
CDK7 Predicts Worse Outcome in Head and Neck Squamous-Cell Cancer

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tumor Material
2.2. Immunohistochemistry
2.3. Digitalization and Evaluation
2.4. Statistics and Visualization
3. Results
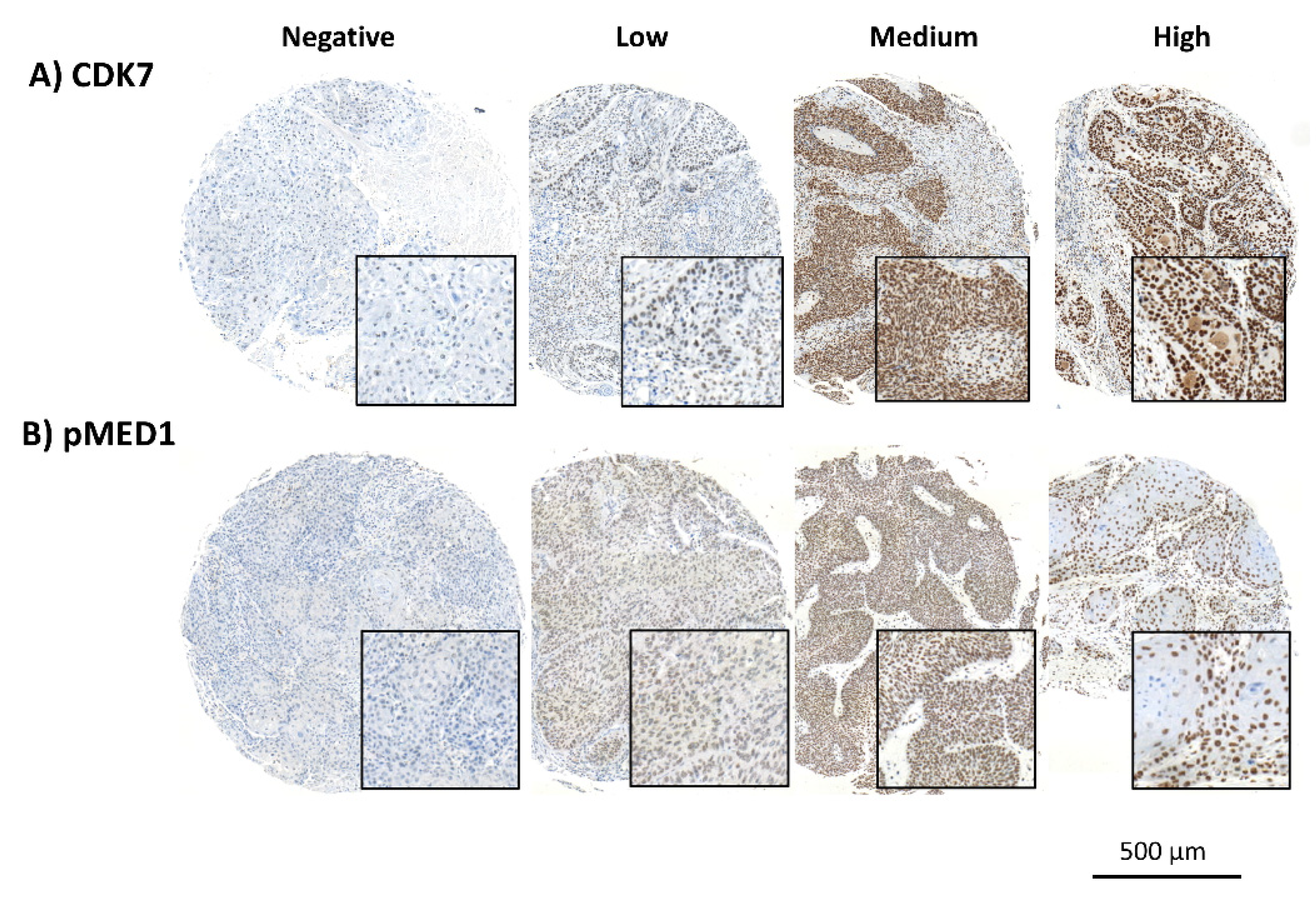
3.1. Distinct Staining
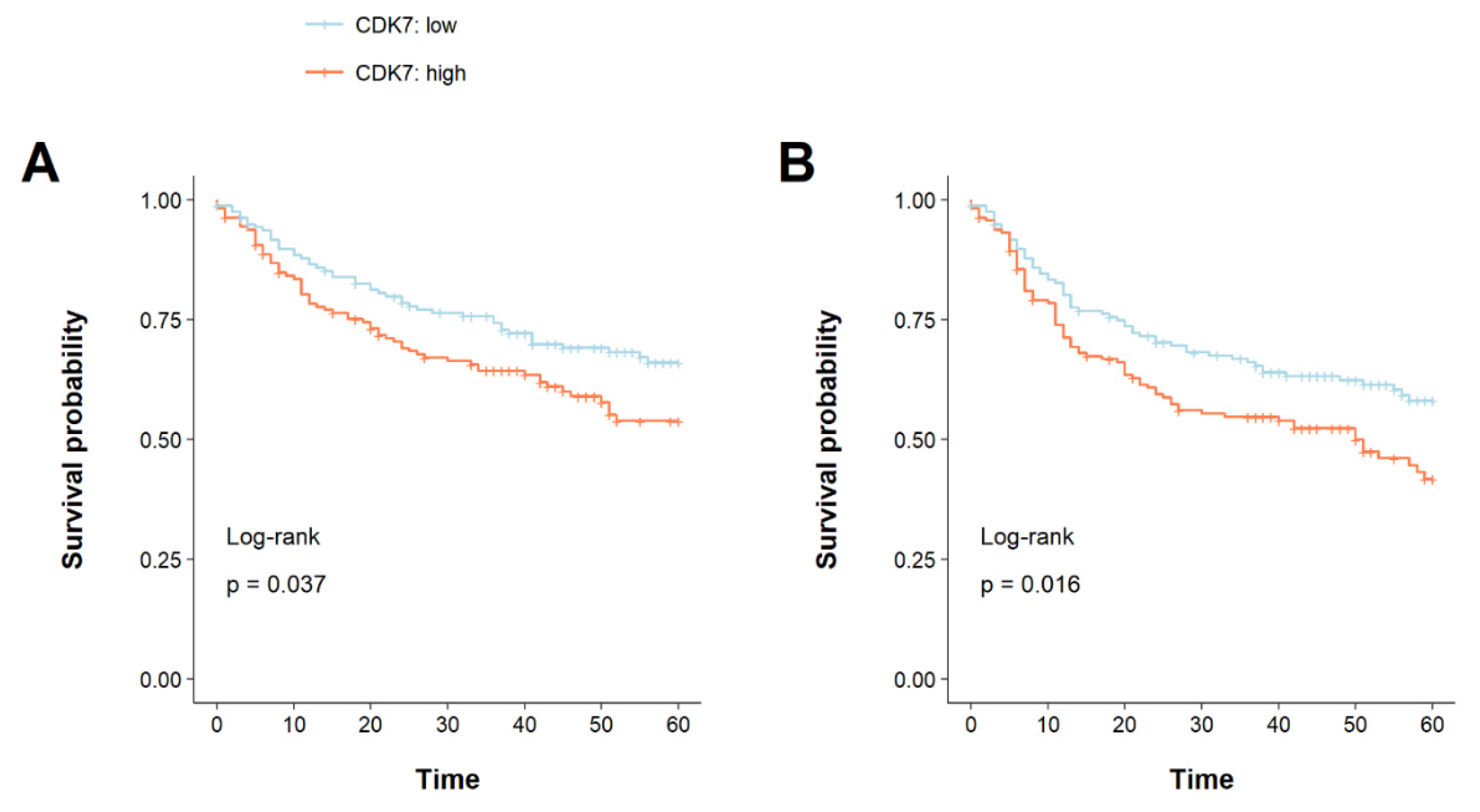
3.2. CDK7 Overexpression Is Associated with a Shorter Overall and Disease-Free Survival
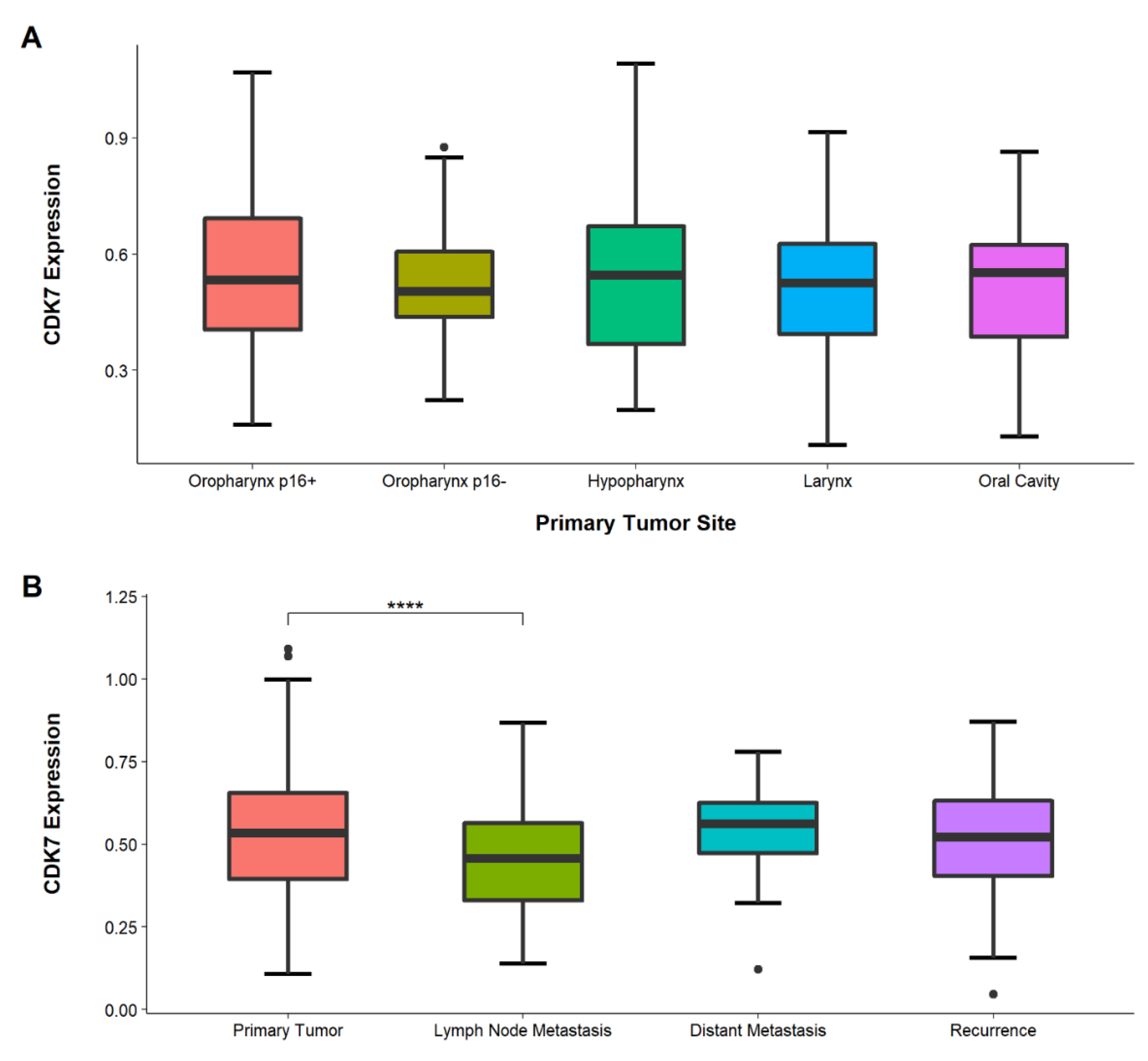
3.3. Congruent CDK7 Expression in Different Primary Tumor Locations
3.4. Lower CDK7 Expression in Lymph-Node Metastasis Than in Primary Tumors
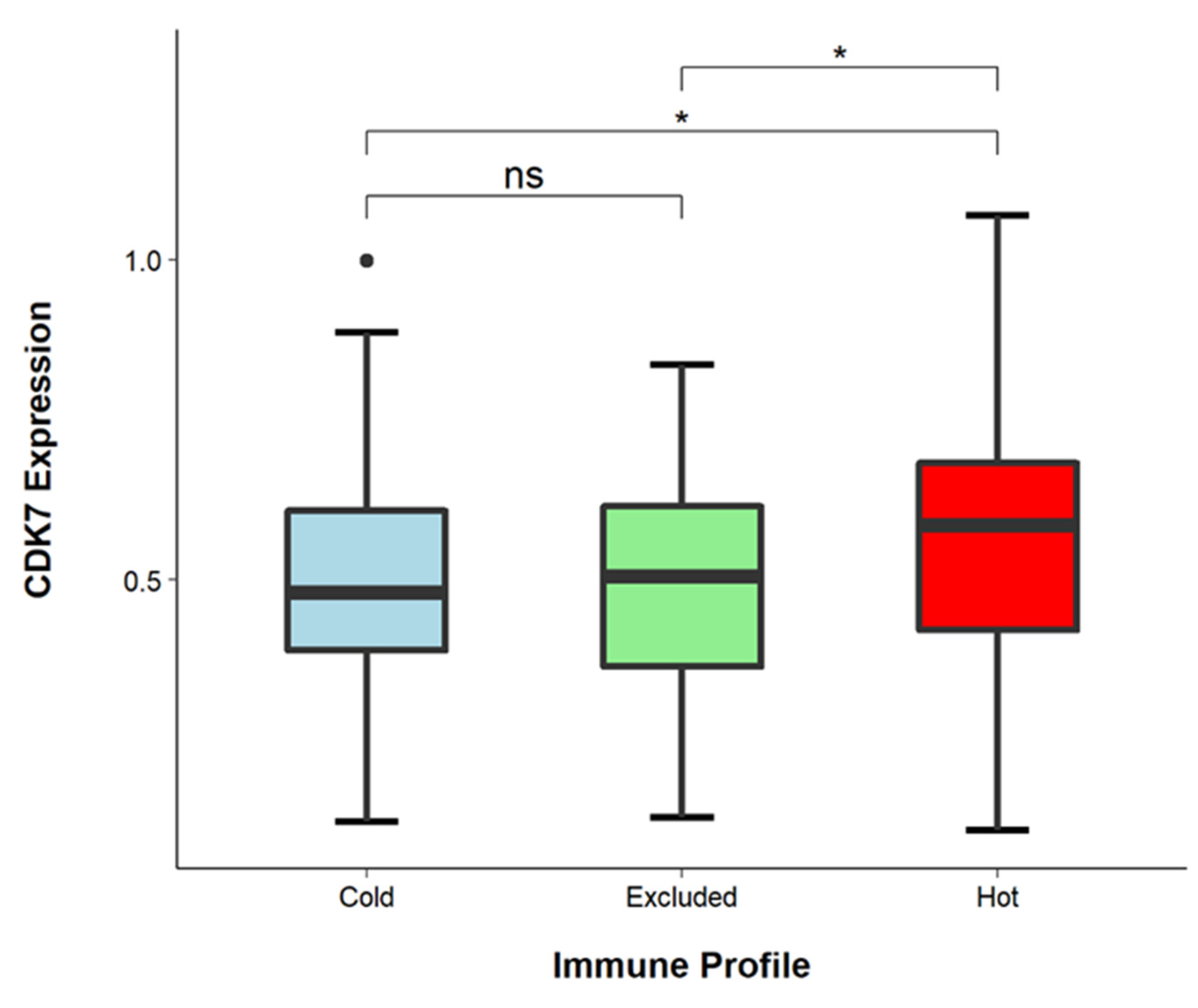
3.5. CDK7 Levels Are Higher in Immune-Infiltrated Tumors
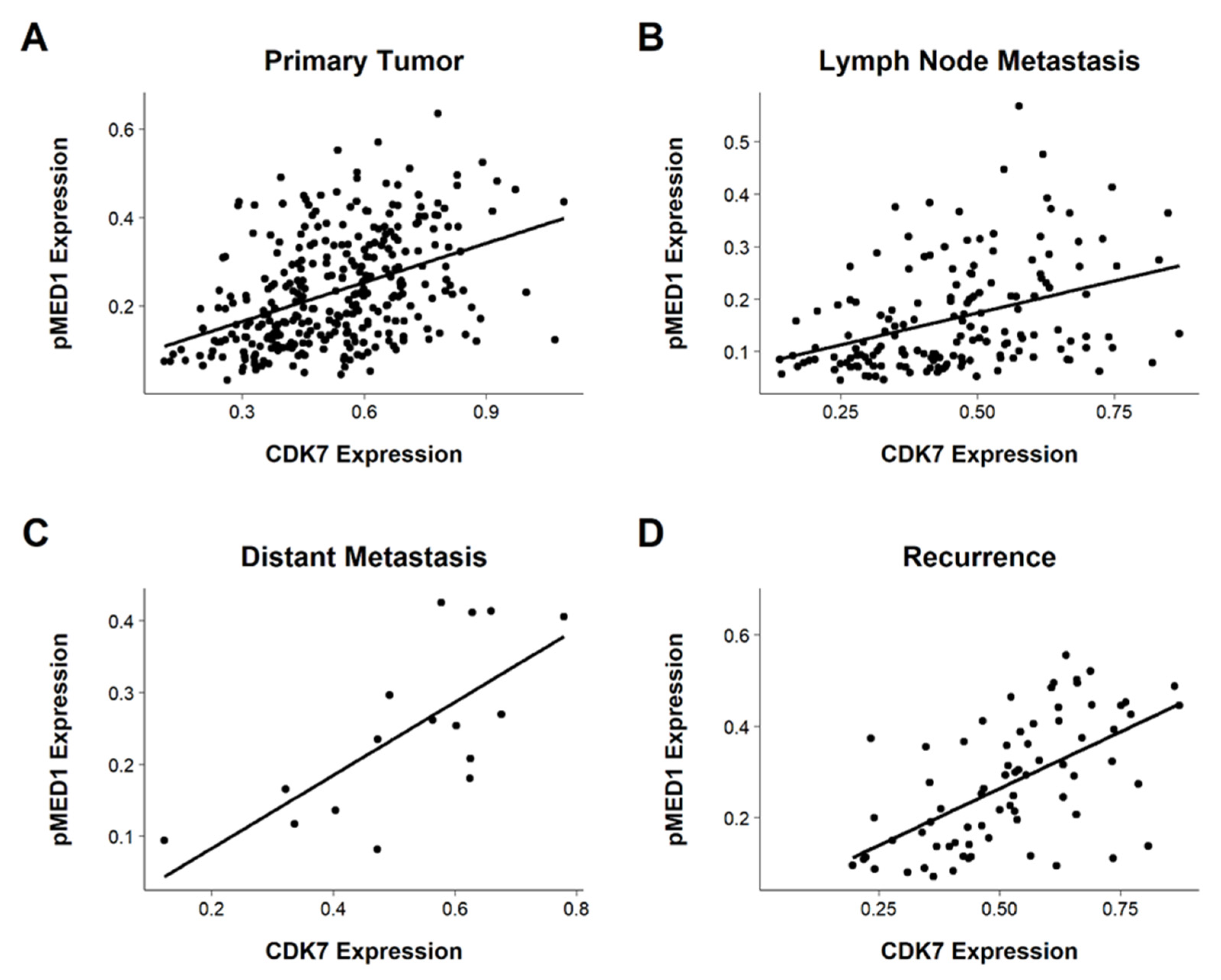
3.6. Correlating CDK7 and pMED1 Expression
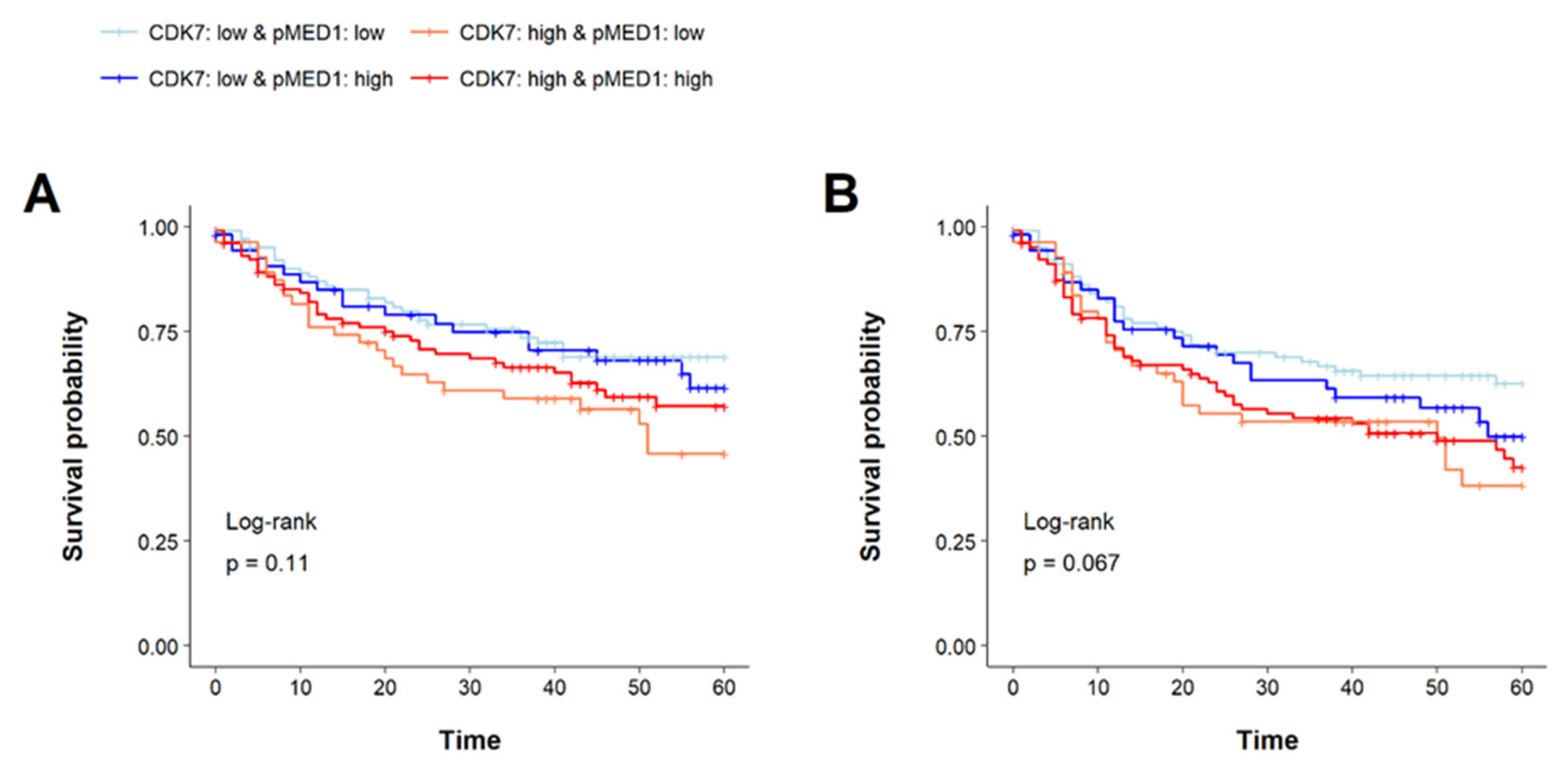
3.7. Prognostic Implications of CDK7 and pMED1 Co-Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| CAK | CDK-activating kinase |
| CDK7 | Cyclin-dependent kinase 7 |
| CI | Confidence interval |
| DAB | 3,3’-diaminobenzidine |
| DFS | Disease-free survival |
| DM | Distant metastasis |
| DSIF | DRB-sensitivity-inducing factor |
| FFPE | Formalin-fixed paraffin-embedded |
| HNSCC | Head and neck squamous-cell cancer |
| HPV | Human papilloma virus |
| HR | Hazard ratio |
| IHC | Immunhistochemistry |
| LN | Lymph node metastasis |
| MAPK-ERK | MAP-kinase |
| NELF | Negative elongation factor |
| NR | Nuclear receptor |
| NSCLC | Non-small-cell lung cancer |
| OS | Overall survival |
| PCC | Pearson correlation coefficient |
| PD-L1 | Programmed death ligand 1 |
| pMED1 | Phospho-mediator of RNA polymerase II transcription subunit 1 |
| PT | Primary tumor |
| RD | Recurrence |
| RNA Pol II | RNA polymerase II |
| ROI | Region of interest |
| SE | Super-enhancer |
| TMA | Tissue microarray |
| TPS | Tumor proportion score |
| UICC | Union internationale contre le cancer |
References
- Chow, L.Q.M. Head and neck cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.I.; Westra, W.H. Molecular pathology of head and neck cancer: Implications for diagnosis, prognosis, and treatment. Annu. Rev. Pathol. 2009, 4, 49–70. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primer 2020, 6, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Denaro, N.; Merlano, M.C.; Russi, E.G. Follow-up in head and neck cancer: Do more does it mean do better? A systematic review and our proposal based on our experience. Clin. Exp. Otorhinolaryngol. 2016, 9, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Canning, M.; Guo, G.; Yu, M.; Myint, C.; Groves, M.W.; Byrd, J.K.; Cui, Y. Heterogeneity of the head and neck squamous cell carcinoma immune landscape and its impact on immunotherapy. Front. Cell Dev. Biol. 2019, 7, 52. [Google Scholar] [CrossRef]
