
Value of c-MET and Associated Signaling Elements for Predicting Outcomes and Targeted Therapy in Penile Cancer

 , , , , , ,
, , , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. Tissue Microarray Construction and Immunohistochemical Staining
2.3. Cell Culture and Induction of Drug Resistance
2.4. Drugs
2.5. Tumor Cell Growth
2.6. SDS-PAGE/Western Blot
2.7. Statistics
3. Results
3.1. Clinicopathological Features of Patients
3.2. Expression of Protein Candidates in the Tumor Tissue
3.3. Biomarker Correlations
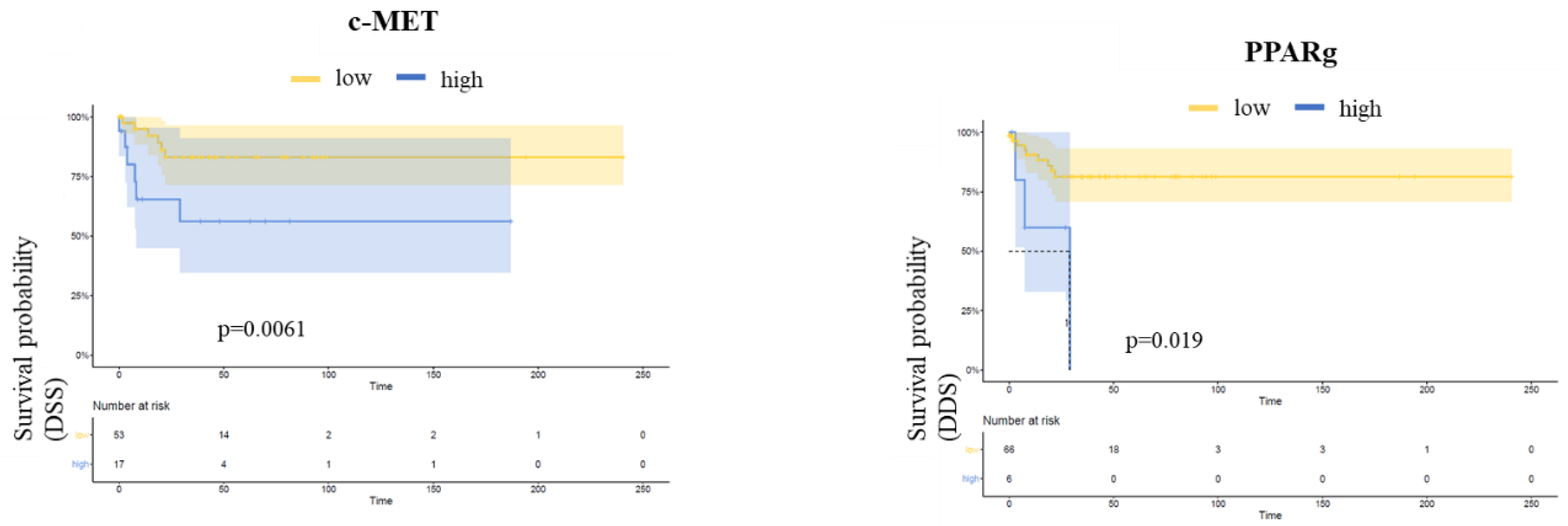
3.4. Follow-Up
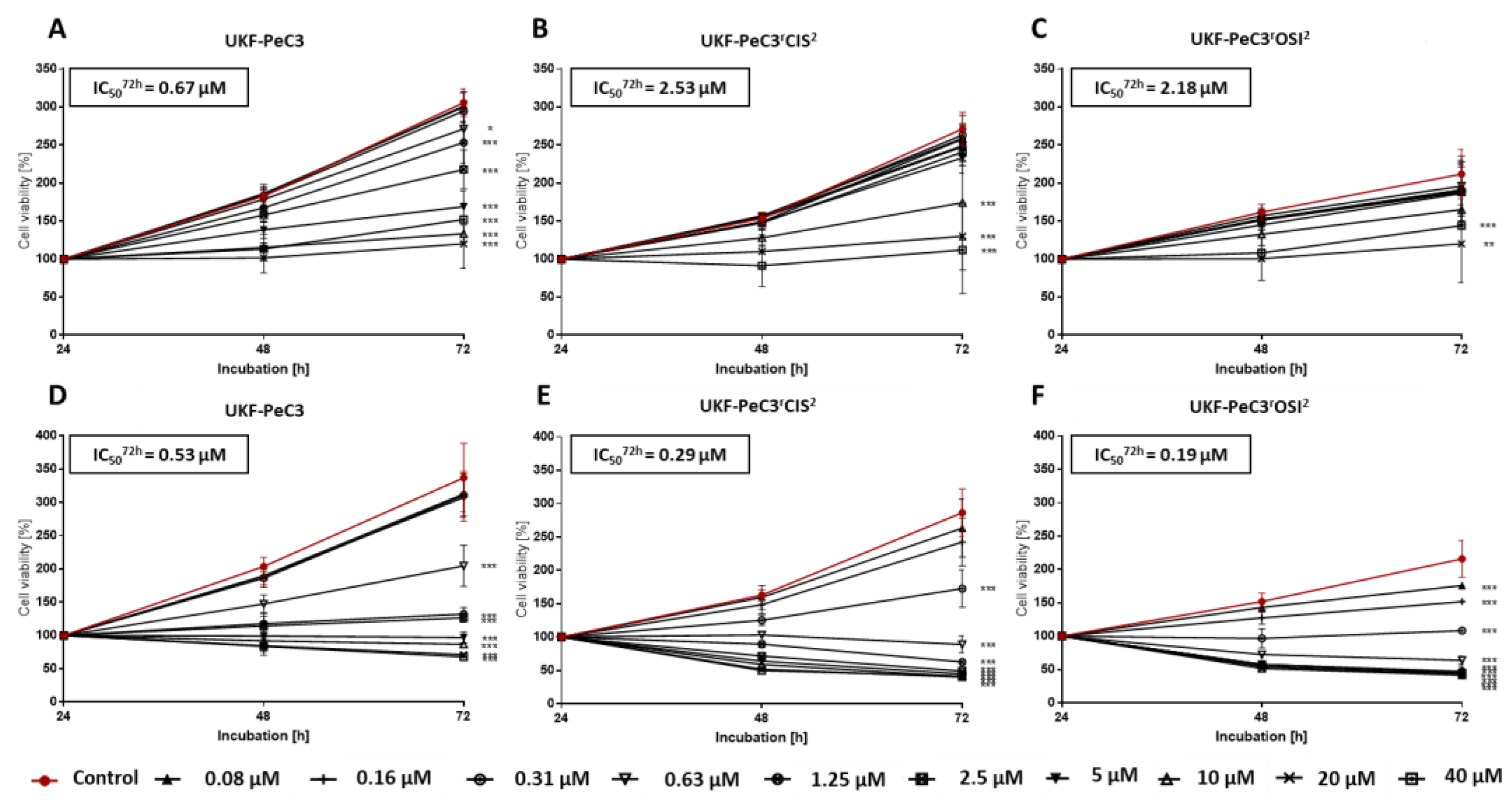
3.5. Induction of Resistance to Cisplatin and Osimertinib
3.6. Expression of Protein Candidates in Cell Lines
3.7. Modulation of Tumor Cell Growth by c-MET Inhibition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, Y.T.; Tan, Y.J.; Oon, C.E. Molecular targeted therapy: Treating cancer with specificity. Eur. J. Pharmacol. 2018, 834, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, M.E.; Ryder, M.; Jimenez, C. Targeted Therapy for Advanced Thyroid Cancer: Kinase Inhibitors and Beyond. Endocr. Rev. 2019, 40, 1573–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barata, P.C.; Rini, B.I. Treatment of renal cell carcinoma: Current status and future directions. CA Cancer J. Clin. 2017, 67, 507–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, J.E.; O’Donnell, P.H.; Balar, A.V.; McGregor, B.A.; Heath, E.I.; Yu, E.Y.; Galsky, M.D.; Hahn, N.M.; Gartner, E.M.; Pinelli, J.M.; et al. Pivotal Trial of Enfortumab Vedotin in Urothelial Carcinoma After Platinum and Anti-Programmed Death 1/Programmed Death Ligand 1 Therapy. J. Clin. Oncol. 2019, 37, 2592–2600. [Google Scholar] [CrossRef]
- Tagawa, S.T.; Balar, A.V.; Petrylak, D.P.; Kalebasty, A.R.; Loriot, Y.; Flechon, A.; Jain, R.K.; Agarwal, N.; Bupathi, M.; Barthelemy, P.; et al. TROPHY-U-01: A Phase II Open-Label Study of Sacituzumab Govitecan in Patients With Metastatic Urothelial Carcinoma Progressing After Platinum-Based Chemotherapy and Checkpoint Inhibitors. J. Clin. Oncol. 2021, 39, 2474–2485. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; do Canto Alvim, L.M.; Rainho, C.A.; Juengel, E.; Blaheta, R.A.; Spiess, P.E.; Rogatto, S.R.; Tsaur, I. Systemic treatment of penile squamous cell carcinoma-hurdles and hopes of preclinical models and clinical regimens: A narrative review. Transl. Androl. Urol. 2021, 10, 4085–4098. [Google Scholar] [CrossRef] [PubMed]
