
Chemotherapy-Free Targeted Anti-BCR-ABL+ Acute Lymphoblastic Leukemia Therapy May Benefit the Heart

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pharmacologic Agents
2.2. Patient Samples
2.3. Cell Lines
2.4. Preparation of Lentiviral Supernatants and Transduction
2.5. Isolation and Culture of Primary Adult Mouse Cardiomyocytes
2.6. Culture and Cardiomyogenic Differentiation of Human Induced Pluripotent Stem Cells (iPSCs)
2.7. Isolation and Culture of Human Mesenchymal Stem Cells (MSCs) from Bone Marrow
2.8. Co-Culture of ALL Cells on MSCs
2.9. Animal Experiments
2.10. In Vivo Bioluminescence Imaging
2.11. Echocardiography
2.12. Histology and Immunostaining
2.13. Immunoblotting
2.14. RNA-Isolation and qRT-PCR
2.15. Statistics
3. Results
3.1. Acute Leukemia-Bearing Mice Developed Profound Cardiac Atrophy Associated with Impaired Cardiac Function
3.2. Selective Ablation of Leukemia with Herpes Simplex Virus-Thymidine Kinase (HSV-TK)/Ganciclovir (GCV) Did Not Recover Cardiac Function and Output
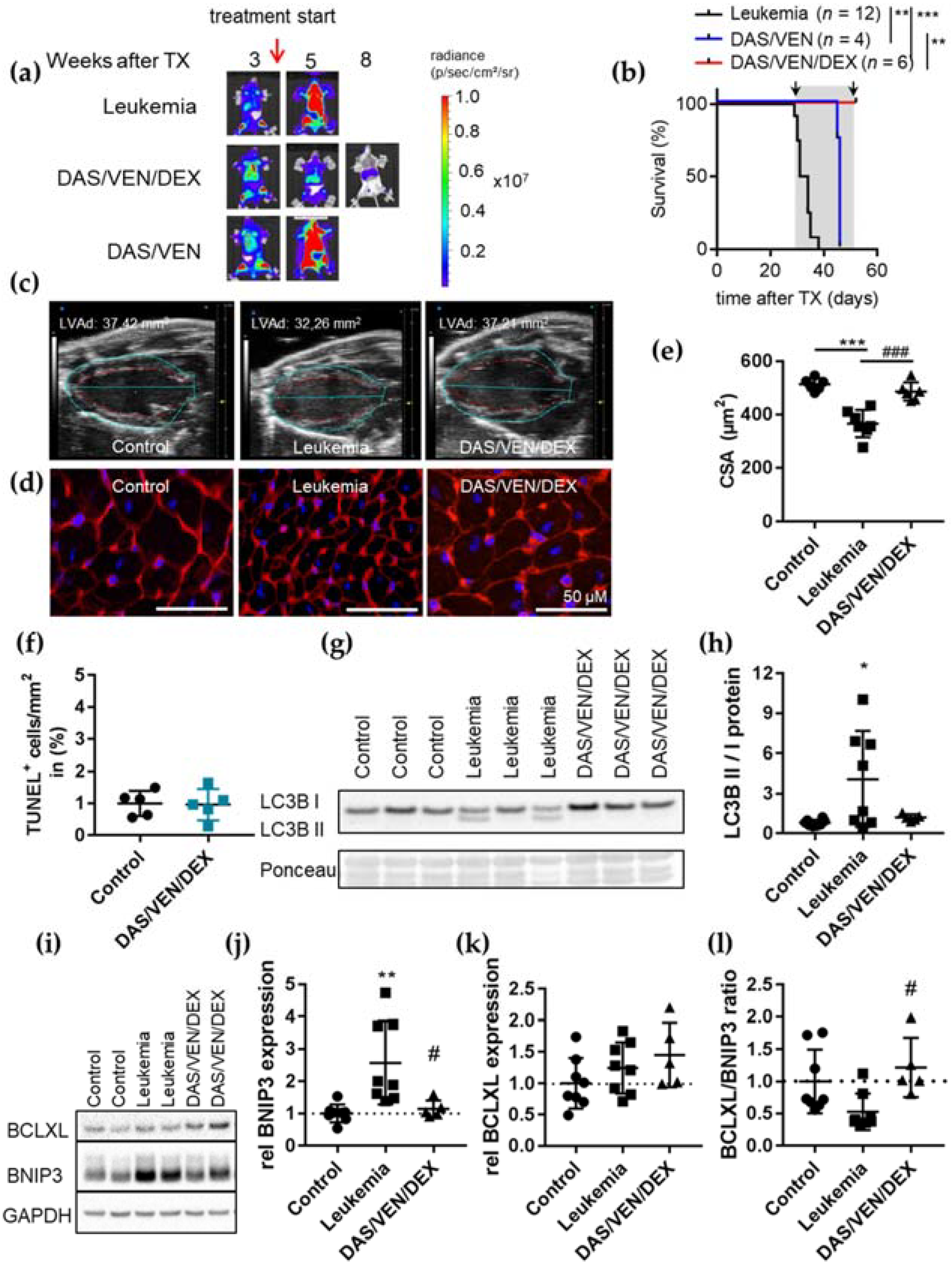
3.3. Combination Therapy of DAS/VEN/DEX Eradicated Leukemia and Improved Cardiac Function and Output
3.4. DAS/VEN/DEX Alters the BCL2 Family Network Differentially in ALL Cells and Cardiomyocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoelzer, D.; Gokbuget, N. Recent approaches in acute lymphoblastic leukemia in adults. Critk. Rev. Oncol. Hematol. 2000, 36, 49–58. [Google Scholar] [CrossRef]
- Moorman, A.V.; Harrison, C.J.; Buck, G.A.; Richards, S.M.; Secker-Walker, L.M.; Martineau, M.; Vance, G.H.; Cherry, A.M.; Higgins, R.R.; Fielding, A.K.; et al. Karyotype is an independent prognostic factor in adult acute lymphoblastic leukemia (ALL): Analysis of cytogenetic data from patients treated on the Medical Research Council (MRC) UKALLXII/Eastern Cooperative Oncology Group (ECOG) 2993 trial. Blood 2007, 109, 3189–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottmann, O.G.; Pfeifer, H. Management of Philadelphia chromosome-positive acute lymphoblastic leukemia (Ph+ ALL). Hematol. Am. Soc. Hematol. Educ. Program 2009, 2009, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravandi, F. Current management of Philadelphia chromosome positive ALL and the role of stem cell transplantation. Hematol. Am. Soc. Hematol. Educ. Program 2017, 2017, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Gökbuget, N.; Kneba, M.; Raff, T.; Trautmann, H.; Bartram, C.-R.; Arnold, R.; Fietkau, R.; Freund, M.; Ganser, A.; Ludwig, W.-D.; et al. Adult patients with acute lymphoblastic leukemia and molecular failure display a poor prognosis and are candidates for stem cell transplantation and targeted therapies. Blood 2012, 120, 1868–1876. [Google Scholar] [CrossRef] [Green Version]