- Bennardo, L.; Bennardo, F.; Giudice, A.; Passante, M.; Dastoli, S.; Morrone, P.; Provenzano, E.; Patruno, C.; Nisticò, S.P. Local chemotherapy as an adjuvant treatment in unresectable squamous cell carcinoma: What do we know so far? Curr. Oncol. 2021, 28, 2317–2325. [Google Scholar] [CrossRef]
- Mehanna, H.; Robinson, M.; Hartley, A.; Kong, A.; Foran, B.; Fulton-Lieuw, T.; Dalby, M.; Mistry, P.; Sen, M.; O’Toole, L.; et al. Radiotherapy plus cisplatin or cetuximab in low-risk human papillomavirus-positive oropharyngeal cancer (De-ESCALaTE HPV): An open-label randomised controlled phase 3 trial. Lancet Lond. Engl. 2019, 393, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, S.; Tang, Y.-J.; Chen, Y.; Zheng, M.; Tang, Y.; Liang, X. The double-edged sword—How human papillomaviruses interact with immunity in head and neck cancer. Front. Immunol. 2019, 10, 653. [Google Scholar] [CrossRef]
- Kim, J.; Cho, Y.-J.; Ryu, J.-Y.; Hwang, I.; Han, H.D.; Ahn, H.J.; Kim, W.Y.; Cho, H.; Chung, J.-Y.; Hewitt, S.M.; et al. CDK7 Is a reliable prognostic factor and novel therapeutic target in epithelial ovarian cancer. Gynecol. Oncol. 2020, 156, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Bryant, A.K.; Sojourner, E.J.; Vitzthum, L.K.; Zakeri, K.; Shen, H.; Nguyen, C.; Murphy, J.D.; Califano, J.A.; Cohen, E.E.W.; Mell, L.K. Prognostic role of P16 in nonoropharyngeal head and neck cancer. J. Natl. Cancer Inst. 2018, 110, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Shen, Z.; Ye, D.; Li, Q.; Deng, H.; Liu, H.; Li, J. The association and clinical significance of CDKN2A promoter methylation in head and neck squamous cell carcinoma: A meta-analysis. Cell. Physiol. Biochem. 2018, 50, 868–882. [Google Scholar] [CrossRef]
- Neskey, D.M.; Osman, A.A.; Ow, T.J.; Katsonis, P.; McDonald, T.; Hicks, S.C.; Hsu, T.-K.; Pickering, C.R.; Ward, A.; Patel, A.; et al. Evolutionary action score of TP53 (EAp53) identifies high risk mutations associated with decreased survival and increased distant metastases in head and neck cancer. Cancer Res. 2015, 75, 1527–1536. [Google Scholar] [CrossRef] [Green Version]