- Christodoulidou, M.; Sahdev, V.; Houssein, S.; Muneer, A. Epidemiology of penile cancer. Curr. Probl. Cancer 2015, 39, 126–136. [Google Scholar] [CrossRef]
- Pagliaro, L.C.; Williams, D.L.; Daliani, D.; Williams, M.B.; Osai, W.; Kincaid, M.; Wen, S.; Thall, P.F.; Pettaway, C.A. Neoadjuvant paclitaxel, ifosfamide, and cisplatin chemotherapy for metastatic penile cancer: A phase II study. J. Clin. Oncol. 2010, 28, 3851–3857. [Google Scholar] [CrossRef]
- Eder, J.P.; Vande Woude, G.F.; Boerner, S.A.; LoRusso, P.M. Novel therapeutic inhibitors of the c-Met signaling pathway in cancer. Clin. Cancer Res. 2009, 15, 2207–2214. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Newton, R.C.; Scherle, P.A. Developing c-MET pathway inhibitors for cancer therapy: Progress and challenges. Trends Mol. Med. 2010, 16, 37–45. [Google Scholar] [CrossRef]
- Thomas, A.; Reetz, S.; Stenzel, P.; Tagscherer, K.; Roth, W.; Schindeldecker, M.; Michaelis, M.; Rothweiler, F.; Cinatl, J., Jr.; Cinatl, J.; et al. Assessment of PI3K/mTOR/AKT Pathway Elements to Serve as Biomarkers and Therapeutic Targets in Penile Cancer. Cancers 2021, 13, 2323. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Kar, M.; Roy, S.; Saha, A.; Padhi, S.; Banerjee, B. Role of beta-catenin in cisplatin resistance, relapse and prognosis of head and neck squamous cell carcinoma. Cell. Oncol. 2018, 41, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Miyazaki, Y.; Hiraoka, S.; Toyota, M.; Nagasawa, Y.; Kondo, S.; Kiyohara, T.; Shinomura, Y.; Matsuzawa, Y. PPARgamma inhibits the expression of c-MET in human gastric cancer cells through the suppression of Ets. Biochem. Biophys. Res. Commun. 1999, 265, 453–456. [Google Scholar] [CrossRef]
- Berger, A.; Brady, N.J.; Bareja, R.; Robinson, B.; Conteduca, V.; Augello, M.A.; Puca, L.; Ahmed, A.; Dardenne, E.; Lu, X.; et al. N-Myc-mediated epigenetic reprogramming drives lineage plasticity in advanced prostate cancer. J. Clin. Investig. 2019, 129, 3924–3940. [Google Scholar] [CrossRef]
- Khan, S.A.; Burke, M.; Zhu, F.; Yang, D.H.; Dubyk, C.; Mehra, R.; Lango, M.J.; Ridge, J.A.; Sher, D.J.; Burtness, B. Survivin expression and impact on head and neck cancer outcomes. Oral Oncol. 2021, 112, 105049. [Google Scholar] [CrossRef]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar]
- Hoo, Z.H.; Candlish, J.; Teare, D. What is an ROC curve? Emerg. Med. J. 2017, 34, 357–359. [Google Scholar] [CrossRef]