- Fielding, A.K.; Richards, S.M.; Chopra, R.; Lazarus, H.M.; Litzow, M.R.; Buck, G.; Durrant, I.J.; Luger, S.M.; Marks, D.I.; Franklin, I.M.; et al. Outcome of 609 adults after relapse of acute lymphoblastic leukemia (ALL); an MRC UKALL12/ECOG 2993 study. Blood 2007, 109, 944–950. [Google Scholar] [CrossRef] [Green Version]
- Davids, M.S.; Letai, A. Targeting the B-cell lymphoma/leukemia 2 family in cancer. J. Clin. Oncol. 2012, 30, 3127–3135. [Google Scholar] [CrossRef] [Green Version]
- Souers, A.J.; Leverson, J.D.; Boghaert, E.R.; Ackler, S.L.; Catron, N.D.; Chen, J.; Dayton, B.D.; Ding, H.; Enschede, S.H.; Fairbrother, W.J.; et al. ABT-199, a potent and selective BCL-2 inhibitor, achieves antitumor activity while sparing platelets. Nat. Med. 2013, 19, 202–208. [Google Scholar] [CrossRef]
- Scherr, M.; Kirchhoff, H.; Battmer, K.; Wohlan, K.; Lee, C.-W.; Ricke-Hoch, M.; Erschow, S.; Law, E.; Kloos, A.; Heuser, M.; et al. Optimized induction of mitochondrial apoptosis for chemotherapy-free treatment of BCR-ABL+acute lymphoblastic leukemia. Leukemia 2019, 33, 1313–1323. [Google Scholar] [CrossRef]
- Kirchhoff, H.; Karsli, U.; Schoenherr, C.; Battmer, K.; Erschow, S.; Talbot, S.R.; Steinemann, D.; Heuser, M.; Heidenreich, O.; Hilfiker-Kleiner, D.; et al. Venetoclax and dexamethasone synergize with inotuzumab ozogamicin–induced DNA damage signaling in B-lineage ALL. Blood 2021, 137, 2657–2661. [Google Scholar] [CrossRef]
- Leonard, J.T.; Rowley, J.S.J.; Eide, C.A.; Traer, E.; Hayes-Lattin, B.; Loriaux, M.; Spurgeon, S.E.; Druker, B.J.; Tyner, J.W.; Chang, B.H. Targeting BCL-2 and ABL/LYN in Philadelphia chromosome–positive acute lymphoblastic leukemia. Sci. Transl. Med. 2016, 8, 354ra114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, B.; Cote, G.M.; Sawyer, D.B.; Chabner, B.A. ERBB2 Inhibition and Heart Failure. N. Engl. J. Med. 2013, 368, 875–876. [Google Scholar] [PubMed]
- Chen, Z.I.; Ai, D.I. Cardiotoxicity associated with targeted cancer therapies. Mol. Clin. Oncol. 2016, 4, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, M.T.; Luu, S.; Gurney, H.; Brown, M.R.; Pearson, S.-A.; Webber, K.; Hunt, L.; Hong, S.; Delaney, G.P.; Vajdic, C.M. Cardiovascular Toxicity of Targeted Therapies for Cancer: An Overview of Systematic Reviews. JNCI Cancer Spectr. 2020, 4, pkaa076. [Google Scholar] [CrossRef]