- Almani, L.; Bellile, E.; Spector, M.; Smith, J.; Brenner, C.; Rozek, L.; Nguyen, A.; Zarins, K.; Thomas, D.; McHugh, J.; et al. Expression of P53 and prognosis in patients with head and neck squamous cell carcinoma (HNSCC). Int. J. Cancer Clin. Res. 2019, 6, 122. [Google Scholar] [CrossRef] [Green Version]
- Bradner, J.E.; Hnisz, D.; Young, R.A. Transcriptional addiction in cancer. Cell 2017, 168, 629–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, S.; George, R.E. Super-enhancer-driven transcriptional dependencies in cancer. Trends Cancer 2017, 3, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galbraith, M.D.; Bender, H.; Espinosa, J.M. Therapeutic targeting of transcriptional cyclin-dependent kinases. Transcription 2019, 10, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Geng, J.; Zhang, L.; Wang, Y.; Niu, N.; Fang, Y.; Liu, F.; Shi, J.; Zhang, Z.-G.; Sun, Y.-W.; et al. THZ1 reveals CDK7-dependent transcriptional addictions in pancreatic cancer. Oncogene 2019, 38, 3932–3945. [Google Scholar] [CrossRef]
- Coin, F.; Egly, J.-M. Revisiting the function of CDK7 in transcription by virtue of a recently described TFIIH kinase inhibitor. Mol. Cell 2015, 59, 513–514. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, T.; Kwiatkowski, N.; Abraham, B.J.; Lee, T.I.; Xie, S.; Yuzugullu, H.; Von, T.; Li, H.; Lin, Z.; et al. CDK7-dependent transcriptional addiction in triple-negative breast cancer. Cell 2015, 163, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.-Y.; Lin, D.-C.; Mayakonda, A.; Hazawa, M.; Ding, L.-W.; Chien, W.-W.; Xu, L.; Chen, Y.; Xiao, J.-F.; Senapedis, W.; et al. Targeting super-enhancer-associated oncogenes in oesophageal squamous cell carcinoma. Gut 2017, 66, 1358–1368. [Google Scholar] [CrossRef] [Green Version]
- Sava, G.P.; Fan, H.; Coombes, R.C.; Buluwela, L.; Ali, S. CDK7 inhibitors as anticancer drugs. Cancer Metastasis Rev. 2020, 39, 805–823. [Google Scholar] [CrossRef]
- Ge, H.; Yao, Y.; Jiang, Y.; Wu, X.; Wang, Y. Pharmacological inhibition of CDK7 by THZ1 impairs tumor growth in P53-mutated HNSCC. Oral Dis. 2021. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Christensen, C.L.; Dries, R.; Oser, M.G.; Deng, J.; Diskin, B.; Li, F.; Pan, Y.; Zhang, X.; Yin, Y.; et al. CDK7 inhibition potentiates genome instability triggering anti-tumor immunity in small cell lung cancer. Cancer Cell 2020, 37, 37–54.e9. [Google Scholar] [CrossRef]