- Michaelis, M.; Wass, M.; Cinatl, J. Drug-adapted cancer cell lines as preclinical models of acquired resistance. Cancer Drug Resist. 2019, 2, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Kotchetkov, R.; Cinatl, J.; Blaheta, R.; Vogel, J.U.; Karaskova, J.; Squire, J.; Driever, P.H.; Klingebiel, T.; Cinatl, J., Jr. Development of resistance to vincristine and doxorubicin in neuroblastoma alters malignant properties and induces additional karyotype changes: A preclinical model. Int. J. Cancer 2003, 104, 36–43. [Google Scholar] [CrossRef]
- Lien, V.T.; Celen, S.; Nuruddin, S.; Attili, B.; Doumont, G.; Van Simaeys, G.; Bormans, G.; Klaveness, J.; Olberg, D.E. Preclinical evaluation of [(18)F] cabozantinib as a PET imaging agent in a prostate cancer mouse model. Nucl. Med. Biol. 2021, 93, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Xie, H.; Dou, G.; Yang, W.; Ge, J.; Zhou, B.; Ren, J.; Li, J.; Wang, J.; Zhang, Z.; et al. Computational study on novel natural inhibitors targeting c-MET. Medicine 2021, 100, e27171. [Google Scholar] [CrossRef] [PubMed]
- Comoglio, P.M.; Trusolino, L.; Boccaccio, C. Known and novel roles of the MET oncogene in cancer: A coherent approach to targeted therapy. Nat. Rev. Cancer 2018, 18, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Monga, S.P.; Mars, W.M.; Pediaditakis, P.; Bell, A.; Mule, K.; Bowen, W.C.; Wang, X.; Zarnegar, R.; Michalopoulos, G.K. Hepatocyte growth factor induces Wnt-independent nuclear translocation of beta-catenin after Met-beta-catenin dissociation in hepatocytes. Cancer Res. 2002, 62, 2064–2071. [Google Scholar] [PubMed]
- Vries, H.M.; Bekers, E.; van Oosterom, M.N.; Karakullukcu, M.B.; van Oosterom, M.N.; Poel, D.; van Leeuwen, F.W.B.; Buckle, T.; Brouwer, O.R. c-MET Receptor-Targeted Fluorescence on the Road to Image-Guided Surgery in Penile Squamous Cell Carcinoma Patients. J. Nucl. Med. 2022, 63, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Gunia, S.; Erbersdobler, A.; Hakenberg, O.W.; Koch, S.; May, M. C-MET is expressed in the majority of penile squamous cell carcinomas and correlates with polysomy-7 but is not associated with MET oncogene amplification, pertinent histopathologic parameters, or with cancer-specific survival. Pathol. Res. Pract. 2013, 209, 215–220. [Google Scholar] [CrossRef]
- Szturz, P.; Budikova, M.; Vermorken, J.B.; Horova, I.; Gal, B.; Raymond, E.; de Gramont, A.; Faivre, S. Prognostic value of c-MET in head and neck cancer: A systematic review and meta-analysis of aggregate data. Oral. Oncol. 2017, 74, 68–76. [Google Scholar] [CrossRef]
