
Comparison of Selected Non-Coding RNAs and Gene Expression Profiles between Common Osteosarcoma Cell Lines
 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
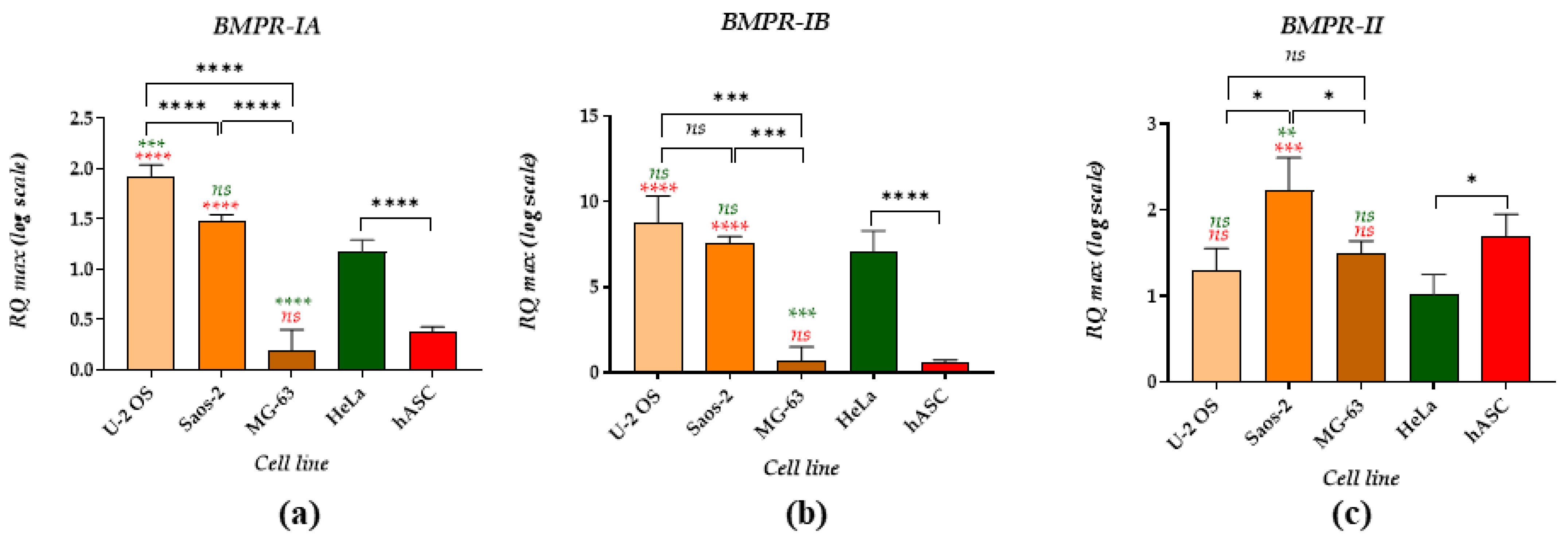
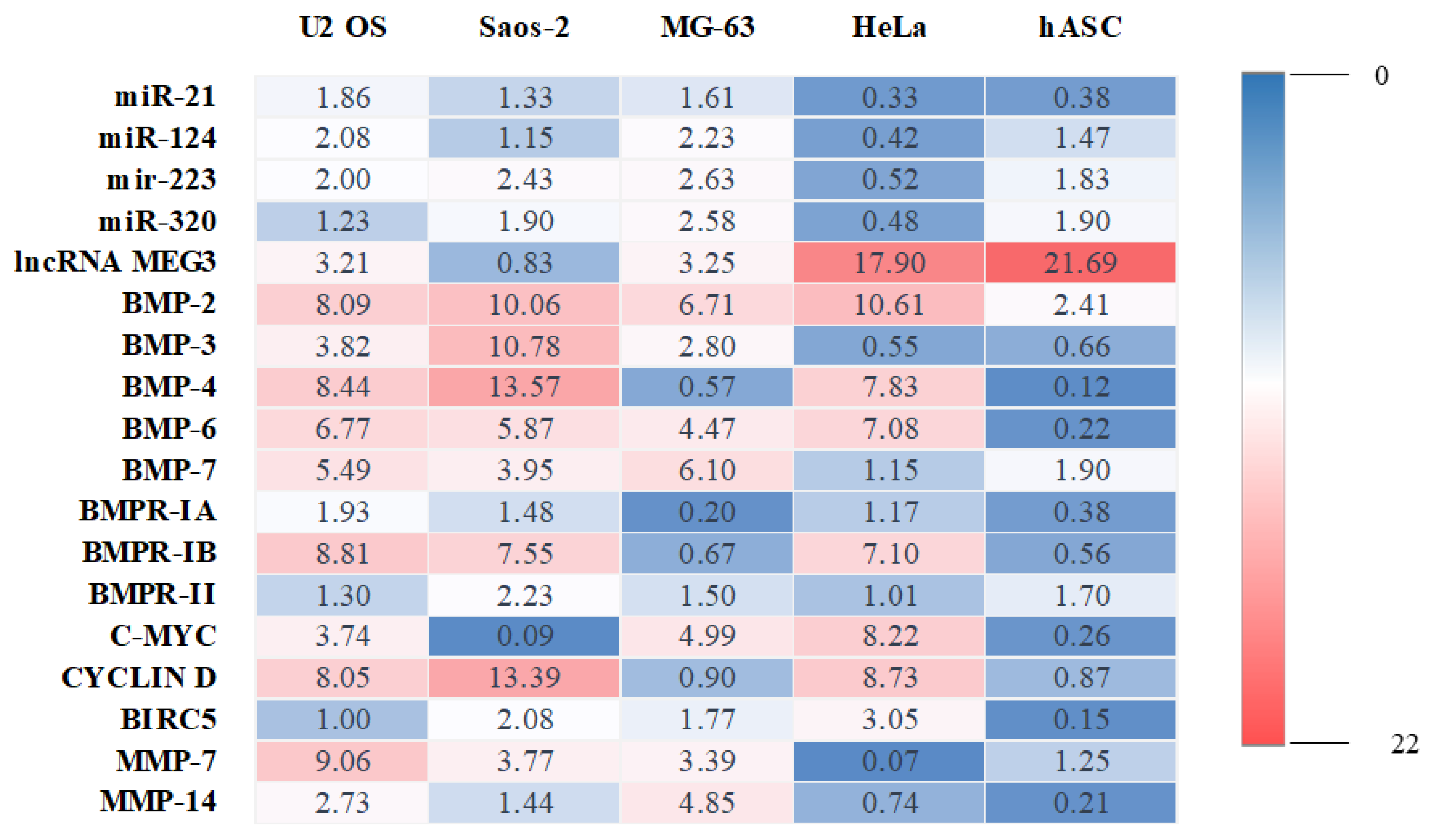
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Quantitative Reverse Transcription PCR (RT-qPCR)
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational Biology of Osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.; Dunning, M.D.; de Brot, S.; Grau-Roma, L.; Mongan, N.P.; Rutland, C.S. Comparative Review of Human and Canine Osteosarcoma: Morphology, Epidemiology, Prognosis, Treatment and Genetics. Acta Vet. Scand. 2017, 59, 71. [Google Scholar] [CrossRef] [PubMed]
- Pautke, C.; Schieker, M.; Tischer, T.; Kolk, A.; Neth, P.; Mutschler, W.; Milz, S. Characterization of Osteosarcoma Cell Lines MG-63, Saos-2 and U-2 OS in Comparison to Human Osteoblasts. Anticancer. Res. 2004, 24, 3743–3748. [Google Scholar]
- Fromigué, O.; Hamidouche, Z.; Marie, P.J. Blockade of the RhoA-JNK-c-Jun-MMP2 Cascade by Atorvastatin Reduces Osteosarcoma Cell Invasion. J. Biol. Chem. 2008, 283, 30549–30556. [Google Scholar] [CrossRef] [PubMed]
- Choong, P.F.; Teh, H.X.; Teoh, H.K.; Ong, H.K.; Choo, K.B.; Sugii, S.; Cheong, S.K.; Kamarul, T. Heterogeneity of Osteosarcoma Cell Lines Led to Variable Responses in Reprogramming. Int. J. Med. Sci. 2014, 11, 1154–1160. [Google Scholar] [CrossRef]
- Włodarski, K.H.; Kochanowska, I.; Pieńikowski, M.; Ostrowski, K. Influence of Osteoprotegerin (OPG) on Experimentally Induced Ectopic Bone. Folia Biol. 2004, 52, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Kochanowska, I.E.; Wlodarski, K.; Wojtowicz, A.; Niemira, K.; Ostrowski, K. Osteogenic Properties of Various HeLa Cell Lines and the BMP Family Genes Expression. Ann. Transplant. 2002, 7, 58–62. [Google Scholar]
- Wang, Y.; Chu, Y.; Yue, B.; Ma, X.; Zhang, G.; Xiang, H.; Liu, Y.; Wang, T.; Wu, X.; Chen, B. Adipose-Derived Mesenchymal Stem Cells Promote Osteosarcoma Proliferation and Metastasis by Activating the STAT3 Pathway. Oncotarget 2017, 8, 23803–23816. [Google Scholar] [CrossRef]
- Wang, Y.; Chu, Y.; Li, K.; Zhang, G.; Guo, Z.; Wu, X.; Qiu, C.; Li, Y.; Wan, X.; Sui, J.; et al. Exosomes Secreted by Adipose-Derived Mesenchymal Stem Cells Foster Metastasis and Osteosarcoma Proliferation by Increasing COLGALT2 Expression. Front. Cell Dev. Biol. 2020, 8, 353. [Google Scholar] [CrossRef]
- Hu, X.; Li, L.; Lu, Y.; Yu, X.; Chen, H.; Yin, Q.; Zhang, Y. MiRNA-21 Inhibition Inhibits Osteosarcoma Cell Proliferation by Targeting PTEN and Regulating the TGF-Β1 Signaling Pathway. Oncol. Lett. 2018, 16, 4337–4342. [Google Scholar] [CrossRef]
