
In Situ N-Glycosylation Signatures of Epithelial Ovarian Cancer Tissue as Defined by MALDI Mass Spectrometry Imaging

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
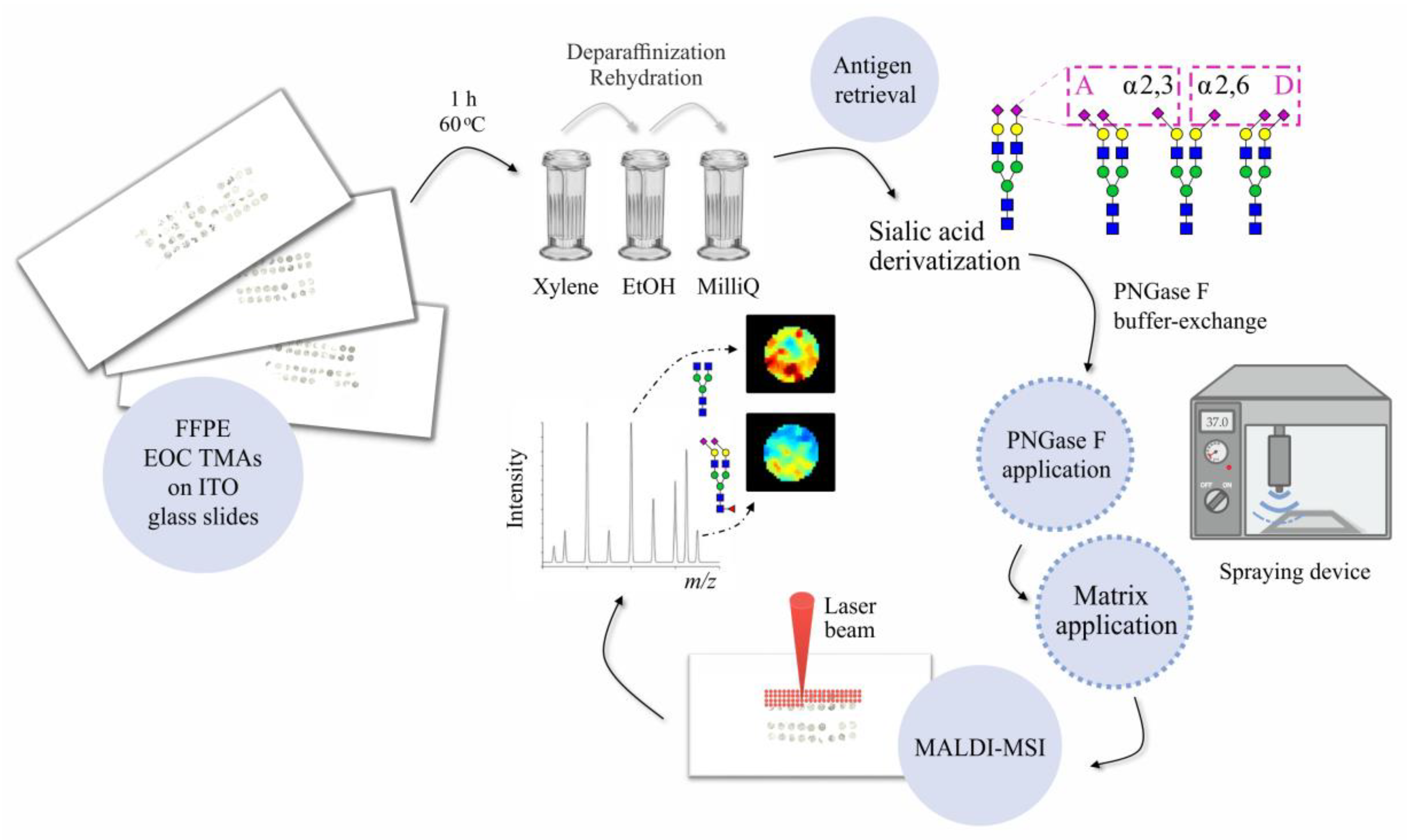
2. Materials and Methods
2.1. Chemicals and Reagents
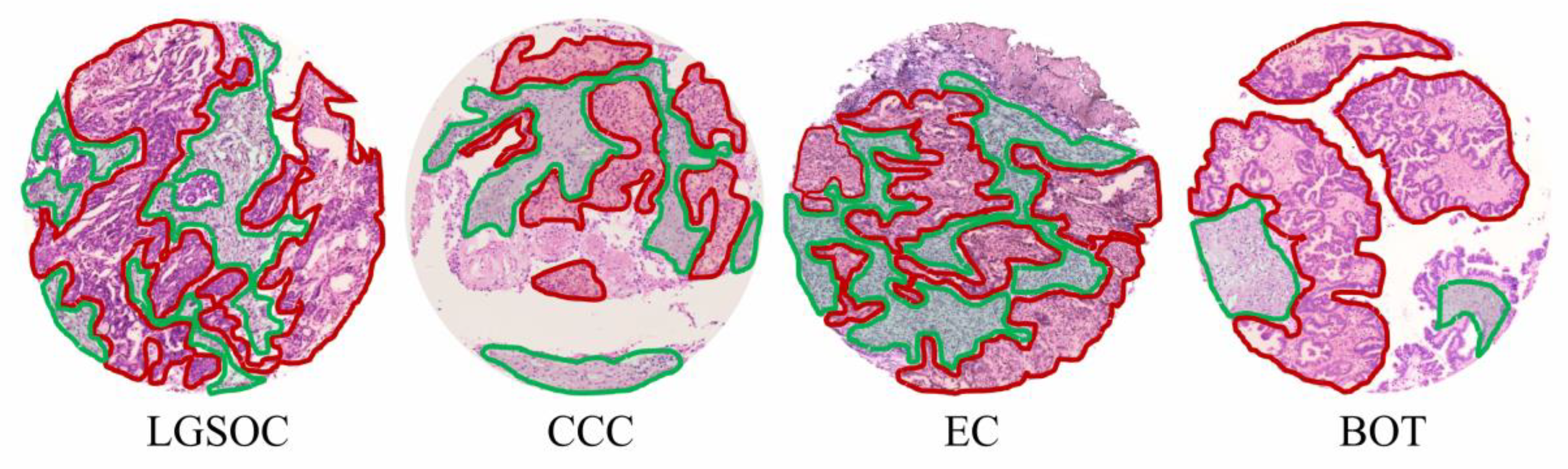
2.2. Tissue Sample Collection and Preparation
2.3. Deparaffinization, Rehydration, and Antigen Retrieval of EOC Tissues
2.4. Chemical Derivatization of Sialic Acids
2.5. In Situ PNGase F Deposition and Digestion
2.6. MALDI Matrix Deposition
2.7. MALDI-MSI Measurement
2.8. Hematoxylin and Eosin Staining
2.9. Data Analysis
3. Results
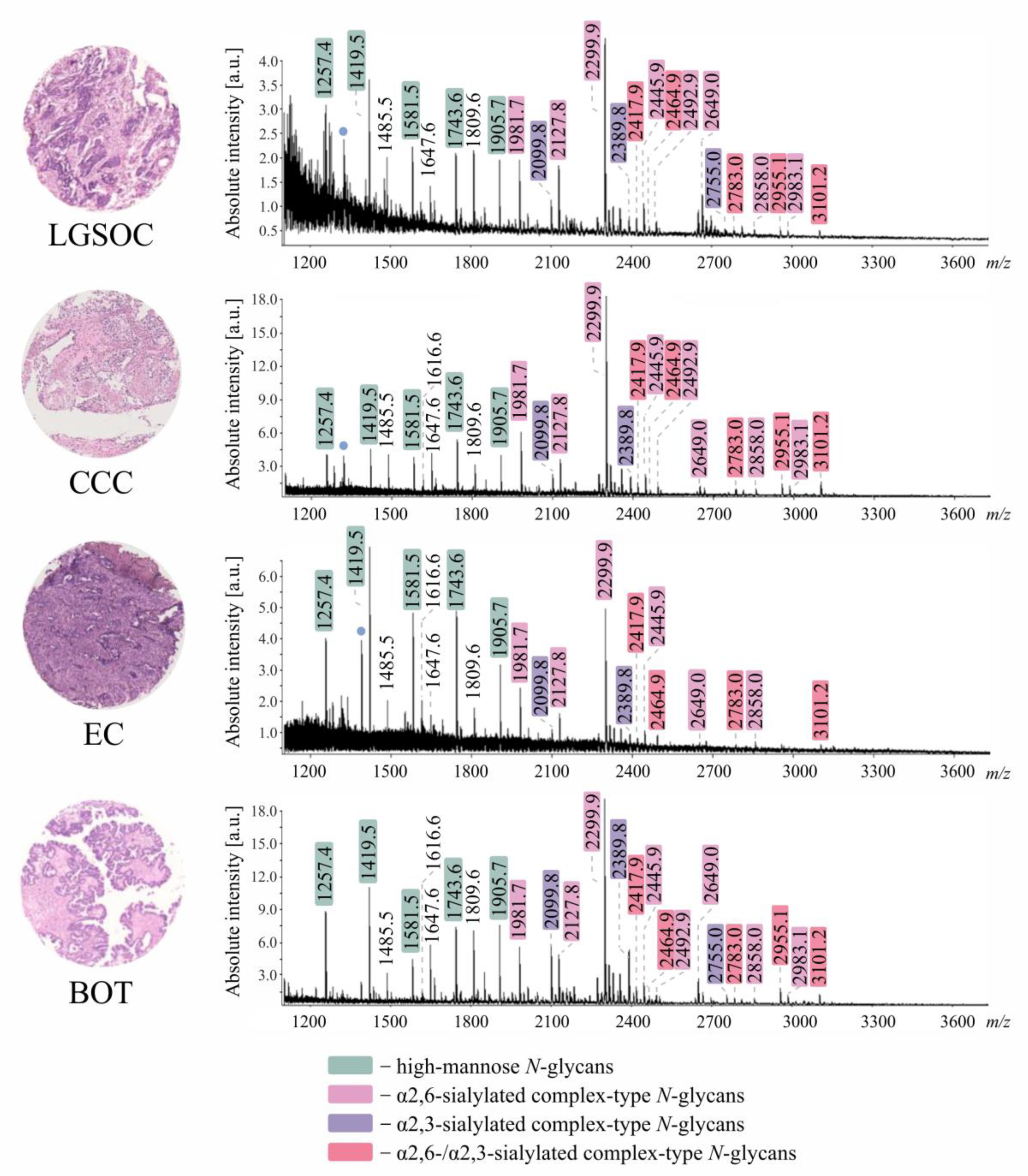
3.1. In Situ N-Glycosylation Profiling of EOC Tissues
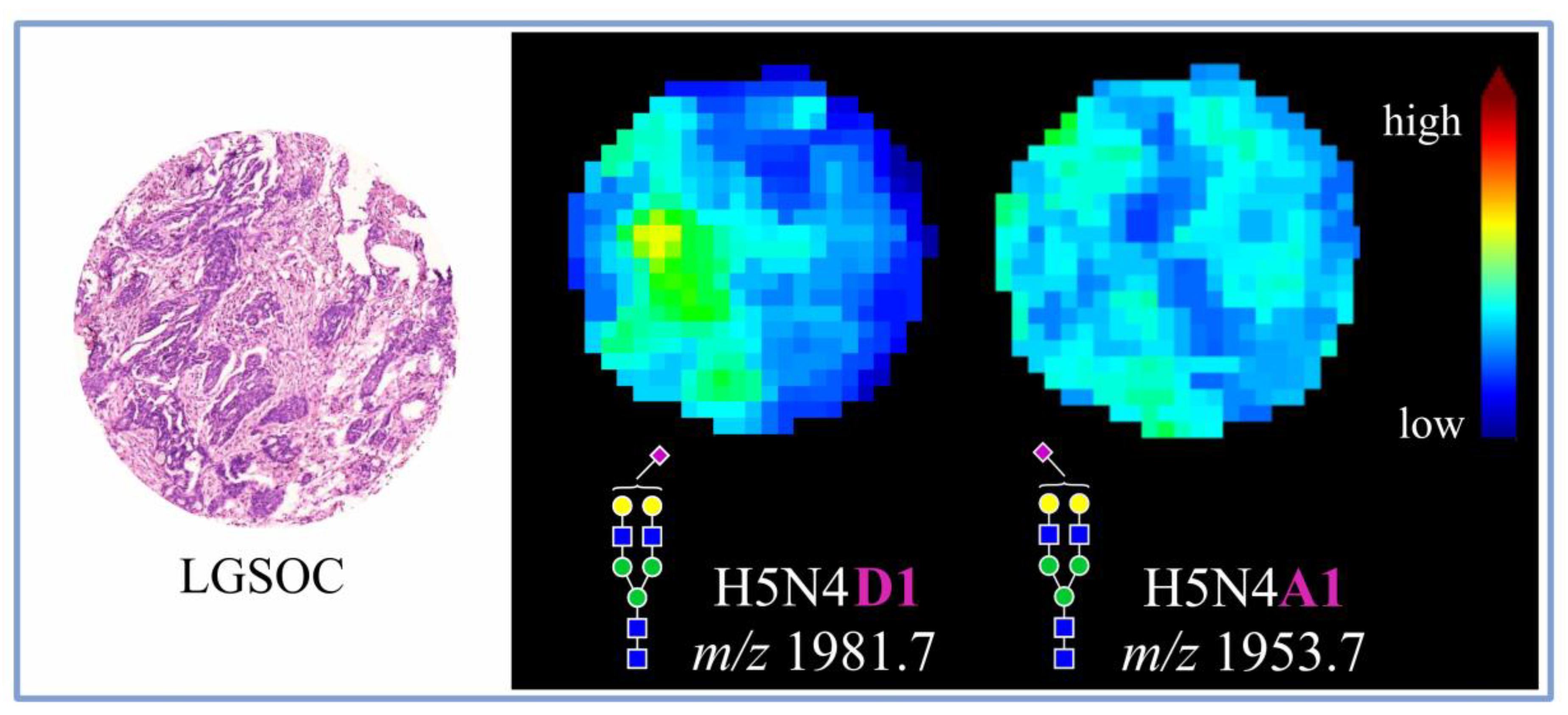
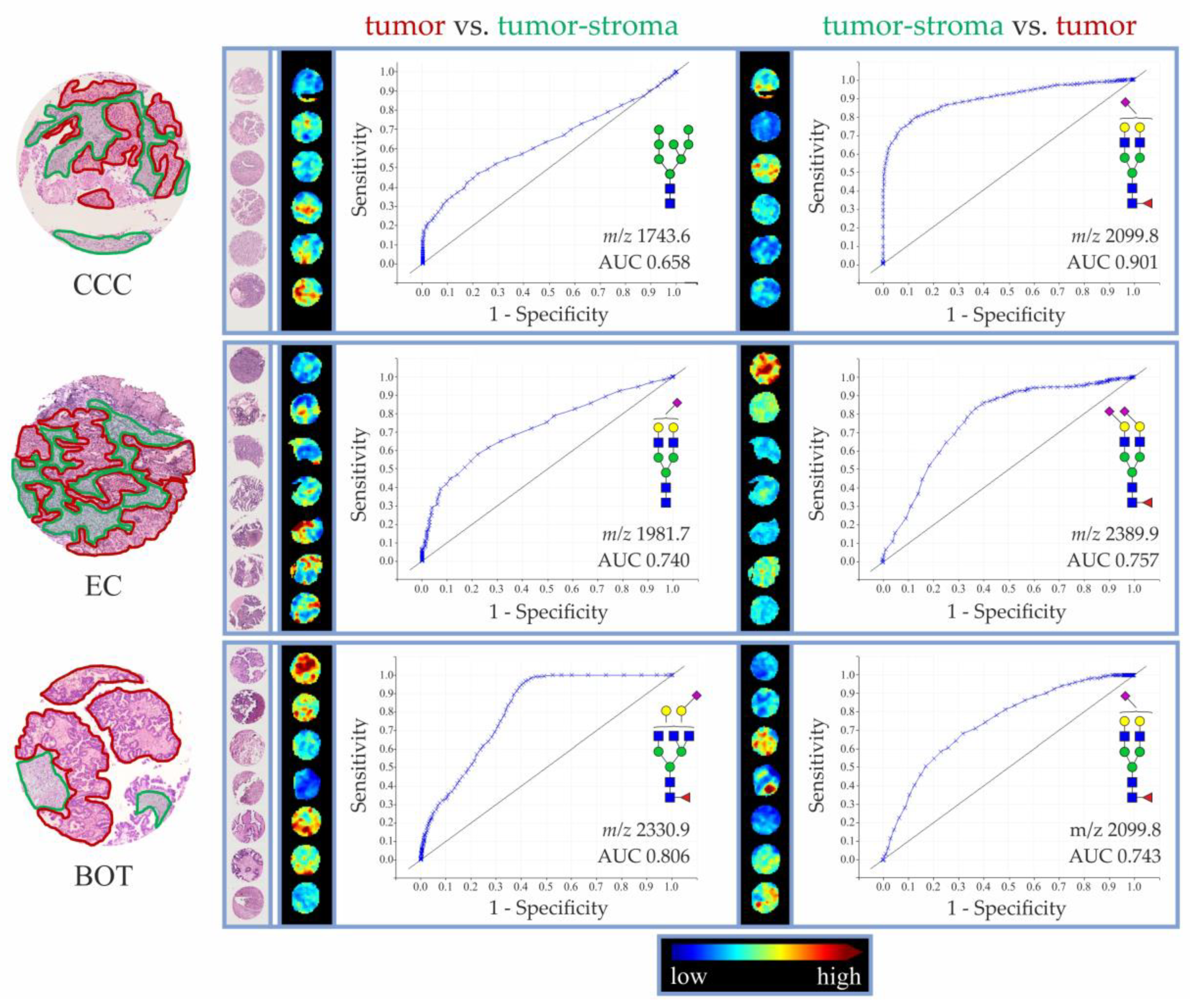
3.2. Discriminatory Power of In Situ Released N-Glycans in TMA Ovarian Cancer Specimens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Cancer Society. Global Cancer Facts & Figures, 4th ed.; American Cancer Society: Atlanta, GA, USA, 2018. [Google Scholar]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanyama, F.M.; Blanchard, V. Glycomic-Based Biomarkers for Ovarian Cancer: Advances and Challenges. Diagnostics 2021, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- Matulonis, U.A.; Sood, A.K.; Fallowfield, L.; Howitt, B.E.; Sehouli, J.; Karlan, B.Y. Ovarian cancer. Nat. Rev. Dis. Prim. 2016, 2, 16061. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Shih Ie, M. The origin and pathogenesis of epithelial ovarian cancer: A proposed unifying theory. Am. J. Surg. Pathol. 2010, 34, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Stanley, P.; Schachter, H.; Taniguchi, N. N-glycans. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Mereiter, S.; Balmana, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef]