- Dotto, G.P.; Rustgi, A.K. Squamous Cell Cancers: A Unified Perspective on Biology and Genetics. Cancer Cell 2016, 29, 622–637. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.L.; Wu, H.F.; Wang, W.J.; Hu, G.M.; Gu, B.; Zhang, M.; Wang, Y.X. C-Met as a potential novel prognostic marker in squamous cell carcinoma and adenocarcinoma of esophagus: Evidence from a meta-analysis. Panminerva Med. 2017, 59, 97–106. [Google Scholar] [CrossRef]
- Thomas, A.; Necchi, A.; Muneer, A.; Tobias-Machado, M.; Tran, A.T.H.; Van Rompuy, A.S.; Spiess, P.E.; Albersen, M. Penile cancer. Nat. Rev. Dis. Primers 2021, 7, 11. [Google Scholar] [CrossRef]
- Gu, W.; Zhu, Y.; Ye, D. Beyond chemotherapy for advanced disease-the role of EGFR and PD-1 inhibitors. Transl. Androl. Urol. 2017, 6, 848–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, G.; Buonerba, C.; Ferro, M.; Calderoni, G.; Bozza, G.; Federico, P.; Tedesco, B.; Ruggieri, V.; Aieta, M. The epidermal growth factor receptors as biological targets in penile cancer. Expert Opin. Biol. Ther. 2015, 15, 473–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.B.; Liu, R.Y.; Peng, Q.H.; Li, Z.S.; Jiang, L.J.; Guo, S.J.; Zhou, Q.H.; Liu, T.Y.; Deng, C.Z.; Yao, K.; et al. EGFR mono-antibody salvage therapy for locally advanced and distant metastatic penile cancer: Clinical outcomes and genetic analysis. Urol. Oncol. 2019, 37, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Remon, J.; Steuer, C.E.; Ramalingam, S.S.; Felip, E. Osimertinib and other third-generation EGFR TKI in EGFR-mutant NSCLC patients. Ann. Oncol. 2018, 29, i20–i27. [Google Scholar] [CrossRef]
- Li, E.; Hu, Z.; Sun, Y.; Zhou, Q.; Yang, B.; Zhang, Z.; Cao, W. Small molecule inhibitor of c-Met (PHA665752) suppresses the growth of ovarian cancer cells and reverses cisplatin resistance. Tumour Biol. 2016, 37, 7843–7852. [Google Scholar] [CrossRef]
- Zhu, K.; Lv, Z.; Xiong, J.; Zheng, H.; Zhang, S.; Jin, H.; Yu, L.; Li, Z.; Zhang, J.; Li, C.; et al. MET inhibitor, capmatinib overcomes osimertinib resistance via suppression of MET/Akt/snail signaling in non-small cell lung cancer and decreased generation of cancer-associated fibroblasts. Aging 2021, 13, 6890–6903. [Google Scholar] [CrossRef]
- Rathi, N.; Maughan, B.L.; Agarwal, N.; Swami, U. Mini-Review: Cabozantinib in the Treatment of Advanced Renal Cell Carcinoma and Hepatocellular Carcinoma. Cancer Manag. Res. 2020, 12, 3741–3749. [Google Scholar] [CrossRef]
