
CYP1B1: A Novel Molecular Biomarker Predicts Molecular Subtype, Tumor Microenvironment, and Immune Response in 33 Cancers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Differential Gene Expression Analysis
2.3. Survival Analysis
2.4. Correlation Analysis between CYP1B1 and Tumor Immune System
2.5. Single-Sample Gene Set Enrichment Analysis (ssGSEA)
2.6. Association between CYP1B1 and Drug Response
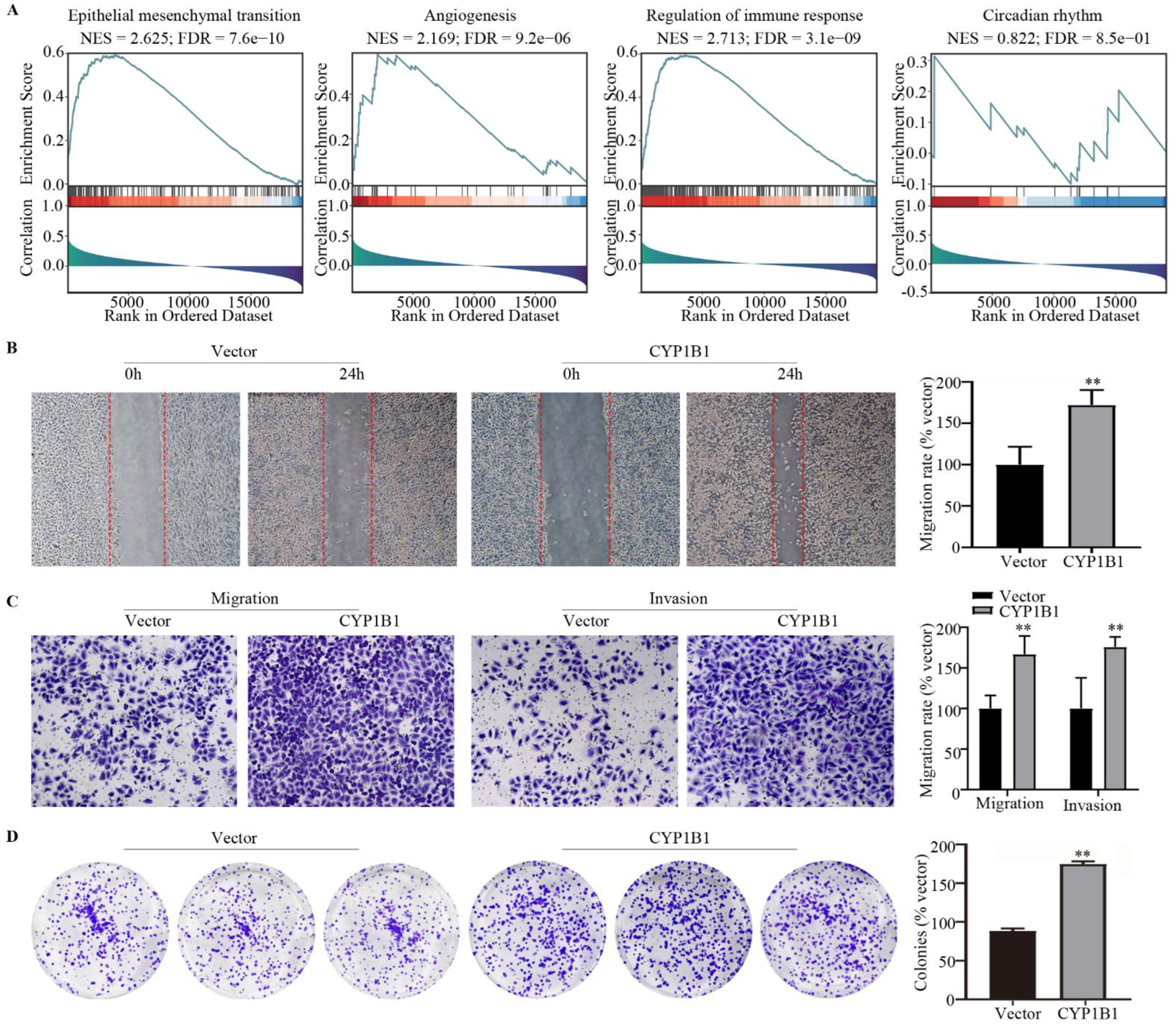
2.7. Gene Set Enrichment Analysis (GSEA)
2.8. Cell Culture
2.9. Wound Healing Assay
2.10. Transwell Assay
2.11. Cell Proliferation Assay
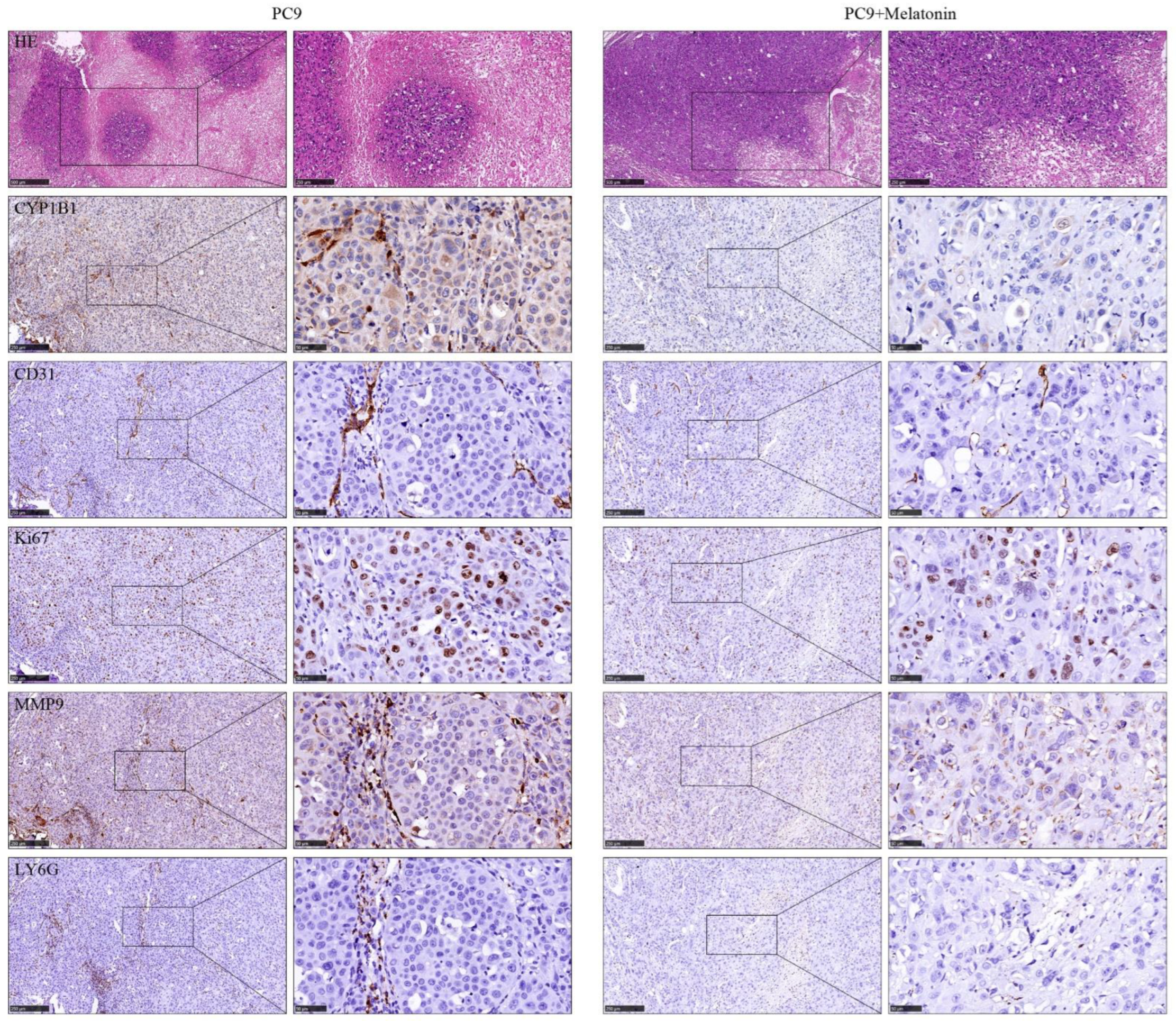
2.12. In Vivo Metastasis Model
2.13. Immunohistochemistry Analysis
2.14. Statistical Analysis
3. Results
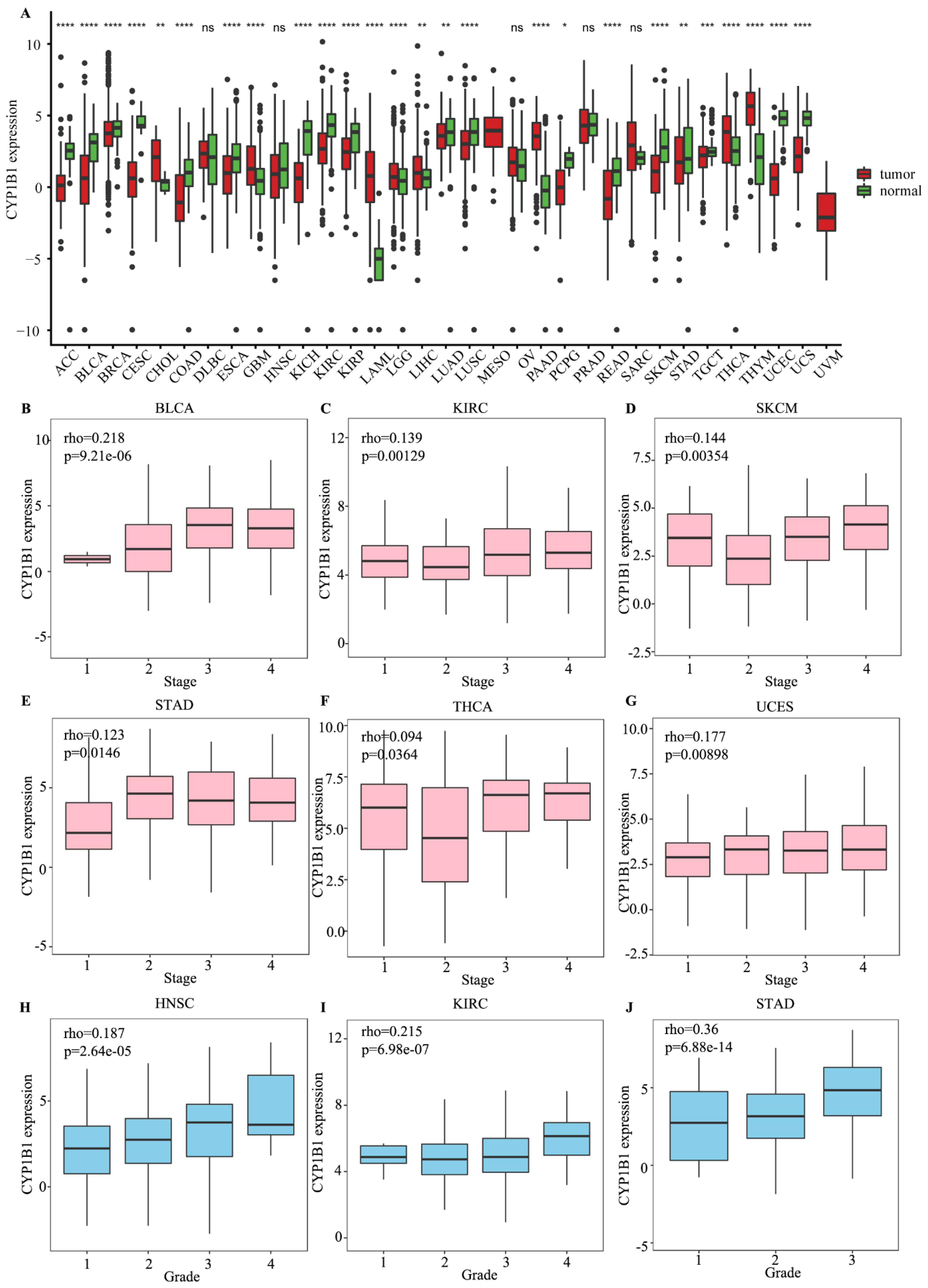
3.1. The Expression, Tumor Stage and Clinical Grade of CYP1B1 in Cancer
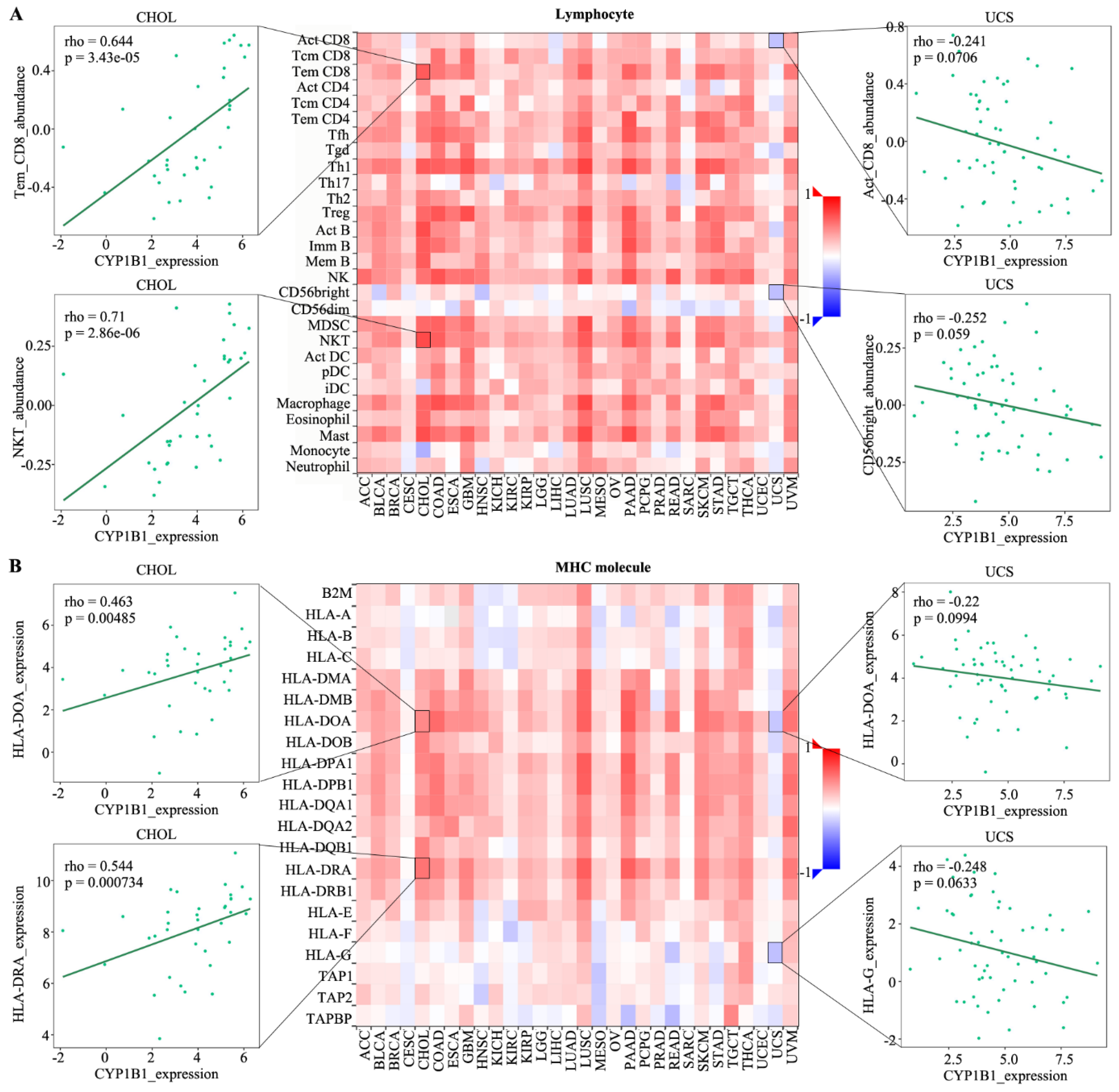
3.2. The Correlation among CYP1B1 Expression and Lymphocyte, Immunomodulator, Chemokine and Receptor
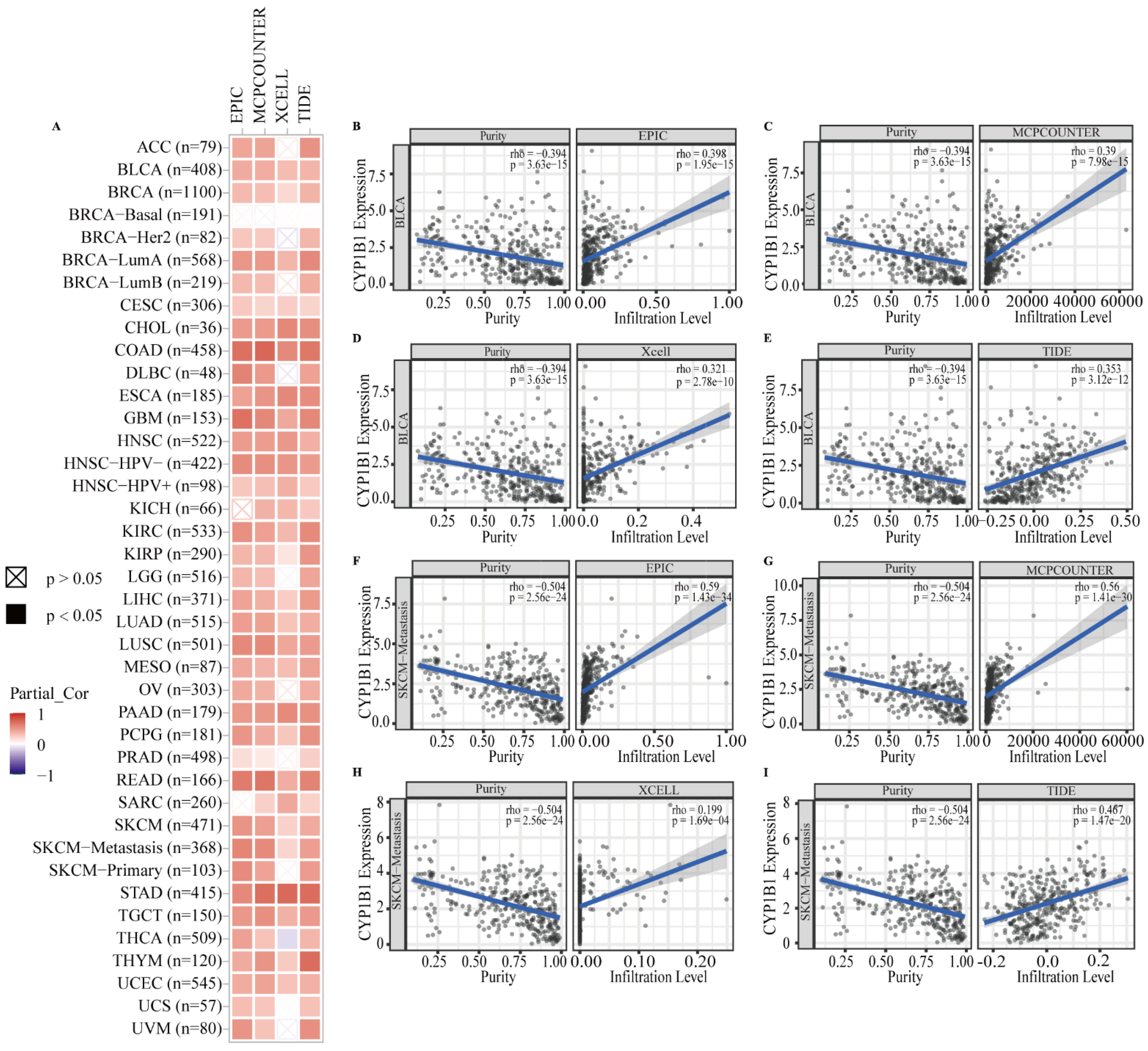
3.3. Correlation between CYP1B1 and CAFs in the Tumor Microenvironment
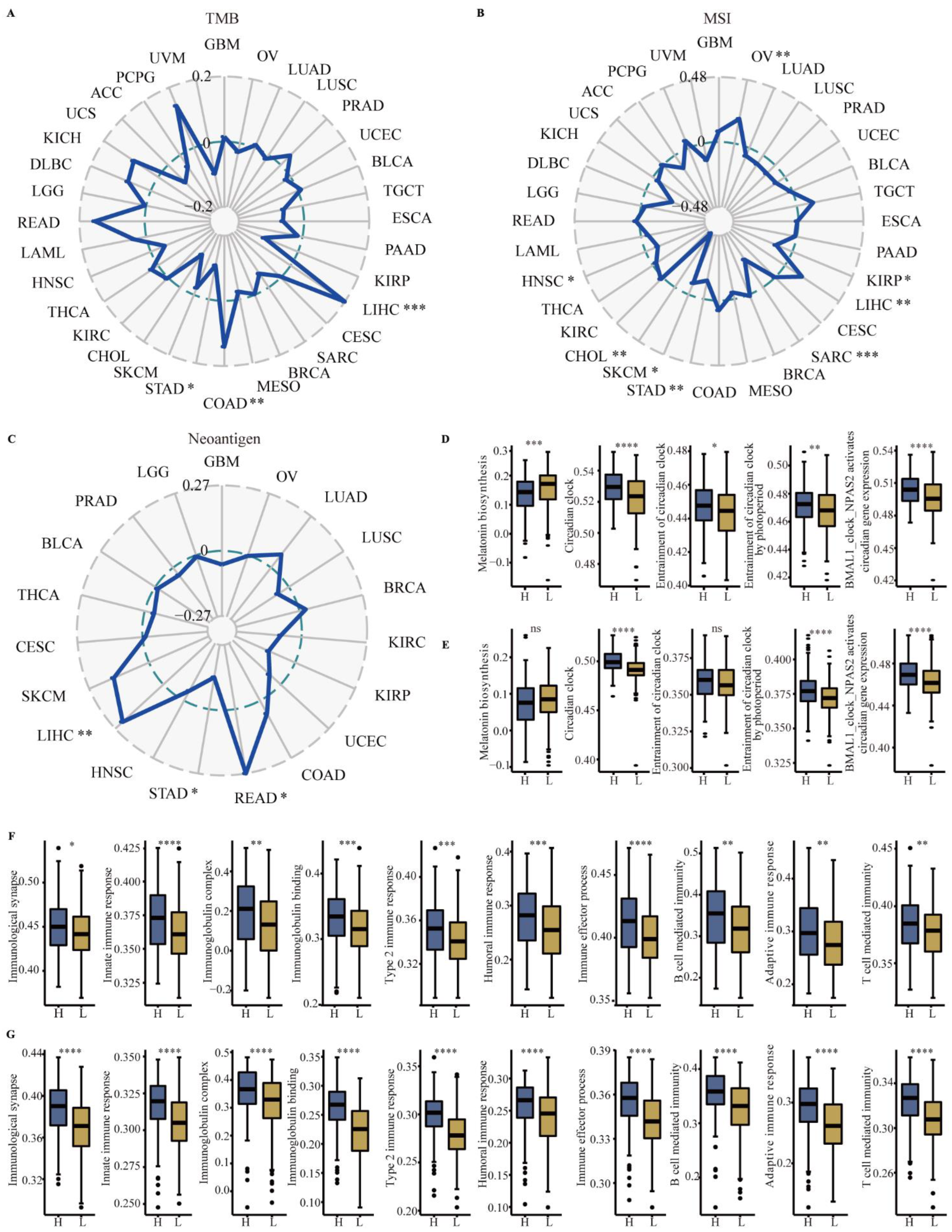
3.4. Correlation between CYP1B1 and TMB, MSI and Neoantigen
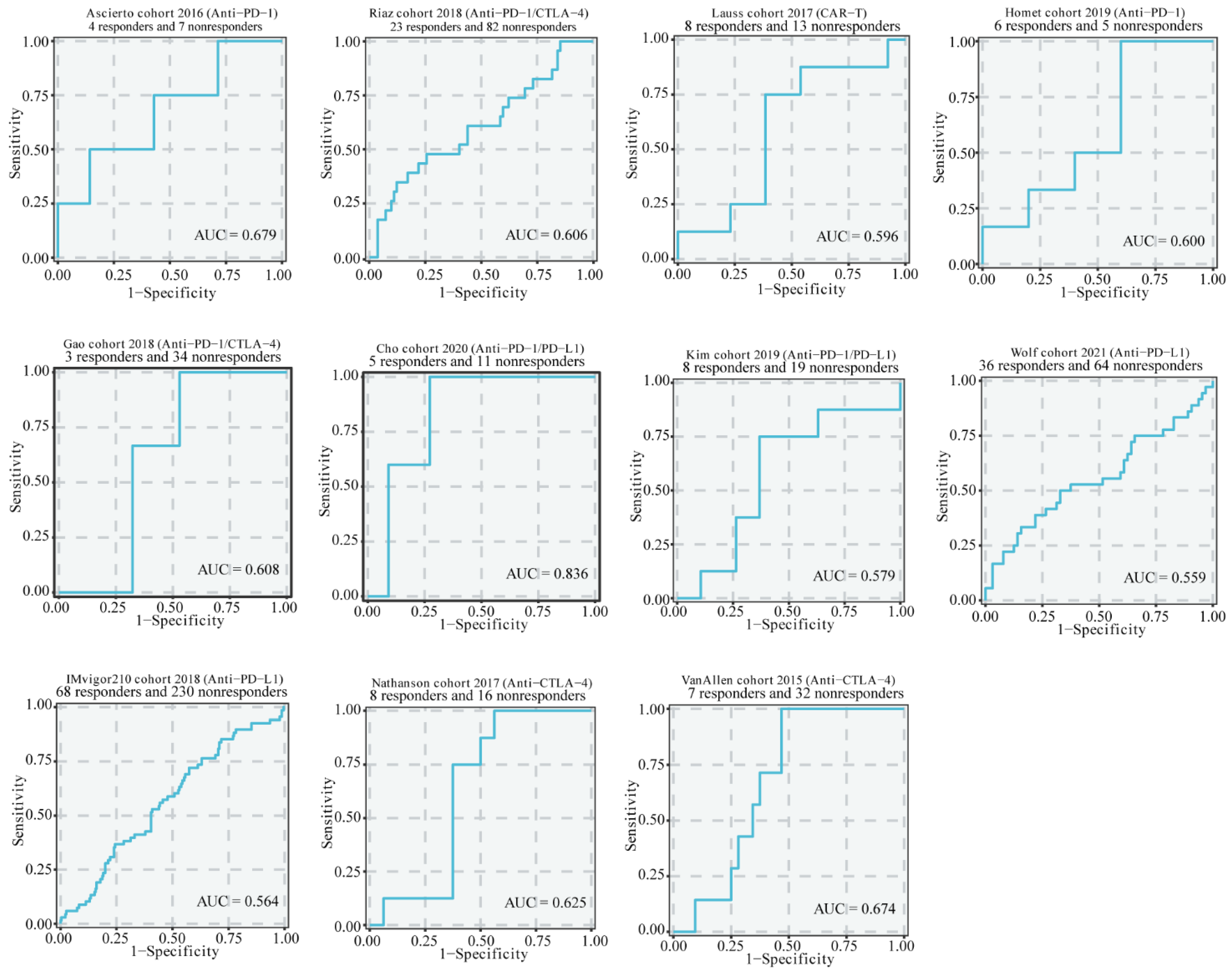
3.5. The Association of CYP1B1 Expression and Therapeutic Response
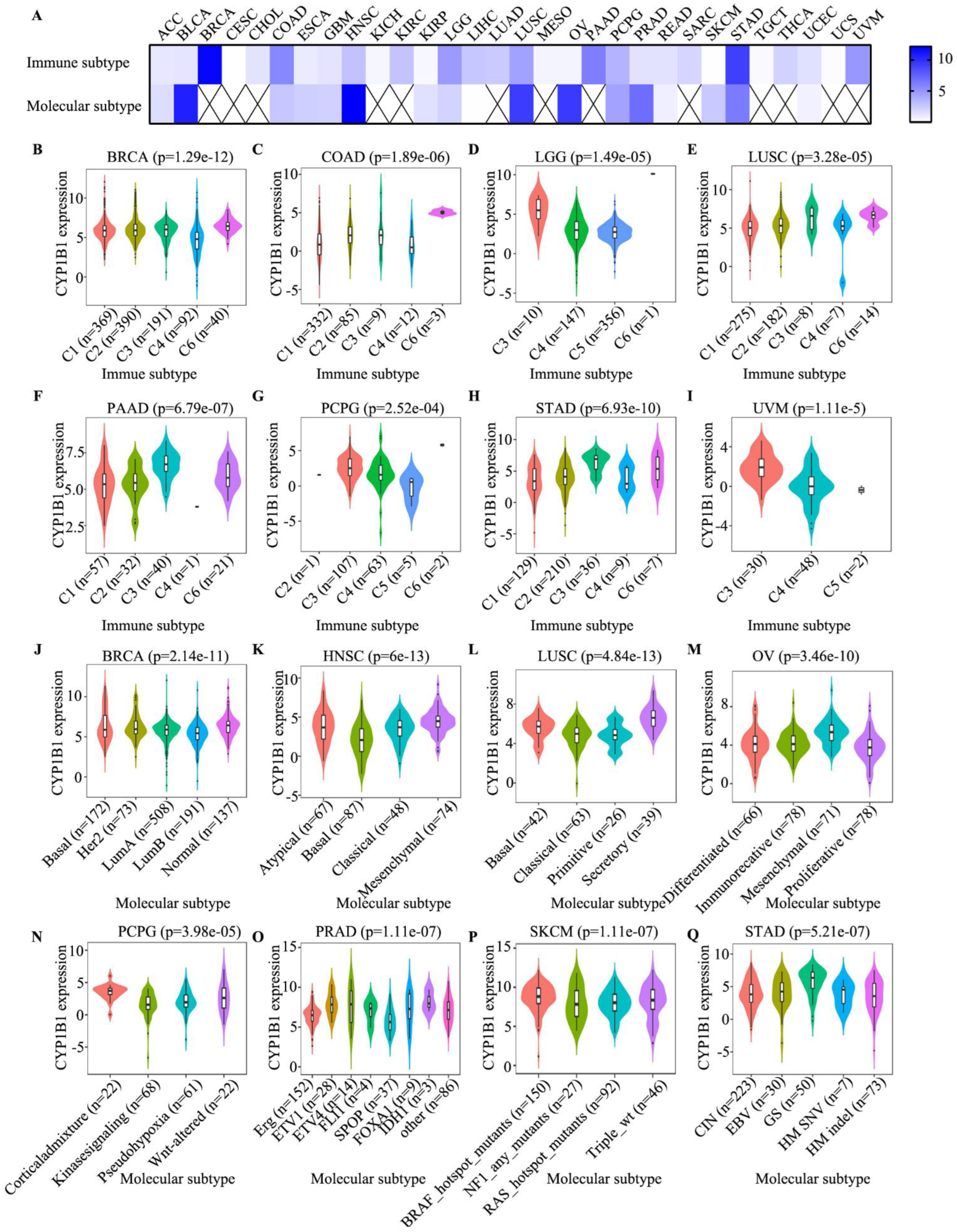
3.6. Identification of Immune Subtype, Molecular Subtype and Immune Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brzezinski, A. Melatonin in humans. N. Engl. J. Med. 1997, 336, 186–195. [Google Scholar] [CrossRef]
- Cagnacci, A. Melatonin in relation to physiology in adult humans. J. Pineal Res. 1996, 21, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Simonneaux, V.; Ribelayga, C. Generation of the melatonin endocrine message in mammals: A review of the complex regulation of melatonin synthesis by norepinephrine, peptides, and other pineal transmitters. Pharmacol. Rev. 2003, 55, 325–395. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Idle, J.R.; Krausz, K.W.; Gonzalez, F.J. Metabolism of melatonin by human cytochromes p450. Drug Metab. Dispos. 2005, 33, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, M.B.; Medvetzky, A.; Weinberger, A.; Driever, W.; Gothilf, Y.; Rath, M.F. Genetic ablation of the Bsx homeodomain transcription factor in zebrafish: Impact on mature pineal gland morphology and circadian behavior. J. Pineal Res. 2022, 72, e12795. [Google Scholar] [CrossRef]
- Kent, B.A.; Rahman, S.A.; St. Hilaire, M.A.; Grant, L.K.; Ruger, M.; Czeisler, C.A.; Lockley, S.W. Circadian lipid and hepatic protein rhythms shift with a phase response curve different than melatonin. Nat. Commun. 2022, 13, 681. [Google Scholar] [CrossRef]
- Hartstein, L.E.; Behn, C.D.; Akacem, L.D.; Stack, N.; Wright, K.P., Jr.; Lebourgeois, M.K. High sensitivity of melatonin suppression response to evening light in preschool-aged children. J. Pineal Res. 2022, 72, e12780. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Choi, W.S. Use of Melatonin in Cancer Treatment: Where Are We? Int. J. Mol. Sci. 2022, 23, 3779. [Google Scholar] [CrossRef]
- Leelaviwat, N.; Mekraksakit, P.; Cross, K.M.; Landis, D.M.; Mclain, M.; Sehgal, L.; Payne, J.D. Melatonin: Translation of Ongoing Studies into Possible Therapeutic Applications Outside Sleep Disorders. Clin. Ther. 2022, 44, 783–812. [Google Scholar] [CrossRef]
- Bantounou, M.; Plascevic, J.; Galley, H.F. Melatonin and Related Compounds: Antioxidant and Anti-Inflammatory Actions. Antioxidants 2022, 11, 532. [Google Scholar] [CrossRef]