- Sheng, S.; Su, W.; Mao, D.; Li, C.; Hu, X.; Deng, W.; Yao, Y.; Ji, Y. MicroRNA-21 Induces Cisplatin Resistance in Head and Neck Squamous Cell Carcinoma. PLoS ONE 2022, 17, e0267017. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liang, Y.; Xu, M.; Xiong, J.; Wang, D.; Ding, Q. MicroRNA-124 Acts as a Tumor-Suppressive MiRNA by Inhibiting the Expression of Snail2 in Osteosarcoma. Oncol. Lett. 2018, 15, 4979–4987. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Xu, X.; Song, Q.; Xu, Y.; Tai, Y.; Goodman, S.B.; Bi, W.; Xu, M.; Jiao, S.; Maloney, W.J.; et al. MiR-223-3p Inhibits Human Osteosarcoma Metastasis and Progression by Directly Targeting CDH6. Mol. Ther. 2018, 26, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, J.; Chen, P.; Song, Y.; An, W.; Zhang, H.; Butegeleqi, B.; Yan, J. MicroRNA-320a Inhibits Invasion and Metastasis in Osteosarcoma by Targeting Cytoplasmic Polyadenylation Element-binding Protein 1. Cancer Med. 2020, 9, 2833–2845. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Liu, X.; Lv, B. Long Non-coding RNA MEG3 Promotes Autophagy and Apoptosis of Nasopharyngeal Carcinoma Cells via PTEN Up-regulation by Binding to MicroRNA-21. J. Cell Mol. Med. 2021, 25, 61–72. [Google Scholar] [CrossRef]
- Shi, Y.; Lv, C.; Shi, L.; Tu, G. MEG3 Inhibits Proliferation and Invasion and Promotes Apoptosis of Human Osteosarcoma Cells. Oncol. Lett. 2018, 15, 1917–1923. [Google Scholar] [CrossRef]
- Mukhametov, U.; Lyulin, S.; Borzunov, D.; Sufianova, G.; Shumadalova, A.; Zhang, D.; Gareev, I. Functions of the Bone Morphogenetic Protein Signaling Pathway through Non-Coding RNAs. Noncoding RNA Res. 2022, 7, 178–183. [Google Scholar] [CrossRef]
- Carreira, A.C.; Lojudice, F.H.; Halcsik, E.; Navarro, R.D.; Sogayar, M.C.; Granjeiro, J.M. Bone Morphogenetic Proteins: Facts, Challenges, and Future Perspectives. J. Dent. Res. 2014, 93, 335–345. [Google Scholar] [CrossRef]
- Katagiri, T.; Watabe, T. Bone Morphogenetic Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) Signaling in Development and Human Diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef]
- Li, H.; Yang, F.; Wang, Z.; Fu, Q.; Liang, A. MicroRNA-21 Promotes Osteogenic Differentiation by Targeting Small Mothers against Decapentaplegic 7. Mol. Med. Rep. 2015, 12, 1561–1567. [Google Scholar] [CrossRef] [PubMed]
- Bjørnland, K.; Flatmark, K.; Pettersen, S.; Aaasen, A.O.; Fodstad, O.; Maelandsmo, G.M. Matrix Metalloproteinases Participate in Osteosarcoma Invasion. J. Surg. Res. 2005, 127, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Kaczorowska, A.; Miękus, N.; Stefanowicz, J.; Adamkiewicz-Drożyńska, E. Selected Matrix Metalloproteinases (MMP-2, MMP-7) and Their Inhibitor (TIMP-2) in Adult and Pediatric Cancer. Diagnostics 2020, 10, 547. [Google Scholar] [CrossRef]
- Liu, Y.; Teng, Z.; Wang, Y.; Gao, P.; Chen, J. Prognostic Significance of Survivin Expression in Osteosarcoma Patients: A Meta-Analysis. Med. Sci. Monit. 2015, 21, 2877–2885. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-N.; Su, X.-X.; Cheng, S.-Q.; Sun, Z.-Y.; Huang, Z.-S.; Ou, T.-M. MYC Modulators in Cancer: A Patent Review. Expert Opin. Ther. Pat. 2019, 29, 353–367. [Google Scholar] [CrossRef]
