
Nrf2 Downregulation Contributes to Epithelial-to-Mesenchymal Transition in Helicobacter pylori-Infected Cells
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Gastric Epithelial Cell Lines’ Culture
2.2. CRISPR-cas9 Mediated Nrf2 Knock-Out
2.3. H. pylori Strains Culture and Co-Culture Model
2.4. ARE-Luciferase and GSH-GSSG Biochemiluminescent Assays
2.5. Cellular ROS Level
2.6. Protein Extraction and Western Blot
2.7. RNA Extraction and RT-qPCR
2.8. Immunofluorescence
2.9. Differential Proteomic Analysis
2.10. Immunohistochemistry on Human Gastric Biopsies
2.11. In Silico Database Analysis
2.12. Statistics
3. Results
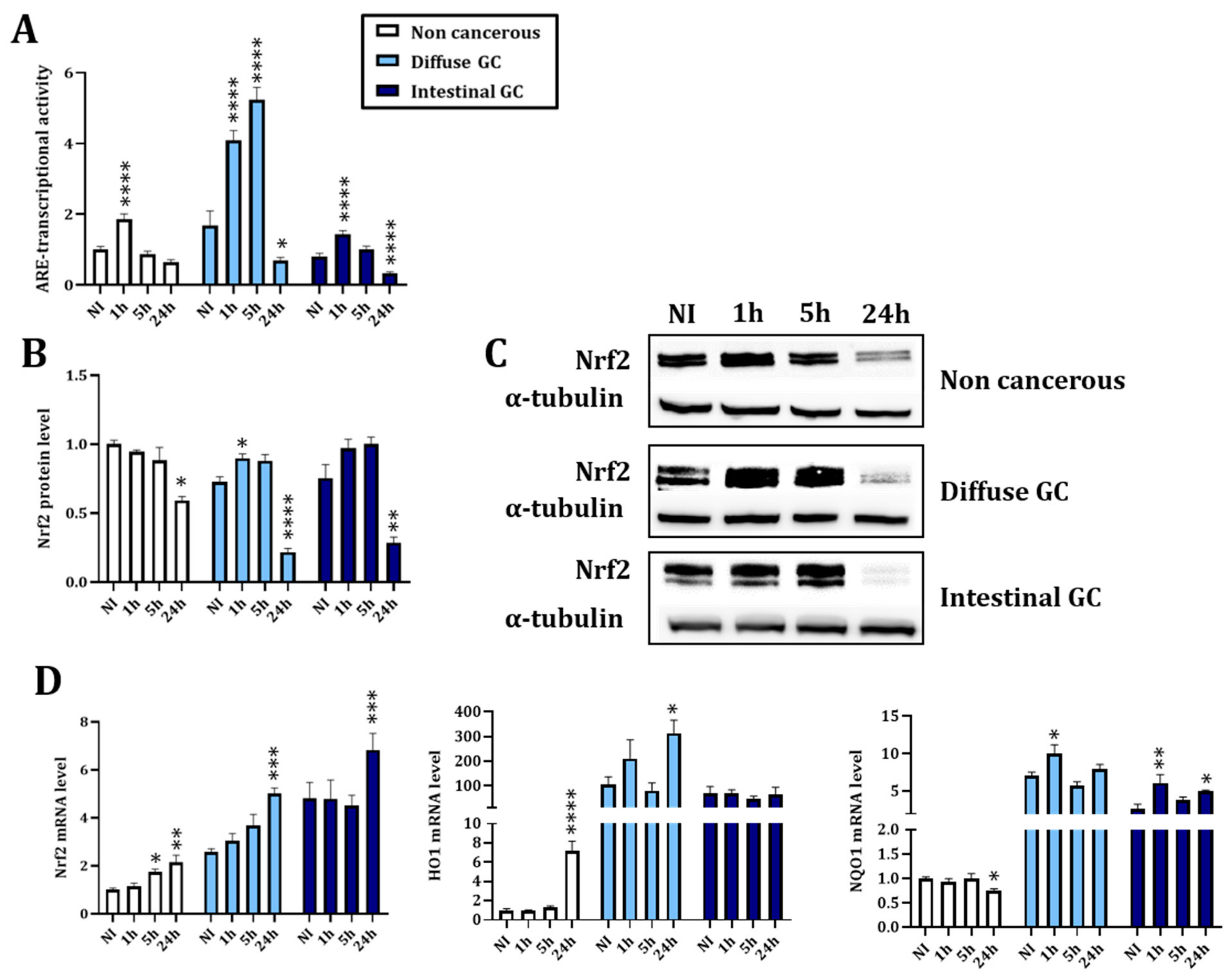
3.1. Nrf2 Signaling Pathway Is Modulated upon Infection with Helicobacter pylori