- Caldemeyer, L.; Dugan, M.; Edwards, J.; Akard, L. Long-Term Side Effects of Tyrosine Kinase Inhibitors in Chronic Myeloid Leukemia. Curr. Hematol. Malig. Rep. 2016, 11, 71–79. [Google Scholar] [CrossRef]
- Singh, A.P.; Umbarkar, P.; Tousif, S.; Lal, H. Cardiotoxicity of the BCR-ABL1 tyrosine kinase inhibitors: Emphasis on ponatinib. Int. J. Cardiol. 2020, 316, 214–221. [Google Scholar] [CrossRef]
- Venturini, L.; Battmer, K.; Castoldi, M.; Schultheis, B.; Hochhaus, A.; Muckenthaler, M.; Ganser, A.; Eder, M.; Scherr, M. Expression of the miR-17-92 polycistron in chronic myeloid leukemia (CML) CD34+ cells. Blood 2007, 109, 4399–4405. [Google Scholar] [CrossRef]
- Thackeray, J.T.; Pietzsch, S.; Stapel, B.; Ricke-Hoch, M.; Lee, C.-W.; Bankstahl, J.P.; Scherr, M.; Heineke, J.; Scharf, G.; Haghikia, A.; et al. Insulin supplementation attenuates cancer-induced cardiomyopathy and slows tumor disease progression. JCI Insight 2017, 2, e93098. [Google Scholar] [CrossRef] [Green Version]
- Kempf, H.; Olmer, R.; Haase, A.; Franke, A.; Bolesani, E.; Schwanke, K.; Robles-Diaz, D.; Coffee, M.; Goehring, G.; Draeger, G.; et al. Bulk cell density and Wnt/TGFbeta signalling regulate mesendodermal patterning of human pluripotent stem cells. Nat. Commun. 2016, 7, 13602. [Google Scholar] [CrossRef] [Green Version]
- Kropp, C.; Kempf, H.; Halloin, C.; Robles-Diaz, D.; Franke, A.; Scheper, T.; Kinast, K.; Knorpp, T.; Joos, T.O.; Haverich, A.; et al. Impact of Feeding Strategies on the Scalable Expansion of Human Pluripotent Stem Cells in Single-Use Stirred Tank Bioreactors. Stem Cells Transl. Med. 2016, 5, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Halloin, C.; Schwanke, K.; Löbel, W.; Franke, A.; Szepes, M.; Biswanath, S.; Wunderlich, S.; Merkert, S.; Weber, N.; Osten, F.; et al. Continuous WNT Control Enables Advanced hPSC Cardiac Processing and Prognostic Surface Marker Identification in Chemically Defined Suspension Culture. Stem Cell Rep. 2019, 13, 775. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, N.; Tejedor, L.S.; Jin, W.; Gudi, V.; Skripuletz, T.; Stein, V.M.; Tipold, A.; Hoffmann, A.; Stangel, M.; Baumgärtner, W.; et al. Mesenchymal Stem Cells Form 3D Clusters Following Intraventricular Transplantation. J. Mol. Neurosci. 