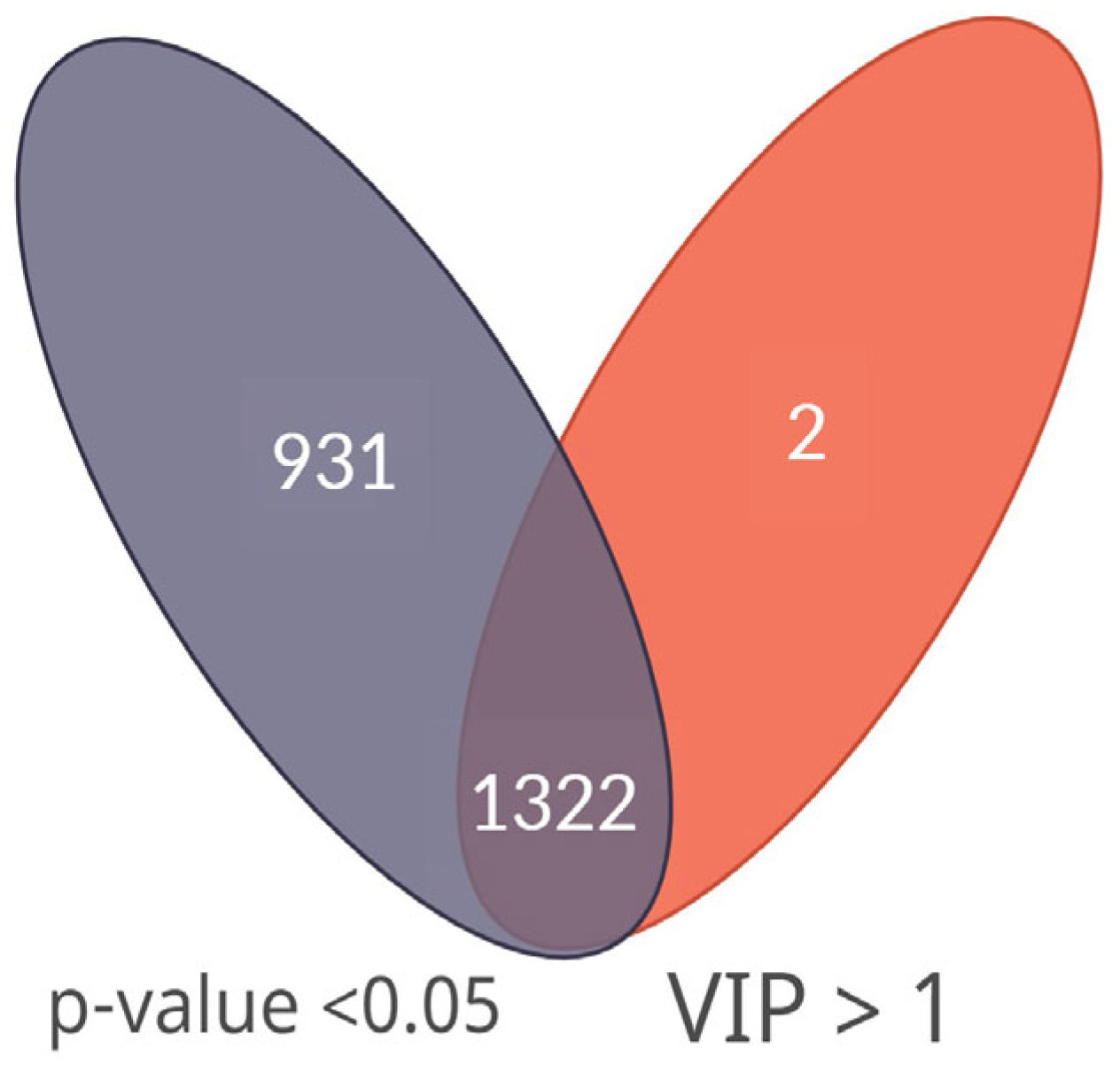
3.2. Statistical Analysis and Metabolite Annotation
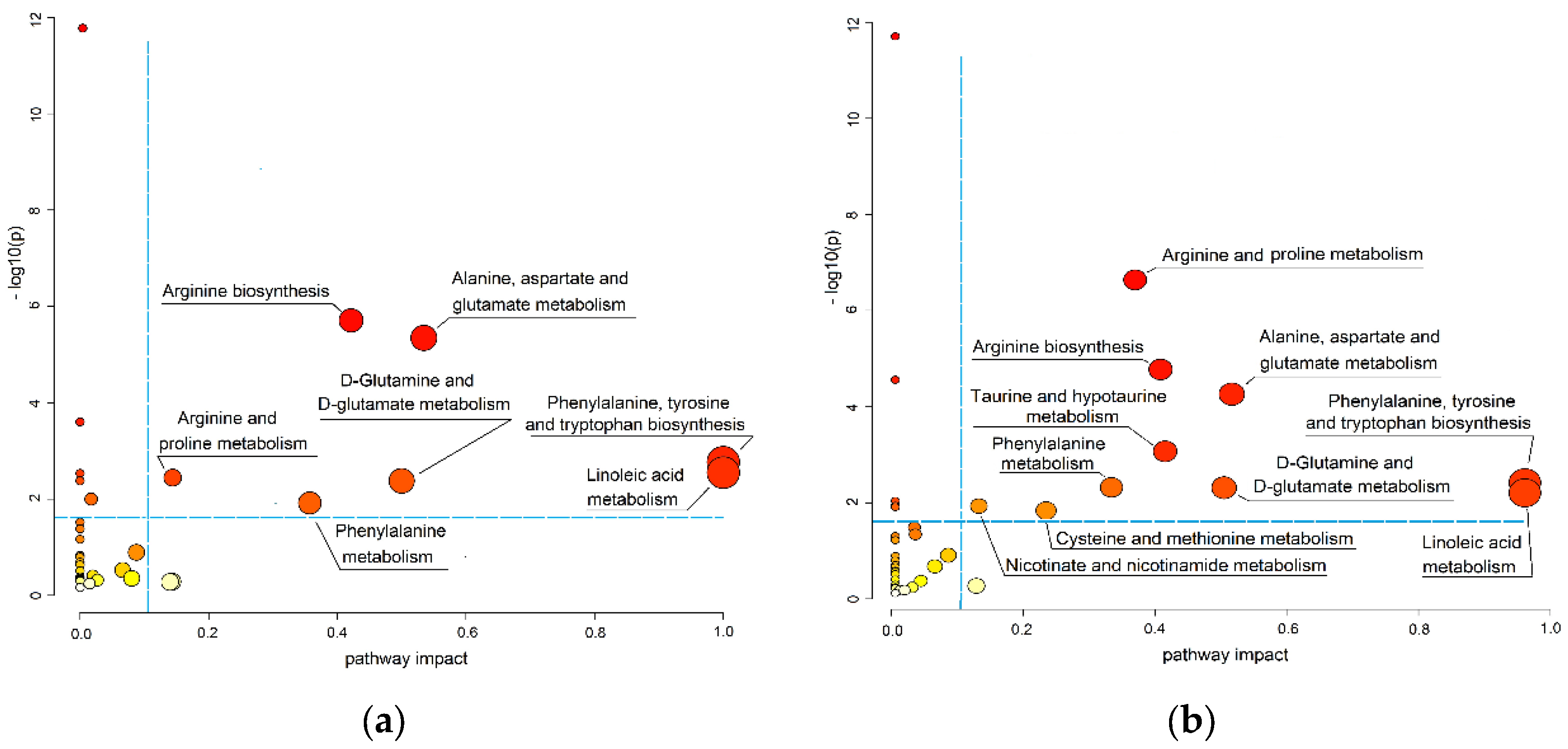
3.3. Pathways Associated with ccRCC Progression
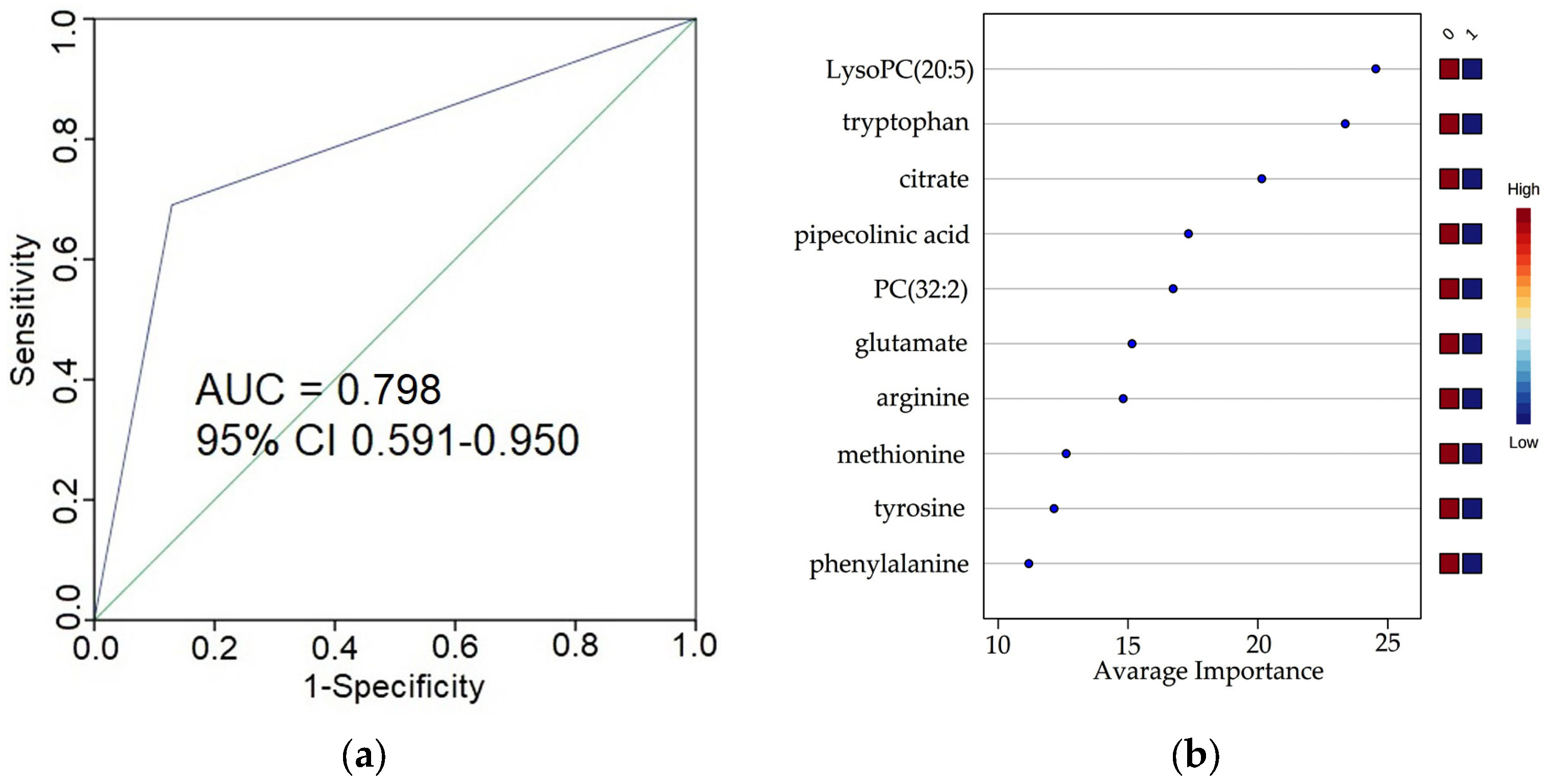
3.4. Predictive Power of the Selected Metabolites for Early Stages ccRCC
3.5. Evaluation of the Diagnostic Model for the ccRCC Advanced Stages
3.6. Evaluation of Specificity of RCC Diagnostic Model on Lung Cancer Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dagenais, G.R.; Leong, D.P.; Rangarajan, S.; Lanas, F.; Lopez-Jaramillo, P.; Gupta, R.; Diaz, R.; Avezum, A.; Oliveira, G.B.F.; Wielgosz, A.; et al. Variations in common diseases, hospital admissions, and deaths in middle-aged adults in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2020, 395, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Wei, E.Y.; Hsieh, J.J. A river model to map convergent cancer evolution and guide therapy in RCC. Nat. Rev. Urol. 2015, 12, 706–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljungberg, B.; Bensalah, K.; Canfield, S.; Dabestani, S.; Hofmann, F.; Hora, M.; Kuczyk, M.A.; Lam, T.; Marconi, L.; Merseburger, A.S.; et al. EAU guidelines on renal cell carcinoma: 2014 update. Eur. Urol. 2015, 67, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Apanovich, N.V.; Peters, M.V.; Korotaeva, A.A.; Apanovich, P.V.; Markova, A.S.; Kamolov, B.S.; Matveev, V.B.; Karpukhin, A.V. Molecular genetic diagnostics of clear cell renal cell carcinoma. Onkourologiya 2016, 12, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.; Gupta, S.; Hudes, G.R.; Egleston, B.L.; Uzzo, R.G.; Kruger, W.D. Serum amino acid levels as a biomarker for renal cell carcinoma. J. Urol. 2011, 186, 1206–1212. [Google Scholar] [CrossRef] [Green Version]
- Zisman, A.; Pantuck, A.J.; Wieder, J.; Chao, D.H.; Dorey, F.; Said, J.W.; DeKernion, J.B.; Figlin, R.A.; Belldegrun, A.S. Risk group assessment and clinical outcome algorithm to predict the natural history of patients with surgically resected renal cell carcinoma. J. Clin. Oncol. 2002, 20, 4559–4566. [Google Scholar] [CrossRef]
- Cochetti, G.; Cari, L.; Maulà, V.; Cagnani, R.; Paladini, A.; Del Zingaro, M.; Nocentini, G.; Mearini, E. Validation in an Independent Cohort of MiR-122, MiR-1271, and MiR-15b as Urinary Biomarkers for the Potential Early Diagnosis of Clear Cell Renal Cell Carcinoma. Cancers 2022, 14, 1112. [Google Scholar] [CrossRef]
- Gupta, A.; Sagar, G.; Siddiqui, Z.; Rao, K.V.S.; Nayak, S.; Saquib, N.; Anand, R. A non-invasive method for concurrent detection of early-stage women-specific cancers. Sci. Rep. 2022, 12, 2301. [Google Scholar] [CrossRef]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems biology and multi-omics integration: Viewpoints from the metabolomics research community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; Wittmann, B.M.; Evans, A.M.; Miller, L.A.D.; Toal, D.R.; Lonergan, S.; Elsea, S.H.; Pappan, K.L. Metabolomics in the clinic: A review of the shared and unique features of untargeted metabolomics for clinical research and clinical testing. J. Mass Spectrom. 