- Tian, H.; Zhou, T.; Chen, H.; Li, C.; Jiang, Z.; Lao, L.; Kahn, S.A.; Duarte, M.E.L.; Zhao, J.; Daubs, M.D.; et al. Bone Morphogenetic Protein-2 Promotes Osteosarcoma Growth by Promoting Epithelial-mesenchymal Transition (EMT) through the Wnt/Β-catenin Signaling Pathway. J. Orthop. Res. 2019, 37, 1638–1648. [Google Scholar] [CrossRef]
- Cai, C.-K.; Zhao, G.-Y.; Tian, L.-Y.; Liu, L.; Yan, K.; Ma, Y.-L.; Ji, Z.-W.; Li, X.-X.; Han, K.; Gao, J.; et al. MiR-15a and MiR-16-1 Downregulate CCND1 and Induce Apoptosis and Cell Cycle Arrest in Osteosarcoma. Oncol. Rep. 2012, 28, 1764–1770. [Google Scholar] [CrossRef]
- He, H.; Ni, J.; Huang, J. Molecular Mechanisms of Chemoresistance in Osteosarcoma (Review). Oncol. Lett. 2014, 7, 1352–1362. [Google Scholar] [CrossRef]
- Li, J.; Yang, Z.; Li, Y.; Xia, J.; Li, D.; Li, H.; Ren, M.; Liao, Y.; Yu, S.; Chen, Y.; et al. Cell Apoptosis, Autophagy and Necroptosis in Osteosarcoma Treatment. Oncotarget 2016, 7, 44763–44778. [Google Scholar] [CrossRef]
- Lauvrak, S.U.; Munthe, E.; Kresse, S.H.; Stratford, E.W.; Namløs, H.M.; Meza-Zepeda, L.A.; Myklebost, O. Functional Characterisation of Osteosarcoma Cell Lines and Identification of MRNAs and MiRNAs Associated with Aggressive Cancer Phenotypes. Br. J. Cancer 2013, 109, 2228–2236. [Google Scholar] [CrossRef]
- Namløs, H.M.; Meza-Zepeda, L.A.; Barøy, T.; Østensen, I.H.G.; Kresse, S.H.; Kuijjer, M.L.; Serra, M.; Bürger, H.; Cleton-Jansen, A.-M.; Myklebost, O. Modulation of the Osteosarcoma Expression Phenotype by MicroRNAs. PLoS ONE 2012, 7, e48086. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, G.; Chen, R.; Hua, Y.; Cai, Z. Mesenchymal Stem Cells in the Osteosarcoma Microenvironment: Their Biological Properties, Influence on Tumor Growth, and Therapeutic Implications. Stem Cell Res. Ther. 2018, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, B.; Bastos, A.R.F.; Brancato, V.; Cerqueira, M.T.; Oliveira, J.M.; Correlo, V.M.; Reis, R.L.; Kundu, S.C. Mechanical Property of Hydrogels and the Presence of Adipose Stem Cells in Tumor Stroma Affect Spheroid Formation in the 3D Osteosarcoma Model. ACS Appl. Mater. Interfaces 2019, 11, 14548–14559. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.; Marcinkowska, K.; Marycz, K.; Wiglusz, R.J.; Śmieszek, A. The Potential Selective Cytotoxicity of Poly (L- Lactic Acid)-Based Scaffolds Functionalized with Nanohydroxyapatite and Europium (III) Ions toward Osteosarcoma Cells. Materials 2019, 12, 3779. [Google Scholar] [CrossRef]
- Punzo, F.; Tortora, C.; Argenziano, M.; Pinto, D.D.; Pota, E.; Martino, M.D.; Paola, A.D.; Rossi, F. Can Denosumab Be Used in Combination with Doxorubicin in Osteosarcoma? Oncotarget 2020, 11, 2763–2773. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, X.; Wang, H. Punicalagin Inhibited Proliferation, Invasion and Angiogenesis of Osteosarcoma through Suppression of NF-κB Signaling. Mol. Med. Rep. 2020, 22, 2386–2394. [Google Scholar] [CrossRef]
- Vanas, V.; Haigl, B.; Stockhammer, V.; Sutterlüty-Fall, H. MicroRNA-21 Increases Proliferation and Cisplatin Sensitivity of Osteosarcoma-Derived Cells. PLoS ONE 2016, 11, e0161023. [Google Scholar] [CrossRef]