- Liu, S.C.; Tsai, C.H.; Wang, Y.H.; Su, C.M.; Wu, H.C.; Fong, Y.C.; Yang, S.F.; Tang, C.H. Melatonin abolished proinflammatory factor expression and antagonized osteoarthritis progression in vivo. Cell Death Dis. 2022, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; He, J.; Liu, C.; Zhang, F.; Zhang, Z.; Chen, H.; Wang, Q.; Ding, X.; Luo, H. Melatonin Attenuates Ropivacaine-Induced Apoptosis by Inhibiting Excessive Mitophagy Through the Parkin/PINK1 Pathway in PC12 and HT22 Cells. Inflammation 2022, 45, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, Y.; Tang, H.; Wang, H.; Jiang, E.; Shao, Z.; Liu, K.; Zhou, X.; Shang, Z. Melatonin Inhibits EMT and PD-L1 Expression through the ERK1/2/FOSL1 Pathway and Regulates Anti-Tumor Immunity in HNSCC. Cancer Sci. 2022, 113, 2232–2245. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Bai, Y.; Wei, Y.; Dong, Y.; Zeng, H.; Reiter, R.J.; Shi, H. Fine-tuning of pathogenesis-related protein 1 (PR1) activity by the melatonin biosynthetic enzyme ASMT2 in defense response to cassava bacterial blight. J. Pineal Res. 2022, 72, e12784. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hao, B.; Zhang, M.; Reiter, R.J.; Lin, S.; Zheng, T.; Chen, X.; Ren, Y.; Yue, L.; Abay, B. Melatonin enhances radiofrequency-induced NK antitumor immunity, causing cancer metabolism reprogramming and inhibition of multiple pulmonary tumor development. Signal Transduct. Target. Ther. 2021, 6, 330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, B.; Li, J.; Mao, X.; He, W.; Chen, Q. Melatonin Inhibits the Progression of Oral Squamous Cell Carcinoma via Inducing miR-25-5p Expression by Directly Targeting NEDD9. Front. Oncol. 2020, 10, 543591. [Google Scholar] [CrossRef]
- Chuffa, L.G.D.A.; Carvalho, R.F.; Justulin, L.A.; Cury, S.S.; Seiva, F.R.F.; Jardim-Perassi, B.V.; de Campos Zuccari, D.A.P.; Reiter, R.J. A meta-analysis of microRNA networks regulated by melatonin in cancer: Portrait of potential candidates for breast cancer treatment. J. Pineal Res. 2020, 69, e12693. [Google Scholar] [CrossRef]