2018, 53, 1143–1154. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Qi, Z.; Zhang, J.; Xue, W.H.; Li, L.F.; Shen, Z.B.; Li, Z.Y.; Yuan, Y.L.; Wang, W.B.; Zhao, J. Untargeted metabolomics analysis of esophageal squamous cell carcinoma discovers dysregulated metabolic pathways and potential diagnostic biomarkers. J. Cancer 2020, 11, 3944–3954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, D.R.; Patel, R.; Kirsch, D.G.; Lewis, C.A.; Vander Heiden, M.G.; Locasale, J.W. Metabolomics in cancer research and emerging applications in clinical oncology. CA Cancer J. Clin. 2021, 71, 333–358. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Soeters, M.R.; Wüst, R.C.I.; Houtkooper, R.H. Metabolic flexibility as an adaptation to energy resources and requirements in health and disease. Endocr. Rev. 2018, 39, 489–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piranavan, P.; Bhamra, M.; Perl, A. Metabolic Targets for Treatment of Autoimmune Diseases. Immunometabolism 2020, 2, e200012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, N.; Lanke, V.; Vinod, P.K. Network-based metabolic characterization of renal cell carcinoma. Sci. Rep. 2020, 10, 5955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.J.; Cheng, E.H. The panoramic view of clear cell renal cell carcinoma metabolism: Values of integrated global cancer metabolomics. Transl. Androl. Urol. 2016, 5, 984–986. [Google Scholar] [CrossRef] [Green Version]
- Mollica, V.; Santoni, M.; Matrana, M.R.; Basso, U.; De Giorgi, U.; Rizzo, A.; Maruzzo, M.; Marchetti, A.; Rosellini, M.; Bleve, S.; et al. Concomitant Proton Pump Inhibitors and Outcome of Patients Treated with Nivolumab Alone or Plus Ipilimumab for Advanced Renal Cell Carcinoma. Target. Oncol. 2022, 17, 61–68. [Google Scholar] [CrossRef]
- Rizzo, A.; Mollica, V.; Santoni, M.; Ricci, A.D.; Rosellini, M.; Marchetti, A.; Montironi, R.; Ardizzoni, A.; Massari, F. Impact of Clinicopathological Features on Survival in Patients Treated with First-line Immune Checkpoint Inhibitors Plus Tyrosine Kinase Inhibitors for Renal Cell Carcinoma: A Meta-analysis of Randomized Clinical Trials. Eur. Urol. Focus 2022, 8, 514–521. [Google Scholar] [CrossRef]
- Lokhov, P.G.; Balashova, E.E.; Trifonova, O.P.; Maslov, D.L.; Ponomarenko, E.A.; Archakov, A.I. Mass Spectrometry-Based Metabolomics Analysis of Obese Patients` Blood Plasma. Int. J. Mol. Sci. 2020, 21, 568. [Google Scholar] [CrossRef]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs—Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Lokhov, P.G.; Kharybin, O.N.; Archakov, A.I. Diagnosis of lung cancer based on direct-infusion electrospray mass spectrometry of blood plasma metabolites. Int. J. Mass Spectrom. 2012, 309, 200–205. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candille, G.; Talagrand, O. Evaluation of probabilistic prediction systems for a scalar variable. Q. J. R. Meteorol. Soc. 2005, 131, 2131–2150. [Google Scholar] [CrossRef]