- Necchi, A.; Mariani, L.; Colecchia, M.; Giannatempo, P.; Raggi, D.; Calareso, G.; Nicolai, N.; Catanzaro, M.; Torelli, T.; Perrone, F.; et al. 927TiP—Cabozantinib in patients with advanced penile squamous cell carcinoma (PSCC): The open-label, single-arm, single-center, phase 2, CaboPen trial. Ann. Oncol. 2017, 28, v329. [Google Scholar] [CrossRef]
- Munshi, N.; Jeay, S.; Li, Y.; Chen, C.R.; France, D.S.; Ashwell, M.A.; Hill, J.; Moussa, M.M.; Leggett, D.S.; Li, C.J. ARQ 197, a novel and selective inhibitor of the human c-Met receptor tyrosine kinase with antitumor activity. Mol. Cancer Ther. 2010, 9, 1544–1553. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.R.; Gajagowni, S.; Bryan, J.N.; Bodenhausen, H.M. Molecular targets for tivantinib (ARQ 197) and vasculogenic mimicry in human melanoma cells. Eur. J. Pharmacol. 2019, 853, 316–324. [Google Scholar] [CrossRef]
- Kuenzi, B.M.; Remsing Rix, L.L.; Kinose, F.; Kroeger, J.L.; Lancet, J.E.; Padron, E.; Rix, U. Off-target based drug repurposing opportunities for tivantinib in acute myeloid leukemia. Sci. Rep. 2019, 9, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remsing Rix, L.L.; Kuenzi, B.M.; Luo, Y.; Remily-Wood, E.; Kinose, F.; Wright, G.; Li, J.; Koomen, J.M.; Haura, E.B.; Lawrence, H.R.; et al. GSK3 alpha and beta are new functionally relevant targets of tivantinib in lung cancer cells. ACS Chem. Biol. 2014, 9, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Laxmidevi, L.B.; Angadi, P.V.; Pillai, R.K.; Chandreshekar, C. Aberrant beta-catenin expression in the histologic differentiation of oral squamous cell carcinoma and verrucous carcinoma: An immunohistochemical study. J. Oral Sci. 2010, 52, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trejo-Solis, C.; Escamilla-Ramirez, A.; Jimenez-Farfan, D.; Castillo-Rodriguez, R.A.; Flores-Najera, A.; Cruz-Salgado, A. Crosstalk of the Wnt/beta-Catenin Signaling Pathway in the Induction of Apoptosis on Cancer Cells. Pharmaceuticals 2021, 14, 871. [Google Scholar] [CrossRef]
- de Galarreta, M.R.; Bresnahan, E.; Molina-Sanchez, P.; Lindblad, K.E.; Maier, B.; Sia, D.; Puigvehi, M.; Miguela, V.; Casanova-Acebes, M.; Dhainaut, M.; et al. beta-Catenin Activation Promotes Immune Escape and Resistance to Anti-PD-1 Therapy in Hepatocellular Carcinoma. Cancer Discov. 2019, 9, 1124–1141. [Google Scholar] [CrossRef]
- Liu, Y.; Ye, X.; Zhang, J.B.; Ouyang, H.; Shen, Z.; Wu, Y.; Wang, W.; Wu, J.; Tao, S.; Yang, X.; et al. PROX1 promotes hepatocellular carcinoma proliferation and sorafenib resistance by enhancing beta-catenin expression and nuclear translocation. Oncogene 2015, 34, 5524–5535. [Google Scholar] [CrossRef]