- Saldova, R.; Royle, L.; Radcliffe, C.M.; Abd Hamid, U.M.; Evans, R.; Arnold, J.N.; Banks, R.E.; Hutson, R.; Harvey, D.J.; Antrobus, R.; et al. Ovarian cancer is associated with changes in glycosylation in both acute-phase proteins and IgG. Glycobiology 2007, 17, 1344–1356. [Google Scholar] [CrossRef] [Green Version]
- Saldova, R.; Wormald, M.R.; Dwek, R.A.; Rudd, P.M. Glycosylation changes on serum glycoproteins in ovarian cancer may contribute to disease pathogenesis. Dis. Markers 2008, 25, 219–232. [Google Scholar] [CrossRef]
- Li, B.; An, H.J.; Kirmiz, C.; Lebrilla, C.B.; Lam, K.S.; Miyamoto, S. Glycoproteomic analyses of ovarian cancer cell lines and sera from ovarian cancer patients show distinct glycosylation changes in individual proteins. J. Proteome Res. 2008, 7, 3776–3788. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Kim, K.; Stroble, C.; Taylor, S.L.; Hong, Q.; Miyamoto, S.; Lebrilla, C.B.; Leiserowitz, G. Protein-Specific Differential Glycosylation of Immunoglobulins in Serum of Ovarian Cancer Patients. J. Proteome Res. 2016, 15, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Weiz, S.; Wieczorek, M.; Schwedler, C.; Kaup, M.; Braicu, E.I.; Sehouli, J.; Tauber, R.; Blanchard, V. Acute-phase glycoprotein N-glycome of ovarian cancer patients analyzed by CE-LIF. Electrophoresis 2016, 37, 1461–1467. [Google Scholar] [CrossRef]
- Tang, H.; Hsueh, P.; Kletter, D.; Bern, M.; Haab, B. The detection and discovery of glycan motifs in biological samples using lectins and antibodies: New methods and opportunities. Adv. Cancer Res. 2015, 126, 167–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballehaninna, U.K.; Chamberlain, R.S. The clinical utility of serum CA 19-9 in the diagnosis, prognosis and management of pancreatic adenocarcinoma: An evidence based appraisal. J. Gastrointest. Oncol. 2012, 3, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Haji-Ghassemi, O.; Blackler, R.J.; Martin Young, N.; Evans, S.V. Antibody recognition of carbohydrate epitopesdagger. Glycobiology 2015, 25, 920–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, I.; Wisztorski, M.; Salzet, M. Tissue imaging using MALDI-MS: A new frontier of histopathology proteomics. Expert Rev. Proteom. 2008, 5, 413–424. [Google Scholar] [CrossRef]