- Nakka, M.; Allen-Rhoades, W.; Li, Y.; Kelly, A.J.; Shen, J.; Taylor, A.M.; Barkauskas, D.A.; Yustein, J.T.; Andrulis, I.L.; Wunder, J.S.; et al. Biomarker Significance of Plasma and Tumor MiR-21, MiR-221, and MiR-106a in Osteosarcoma. Oncotarget 2017, 8, 96738–96752. [Google Scholar] [CrossRef]
- Sekar, D.; Mani, P.; Biruntha, M.; Sivagurunathan, P.; Karthigeyan, M. Dissecting the Functional Role of MicroRNA 21 in Osteosarcoma. Cancer Gene Ther. 2019, 26, 179–182. [Google Scholar] [CrossRef]
- Lv, C.; Hao, Y.; Tu, G. MicroRNA-21 Promotes Proliferation, Invasion and Suppresses Apoptosis in Human Osteosarcoma Line MG63 through PTEN/Akt Pathway. Tumor Biol. 2016, 37, 9333–9342. [Google Scholar] [CrossRef]
- Hua, Y.; Jin, Z.; Zhou, F.; Zhang, Y.-Q.; Zhuang, Y. The Expression Significance of Serum MiR-21 in Patients with Osteosarcoma and Its Relationship with Chemosensitivity. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2989–2994. [Google Scholar] [PubMed]
- Fehr, J.E.; Trotter, G.W.; Oxford, J.T.; Hart, D.A. Comparison of Northern Blot Hybridization and a Reverse Transcriptase-Polymerase Chain Reaction Technique for Measurement of MRNA Expression of Metalloproteinases and Matrix Components in Articular Cartilage and Synovial Membrane from Horses with Osteoarthritis. Am. J. Vet. Res. 2000, 61, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wu, W.; Jing, D.; Yang, L.; Guo, H.; Wang, L.; Zhang, W.; Pu, F.; Shao, Z. Engineered Exosome as Targeted LncRNA MEG3 Delivery Vehicles for Osteosarcoma Therapy. J. Control. Release 2022, 343, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.-Z.; Guo, X.-J.; Zhao, Y.-M.; Fang, Y. Decreased Expression of Long Non-Coding RNA MEG3 Acts as a Potential Predictor Biomarker in Progression and Poor Prognosis of Osteosarcoma. Int. J. Clin. Exp. Pathol. 2015, 8, 15138–15142. [Google Scholar]
- Smieszek, A.; Marcinkowska, K.; Pielok, A.; Sikora, M.; Valihrach, L.; Marycz, K. The Role of MiR-21 in Osteoblasts–Osteoclasts Coupling In Vitro. Cells 2020, 9, 479. [Google Scholar] [CrossRef]
- Seweryn, A.; Pielok, A.; Lawniczak-Jablonska, K.; Pietruszka, R.; Marcinkowska, K.; Sikora, M.; Witkowski, B.S.; Godlewski, M.; Marycz, K.; Smieszek, A. Zirconium Oxide Thin Films Obtained by Atomic Layer Deposition Technology Abolish the Anti-Osteogenic Effect Resulting from MiR-21 Inhibition in the Pre-Osteoblastic MC3T3 Cell Line. Int. J. Nanomed. 2020, 15, 1595–1610. [Google Scholar] [CrossRef]
- Son, Y.H.; Ka, S.; Kim, A.Y.; Kim, J.B. Regulation of Adipocyte Differentiation via MicroRNAs. Endocrinol. Metab. 2014, 29, 122–135. [Google Scholar] [CrossRef]
- Tang, J.; Lin, X.; Zhong, J.; Xu, F.; Wu, F.; Liao, X.; Cui, R.-R.; Li, F.; Yuan, L. MiR-124 Regulates the Osteogenic Differentiation of Bone Marrow-derived Mesenchymal Stem Cells by Targeting Sp7. Mol. Med. Rep. 2019, 19, 3807–3814. [Google Scholar] [CrossRef]