- Wu, K.; Bonavida, B. The activated NF-kappaB-Snail-RKIP circuitry in cancer regulates both the metastatic cascade and resistance to apoptosis by cytotoxic drugs. Crit. Rev. Immunol. 2009, 29, 241–254. [Google Scholar] [CrossRef]
- Rodel, F.; Sprenger, T.; Kaina, B.; Liersch, T.; Rodel, C.; Fulda, S.; Hehlgans, S. Survivin as a prognostic/predictive marker and molecular target in cancer therapy. Curr. Med. Chem. 2012, 19, 3679–3688. [Google Scholar] [CrossRef]
- Forootan, F.S.; Forootan, S.S.; Malki, M.I.; Chen, D.; Li, G.; Lin, K.; Rudland, P.S.; Foster, C.S.; Ke, Y. The expression of C-FABP and PPARgamma and their prognostic significance in prostate cancer. Int. J. Oncol. 2014, 44, 265–275. [Google Scholar] [CrossRef]
- Elix, C.; Pal, S.K.; Jones, J.O. The role of peroxisome proliferator-activated receptor gamma in prostate cancer. Asian J. Androl. 2018, 20, 238–243. [Google Scholar] [CrossRef]
- Steinestel, J.; Al Ghazal, A.; Arndt, A.; Schnoeller, T.J.; Schrader, A.J.; Moeller, P.; Steinestel, K. The role of histologic subtype, p16(INK4a) expression, and presence of human papillomavirus DNA in penile squamous cell carcinoma. BMC Cancer 2015, 15, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winters, B.R.; Mossanen, M.; Holt, S.K.; Lin, D.W.; Wright, J.L. Predictors of Nodal Upstaging in Clinical Node Negative Patients With Penile Carcinoma: A National Cancer Database Analysis. Urology 2016, 96, 29–34. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Cohort (n = 94) | |
|---|---|
| Age at diagnosis | |
| Mean (SD) | 64.9 (11.8) |
| Median [Min, Max] | 67.0 [31.0, 90.0] |
| ≤65 | 45 (47.9%) |
| >65 | 49 (52.1%) |
| Primary surgery | |
| Circumcision | 8 (8.5%) |
| Tumor excision | 5 (5.3%) |
| Partial penectomy | 55 (58.5%) |
| Total penectomy | 25 (26.6%) |
| Missing | 1 (1.1%) |
| Tumor grade | |
| Low (G1/G2) | 70 (74.5%) |
| High (G3/G4) | 22 (23.4%) |
| Missing | 2 (2.1%) |
| pT stage | |
| pT1 | 36 (38.3%) |
| pT2 | 30 (31.9%) |
| pT3 | 24 (25.5%) |
| Missing | 4 (4.3%) |
| HPV status | |
| Negative | 65 (69.1%) |
| Positive | 24 (25.5%) |
| Missing | 5 (5.3%) |
| pN stage | |
| NX-0 | 77 (81.9%) |
| N1 | 4 (4.3%) |
| N2 | 8 (8.5%) |
| N3 | 5 (5.3%) |
| Recurrence status | |
| No | 66 (70.2%) |
| Yes | 28 (29.8%) |
| Recurrence site | |
| None | 66 (70.2%) |
| Local | 18 (19.2%) |
| Systemic | 10 (10.6%) |
| Adjuvant therapy | |
| None | 68 (72.3%) |
| CTX | 16 (17.0%) |
| Radiation | 1 (1.1%) |