- Goto-Inoue, N.; Hayasaka, T.; Zaima, N.; Setou, M. Imaging mass spectrometry for lipidomics. Biochim. Biophys. Acta 2011, 1811, 961–969. [Google Scholar] [CrossRef]
- Fujimura, Y.; Miura, D. MALDI Mass Spectrometry Imaging for Visualizing In Situ Metabolism of Endogenous Metabolites and Dietary Phytochemicals. Metabolites 2014, 4, 319–346. [Google Scholar] [CrossRef]
- Powers, T.W.; Holst, S.; Wuhrer, M.; Mehta, A.S.; Drake, R.R. Two-Dimensional N-glycan Distribution Mapping of Hepatocellular Carcinoma Tissues by MALDI-Imaging Mass Spectrometry. Biomolecules 2015, 5, 2554–2572. [Google Scholar] [CrossRef] [Green Version]
- Everest-Dass, A.V.; Briggs, M.T.; Kaur, G.; Oehler, M.K.; Hoffmann, P.; Packer, N.H. N-glycan MALDI Imaging Mass Spectrometry on Formalin-Fixed Paraffin-Embedded Tissue Enables the Delineation of Ovarian Cancer Tissues. Mol. Cell. Proteom. 2016, 15, 3003–3016. [Google Scholar] [CrossRef] [Green Version]
- Holst, S.; Heijs, B.; de Haan, N.; van Zeijl, R.J.; Briaire-de Bruijn, I.H.; van Pelt, G.W.; Mehta, A.S.; Angel, P.M.; Mesker, W.E.; Tollenaar, R.A.; et al. Linkage-Specific in Situ Sialic Acid Derivatization for N-glycan Mass Spectrometry Imaging of Formalin-Fixed Paraffin-Embedded Tissues. Anal. Chem. 2016, 88, 5904–5913. [Google Scholar] [CrossRef]
- Briggs, M.T.; Condina, M.R.; Ho, Y.Y.; Everest-Dass, A.V.; Mittal, P.; Kaur, G.; Oehler, M.K.; Packer, N.H.; Hoffmann, P. MALDI Mass Spectrometry Imaging of Early- and Late-Stage Serous Ovarian Cancer Tissue Reveals Stage-Specific N-glycans. Proteomics 2019, 19, e1800482. [Google Scholar] [CrossRef] [Green Version]