- Wang, L.; Kang, F.-B.; Sun, N.; Wang, J.; Chen, W.; Li, D.; Shan, B.-E. The Tumor Suppressor MiR-124 Inhibits Cell Proliferation and Invasion by Targeting B7-H3 in Osteosarcoma. Tumor Biol. 2016, 37, 14939–14947. [Google Scholar] [CrossRef]
- Cong, C.; Wang, W.; Tian, J.; Gao, T.; Zheng, W.; Zhou, C. Identification of Serum MiR-124 as a Biomarker for Diagnosis and Prognosis in Osteosarcoma. Cancer Biomark 2018, 21, 449–454. [Google Scholar] [CrossRef]
- Geng, S.; Zhang, X.; Chen, J.; Liu, X.; Zhang, H.; Xu, X.; Ma, Y.; Li, B.; Zhang, Y.; Bi, Z.; et al. The Tumor Suppressor Role of MiR-124 in Osteosarcoma. PLoS ONE 2014, 9, e91566. [Google Scholar] [CrossRef] [PubMed]
- Lucero, C.M.J.; Vega, O.A.; Osorio, M.M.; Tapia, J.C.; Antonelli, M.; Stein, G.S.; Van Wijnen, A.J.; Galindo, M.A. The Cancer-Related Transcription Factor Runx2 Modulates Cell Proliferation in Human Osteosarcoma Cell Lines. J. Cell Physiol. 2013, 228, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cai, D.; Meng, L.; Wang, B. MicroRNA-124 Inhibits Proliferation, Invasion, Migration and Epithelial-Mesenchymal Transition of Cervical Carcinoma Cells by Targeting Astrocyte-Elevated Gene-1. Oncol. Rep. 2016, 36, 2321–2328. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Liu, Y.; Liao, W.; Liu, R.; Shi, P.; Wang, L. MiRNA-223 Is a Potential Diagnostic and Prognostic Marker for Osteosarcoma. J. Bone Oncol. 2016, 5, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Gao, Y.; Zhou, J.; Wang, J.; Zheng, F.; Guo, F.; Chang, A.; Li, X.; Wang, B. MiR-223 Regulates Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells Through a C/EBPs/MiR-223/FGFR2 Regulatory Feedback Loop. STEM CELLS 2015, 33, 1589–1600. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, Y.; Chen, Q.; Qiu, N.; Zhao, Y.; You, X. MiR-223 Inhibited Cell Metastasis of Human Cervical Cancer by Modulating Epithelial-Mesenchymal Transition. Int. J. Clin. Exp. Pathol. 2015, 8, 11224–11229. [Google Scholar]
- Luo, L.; Yang, R.; Zhao, S.; Chen, Y.; Hong, S.; Wang, K.; Wang, T.; Cheng, J.; Zhang, T.; Chen, D. Decreased MiR-320 Expression Is Associated with Breast Cancer Progression, Cell Migration, and Invasiveness via Targeting Aquaporin 1. Acta Biochim. Biophys. Sin. 2018, 50, 473–480. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Z. MicroRNA-320 Suppresses Cervical Cancer Cell Viability, Migration and Invasion via Directly Targeting FOXM1. Oncol. Lett. 2017, 14, 3809–3816. [Google Scholar] [CrossRef]
- Pan, C.; Gao, H.; Zheng, N.; Gao, Q.; Si, Y.; Zhao, Y. MiR-320 Inhibits the Growth of Glioma Cells through Downregulating PBX3. Biol. Res. 2017, 50, 31. [Google Scholar] [CrossRef]
- Wu, H.; Li, W.; Zhang, M.; Zhu, S.; Zhang, D.; Wang, X. Inhibitory Roles of MiR-320 in Osteosarcoma via Regulating E2F1. J. Cancer Res. Ther. 2016, 12, 68–71. [Google Scholar] [CrossRef]
- Cheng, C.; Chen, Z.-Q.; Shi, X.-T. MicroRNA-320 Inhibits Osteosarcoma Cells Proliferation by Directly Targeting Fatty Acid Synthase. Tumor Biol. 2014, 35, 4177–4183. [Google Scholar] [CrossRef] [PubMed]