| CTX and Radiation | 2 (2.1%) |
| Missing | 7 (7.4%) |
| Tumor-dependent death | |
| No | 61 (64.9%) |
| Yes | 12 (12.8%) |
| Missing | 21 (22.3%) |
| Characteristic | OS | RFS | DSS | |||
|---|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | HR (95% CI) | p | |
| PPARg | ||||||
| Low expression | 1.00 (reference) | 1.00 (reference) | 1.00 (reference) | |||
| High expression | 2.77 (0.74–10.36) | 0.10 | 1.59 (0.41–6.13) | 0.50 | 5.18 (0.95–28.32) | 0.06 |
| β-catenin | ||||||
| Low expression | 1.00 (reference) | 1.00 (reference) | 1.00 (reference) | |||
| High expression | 0.76 (0.30–1.89) | 0.60 | 1.34 (0.52–3.40) | 0.50 | 0.74 (0.16–3.42) | 0.70 |
| c-MET | ||||||
| Low expression | 1.00 (reference) | 1.00 (reference) | 1.00 (reference) | |||
| High expression | 1.38 (0.57–3.37) | 0.50 | 1.05 (0.38–2.92) | 0.90 | 5.03 (1.08 -23.32) | 0.04 |
| Grading | ||||||
| low (G1/G2) | 1.00 (reference) | 1.00 (reference) | 1.00 (reference) | |||
| high (G1/G2) | 1.72 (0.72–4.10) | 0.20 | 1.45 (0.48–4.32) | 0.50 | 0.40 (0.08–2.13) | 0.28 |
| T-Stage | ||||||
| pT1/pT2 | 1.00 (reference) | 1.00 (reference) | 1.00 (reference) | |||
| pT3 | 0.89 (0.34–2.33) | 0.80 | 0.73 (0.27–1.97) | 0.50 | 0.56 (0.11–2.85) | 0.48 |
| N-Stage | ||||||
| N0 | 1.00 (reference) | 1.00 (reference) | 1.00 (reference) | |||
| N1/N2/N3 | 1.84 (0.67–5.04) | 0.20 | 1.11 (0.33–3.80) | 0.90 | 14.09 (3.29–60.43) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, A.; Slade, K.S.; Blaheta, R.A.; Markowitsch, S.D.; Stenzel, P.; Tagscherer, K.E.; Roth, W.; Schindeldecker, M.; Michaelis, M.; Rothweiler, F.; et al. Value of c-MET and Associated Signaling Elements for Predicting Outcomes and Targeted Therapy in Penile Cancer. Cancers 2022, 14, 1683. https://doi.org/10.3390/cancers14071683
Thomas A, Slade KS, Blaheta RA, Markowitsch SD, Stenzel P, Tagscherer KE, Roth W, Schindeldecker M, Michaelis M, Rothweiler F, et al. Value of c-MET and Associated Signaling Elements for Predicting Outcomes and Targeted Therapy in Penile Cancer. Cancers. 2022; 14(7):1683. https://doi.org/10.3390/cancers14071683
Chicago/Turabian StyleThomas, Anita, Kimberly Sue Slade, Roman A. Blaheta, Sascha D. Markowitsch, Philipp Stenzel, Katrin E. Tagscherer, Wilfried Roth, Mario Schindeldecker, Martin Michaelis, Florian Rothweiler, and et al. 2022. "Value of c-MET and Associated Signaling Elements for Predicting Outcomes and Targeted Therapy in Penile Cancer" Cancers 14, no. 7: 1683. https://doi.org/10.3390/cancers14071683