- Schultz, M.J.; Swindall, A.F.; Bellis, S.L. Regulation of the metastatic cell phenotype by sialylated glycans. Cancer Metastasis Rev. 2012, 31, 501–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrobono, S.; Stecca, B. Aberrant Sialylation in Cancer: Biomarker and Potential Target for Therapeutic Intervention? Cancers 2021, 13, 2014. [Google Scholar] [CrossRef] [PubMed]
- Ceroni, A.; Maass, K.; Geyer, H.; Geyer, R.; Dell, A.; Haslam, S.M. GlycoWorkbench: A tool for the computer-assisted annotation of mass spectra of glycans. J. Proteome Res. 2008, 7, 1650–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biskup, K.; Braicu, E.I.; Sehouli, J.; Fotopoulou, C.; Tauber, R.; Berger, M.; Blanchard, V. Serum glycome profiling: A biomarker for diagnosis of ovarian cancer. J. Proteome Res. 2013, 12, 4056–4063. [Google Scholar] [CrossRef] [PubMed]
- Biskup, K.; Braicu, E.I.; Sehouli, J.; Tauber, R.; Blanchard, V. The ascites N-glycome of epithelial ovarian cancer patients. J. Proteom. 2017, 157, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Dedova, T.; Braicu, E.I.; Sehouli, J.; Blanchard, V. Sialic Acid Linkage Analysis Refines the Diagnosis of Ovarian Cancer. Front. Oncol. 2019, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Kuzmanov, U.; Jiang, N.; Smith, C.R.; Soosaipillai, A.; Diamandis, E.P. Differential N-glycosylation of kallikrein 6 derived from ovarian cancer cells or the central nervous system. Mol. Cell. Proteom. 2009, 8, 791–798. [Google Scholar] [CrossRef] [Green Version]
- Young, C.; Condina, M.R.; Briggs, M.T.; Moh, E.S.X.; Kaur, G.; Oehler, M.K.; Hoffmann, P. In-House Packed Porous Graphitic Carbon Columns for Liquid Chromatography-Mass Spectrometry Analysis of N-glycans. Front. Chem. 2021, 9, 653959. [Google Scholar] [CrossRef]