- Laxman, N.; Mallmin, H.; Nilsson, O.; Kindmark, A. MiR-203 and MiR-320 Regulate Bone Morphogenetic Protein-2-Induced Osteoblast Differentiation by Targeting Distal-Less Homeobox 5 (Dlx5). Genes 2016, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Hamam, D.; Ali, D.; Vishnubalaji, R.; Hamam, R.; Al-Nbaheen, M.; Chen, L.; Kassem, M.; Aldahmash, A.; Alajez, N.M. MicroRNA-320/RUNX2 Axis Regulates Adipocytic Differentiation of Human Mesenchymal (Skeletal) Stem Cells. Cell Death Dis. 2014, 5, e1499. [Google Scholar] [CrossRef]
- Sulzbacher, I.; Birner, P.; Trieb, K.; Pichlbauer, E.; Lang, S. The Expression of Bone Morphogenetic Proteins in Osteosarcoma and Its Relevance as a Prognostic Parameter. J. Clin. Pathol. 2002, 55, 381–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haudenschild, D.R.; Palmer, S.M.; Moseley, T.A.; You, Z.; Reddi, A.H. Bone Morphogenetic Protein (BMP)-6 Signaling and BMP Antagonist Noggin in Prostate Cancer. Cancer Res. 2004, 64, 8276–8284. [Google Scholar] [CrossRef] [PubMed]
- Kochanowska, I.E.; Wlodarski, K.; Wojtowicz, A.; Kinsner, A.; Ostrowski, K. BMP-4 and BMP-6 Involvement in the Osteogenic Properties of the HeLa Cell Line. Exp. Biol. Med. 2002, 227, 57–62. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, J.; Zhou, Y.; Wu, G. The Roles of Bone Morphogenetic Proteins and Their Signaling in the Osteogenesis of Adipose-Derived Stem Cells. Tissue Eng. Part. B Rev. 2014, 20, 84–92. [Google Scholar] [CrossRef]
- Yadin, D.; Knaus, P.; Mueller, T.D. Structural Insights into BMP Receptors: Specificity, Activation and Inhibition. Cytokine Growth Factor Rev. 2016, 27, 13–34. [Google Scholar] [CrossRef]
- Guo, W.; Gorlick, R.; Ladanyi, M.; Meyers, P.A.; Huvos, A.G.; Bertino, J.R.; Healey, J.H. Expression of Bone Morphogenetic Proteins and Receptors in Sarcomas. Clin. Orthop. Relat. Res. 1999, 175–183. [Google Scholar] [CrossRef]
- Schleinitz, D.; Klöting, N.; Böttcher, Y.; Wolf, S.; Dietrich, K.; Tönjes, A.; Breitfeld, J.; Enigk, B.; Halbritter, J.; Körner, A.; et al. Genetic and Evolutionary Analyses of the Human Bone Morphogenetic Protein Receptor 2 (BMPR2) in the Pathophysiology of Obesity. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Bach, D.-H.; Park, H.J.; Lee, S.K. The Dual Role of Bone Morphogenetic Proteins in Cancer. Mol. Ther. Oncolytics 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Badal, S.S.; Wang, Y.; Chang, B.H.J.; Rodriguez, A.; Danesh, F.R. MicroRNA-22 Is a Master Regulator of Bone Morphogenetic Protein-7/6 Homeostasis in the Kidney. J. Biol. Chem. 2013, 288, 36202–36214. [Google Scholar] [CrossRef]
- Davis, B.N.; Hilyard, A.C.; Lagna, G.; Hata, A. SMAD Proteins Control DROSHA-Mediated MicroRNA Maturation. Nature 2008, 454, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lim, M.; Pham, L.K.; Kendall, A.H.; Altieri, D.C.; Roy-Burman, P. Bone Morphogenetic Protein 7 Protects Prostate Cancer Cells from Stress-Induced Apoptosis via Both Smad and c-Jun NH2-Terminal Kinase Pathways. Available online: https://pubmed.ncbi.nlm.nih.gov/16618753/ (accessed on 16 September 2020).