2018, 65, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Morton, D.; Griffiths, P. Guidelines on the recognition of pain, distress and discomfort in experimental animals and an hypothesis for assessment. Vet. Rec. 1985, 116, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Bomken, S.; Buechler, L.; Rehe, K.; Ponthan, F.; Elder, A.; Blair, H.; Bacon, C.M.; Vormoor, J.; Heidenreich, O. Lentiviral marking of patient-derived acute lymphoblastic leukaemic cells allows in vivo tracking of disease progression. Leukemia 2012, 27, 718–721. [Google Scholar] [CrossRef] [Green Version]
- Pincha, M.; Salguero, G.; Wedekind, D.; Sundarasetty, B.S.; Lin, A.; Kasahara, N.; Brugman, M.H.; Jirmo, A.C.; Modlich, U.; Gutzmer, R.; et al. Lentiviral vectors for induction of self-differentiation and conditional ablation of dendritic cells. Gene Ther. 2011, 18, 750–764. [Google Scholar] [CrossRef]
- Hilfiker-Kleiner, D.; Shukla, P.; Klein, G.; Schaefer, A.; Stapel, B.; Hoch, M.; Muller, W.; Scherr, M.; Theilmeier, G.; Ernst, M.; et al. Continuous Glycoprotein-130–Mediated Signal Transducer and Activator of Transcription-3 Activation Promotes Inflammation, Left Ventricular Rupture, and Adverse Outcome in Subacute Myocardial Infarction. Circulation 2010, 122, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Pietzsch, S.; Ricke-Hoch, M.; Stapel, B.; Hilfiker-Kleiner, D. Modulation of cardiac AKT and STAT3 signalling in preclinical cancer models and their impact on the heart. Biochim. Biophys. Acta 2020, 1867, 118519. [Google Scholar] [CrossRef]
- Schips, T.G.; Wietelmann, A.; Höhn, K.; Schimanski, S.; Walther, P.; Braun, T.; Wirth, T.; Maier, H.J. FoxO3 induces reversible cardiac atrophy and autophagy in a transgenic mouse model. Cardiovasc. Res. 2011, 91, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Madden, S.D.; Donovan, M.; Cotter, T.G. Key apoptosis regulating proteins are down-regulated during postnatal tissue development. Int. J. Dev. Biol. 2007, 51, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Veis, D.J.; Sorenson, C.M.; Shutter, J.R.; Korsmeyer, S.J. Bcl-2-deficient mice demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell 1993, 75, 229–240. [Google Scholar] [CrossRef]
- Rasmussen, M.; Taneja, N.; Neininger, A.; Wang, L.; Robertson, G.L.; Riffle, S.N.; Shi, L.; Knollmann, B.C.; Burnette, D.T.; Gama, V. MCL-1 Inhibition by Selective BH3 Mimetics Disrupts Mitochondrial Dynamics Causing Loss of Viability and Functionality of Human Cardiomyocytes. iScience 2020, 23, 101015. [Google Scholar] [CrossRef]