- Blagus, R.; Lusa, L. Class prediction for high-dimensional class-imbalanced data. BMC Bioinform. 2010, 11, 523. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The human metabolome database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, A.; Hossain, L.; Leydesdorff, L. Betweenness centrality as a driver of preferential attachment in the evolution of research collaboration networks. J. Informetr. 2012, 6, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.A.M.; Graham, B.; Webb, K.; Ashton, L.V.; Harton, M.; Luetkemeyer, A.; Bokatzian, S.; Almubarak, R.; Mahapatra, S.; Hovind, L.; et al. A pilot metabolomics study of tuberculosis immune reconstitution inflammatory syndrome. Int. J. Infect. Dis. 2019, 84, 30–38. [Google Scholar] [CrossRef]
- Chen, Y.; Li, E.M.; Xu, L.Y. Guide to Metabolomics Analysis: A Bioinformatics Workflow. Metabolites 2022, 12, 357. [Google Scholar] [CrossRef] [PubMed]
- Sarvin, B.; Lagziel, S.; Sarvin, N.; Mukha, D.; Kumar, P.; Aizenshtein, E.; Shlomi, T. Fast and sensitive flow-injection mass spectrometry metabolomics by analyzing sample-specific ion distributions. Nat. Commun. 2020, 11, 3186. [Google Scholar] [CrossRef] [PubMed]
- Langford, V.S.; Perkins, M.J. Demystifying Sample Preparation for Headspace Analysis Using Direct Injection Mass Spectrometry. Column 2022, 18, 24–28. [Google Scholar]
- Lebrón-Aguilar, R.; Soria, A.C.; Quintanilla-López, J.E. Comprehensive evaluation of direct injection mass spectrometry for the quantitative profiling of volatiles in food samples. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southam, A.D.; Weber, R.J.M.; Engel, J.; Jones, M.R.; Viant, M. A complete workflow for high-resolution spectral-stitching nanoelectrospray direct-infusion mass-spectrometry-based metabolomics and lipidomics. Nat. Protoc. 2017, 12, 310–328. [Google Scholar] [CrossRef]
- Kirwan, J.A.; Weber, R.J.M.; Broadhurst, D.I.; Viant, M.R. Direct infusion mass spectrometry metabolomics dataset: A benchmark for data processing and quality control. Sci. Data 2014, 1, 140012. [Google Scholar] [CrossRef] [Green Version]
- Korangath, P.; Teo, W.W.; Sadik, H.; Han, L.; Mori, N.; Huijts, C.M.; Wildes, F.; Bharti, S.; Zhang, Z.; Santa-Maria, C.A.; et al. Targeting glutamine metabolism in breast cancer with aminooxyacetate. Clin. Cancer Res. 2015, 21, 3263–3273. [Google Scholar] [CrossRef] [Green Version]