- Chan, J.A.; Krichevsky, A.M.; Kosik, K.S. MicroRNA-21 Is an Antiapoptotic Factor in Human Glioblastoma Cells. Cancer Res. 2005, 65, 6029–6033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappellen, D.; Schlange, T.; Bauer, M.; Maurer, F.; Hynes, N.E. Novel C-MYC Target Genes Mediate Differential Effects on Cell Proliferation and Migration. EMBO Rep. 2007, 8, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Wang, Y.; Bi, W. C-Myc Overexpression Promotes Osteosarcoma Cell Invasion via Activation of MEK-ERK Pathway. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2012, 20, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Li, Z.; Yu, Z.; Yan, F.; Liu, Y.; Lu, X.; Yang, W. MicroRNA-33b Suppresses Migration and Invasion by Targeting c-Myc in Osteosarcoma Cells. PLoS ONE 2014, 9, e115300. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, G.; Wang, X.; Liu, Y. A Furin Inhibitor Downregulates Osteosarcoma Cell Migration by Downregulating the Expression Levels of MT1-MMP via the Wnt Signaling Pathway. Oncol. Lett. 2014, 7, 1033–1038. [Google Scholar] [CrossRef]
- Huang, Y.-Z.; Zhang, J.; Shao, H.-Y.; Chen, J.-P.; Zhao, H.-Y. MicroRNA-191 Promotes Osteosarcoma Cells Proliferation by Targeting Checkpoint Kinase 2. Tumor Biol. 2015, 36, 6095–6101. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, W.; Chen, B.; Bai, P.-D.; Wang, X.-G.; Xing, J.-C. Up-Regulation of MiR-124 Inhibits Invasion and Proliferation of Prostate Cancer Cells through Mediating JAK-STAT3 Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2338–2345. [Google Scholar]
- Melnik, S.; Werth, N.; Boeuf, S.; Hahn, E.-M.; Gotterbarm, T.; Anton, M.; Richter, W. Impact of C-MYC Expression on Proliferation, Differentiation, and Risk of Neoplastic Transformation of Human Mesenchymal Stromal Cells. Stem Cell Res. Ther. 2019, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Paula, A.C.; Martins, T.M.; Zonari, A.; Frade, S.P.; Angelo, P.C.; Gomes, D.A.; Goes, A.M. Human Adipose Tissue-Derived Stem Cells Cultured in Xeno-Free Culture Condition Enhance c-MYC Expression Increasing Proliferation but Bypassing Spontaneous Cell Transformation. Stem Cell Res. Ther. 2015, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Liu, T.; Song, K. Growth Characteristics of Human Adipose-Derived Stem Cells During Long Time Culture Regulated by Cyclin A and Cyclin D1. Appl. Biochem. Biotechnol. 2012, 168, 2230–2244. [Google Scholar] [CrossRef] [PubMed]
- Trieb, K.; Lehner, R.; Stulnig, T.; Sulzbacher, I.; Shroyer, K.R. Survivin Expression in Human Osteosarcoma Is a Marker for Survival. Eur. J. Surg. Oncol. (EJSO) 2003, 29, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Xiong, Y.; Watari, H.; Hanley, S.J.; Konno, Y.; Ihira, K.; Suzuki, F.; Yamada, T.; Kudo, M.; Yue, J.; et al. Suppression of IASPP-Dependent Aggressiveness in Cervical Cancer through Reversal of Methylation Silencing of MicroRNA-124. Sci. Rep. 2016, 6, 35480. [Google Scholar] [CrossRef]
- Ejarque, M.; Ceperuelo-Mallafré, V.; Serena, C.; Pachón, G.; Núñez-Álvarez, Y.; Terrón-Puig, M.; Calvo, E.; Núñez-Roa, C.; Oliva-Olivera, W.; Tinahones, F.J.; et al. Survivin, a Key Player in Cancer Progression, Increases in Obesity and Protects Adipose Tissue Stem Cells from Apoptosis. Cell Death Dis. 2017, 8, e2802. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X.; Yi, Z.; Ma, B.; Wang, H.; Pu, Y.; Wang, J.; Wang, S. WNT5A Promotes Migration and Invasion of Human Osteosarcoma Cells via SRC/ERK/MMP-14 Pathway: WNT5A Promotes Migration and Invasion. Cell Biol. Int. 2018, 42, 598–607. [Google Scholar] [CrossRef]