- Anugraham, M.; Jacob, F.; Nixdorf, S.; Everest-Dass, A.V.; Heinzelmann-Schwarz, V.; Packer, N.H. Specific glycosylation of membrane proteins in epithelial ovarian cancer cell lines: Glycan structures reflect gene expression and DNA methylation status. Mol. Cell. Proteom. 2014, 13, 2213–2232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, Y.; Qian, Y.; Wu, X.; Zhang, Z.; Liu, X.; Zhao, R.; Zhou, L.; Ruan, Y.; Xu, J.; et al. Discovery of specific metastasis-related N-glycan alterations in epithelial ovarian cancer based on quantitative glycomics. PLoS ONE 2014, 9, e87978. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Deng, Z.; Huang, C.; Wu, H.; Zhao, X.; Li, Y. Mass spectrometric profiling reveals association of N-glycan patterns with epithelial ovarian cancer progression. Tumour Biol. 2017, 39, 1010428317716249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.D.; Phoomak, C.; Xu, G.; Olney, L.P.; Tran, K.A.; Park, S.S.; Haigh, N.E.; Luxardi, G.; Lert-Itthiporn, W.; Shimoda, M.; et al. Metastasis of cholangiocarcinoma is promoted by extended high-mannose glycans. Proc. Natl. Acad. Sci. USA 2020, 117, 7633–7644. [Google Scholar] [CrossRef] [PubMed]
- Hamester, F.; Legler, K.; Wichert, B.; Kelle, N.; Eylmann, K.; Rossberg, M.; Ding, Y.; Kurti, S.; Schmalfeldt, B.; Milde-Langosch, K.; et al. Prognostic relevance of the Golgi mannosidase MAN1A1 in ovarian cancer: Impact of N-glycosylation on tumour cell aggregation. Br. J. Cancer 2019, 121, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Swindall, A.F.; Bellis, S.L. Sialylation of the Fas death receptor by ST6Gal-I provides protection against Fas-mediated apoptosis in colon carcinoma cells. J. Biol. Chem. 2011, 286, 22982–22990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuo, Y.; Chammas, R.; Bellis, S.L. Sialylation of beta1 integrins blocks cell adhesion to galectin-3 and protects cells against galectin-3-induced apoptosis. J. Biol. Chem. 2008, 283, 22177–22185. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Christie, D.R.; Shaikh, F.M.; Lucas, J.A., IV; Lucas, J.A., III; Bellis, S.L. ST6Gal-I expression in ovarian cancer cells promotes an invasive phenotype by altering integrin glycosylation and function. J. Ovarian Res. 2008, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Kemmner, W.; Grigull, S.; Schlag, P.M. Cell surface alpha 2,6 sialylation affects adhesion of breast carcinoma cells. Exp. Cell Res. 2002, 276, 101–110. [Google Scholar] [CrossRef]
- Schultz, M.J.; Swindall, A.F.; Wright, J.W.; Sztul, E.S.; Landen, C.N.; Bellis, S.L. ST6Gal-I sialyltransferase confers cisplatin resistance in ovarian tumor cells. J. Ovarian Res. 2013, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Espina, V.; Wulfkuhle, J.D.; Calvert, V.S.; VanMeter, A.; Zhou, W.; Coukos, G.; Geho, D.H.; Petricoin, E.F., 3rd; Liotta, L.A. Laser-capture microdissection. Nat. Protoc. 2006, 1, 586–603. [Google Scholar] [CrossRef] [PubMed]
- Klein, O.; Kanter, F.; Kulbe, H.; Jank, P.; Denkert, C.; Nebrich, G.; Schmitt, W.D.; Wu, Z.; Kunze, C.A.; Sehouli, J.; et al. MALDI-Imaging for Classification of Epithelial Ovarian Cancer Histotypes from a Tissue Microarray Using Machine Learning Methods. Proteom. Clin. Appl. 2019, 13, e1700181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selman, M.H.; Hemayatkar, M.; Deelder, A.M.; Wuhrer, M. Cotton HILIC SPE microtips for microscale purification and enrichment of glycans and glycopeptides. Anal. Chem. 2011, 83, 2492–2499. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z [M + Na]+ | Composition | Neg | LGSOC | CCC | EC | BOT |
|---|---|---|---|---|---|---|
| 1136.40 | H3N3 | |||||
| 1257.42 | H5N2 | ✓ | ✓ | ✓ | ✓ | |
| 1282.45 | H3N3F1 | |||||
| 1298.45 | H4N3 | |||||
| 1339.48 | H3N4 | |||||
| 1419.48 | H6N2 | ✓ | ✓ | ✓ | ✓ | |
| 1444.51 | H4N3F1 | ✓ | ||||
| 1485.53 | H3N4F1 | ✓ | ✓ | ✓ | ✓ | |
| 1501.53 | H4N4 | ✓ | ✓ | |||
| 1581.53 | H7N2 | ✓ | ✓ | ✓ | ✓ | |
| 1616.60 | H4N3D1 | ✓ | ✓ | |||
| 1647.59 | H4N4F1 | ✓ | ✓ | ✓ | ||
| 1663.58 | H5N4 | ✓ | ✓ | ✓ | ||
| 1688.61 | H3N5F1 | ✓ | ||||