- Green, C.R.; Wallace, M.; Divakaruni, A.S.; Phillips, S.A.; Murphy, A.N.; Ciaraldi, T.P.; Metallo, C.M. Branched-chain amino acid catabolism fuels adipocyte differentiation and lipogenesis. Nat. Chem. Biol. 2016, 12, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.A.; Vidoni, S.; Durigon, R.; Pearce, S.F.; Rorbach, J.; He, J.; Brea-Calvo, G.; Minczuk, M.; Reyes, A.; Holt, I.J.; et al. Amino acid starvation has opposite effects on mitochondrial and cytosolic protein synthesis. PLoS ONE 2014, 9, e93597. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, A.E.; Deshmukh, M. Glucose metabolism inhibits apoptosis in neurons and cancer cells by redox inactivation of cytochrome c. Nat. Cell Biol. 2008, 10, 1477–1483. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Plass, C.; Gerhauser, C. Cancer Chemoprevention by Targeting the Epigenome. Curr. Drug Targets 2011, 12, 1925–1956. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, Y.; Higashiyama, M.; Gochi, A.; Akaike, M.; Ishikawa, T.; Miura, T.; Saruki, N.; Bando, E.; Kimura, H.; Imamura, F.; et al. Plasma free amino acid profiling of five types of cancer patients and its application for early detection. PLoS ONE 2011, 6, e24143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucarelli, G.; Loizzo, D.; Franzin, R.; Battaglia, S.; Ferro, M.; Cantiello, F.; Castellano, G.; Bettocchi, C.; Ditonno, P.; Battaglia, M. Metabolomic insights into pathophysiological mechanisms and biomarker discovery in clear cell renal cell carcinoma. Expert Rev. Mol. Diagn. 2019, 19, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Tan, V.Y.; Corbin, L.J.; Smith-Byrne, K.; Alcala, K.; Langenberg, C.; Stewart, I.D.; Butterworth, A.S.; Surendran, P.; Achaintre, D.; et al. The blood metabolome of incident kidney cancer: A case-control study nested within the MetKid consortium. PLoS Med. 2021, 18, e1003786. [Google Scholar] [CrossRef]
- Jing, L.; Guigonis, J.M.; Borchiellini, D.; Durand, M.; Pourcher, T.; Ambrosetti, D. LC-MS based metabolomic profiling for renal cell carcinoma histologic subtypes. Sci. Rep. 2019, 9, 15635. [Google Scholar] [CrossRef] [Green Version]
- Nizioł, J.; Bonifay, V.; Ossoliński, K.; Ossoliński, T.; Ossolińska, A.; Sunner, J.; Beech, I.; Arendowski, A.; Ruman, T. Metabolomic study of human tissue and urine in clear cell renal carcinoma by LC-HRMS and PLS-DA. Anal. Bioanal. Chem. 2018, 410, 3859–3869. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, M.; Liu, X.; Sun, H.; Guo, Z.; Tang, X.; Wang, Z.; Li, J.; He, L.; Zhang, W.; et al. Investigation of Plasma Metabolic and Lipidomic Characteristics of a Chinese Cohort and a Pilot Study of Renal Cell Carcinoma Biomarker. Front. Oncol. 2020, 10, 1507. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, M.; Cheng, X.; Liu, X.; Sun, H.; Guo, Z.; Li, J.; Tang, X.; Wang, Z.; Sun, W.; et al. LC-MS-Based Plasma Metabolomics and Lipidomics Analyses for Differential Diagnosis of Bladder Cancer and Renal Cell Carcinoma. Front Oncol. 2020, 15, 717. [Google Scholar] [CrossRef]