- Fathizadeh, H.; Mirzaei, H.; Asemi, Z. Melatonin: An anti-tumor agent for osteosarcoma. Cancer Cell Int. 2019, 19, 319. [Google Scholar] [CrossRef]
- Ross, J. Comments on the article “Persistent confusion of total entropy and chemical system entropy in chemical thermodynamics” [(1996) Proc. Natl. Acad. Sci. USA 93, 7452–7453]. Proc. Natl. Acad. Sci. USA 1996, 93, 14314; discussion 14315. [Google Scholar] [CrossRef] [Green Version]
- Crewe, H.K.; Notley, L.M.; Wunsch, R.M.; Lennard, M.S.; Gillam, E.M.J. Metabolism of tamoxifen by recombinant human cytochrome P450 enzymes: Formation of the 4-hydroxy, 4′-hydroxy and N-desmethyl metabolites and isomerization of trans-4-hydroxytamoxifen. Drug Metab. Dispos. 2002, 30, 869–874. [Google Scholar] [CrossRef]
- Shou, M.; Korzekwa, K.R.; Krausz, K.W.; Buters, J.T.; Grogan, J.; Goldfarb, I.; Hardwick, J.P.; Gonzalez, F.J.; Gelboin, H.V. Specificity of cDNA-expressed human and rodent cytochrome P450s in the oxidative metabolism of the potent carcinogen 7,12-dimethylbenz[a]anthracene. Mol. Carcinog. 1996, 17, 241–249. [Google Scholar] [CrossRef]
- Shimada, T.; Gillam, E.M.; Sutter, T.R.; Strickland, P.T.; Guengerich, F.P.; Yamazaki, H. Oxidation of xenobiotics by recombinant human cytochrome P450 1B1. Drug Metab. Dispos. 1997, 25, 617–622. [Google Scholar] [PubMed]
- Tanaka, Y.; Sasaki, M.; Kaneuchi, M.; Shiina, H.; Lgawa, M.; Dahiya, R. Polymorphisms of the CYP1B1 gene have higher risk for prostate cancer. Biochem. Biophys. Res. Commun. 2002, 296, 820–826. [Google Scholar] [CrossRef]
- Mcgrath., M.; Hankinson, S.E.; Arbeitman, L.; Colditz, G.A.; Hunter, D.J.; Vivo, I.D. Cytochrome P450 1B1 and catechol-O-methyltransferase polymorphisms and endometrial cancer susceptibility. Carcinogenesis 2004, 25, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Long, B.; Qin, X.; Li, W.; Zhou, Y. Cytochrome P1B1 (CYP1B1) polymorphisms and cancer risk: A meta-analysis of 52 studies. Toxicology 2015, 327, 77–86. [Google Scholar] [CrossRef]