- Cyr-Depauw, C.; Northey, J.J.; Tabariès, S.; Annis, M.G.; Dong, Z.; Cory, S.; Hallett, M.; Rennhack, J.P.; Andrechek, E.R.; Siegel, P.M. Chordin-Like 1 Suppresses Bone Morphogenetic Protein 4-Induced Breast Cancer Cell Migration and Invasion. Mol. Cell. Biol. 2016, 36, 1509–1525. [Google Scholar] [CrossRef]
- Xu, J.; Wu, S.; Shi, X. Expression of Matrix Metalloproteinase Regulator, RECK, and Its Clinical Significance in Osteosarcoma. J. Orthop. Res. 2010, 28, 1621–1625. [Google Scholar] [CrossRef]
- Chomczynski, P. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Smieszek, A.; Kornicka, K.; Szłapka-Kosarzewska, J.; Androvic, P.; Valihrach, L.; Langerova, L.; Rohlova, E.; Kubista, M.; Marycz, K. Metformin Increases Proliferative Activity and Viability of Multipotent Stromal Stem Cells Isolated from Adipose Tissue Derived from Horses with Equine Metabolic Syndrome. Cells 2019, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence 5’-3’ | Loci | Amplicon | Accession No. |
|---|---|---|---|---|
| Length [bp] | ||||
| BMP-2 | F: TAGACCTGTATCGCAGGCAC | 1024-1043 | 193 | NM_001200.3 |
| R: AACTCCTCCGTGGGGATAGA | 1216-1197 | |||
| BMP-3 | F: GGTAGACTTTGCAGATATTGGCTGG | 1448-1472 | 148 | NM_001201.3 |
| R: AGCTCTCACTATACTCTGGATGGTA | 1595-1571 | |||
| BMP-4 | F: CTGCTCCGGCTGAGTATCTA | 181-200 | 192 | NM_001202.5 |
| R: GTTGCTCGGGATGGCACTAC | 372-353 | |||
| BMP-6 | F: TCAACCGCAAGAGCCTTCTG | 1402-1421 | 164 | NM_001718.5 |
| R: TTTGTGGTGTCGCTGACGAG | 1565-1546 | |||
| BMP-7 | F: CAAGGCCGTCTTCAGTACCC | 850-869 | 145 | NM_001719.2 |
| R: CTCTCGATGGTGGTAGCGTG | 994-975 | |||
| BMPR-IA | F: TAAAGGTGACAGTACACAGGAAACA | 511-535 | 298 | NM_004329.2 |
| R: TCTATGATGGCAAAGCAATGTCC | 808-786 | |||
| BMPR-IB | F: TACAAGCCTGCCATAAGTGAGAAGC | 236-258 | 209 | NM_001203.2 |
| R: ATCATCGTGAAACAATATCCGTCTG | 444-420 | |||
| BMPR-II | F: TCCTCTCATCAGCCATTTGTCCTTTC | 998-1023 | 457 | NM_001204.6 |
| R: AGTTACTACACATTCTTCATAG | 1454-1433 | |||
| GAPDH | F: GTCAGTGGTGGACCTGACCT | 894-913 | 256 | NM_001289746.1 |
| R: CACCACCCTGTTGCTGTAGC | 1149-1130 | |||
| CCND1 | F: GATGCCAACCTCCTCAACGA | 264-283 | 211 | NM_053056.2 |
| R: GGAAGCGGTCCAGGTAGTTC | 474-455 | |||
| C-MYC | F: CTTCTCTCCGTCCTCGGATTCT | 1847-1868 | 204 | NM_001354870.1 |
| R: GAAGGTGATCCAGACTCTGACCTT | 2050-2027 | |||
| BIRC5 | F: ACCGCATCTCTACATTCAAG | 114-143 | 113 | NM_001168.3 |
| R: CAAGTCTGGCTCGTTCTC | 226-209 | |||
| MMP-7 | F: TGTATGGGGAACTGCTGACA | 488-507 | 151 | NM_002423.5 |
| R: GCGTTCATCCTCATCGAAGT | 638-619 | |||
| MMP-14 | F: TCGGCCCAAAGCAGCAGCTTC | 312-332 | 180 | NM_004995.4 |
| R: CTTCATGGTGTCTGCATCAGC | 491-471 | |||
| lncRNA MEG 3 | F: GAGTGTTTCCCTCCCCAAG | 474 -493 | 203 | NR_046470.2 |
| R: GCGTGCCTTTGGTGATTCAG | 676-657 | |||
| miR-21a-5p | TAGCTTATCAGACTGATGTTGA | 18-39 | - | MIMAT0000530 |
| miR-124-3p | TAAGGCACGCGGTGAATGCC | 44-63 | - | MIMAT0000134 |
| miR-203a-3p | GTGAAATGTTTAGGACCACTAG | 65-86 | - | MI0000283 |
| miR-320-3p | AAAAGCTGGGTTGAGAGGGCGA | 48-69 | - | MI0000704 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikora, M.; Krajewska, K.; Marcinkowska, K.; Raciborska, A.; Wiglusz, R.J.; Śmieszek, A. Comparison of Selected Non-Coding RNAs and Gene Expression Profiles between Common Osteosarcoma Cell Lines. Cancers 2022, 14, 4533. https://doi.org/10.3390/cancers14184533
Sikora M, Krajewska K, Marcinkowska K, Raciborska A, Wiglusz RJ, Śmieszek A. Comparison of Selected Non-Coding RNAs and Gene Expression Profiles between Common Osteosarcoma Cell Lines. Cancers. 2022; 14(18):4533. https://doi.org/10.3390/cancers14184533
Chicago/Turabian StyleSikora, Mateusz, Katarzyna Krajewska, Klaudia Marcinkowska, Anna Raciborska, Rafał Jakub Wiglusz, and Agnieszka Śmieszek. 2022. "Comparison of Selected Non-Coding RNAs and Gene Expression Profiles between Common Osteosarcoma Cell Lines" Cancers 14, no. 18: 4533. https://doi.org/10.3390/cancers14184533