- Falegan, O.S.; Ball, M.W.; Shaykhutdinov, R.A.; Pieroraio, P.M.; Farshidfar, F.; Vogel, H.J.; Allaf, M.E.; Hyndman, M.E. Urine and serum metabolomics analyses may distinguish between stages of renal cell carcinoma. Metabolites 2017, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Dong, B.; Liu, X.; Xuan, H.; Huang, Y.; Lin, D. Metabonomic profiling of renal cell carcinoma: High-resolution proton nuclear magnetic resonance spectroscopy of human serum with multivariate data analysis. Anal. Chim. Acta 2008, 624, 269–277. [Google Scholar] [CrossRef]
- Nizioł, J.; Ossoliński, K.; Tripet, B.P.; Copié, V.; Arendowski, A.; Ruman, T. Nuclear magnetic resonance and surface-assisted laser desorption/ionization mass spectrometry-based metabolome profiling of urine samples from kidney cancer patients. J. Pharm. Biomed. Anal. 2021, 193, 113752. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kawasaki, Y.; Maekawa, M.; Takasaki, S.; Shimada, S.; Morozumi, K.; Sato, M.; Kawamorita, N.; Yamashita, S.; Mitsuzuka, K.; et al. Accurate quantification of urinary metabolites for predictive models manifest clinicopathology of renal cell carcinoma. Cancer Sci. 2020, 111, 2570–2578. [Google Scholar] [CrossRef] [PubMed]
- Bifarin, O.O.; Gaul, D.A.; Sah, S.; Arnold, R.S.; Ogan, K.; Master, V.A.; Roberts, D.L.; Bergquist, S.H.; Petros, J.A.; Edison, A.S.; et al. Urine-based metabolomics and machine learning reveals metabolites associated with renal cell carcinoma stage. Cancers 2021, 13, 6253. [Google Scholar] [CrossRef] [PubMed]
- Skene, D.J.; Middleton, B.; Fraser, C.K.; Pennings, J.L.A.; Kuchel, T.R.; Rudiger, S.R.; Bawden, C.S.; Morton, A.J. Metabolic profiling of presymptomatic Huntington’s disease sheep reveals novel biomarkers. Sci. Rep. 2017, 7, 43030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Hasim, A.; Mamtimin, B.; Kong, B.; Zhang, H.P.; Sheyhidin, I. Plasma free amino acid profiling of esophageal cancer using high-performance liquid chromatography spectroscopy. World J. Gastroenterol. 2014, 20, 8653–8659. [Google Scholar] [CrossRef]
- Ihata, Y.; Miyagi, E.; Numazaki, R.; Muramatsu, T.; Imaizumi, A.; Yamamoto, H.; Yamakado, M.; Okamoto, N.; Hirahara, F. Amino acid profile index for early detection of endometrial cancer: Verification as a novel diagnostic marker. Int. J. Clin. Oncol. 2014, 19, 364–372. [Google Scholar] [CrossRef]
- Lin, L.; Huang, Z.; Gao, Y.; Chen, Y.; Hang, W.; Xing, J.; Yan, X. LC-MS-based serum metabolic profiling for genitourinary cancer classification and cancer type-specific biomarker discovery. Proteomics 2012, 12, 2238–2246. [Google Scholar] [CrossRef]
- Du, T.; Han, J. Arginine Metabolism and Its Potential in Treatment of Colorectal Cancer. Front. Cell Dev. Biol. 2021, 9, 658861. [Google Scholar] [CrossRef]
- Matos, A.; Carvalho, M.; Bicho, M.; Ribeiro, R. Arginine and arginases modulate metabolism, tumor microenvironment and prostate cancer progression. Nutrients 2021, 13, 4503. [Google Scholar] [CrossRef]
- Li, C.; Zhao, H. Tryptophan and Its Metabolites in Lung Cancer: Basic Functions and Clinical Significance. Front. Oncol. 2021, 11, 707277. [Google Scholar] [CrossRef]