- Kwiatkowski, N.; Zhang, T.; Rahl, P.B.; Abraham, B.J.; Reddy, J.; Ficarro, S.B.; Dastur, A.; Amzallag, A.; Ramaswamy, S.; Tesar, B.; et al. Targeting transcription regulation in cancer with a covalent CDK7 inhibitor. Nature 2014, 511, 616–620. [Google Scholar] [CrossRef] [Green Version]
- Ur Rasool, R.; Natesan, R.; Deng, Q.; Aras, S.; Lal, P.; Sander Effron, S.; Mitchell-Velasquez, E.; Posimo, J.M.; Carskadon, S.; Baca, S.C.; et al. CDK7 inhibition suppresses castration-resistant prostate cancer through MED1 inactivation. Cancer Discov. 2019, 9, 1538–1555. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ge, H.; Jiang, Y.; Huang, R.; Wu, Y.; Wang, D.; Guo, S.; Li, S.; Wang, Y.; Jiang, H.; et al. Combinational therapeutic targeting of BRD4 and CDK7 synergistically induces anticancer effects in head and neck squamous cell carcinoma. Cancer Lett. 2020, 469, 510–523. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.; Mei, H.; Zhang, D.; Wu, G.; Zhang, T.; Lin, Z. Antitumor effects of a covalent cyclin-dependent kinase 7 inhibitor in colorectal cancer. Anticancer. Drugs 2019, 30, 466–474. [Google Scholar] [CrossRef]
- Belakavadi, M.; Pandey, P.K.; Vijayvargia, R.; Fondell, J.D. MED1 phosphorylation promotes its association with mediator: Implications for nuclear receptor signaling. Mol. Cell. Biol. 2008, 28, 3932–3942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gade, P.; Singh, A.K.; Roy, S.K.; Reddy, S.P.; Kalvakolanu, D.V. Down regulation of the transcriptional mediator subunit Med1 contributes to the loss of expression of metastasis associated Dapk1 in human cancers and cancer cells. Int. J. Cancer J. Int. Cancer 2009, 125, 1566–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.; Son, C.H.; Um, S.J.; Kwon, H.C.; Lee, K.E.; Choi, P.J.; Roh, M.S. A different TRAP220 expression in distinct histologic subtypes of lung adenocarcinoma and the prognostic significance. Lung Cancer Amst. Neth. 2011, 71, 312–318. [Google Scholar] [CrossRef]
- Kim, H.-J.; Roh, M.S.; Son, C.H.; Kim, A.J.; Jee, H.J.; Song, N.; Kim, M.; Seo, S.-Y.; Yoo, Y.H.; Yun, J. Loss of Med1/TRAP220 promotes the invasion and metastasis of human non-small-cell lung cancer cells by modulating the expression of metastasis-related genes. Cancer Lett. 2012, 321, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, M.; Zhang, X.; Huang, H.; Huang, J.; Ke, J.; Ding, H.; Xiao, J.; Shan, X.; Liu, Q.; et al. Upregulation of CDK7 in gastric cancer cell promotes tumor cell proliferation and predicts poor prognosis. Exp. Mol. Pathol. 