- Zhang, J.; Ney, P.A. Role of BNIP3 and NIX in cell death, autophagy, and mitophagy. Cell Death Differ. 2009, 16, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Ray, R.; Dubik, D.; Shi, L.; Cizeau, J.; Bleackley, R.C.; Saxena, S.; Gietz, R.D.; Greenberg, A.H. The E1B 19K/Bcl-2–binding Protein Nip3 is a Dimeric Mitochondrial Protein that Activates Apoptosis. J. Exp. Med. 1997, 186, 1975–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanna, R.A.; Quinsay, M.N.; Orogo, A.M.; Giang, K.; Rikka, S.; Gustafsson, Å.B. Microtubule-associated Protein 1 Light Chain 3 (LC3) Interacts with Bnip3 Protein to Selectively Remove Endoplasmic Reticulum and Mitochondria via Autophagy. J. Biol. Chem. 2012, 287, 19094–19104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murai, M.; Toyota, M.; Satoh, A.; Suzuki, H.; Akino, K.; Mita, H.; Sasaki, Y.; Ishida, T.; Shen, L.; Garciamanero, G.; et al. Aberrant DNA methylation associated with silencing BNIP3 gene expression in haematopoietic tumours. Br. J. Cancer 2005, 92, 1165–1172. [Google Scholar] [CrossRef] [Green Version]
- Heller, G.; Schmidt, W.; Ziegler, B.; Holzer, S.; Müllauer, L.; Bilban, M.; Zielinski, C.C.; Drach, J.; Zöchbauer-Müller, S. Genome-Wide Transcriptional Response to 5-Aza-2′-Deoxycytidine and Trichostatin A in Multiple Myeloma Cells. Cancer Res. 2008, 68, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okami, J.; Simeone, D.M.; Logsdon, C.D. Silencing of the Hypoxia-Inducible Cell Death Protein BNIP3 in Pancreatic Cancer. Cancer Res. 2004, 64, 5338–5346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, R.; Oakley, R.H.; Cruz-Topete, D.; Cidlowski, J.A. Dual Role for Glucocorticoids in Cardiomyocyte Hypertrophy and Apoptosis. Endocrinology 2012, 153, 5346–5360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saruta, T. Mechanism of Glucocorticoid-Induced Hypertension. Hypertens. Res. 1996, 19, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, B.J. Hypertrophic cardiomyopathy: A systematic review. JAMA 2002, 287, 1308–1320. [Google Scholar] [CrossRef] [Green Version]
- De, P.; Roy, S.G.; Kar, D.; Bandyopadhyay, A. Excess of glucocorticoid induces myocardial remodeling and alteration of calcium signaling in cardiomyocytes. J. Endocrinol. 2011, 209, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Strom, J.; Chen, Q.M. Dexamethasone induces transcriptional activation of Bcl-xL gene and inhibits cardiac injury by myocardial ischemia. Eur. J. Pharmacol. 