- Ayob, A.Z.; Ramasamy, T.S. Cancer stem cells as key drivers of tumour progression. J. Biomed. Sci. 2018, 25, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proenza, A.M.; Oliver, J.; Palou, A.; Roca, P. Breast and lung cancer are associated with a decrease in blood cell amino acid content. J. Nutr. Biochem. 2003, 14, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Pian, C.; Xu, M.; Kong, L.; Li, Z.; Ji, J.; Chen, Y.; Zhang, L. Revealing prognosis-related pathways at the individual level by a comprehensive analysis of different cancer transcription data. Genes 2020, 11, 1281. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Ji, J.; Zhao, L.; Chen, M.; Shi, A.; Pan, L.; Huang, Y.; Zhang, H.; Dong, B.; Gao, H. Prediction and diagnosis of renal cell carcinoma using nuclear magnetic resonance-based serum metabolomics and self-organizing maps. Oncotarget 2016, 7, 59189–59198. [Google Scholar] [CrossRef] [Green Version]
- Zira, A.N.; Theocharis, S.E.; Mitropoulos, D.; Migdalis, V.; Mikros, E. 1H NMR metabonomic analysis in renal cell carcinoma: A possible diagnostic tool. J. Proteome Res. 2010, 9, 4038–4044. [Google Scholar] [CrossRef]
- Reznik, E.; Luna, A.; Aksoy, B.A.; Liu, E.M.; La, K.; Ostrovnaya, I.; Creighton, C.J.; Hakimi, A.A.; Sander, C. A Landscape of Metabolic Variation across Tumor Types. Cell Syst. 2018, 6, 303–313. [Google Scholar] [CrossRef]
- He, L.; Chen, J.; Xu, F.; Li, J. Prognostic Implication of a Metabolism-Associated Gene Signature in Lung Adenocarcinoma. Mol. Ther. Oncolytics 2020, 19, 265–277. [Google Scholar] [CrossRef]
- Nizioł, J.; Ossoliński, K.; Tripet, B.P.; Copié, V.; Arendowski, A.; Ruman, T. Nuclear magnetic resonance and surface-assisted laser desorption/ionization mass spectrometry-based serum metabolomics of kidney cancer. Anal. Bioanal. Chem. 2020, 412, 5827–5841. [Google Scholar] [CrossRef]
- Deja, S.; Litarski, A.; Mielko, K.A.; Pudełko-Malik, N.; Wojtowicz, W.; Zabek, A.; Szydełko, T.; Młynarz, P. Gender-Specific Metabolomics Approach to Kidney Cancer. Metabolites 2021, 10, 767. [Google Scholar] [CrossRef]
- Desaire, H. How (Not) to Generate a Highly Predictive Biomarker Panel Using Machine Learning. J. Proteome Res. 2022, 21, 2071–2074. [Google Scholar] [CrossRef]
- Broadhurst, D.I.; Kell, D.B. Statistical strategies for avoiding false discoveries in metabolomics and related experiments. Metabolomics 2006, 2, 171–196. [Google Scholar] [CrossRef]
- Ransohoff, D.F. Rules of evidence for cancer molecular-marker discovery and validation. Nat. Rev. Cancer 2004, 4, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Liu, X.; Sun, H.; Guo, Z.; Tang, X.; Wang, Z.; Li, J.; Li, H.; Sun, W.; et al. Urine Metabolomics for Renal Cell Carcinoma (RCC) Prediction: Tryptophan Metabolism as an Important Pathway in RCC. Front. Oncol. 2019, 17, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, B.; Schafferer, S.; Tang, Q.; Scheffler, M.; Nees, J.; Heil, J.; Schott, S.; Golatta, M.; Wallwiener, M.; Sohn, C.; et al. A plasma metabolite panel as biomarkers for early primary breast cancer detection. Int. J. Cancer 2019, 144, 2833–2842. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort | Total Number | Age (years) | BMI (kg/m2) | Male | Female | Male/Female | ||