2016, 100, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, X.; Wang, Y.; Shi, H.; Guan, C.; Yao, L.; Huang, X.; Ding, Z.; Huang, Y.; Wang, H.; et al. Low expression of cyclinh and cyclin-dependent kinase 7 can decrease the proliferation of human esophageal squamous cell carcinoma. Dig. Dis. Sci. 2013, 58, 2028–2037. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, R.; Wu, Y.; Diao, P.; Zhang, W.; Li, J.; Li, Z.; Wang, Y.; Cheng, J.; Yang, J. Overexpression of CDK7 is associated with unfavourable prognosis in oral squamous cell carcinoma. Pathology 2019, 51, 74–80. [Google Scholar] [CrossRef]
- Klümper, N.; Syring, I.; Vogel, W.; Schmidt, D.; Müller, S.C.; Ellinger, J.; Shaikhibrahim, Z.; Brägelmann, J.; Perner, S. Mediator complex subunit MED1 protein expression is decreased during bladder cancer progression. Front. Med. 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Idel, C.; Ribbat-Idel, J.; Kuppler, P.; Krupar, R.; Offermann, A.; Vogel, W.; Rades, D.; Kirfel, J.; Wollenberg, B.; Perner, S. EVI1 as a marker for lymph node metastasis in HNSCC. Int. J. Mol. Sci. 2020, 21, 854. [Google Scholar] [CrossRef] [Green Version]
- Klapper, L.; Ribbat-Idel, J.; Kuppler, P.; Paulsen, F.-O.; Bruchhage, K.-L.; Rades, D.; Offermann, A.; Kirfel, J.; Wollenberg, B.; Idel, C.; et al. NR2F6 as a prognostic biomarker in HNSCC. Int. J. Mol. Sci. 2020, 21, 5527. [Google Scholar] [CrossRef]
- Paulsen, F.-O.; Idel, C.; Ribbat-Idel, J.; Kuppler, P.; Klapper, L.; Rades, D.; Bruchhage, K.-L.; Wollenberg, B.; Brägelmann, J.; Perner, S.; et al. CDK19 as a potential HPV-independent biomarker for recurrent disease in HNSCC. Int. J. Mol. Sci. 2020, 21, 5508. [Google Scholar] [CrossRef] [PubMed]
- Ribbat-Idel, J.; Perner, S.; Kuppler, P.; Klapper, L.; Krupar, R.; Watermann, C.; Paulsen, F.-O.; Offermann, A.; Bruchhage, K.-L.; Wollenberg, B.; et al. Immunologic “cold” squamous cell carcinomas of the head and neck are associated with an unfavorable prognosis. Front. Med. 2021, 8, 622330. [Google Scholar] [CrossRef] [PubMed]