| 1704.61 | H4N5 | ✓ | ||||
| 1743.58 | H8N2 | ✓ | ✓ | ✓ | ✓ | |
| 1762.65 | H4N3F1D1 | ✓ | ||||
| 1778.64 | H5N3D1 | ✓ | ||||
| 1809.64 | H5N4F1 | ✓ | ✓ | ✓ | ✓ | |
| 1819.68 | H4N4D1 | ✓ | ||||
| 1825.63 | H6N4 | ✓ | ||||
| 1850.67 | H4N5F1 | ✓ | ✓ | ✓ | ||
| 1866.66 | H5N5 | ✓ | ||||
| 1891.69 | H3N6F1 | |||||
| 1905.63 | H9N2 | ✓ | ✓ | ✓ | ✓ | |
| 1937.70 | H4N4F1A1 | |||||
| 1953.70 | H5N4A1 | ✓ | ✓ | ✓ | ||
| 1965.73 | H4N4F1D1 | ✓ | ✓ | |||
| 1981.73 | H5N4D1 | ✓ | ✓ | ✓ | ✓ | |
| 2012.72 | H5N5F1 | ✓ | ✓ | ✓ | ✓ | |
| 2028.71 | H6N5 | ✓ | ||||
| 2053.75 | H4N6F1 | |||||
| 2099.76 | H5N4F1A1 | ✓ | ✓ | ✓ | ✓ | |
| 2127.79 | H5N4F1D1 | ✓ | ✓ | ✓ | ✓ | |
| 2168.82 | H4N5F1D1 | ✓ | ✓ | |||
| 2174.77 | H6N5F1 | ✓ | ✓ | |||
| 2184.80 | H5N5D1 | ✓ | ✓ | ✓ | ||
| 2243.81 | H5N4A2 | ✓ | ||||
| 2271.85 | H5N4A1D1 | ✓ | ✓ | ✓ | ||
| 2299.88 | H5N4D2 | ✓ | ✓ | ✓ | ✓ | |
| 2330.87 | H5N5F1D1 | ✓ | ✓ | ✓ | ✓ | |
| 2346.86 | H6N5D1 | ✓ | ✓ | ✓ | ||
| 2389.88 | H5N4F1A2 | ✓ | ✓ | ✓ | ✓ | |
| 2417.91 | H5N4F1A1D1 | ✓ | ✓ | ✓ | ||
| 2445.94 | H5N4F1D2 | ✓ | ✓ | ✓ | ✓ | |
| 2464.89 | H6N5F1A1 | ✓ | ✓ | ✓ | ||
| 2492.91 | H6N5F1D1 | ✓ | ✓ | ✓ | ✓ | |
| 2502.95 | H5N5D2 | ✓ | ✓ | ✓ | ||
| 2539.90 | H7N6F1 | |||||
| 2636.97 | H6N5A1D1 | ✓ | ✓ | |||
| 2649.00 | H5N5F1D2 | ✓ | ✓ | ✓ | ||
| 2665.00 | H6N5D2 | ✓ | ✓ | |||
| 2696.00 | H6N6F1D1 | ✓ | ✓ | ✓ | ||
| 2755.00 | H6N5F1A2 | ✓ | ✓ | |||
| 2783.03 | H6N5F1A1D1 | ✓ | ✓ | ✓ | ||
| 2811.06 | H6N5F1D2 | ✓ | ✓ | ✓ | ||
| 2830.01 | H7N6F1A1 | |||||
| 2858.05 | H7N6F1D1 | ✓ | ✓ | ✓ | ✓ | |
| 2955.11 | H6N5A1D2 | ✓ | ✓ | ✓ | ||
| 2983.14 | H6N5D3 | ✓ | ✓ | ✓ | ||
| 3045.12 | H6N5F1A3 | ✓ | ✓ | |||
| 3073.14 | H6N5F1A2D1 | ✓ | ✓ | |||
| 3101.17 | H6N5F1A1D2 | ✓ | ✓ | ✓ | ✓ | |
| 3148.16 | H7N6F1A1D1 | ✓ | ✓ | |||
| 3158.19 | H6N6A1D2 | ✓ | ✓ |
| Tumor vs. Tumor-stroma | |||||||||||
| LGSOC | CCC | EC | BOT | ||||||||
| m/z | Composition | AUC | m/z | Composition | AUC | m/z | Composition | AUC | m/z | Composition | AUC |