|---|---|---|---|---|---|---|---|---|
| Number | Age (years) | Number | Age (years) | |||||
| Control | 51 | 56.5 ± 7.7 1 | 30.5 ± 2.7 1 | 23 | 52.5 ± 6.9 | 28 | 59.1 ± 7.1 | 45%/55% |
| ccRCC (I–II stages) | 39 | 60.0 ± 7.9 | 32.6 ± 3.4 | 18 | 57.1 ± 9.1 | 21 | 62.0 ± 6.5 | 46%/54% |
| ccRCC (III–IV stages) | 22 | 58.3 ± 7.0 | 31.2 ± 2.8 | 17 | 57.5 ± 7.5 | 5 | 61.3 ± 4.1 | 77%/23% |
| pRCC and chrRCC (I–II stages) | 12 | 58.2 ± 10.2 | 32.1 ± 2.6 | 4 | 57.3 ± 11.8 | 8 | 58.6 ± 9.4 | 33%/67% |
| Lung cancer 2 (I–II stages) | 25 | 61.6 ± 4.2 | 30.3 ± 2.8 | 16 | 60.4 ± 3.5 | 9 | 62.3 ± 5.4 | 64%/36% |
| Controls | Cancer Patients | |
|---|---|---|
| non-cancer volunteers | vs | patients with ccRCC (I–II stages) |
| patients with ccRCC (III–IV stages) | ||
| patients with pRCC and chrRCC (I–II stages) | ||
| № | Pathway Name 1 | Total | Hits | p-Value | −log(p) | Impact |
|---|---|---|---|---|---|---|
| ccRCC and pRCC/chrRCC patients (early stages) | ||||||
| 1 | Aminoacyl-tRNA biosynthesis | 48 | 13 | 6.29 × 10−14 | 13.20 | 0.00 |
| 2 | Arginine biosynthesiss 2 | 14 | 5 | 1.96 × 10−6 | 5.71 | 0.42 |
| 3 | Alanine, aspartate, and glutamate metabolism 2 | 28 | 6 | 4.51 × 10−6 | 5.35 | 0.53 |
| 4 | Valine, leucine, and isoleucine biosynthesis | 8 | 3 | 2.49 × 10−4 | 3.60 | 0.00 |
| 5 | Phenylalanine, tyrosine, and tryptophan biosynthesis 2 | 4 | 2 | 1.72 × 10−3 | 2.76 | 1.00 |
| 6 | Linoleic acid metabolism 2 | 5 | 2 | 2.83 × 10−3 | 2.55 | 1.00 |
| 7 | Biosynthesis of unsaturated fatty acids | 36 | 4 | 2.94 × 10−3 | 2.53 | 0.00 |
| 8 | Arginine and proline metabolism 2 | 38 | 4 | 3.60 × 10−3 | 2.44 | 0.14 |
| 9 | Nitrogen metabolism | 6 | 2 | 4.20 × 10−3 | 2.38 | 0.00 |
| 10 | D-Glutamine and D-glutamate metabolism 2 | 6 | 2 | 4.20 × 10−3 | 2.38 | 0.50 |
| 11 | Phenylalanine metabolism 2 | 10 | 2 | 1.21 × 10−2 | 1.92 | 0.36 |
| 12 | Glyoxylate and dicarboxylate metabolism | 32 | 3 | 1.67 × 10−2 | 1.78 | 0.03 |
| 13 | Histidine metabolism | 16 | 2 | 3.02 × 10−2 | 1.52 | 0.00 |
| 14 | Lysine degradation | 25 | 2 | 6.85 × 10−2 | 1.16 | 0.00 |
| 15 | Glycerophospholipids metabolism | 36 | 2 | 1.28 × 10−1 | 0.89 | 0.11 |
| ccRCC patients (advanced stages) | ||||||
| 16 | Aminoacyl-tRNA biosynthesis | 48 | 14 | 1.29 × 10−12 | 11.89 | 0.00 |
| 17 | Arginine and proline metabolism 2 | 38 | 9 | 1.88 × 10−7 | 6.73 | 0.38 |
| 18 | Arginine biosynthesis 2 | 14 | 5 | 1.50 × 10−5 | 4.82 | 0.42 |
| 19 | Valine, leucine, and isoleucine biosynthesis | 8 | 4 | 2.47 × 10−5 | 4.61 | 0.00 |