- Ribbat-Idel, J.; Dressler, F.F.; Krupar, R.; Watermann, C.; Paulsen, F.-O.; Kuppler, P.; Klapper, L.; Offermann, A.; Wollenberg, B.; Rades, D.; et al. Performance of different diagnostic PD-L1 clones in head and neck squamous cell carcinoma. Front. Med. 2021, 8, 640515. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Goltz, D.; Shaikhibrahim, Z.; Vogel, W.; Böhm, D.; Scheble, V.; Sotlar, K.; Fend, F.; Tan, S.-H.; Dobi, A.; et al. ERG protein expression and genomic rearrangement status in primary and metastatic prostate cancer—A comparative study of two monoclonal antibodies. Prostate Cancer Prostatic Dis. 2012, 15, 165–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheble, V.J.; Braun, M.; Beroukhim, R.; Mermel, C.H.; Ruiz, C.; Wilbertz, T.; Stiedl, A.-C.; Petersen, K.; Reischl, M.; Kuefer, R.; et al. ERG rearrangement is specific to prostate cancer and does not occur in any other common tumor. Mod. Pathol. 2010, 23, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Germer, K.; Wu, T.; Wang, J.; Luo, J.; Wang, S.; Wang, Q.; Zhang, X. Cross-talk between HER2 and MED1 regulates tamoxifen resistance of human breast cancer cells. Cancer Res. 2012, 72, 5625–5634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweitzer, A.; Knauer, S.K.; Stauber, R.H. Nuclear receptors in head and neck cancer: Current knowledge and perspectives. Int. J. Cancer 2010, 126, 801–809. [Google Scholar] [CrossRef]
- Dong, J.; Li, J.; Li, Y.; Ma, Z.; Yu, Y.; Wang, C.-Y. Transcriptional super-enhancers control cancer stemness and metastasis genes in squamous cell carcinoma. Nat. Commun. 2021, 12, 3974. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, R.; Lin, Z.; Zhang, S.; Chen, Y.; Tang, J.; Hong, J.; Zhou, X.; Zong, Y.; Xu, Y.; et al. CDK7 inhibitor THZ1 enhances AntiPD-1 therapy efficacy via the P38α/MYC/PD-L1 signaling in non-small cell lung cancer. J. Hematol. Oncol.J Hematol. Oncol. 2020, 13, 99. [Google Scholar] [CrossRef]
- Idel, C.; Ribbat-Idel, J.; Klapper, L.; Krupar, R.; Bruchhage, K.-L.; Dreyer, E.; Rades, D.; Polasky, C.; Offermann, A.; Kirfel, J.; et al. Spatial distribution of immune cells in head and neck squamous cell carcinomas. Front. Oncol. 2021, 11, 712788. [Google Scholar] [CrossRef]
- Patel, H.; Periyasamy, M.; Sava, G.P.; Bondke, A.; Slafer, B.W.; Kroll, S.H.B.; Barbazanges, M.; Starkey, R.; Ottaviani, S.; Harrod, A.; et al. ICEC0942, an orally bioavailable selective inhibitor of CDK7 for cancer treatment. Mol. Cancer Ther. 2018, 17, 1156–1166. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | CDK7 Expression | Total (n = 326) | p Value | |
|---|---|---|---|---|
| CDK7 Low (n = 161) | CDK7 High (n = 165) | |||
| Age | 0.355 | |||
| Missing | 0 | 2 | 2 | |
| Mean (SD) | 61.826 (10.126) | 62.890 (10.527) | 62.361 (10.328) | |
| Sex | 0.882 | |||