| - | - | - | 1743.6 | H8N2 | 0.658 | 1981.7 | H5N4D1 | 0.740 | 2330.9 | H5N5F1D1 | 0.806 |
| 2330.9 | H5N5F1D1 | 0.735 | 2649.0 | H5N5F1D2 | 0.775 | ||||||
| 2445.9 | H5N4F1D2 | 0.728 | 1905.6 | H9N2 | 0.759 | ||||||
| 2127.8 | H5N4F1D1 | 0.727 | 1419.5 | H6N2 | 0.758 | ||||||
| 1647.6 | H4N4F1 | 0.722 | 2346.9 | H6N5D1 | 0.732 | ||||||
| 2492.9 | H6N5F1D1 | 0.682 | 2696.0 | H6N6F1D1 | 0.724 | ||||||
| 1419.5 | H6N2 | 0.670 | 1581.5 | H7N2 | 0.719 | ||||||
| 1581.5 | H7N2 | 0.667 | 3101.2 | H6N5F1A1D2 | 0.700 | ||||||
| Tumor-stroma vs. Tumor | |||||||||||
| LGSOC | CCC | EC | BOT | ||||||||
| m/z | Composition | AUC | m/z | Composition | AUC | m/z | Composition | AUC | m/z | Composition | AUC |
| - | - | - | 2099.8 | H5N4F1A1 | 0.901 | 2389.9 | H5N4F1A2 | 0.757 | 2099.8 | H5N4F1A1 | 0.743 |
| 1953.7 | H5N4A1 | 0.825 | 2099.8 | H5N4A1 | 0.734 | 1809.6 | H5N4F1 | 0.670 | |||
| 1981.7 | H5N4D1 | 0.822 | 2012.7 | H5N5F1 | 0.662 | ||||||
| 2955.1 | H6N5A1D2 | 0.815 | |||||||||
| 2665.0 | H6N5D2 | 0.774 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzeski, M.; Taube, E.T.; Braicu, E.I.; Sehouli, J.; Blanchard, V.; Klein, O. In Situ N-Glycosylation Signatures of Epithelial Ovarian Cancer Tissue as Defined by MALDI Mass Spectrometry Imaging. Cancers 2022, 14, 1021. https://doi.org/10.3390/cancers14041021
Grzeski M, Taube ET, Braicu EI, Sehouli J, Blanchard V, Klein O. In Situ N-Glycosylation Signatures of Epithelial Ovarian Cancer Tissue as Defined by MALDI Mass Spectrometry Imaging. Cancers. 2022; 14(4):1021. https://doi.org/10.3390/cancers14041021
Chicago/Turabian StyleGrzeski, Marta, Eliane T. Taube, Elena I. Braicu, Jalid Sehouli, Véronique Blanchard, and Oliver Klein. 2022. "In Situ N-Glycosylation Signatures of Epithelial Ovarian Cancer Tissue as Defined by MALDI Mass Spectrometry Imaging" Cancers 14, no. 4: 1021. https://doi.org/10.3390/cancers14041021