- Mohamed, H.T.; Gadalla, R.; El-husseiny, N.; Hassan, H.; Wang, Z.; Ibrahim, S.A.; EI-Shinawi, M.; Sherr, D.H.; Mohamed, M.M. Inflammatory breast cancer: Activation of the aryl hydrocarbon receptor and its target CYP1B1 correlates closely with Wnt5a/b-beta-catenin signalling, the stem cell phenotype and disease progression. J. Adv. Res. 2019, 16, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.J.; Baek, H.S.; Ye, D.J.; Shin, S.; Kim, D.; Chun, Y.J. CYP1B1 Enhances Cell Proliferation and Metastasis through Induction of EMT and Activation of Wnt/beta-Catenin Signaling via Sp1 Upregulation. PLoS ONE 2016, 11, e0151598. [Google Scholar] [CrossRef] [Green Version]
- D’uva, G.; Baci, D.; Albini, A.; Noonan, D.M. Cancer chemoprevention revisited: Cytochrome P450 family 1B1 as a target in the tumor and the microenvironment. Cancer Treat. Rev. 2018, 63, 1–18. [Google Scholar] [CrossRef]
- Hu, B.; Wu, C.; Mao, H.; Gu, H.; Dong, H.; Yan, J.; Qi, Z.; Yuan, L.; Dong, Q.; Long, J. Subpopulations of cancer-associated fibroblasts link the prognosis and metabolic features of pancreatic ductal adenocarcinoma. Ann. Transl. Med. 2022, 10, 262. [Google Scholar] [CrossRef]
- Saw, P.E.; Chen, J.; Song, E. Targeting CAFs to overcome anticancer therapeutic resistance. Trends Cancer 2022, 8, 527–555. [Google Scholar] [CrossRef]
- Ganguly, K.; Shah, A.; Atri, P.; Rauth, S.; Ponnusamy, M.P.; Kumar, S.; Batra, S.K. Chemokine-mucinome interplay in shaping the heterogeneous tumor microenvironment of pancreatic cancer. Semin. Cancer Biol. 2022, 86, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, H.; Du, K.; Xie, T.; Wang, B.; Chen, C.; Reiter, R.J.; Cen, B.; Yuan, Y. Pan-cancer analyses reveal genomics and clinical characteristics of the melatonergic regulators in cancer. J. Pineal Res. 2021, 71, e12758. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.Y.; Seo, Y.; Oh, S.J.; Ahn, J.S.; Song, M.H.; Kang, M.J.; Oh, J.M.; Lee, D.; Kim, Y.H.; Sung, E.S.; et al. Melatonin and verteporfin synergistically suppress the growth and stemness of head and neck squamous cell carcinoma through the regulation of mitochondrial dynamics. J. Pineal Res. 2022, 72, e12779. [Google Scholar] [CrossRef] [PubMed]
- Bilska, B.; Schedel, F.; Piotrowska, A.; Stefan, J.; Zmijewski, M.; Pyza, E.; Reiter, R.J.; Steinbrink, K.; Slominski, A.T.; Tulic, M.K.; et al. Mitochondrial function is controlled by melatonin and its metabolites in vitro in human melanoma cells. J. Pineal Res. 2021, 70, e12728. [Google Scholar] [CrossRef]
- Kaminsky, L.S.; Spivack, S.D. Cytochromes P450 and cancer. Mol. Aspects Med. 1999, 20, 70–84, 137. [Google Scholar] [PubMed]
- Silvestri, L.; Sonzogni, L.; De Silvestri, A.; Gritti, C.; Foti, L.; Zavaglia, C.; Leveri, M.; Cividini, A.; Mondelli, M.U.; Civardi, E.; et al. CYP enzyme polymorphisms and susceptibility to HCV-related chronic liver disease and liver cancer. Int. J. Cancer 2003, 104, 310–317. [Google Scholar] [CrossRef]
- Murray, G.I.; Taylor, M.C.; Mcfadyen, M.C.; McKay, J.A.; Greenlee, W.F.; Burke, M.D.; Melvin, W.T. Tumor-specific expression of cytochrome P450 CYP1B1. Cancer Res. 1997, 57, 3026–3031. [Google Scholar] [PubMed]
- Luby, T.M.; Cole, G.; Baker, L.; Kornher, J.S.; Ramstedt, U.; Hedley, M.L. Repeated immunization with plasmid DNA formulated in poly(lactide-co-glycolide) microparticles is well tolerated and stimulates durable T cell responses to the tumor-associated antigen cytochrome P450 1B1. Clin. Immunol. 2004, 112, 45–53. [Google Scholar] [CrossRef]