| Missing | 1 | 0 | 1 | |
| Female | 36 (22.5%) | 36 (21.8%) | 72 (22.2%) | |
| Male | 124 (77.5%) | 129 (78.2%) | 253 (77.8%) | |
| Alcohol | 0.384 | |||
| Missing | 5 | 6 | 11 | |
| No | 91 (58.3%) | 85 (53.5%) | 176 (55.9%) | |
| Yes | 65 (41.7%) | 74 (46.5%) | 139 (44.1%) | |
| Nicotine | 0.854 | |||
| Missing | 11 | 6 | 17 | |
| No | 16 (10.7%) | 18 (11.3%) | 34 (11.0%) | |
| Yes | 134 (89.3%) | 141 (88.7%) | 275 (89.0%) | |
| Packyears | 0.608 | |||
| Missing | 11 | 6 | 17 | |
| Mean (SD) | 39.173 (26.669) | 37.673 (24.659) | 38.401 (25.624) | |
| p16 | 0.618 | |||
| Negative | 121 (75.2%) | 120 (72.7%) | 241 (73.9%) | |
| Positive | 40 (24.8%) | 45 (27.3%) | 85 (26.1%) | |
| Location | 0.716 | |||
| Missing | 5 | 4 | 9 | |
| CUP | 1 (0.6%) | 0 (0.0%) | 1 (0.3%) | |
| Hypopharynx | 20 (12.8%) | 24 (14.9%) | 44 (13.9%) | |
| Larynx | 47 (30.1%) | 46 (28.6%) | 93 (29.3%) | |
| Oral Cavity | 33 (21.2%) | 41 (25.5%) | 74 (23.3%) | |
| Oropharynx p16- | 28 (17.9%) | 22 (13.7%) | 50 (15.8%) | |
| Oropharynx p16+ | 27 (17.3%) | 28 (17.4%) | 55 (17.4%) | |
| Recurrence | 0.431 | |||
| No | 123 (76.4%) | 132 (80.0%) | 255 (78.2%) | |
| Yes | 38 (23.6%) | 33 (20.0%) | 71 (21.8%) | |
| T-Stage | 0.816 | |||
| Missing | 2 | 0 | 2 | |
| T (1,2) | 83 (52.2%) | 84 (50.9%) | 167 (51.5%) | |
| T (3,4) | 76 (47.8%) | 81 (49.1%) | 157 (48.5%) | |
| N-Stage | 0.228 | |||
| Missing | 1 | 1 | 2 | |
| N- | 77 (48.1%) | 68 (41.5%) | 145 (44.8%) | |
| N+ | 83 (51.9%) | 96 (58.5%) | 179 (55.2%) | |
| M-Stage | 0.112 | |||
| Missing | 0 | 1 | 1 | |
| M- | 145 (90.1%) | 138 (84.1%) | 283 (87.1%) | |
| M+ | 16 (9.9%) | 26 (15.9%) | 42 (12.9%) | |
| UICC Stage | 0.566 | |||
| Missing | 1 | 0 | 1 | |
| UICC (1,2) | 60 (37.5%) | 67 (40.6%) | 127 (39.1%) | |
| UICC (3,4) | 100 (62.5%) | 98 (59.4%) | 198 (60.9%) | |
| Grading | 0.953 | |||
| Missing | 3 | 0 | 3 | |
| G (1,2) | 123 (77.8%) | 128 (77.6%) | 251 (77.7%) | |
| G (3,4) | 35 (22.2%) | 37 (22.4%) | 72 (22.3%) | |
| 5-Year Overall Survival | Univariable Survival Analysis | Multivariable Survival Analysis | ||||
| Variable | HR | 95% CI | p Value | HR | 95% CI | p Value |
| Alcohol consumption | 1.74 | 1.23–2.45 | 0.002 | 1.17 | 0.78–1.76 | 0.457 |
| Packyears | 1.01 | 1.00–1.02 | <0.001 | 1.01 | 1.00–1.02 | 0.018 * |
| p16 status | 0.48 | 0.31–0.74 | <0.001 | 0.70 | 0.40–1.22 | 0.205 |
| T-stage | 2.29 | 1.61–3.25 | <0.001 | 1.24 | 0.73–2.12 | 0.422 |