| 20 | Alanine, aspartate, and glutamate metabolism 2 | 28 | 6 | 4.98 × 10−5 | 4.30 | 0.53 |
| 21 | Taurine and hypotaurine metabolism 2 | 8 | 3 | 8.16 × 10−4 | 3.09 | 0.43 |
| 22 | Phenylalanine, tyrosine, and tryptophan biosynthesis 2 | 4 | 2 | 3.77 × 10−3 | 2.42 | 1.00 |
| 23 | Linoleic acid metabolism 2 | 5 | 2 | 6.18 × 10−3 | 2.21 | 1.00 |
| 24 | Nitrogen metabolism | 6 | 2 | 9.12 × 10−3 | 2.04 | 0.00 |
| 25 | D-Glutamine and D-glutamate metabolism 2 | 6 | 2 | 9.13 × 10−3 | 2.04 | 0.50 |
| 26 | Biosynthesis of unsaturated fatty acids | 36 | 4 | 1.23 × 10−2 | 1.91 | 0.00 |
| 27 | Phenylalanine metabolism 2 | 10 | 2 | 2.56 × 10−2 | 1.59 | 0.36 |
| 28 | Glutathione metabolism | 28 | 3 | 3.33 × 10−2 | 1.47 | 0.03 |
| 29 | Glyoxylate and dicarboxylate metabolism | 32 | 3 | 4.47 × 10−2 | 1.35 | 0.03 |
| 30 | Glycine, serine, and threonine metabolism | 33 | 3 | 4.77 × 10−2 | 1.32 | 0.00 |
| 31 | Cysteine and methionine metabolism 2 | 33 | 3 | 4.77 × 10−2 | 1.32 | 0.26 |
| 32 | Nicotinate and nicotinamide metabolism 2 | 14 | 2 | 4.95 × 10−2 | 1.27 | 0.14 |
| 33 | Histidine metabolism | 16 | 2 | 6.20 × 10−2 | 1.21 | 0.00 |
| 34 | Lysine degradation | 25 | 2 | 1.34 × 10−1 | 0.87 | 0.00 |
| Metabolites | Pathway Name 1 |
|---|---|
| pipecolinic acid | arginine and proline metabolism |
| glutamate | alanine, aspartate, and glutamate metabolism |
| arginine biosynthesis | |
| arginine and proline metabolism | |
| glutamine and glutamate metabolism | |
| methionine | cysteine and methionine metabolism |
| arginine | arginine biosynthesis |
| arginine and proline metabolism | |
| tyrosine | phenylalanine, tyrosine, and tryptophanbiosynthesis |
| phenylalanine metabolism | |
| phenylalanine | phenylalanine, tyrosine, and tryptophanbiosynthesis |
| phenylalanine metabolism | |
| tryptophan | phenylalanine, tyrosine, and tryptophanbiosynthesis |
| citrate | alanine, aspartate, and glutamate metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maslov, D.L.; Trifonova, O.P.; Lichtenberg, S.; Balashova, E.E.; Mamedli, Z.Z.; Alferov, A.A.; Stilidi, I.S.; Lokhov, P.G.; Kushlinskii, N.E.; Archakov, A.I. Blood Plasma Metabolome Profiling at Different Stages of Renal Cell Carcinoma. Cancers 2023, 15, 140. https://doi.org/10.3390/cancers15010140
Maslov DL, Trifonova OP, Lichtenberg S, Balashova EE, Mamedli ZZ, Alferov AA, Stilidi IS, Lokhov PG, Kushlinskii NE, Archakov AI. Blood Plasma Metabolome Profiling at Different Stages of Renal Cell Carcinoma. Cancers. 2023; 15(1):140. https://doi.org/10.3390/cancers15010140
Chicago/Turabian StyleMaslov, Dmitry L., Oxana P. Trifonova, Steven Lichtenberg, Elena E. Balashova, Zaman Z. Mamedli, Aleksandr A. Alferov, Ivan S. Stilidi, Petr G. Lokhov, Nikolay E. Kushlinskii, and Alexander I. Archakov. 2023. "Blood Plasma Metabolome Profiling at Different Stages of Renal Cell Carcinoma" Cancers 15, no. 1: 140. https://doi.org/10.3390/cancers15010140