- Mitsui, Y.; Chang, I.; Fukuhara, S.; Hiraki, M.; Arichi, N.; Yasumoto, H.; Hirata, H.; Yamamura, S.; Shahryari, V.; Deng, G.; et al. CYP1B1 promotes tumorigenesis via altered expression of CDC20 and DAPK1 genes in renal cell carcinoma. BMC Cancer 2015, 15, 942. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.W.; Zheng, Z.Q.; Wang, Z.X.; Zhou, G.Q.; Chen, L.; Mao, Y.P.; Lin, A.H.; Reiter, R.J.; Ma, J.; Chen, Y.P.; et al. Pan-cancer genomic analyses reveal prognostic and immunogenic features of the tumor melatonergic microenvironment across 14 solid cancer types. J. Pineal Res. 2019, 66, e12557. [Google Scholar] [CrossRef]
- Maecker, B.; Sherr, D.H.; Vonderheide, R.H.; von Bergwelt-Baildon, M.S.; Hirano, N.; Anderson, K.S.; Xia, Z.; Butler, M.O.; Wucherpfennig, K.W.; O’Hara, C.; et al. The shared tumor-associated antigen cytochrome P450 1B1 is recognized by specific cytotoxic T cells. Blood 2003, 102, 3287–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maecker, B.; von Bergwelt-Baildon, M.S.; SHERR, D.H.; Nadler, L.M.; Schultze, J.L. Identification of a new HLA-A*0201-restricted cryptic epitope from CYP1B1. Int. J. Cancer. 2005, 115, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J. Crown J Biomarkers for Predicting Response to Immunotherapy with Immune Checkpoint Inhibitors in Cancer Patients. Clin. Chem. 2019, 65, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, K.; Sun, C.; Kobayashi, Y.; Kanaseki, T.; Tokita, S.; Komatsu, T.; Maejima, K.; Futami, J.; Nomura, S.; Udaka, K.; et al. Identification of Neoantigens in Two Murine Gastric Cancer Cell Lines Leading to the Neoantigen-Based Immunotherapy. Cancers 2021, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Mu, Y.; Qi, C.; Wang, J.; Xi, G.; Guo, J.; Mi, R.; Zhao, F. CYP1B1 enhances the resistance of epithelial ovarian cancer cells