| M-stage | 1.98 | 1.31–2.99 | <0.001 | 2.10 | 1.32–3.35 | 0.002 * |
| UICC | 2.83 | 1.91–4.19 | <0.001 | 1.87 | 1.00–3.50 | 0.050 |
| CDK7 | 1.48 | 1.02–2.15 | 0.038 | 1.50 | 1.01–2.22 | 0.045 * |
| 5-Year Disease-Free Survival | Univariable Survival Analysis | Multivariable Survival Analysis | ||||
| Variable | HR | 95% CI | p Value | HR | 95% CI | p Value |
| Age | 1.02 | 1.00–1.03 | 0.012 | 1.03 | 1.01–1.05 | 0.007 * |
| Alcohol consumption | 1.47 | 1.09–1.98 | 0.011 | 1.25 | 0.86–1.82 | 0.250 |
| Packyears | 1.01 | 1.00–1.01 | 0.022 | 1.00 | 1.00–1.01 | 0.453 |
| p16 status | 0.47 | 0.32–0.68 | <0.001 | 0.48 | 0.29–0.81 | 0.006 * |
| T-stage | 2.13 | 1.57–2.87 | <0.001 | 1.44 | 0.88–2.36 | 0.144 |
| M-stage | 1.66 | 1.14–2.43 | 0.008 | 1.80 | 1.14–2.84 | 0.011 * |
| UICC | 2.29 | 1.66–3.16 | <0.001 | 1.40 | 0.80–2.43 | 0.237 |
| CDK7 | 1.51 | 1.08–2.10 | 0.016 | 1.50 | 1.05–2.14 | 0.024 * |
| Clinicopathological Feature | n | p Value |
|---|---|---|
| Sex | Male (n = 72) vs. Female (n = 253) | 0.59 |
| Age | ≤62 years (n = 152) vs. >62 years (n = 172) | 0.17 |
| Alcohol consumption | No (n = 176) vs. Yes (n = 139) | 0.14 |
| Nicotine consumption | No (n = 34) vs. Yes (n = 275) | 0.4 |
| Recurrent disease | No (n = 255) vs. Yes (n = 71) | 0.75 |
| p16 status | Negative (n = 241) vs. Positive (n = 85) | 0.15 |
| T-stage | T (1,2) (n = 167) vs. T (3,4) (n = 157) | 0.73 |
| N-stage | N0 (n = 145) vs. N+ (n = 179) | 0.06 |
| M-stage | M0 (n = 283) vs. M+ (n = 42) | 0.06 |
| UICC-stage | UICC (I,II) (n = 127) vs. UICC (III,IV) (n = 198) | 0.89 |
| Grading | G (1,2) (n = 251) vs. G (3,4) (n = 72) | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jagomast, T.; Idel, C.; Klapper, L.; Kuppler, P.; Offermann, A.; Dreyer, E.; Bruchhage, K.-L.; Ribbat-Idel, J.; Perner, S. CDK7 Predicts Worse Outcome in Head and Neck Squamous-Cell Cancer. Cancers 2022, 14, 492. https://doi.org/10.3390/cancers14030492
Jagomast T, Idel C, Klapper L, Kuppler P, Offermann A, Dreyer E, Bruchhage K-L, Ribbat-Idel J, Perner S. CDK7 Predicts Worse Outcome in Head and Neck Squamous-Cell Cancer. Cancers. 2022; 14(3):492. https://doi.org/10.3390/cancers14030492
Chicago/Turabian StyleJagomast, Tobias, Christian Idel, Luise Klapper, Patrick Kuppler, Anne Offermann, Eva Dreyer, Karl-Ludwig Bruchhage, Julika Ribbat-Idel, and Sven Perner. 2022. "CDK7 Predicts Worse Outcome in Head and Neck Squamous-Cell Cancer" Cancers 14, no. 3: 492. https://doi.org/10.3390/cancers14030492