2011, 668, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.M.; Alexander, D.; Sun, H.; Xie, L.; Lin, Y.; Terrand, J.; Morrissy, S.; Purdom, S. Corticosteroids Inhibit Cell Death Induced by Doxorubicin in Cardiomyocytes: Induction of Antiapoptosis, Antioxidant, and Detoxification Genes. Mol. Pharmacol. 2005, 67, 1861–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Q.-G.; Na, T.; Guo, Y.-M.; Bi, Y.-T.; Zhang, H.-Y.; Dai, D.-Z. Improvement of chronic heart failure by dexamethasone is not associated with downregulation of leptin in rats. Acta Pharmacol. Sin. 2007, 28, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, N.; Yang, C.; Xu, A. Dexamethasone treatment improves sarcoplasmic reticulum function and contractile performance in aged myocardium. Mol. Cell. Biochem. 2004, 266, 31–36. [Google Scholar] [CrossRef]
- Pietzsch, S.; Wohlan, K.; Thackeray, J.T.; Heimerl, M.; Schuchardt, S.; Scherr, M.; Ricke-Hoch, M.; Hilfiker-Kleiner, D. Anthracycline-free tumor elimination in mice leads to functional and molecular cardiac recovery from cancer-induced alterations in contrast to long-lasting doxorubicin treatment effects. Basic Res. Cardiol. 2021, 116, 1–20. [Google Scholar] [CrossRef]
- McGowan, J.V.; Chung, R.; Maulik, A.; Piotrowska, I.; Walker, J.M.; Yellon, D.M. Anthracycline Chemotherapy and Cardiotoxicity. Cardiovasc. Drugs Ther. 2017, 31, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.K.; Chanson, D.; Teh, J.B.; Iukuridze, A.; Peng, K.; Forman, S.J.; Nakamura, R.; Wong, F.L.; Cai, L.; Armenian, S.H. Atrial Fibrillation in Patients Undergoing Allogeneic Hematopoietic Cell Transplantation. J. Clin. Oncol. 2021, 39, 902–910. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| mRNA | Sense Primers (5′ to 3′) | Antisense Primers (5′ to 3′) |
|---|---|---|
| mmu 18S | GTAACCCGTTGAACCCCATT | CCATCCAATCGGTAGTAGCG |
| mmu BCLXL | GACAAGGAGATGCAGGTATTGG | TCCCGTAGAGATCCACAAAAGT |
| mmu BNIP3 | CAGCATGAGAAACACAAGCGT | GTTGTCAGACGCCTTCCAATG |
| mmu LC3B | CATGCCGTCCGAGAAGACCT | TCGCTCTATAATCACTGGGATCT |
| Parameters | Age-Matched Healthy Control for BV173 (n = 10) | BV173 (n = 7) | Age-Matched Healthy Control for PDX (n = 9) | PDX (n = 5) |
|---|---|---|---|---|
| BW (g) | 24.12 ± 1.68 | 18.53 ± 1.85 * | 28.06 ± 1.01 | 22.10 ± 1.81 *** |
| HW (mg) | 98.46 ± 0.88 | 68.46 ± 8.29 *** | 102.46 ± 6.83 | 86.20 ± 5.07 ** |
| HW/BW (mg/g) | 4.29 ± 0.55 | 3.7 ± 0.47 | 3.65 ± 0.15 | 3.92 ± 0.39 |
| LVEDD (mm) | 7.71 ± 0.64 | 6.65 ± 0.36 ** | 7.56 ± 0.37 | 7.19 ± 0.78 |