to paclitaxel in vivo and in vitro. Int. J. Mol. Med. 2015, 35, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Wang, S.; Cao, C.; Ye, W.; Wang, M.; Zhou, C.; Chen, W.; Zhang, X.; Zhang, K.; Zhou, W. alpha-naphthoflavone-derived cytochrome P450 (CYP)1B1 degraders specific for sensitizing CYP1B1-mediated drug resistance to prostate cancer DU145: Structure activity relationship. Bioorg. Chem. 2021, 116, 105295. [Google Scholar] [CrossRef]
- Lin, Q.; Cao, J.; Du, X.; Yang, K.; Yang, X.; Liang, Z.; Shi, J.; Zhang, J. CYP1B1-catalyzed 4-OHE2 promotes the castration resistance of prostate cancer stem cells by estrogen receptor alpha-mediated IL6 activation. Cell Commun. Signal. 2022, 20, 31. [Google Scholar] [CrossRef]
- Gribben, J.G.; Ryan, D.P.; Boyajian, R.; Urban, R.G.; Hedley, M.L.; Beach, K.; Nealon, P.; Matulonis, U.; Campos, S.; Gilligan, T.D.; et al. Unexpected association between induction of immunity to the universal tumor antigen CYP1B1 and response to next therapy. Clin. Cancer Res. 2005, 11, 4430–4436. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, B.; Liu, G.; Dai, Z.; Wang, L.; Lin, B.; Zhang, J. CYP1B1: A Novel Molecular Biomarker Predicts Molecular Subtype, Tumor Microenvironment, and Immune Response in 33 Cancers. Cancers 2022, 14, 5641. https://doi.org/10.3390/cancers14225641
Yuan B, Liu G, Dai Z, Wang L, Lin B, Zhang J. CYP1B1: A Novel Molecular Biomarker Predicts Molecular Subtype, Tumor Microenvironment, and Immune Response in 33 Cancers. Cancers. 2022; 14(22):5641. https://doi.org/10.3390/cancers14225641
Chicago/Turabian StyleYuan, Benchao, Guihong Liu, Zili Dai, Li Wang, Baisheng Lin, and Jian Zhang. 2022. "CYP1B1: A Novel Molecular Biomarker Predicts Molecular Subtype, Tumor Microenvironment, and Immune Response in 33 Cancers" Cancers 14, no. 22: 5641. https://doi.org/10.3390/cancers14225641