| LVESD (mm) | 6.35 ± 0.51 | 5.33 ± 0.68 | 6.48 ± 0.45 | 6.41 ± 0.31 |
| EDA (mm2) | 0.20 ± 0.02 | 0.16 ± 0.015 ** | 0.20 ± 0.01 | 0.16 ± 0.03 * |
| ESA (mm2) | 0.09 ± 0.02 | 0.07 ± 0.02 | 0.11 ± 0.02 | 0.10 ± 0.02 |
| HR (bpm) | 511 ± 30 | 499 ± 34 | 506 ± 37 | 492 ± 36 |
| EVd (µL) | 43.87 ± 7.75 | 31.90 ± 5.58 * | 43.44 ± 5.39 | 32.39 ± 9.84 * |
| EVs (µL) | 11.36 ± 3.24 | 9.09 ± 4.33 | 15.75 ± 3.80 | 12.58 ± 4.60 |
| FAC (%) | 54.36 ± 5.56 | 53.28 ± 11.56 | 44.80 ± 7.80 | 41.60 ± 7.96 |
| EF (%) | 74.14 ± 5.58 | 71.84 ± 11.96 | 63.66 ± 7.80 | 61.51 ± 9.72 |
| Parameters | Before Treatment | Four Weeks Treatment | ||
|---|---|---|---|---|
| Control (n = 14) | Leukemia (n = 5) | Control (n = 6) | GCV (n = 5) | |
| LVEDD (mm) | 7.62 ± 0.39 | 7.26 ± 0.48 | 7.98 ± 0.15 | 7.12 ± 0.62 |
| LVESD (mm) | 6.12 ± 0.89 | 6.44 ± 0.70 | 6.63 ± 0.48 | 5.97 ± 0.16 * |
| EDA (mm2) | 0.19 ± 0.02 | 0.16 ± 0.05 * | 0.20 ± 0.03 | 0.15 ± 0.02 * |
| ESA (mm2) | 0.09±0.02 | 0.08 ± 0.02 | 0.09 ± 0.02 | 0.07 ± 0.01 * |
| HR (bpm) | 505 ± 38 | 507 ± 28 | 500 ± 32 | 541 ± 13 * |
| LV CO (mL/min) | 14.84 ± 2.44 | 11.8 ± 5.15 | 16.34 ± 4.29 | 11.19 ± 2.33 * |
| ESV (µL) | 29.49 ± 4.87 | 23.3 ± 10.11 | 32.47 ± 7.32 | 20.72 ± 4.42 * |
| EVd (µL) | 41.40 ± 7.91 | 31.73 ± 13.5 | 43.82 ± 8.53 | 27.53 ± 5.82 ** |
| EVs (µL) | 11.91 ± 5.15 | 8.43 ± 3.99 | 11.35 ± 3.74 | 6.81 ± 1.82 * |
| FAC (%) | 52.37 ± 10.10 | 51.22 ± 6.44 | 54.42 ± 6.88 | 54.82 ± 3.16 |
| EF (%) | 72.03 ± 9.31 | 72.35 ± 7.5 | 74.04 ± 7.05 | 75.31 ± 3.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchhoff, H.; Ricke-Hoch, M.; Wohlan, K.; Pietzsch, S.; Karsli, Ü.; Erschow, S.; Zweigerdt, R.; Ganser, A.; Eder, M.; Scherr, M.; et al. Chemotherapy-Free Targeted Anti-BCR-ABL+ Acute Lymphoblastic Leukemia Therapy May Benefit the Heart. Cancers 2022, 14, 983. https://doi.org/10.3390/cancers14040983
Kirchhoff H, Ricke-Hoch M, Wohlan K, Pietzsch S, Karsli Ü, Erschow S, Zweigerdt R, Ganser A, Eder M, Scherr M, et al. Chemotherapy-Free Targeted Anti-BCR-ABL+ Acute Lymphoblastic Leukemia Therapy May Benefit the Heart. Cancers. 2022; 14(4):983. https://doi.org/10.3390/cancers14040983
Chicago/Turabian StyleKirchhoff, Hanna, Melanie Ricke-Hoch, Katharina Wohlan, Stefan Pietzsch, Ümran Karsli, Sergej Erschow, Robert Zweigerdt, Arnold Ganser, Matthias Eder, Michaela Scherr, and et al. 2022. "Chemotherapy-Free Targeted Anti-BCR-ABL+ Acute Lymphoblastic Leukemia Therapy May Benefit the Heart" Cancers 14, no. 4: 983. https://doi.org/10.3390/cancers14040983