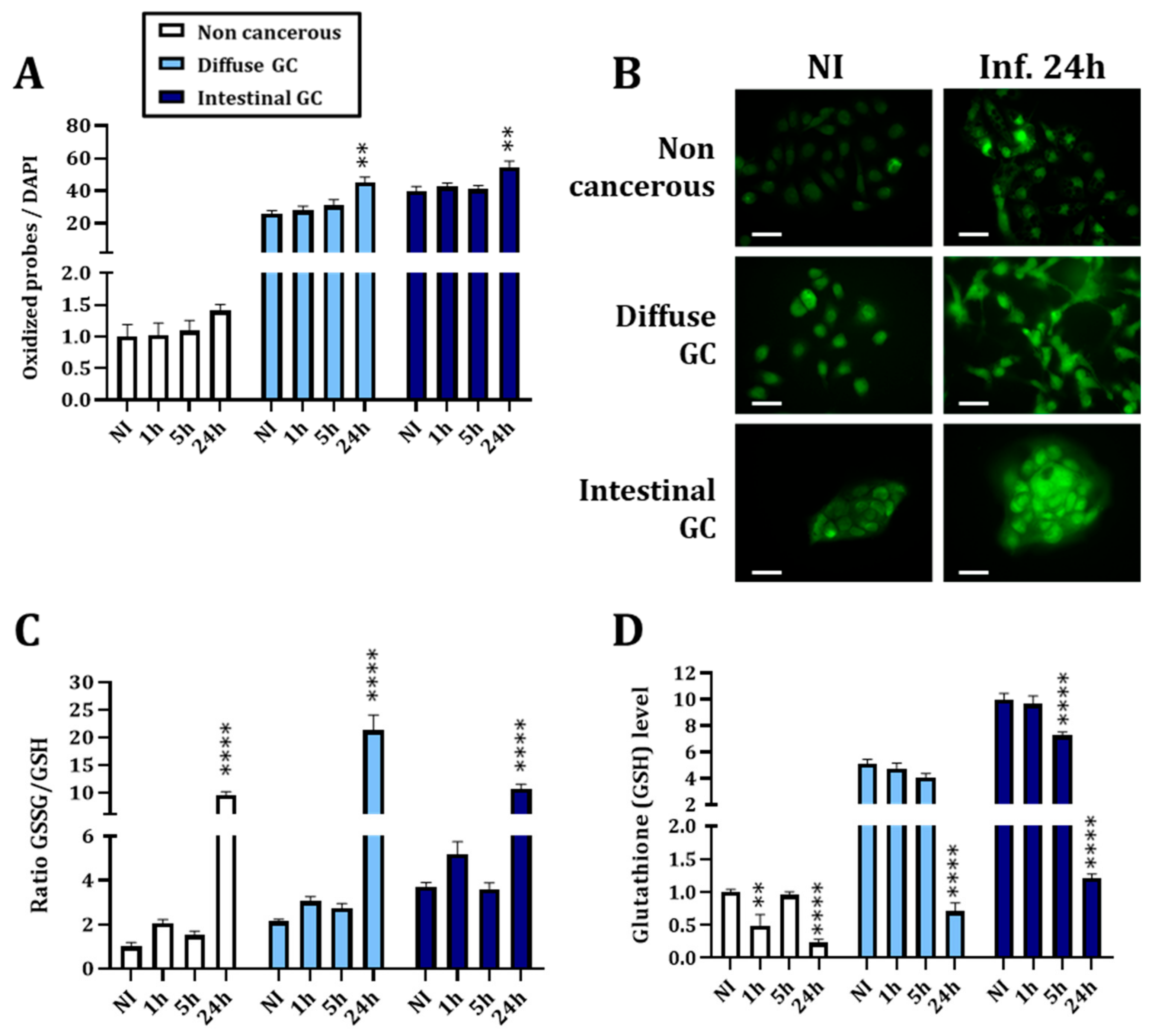
3.1.1. H. pylori Infection Kinetics Is Associated with a Decreased Nrf2 Activity Consistent with Increased Oxidative Stress at 24 h Post Infection
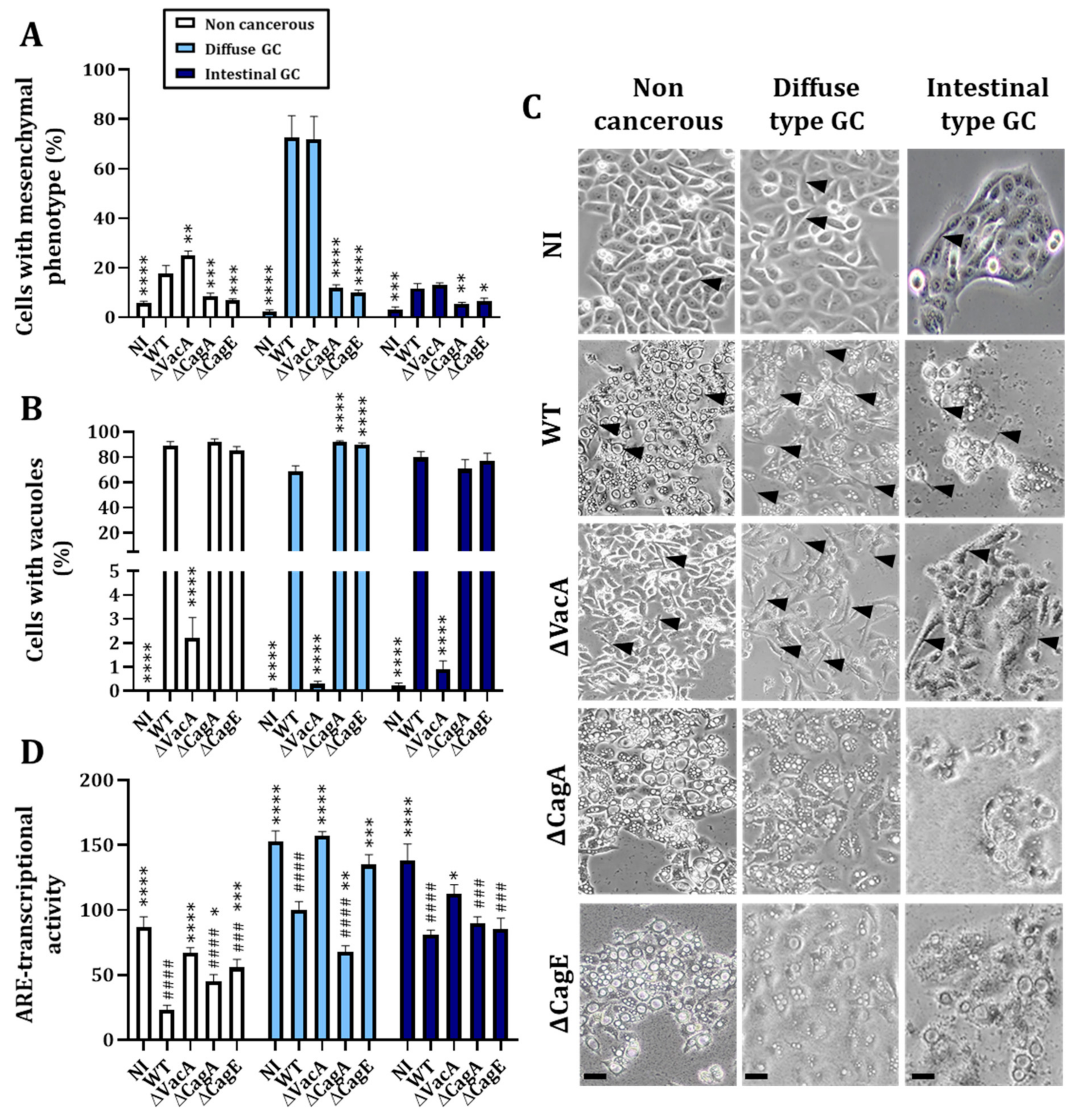
3.1.2. H. pylori Virulence Factor VacA Is Involved in the Regulation of Nrf2 Activity
3.2. Nrf2 Impairment Favors H. pylori-Induced Epithelial-to-Mesenchymal Transition
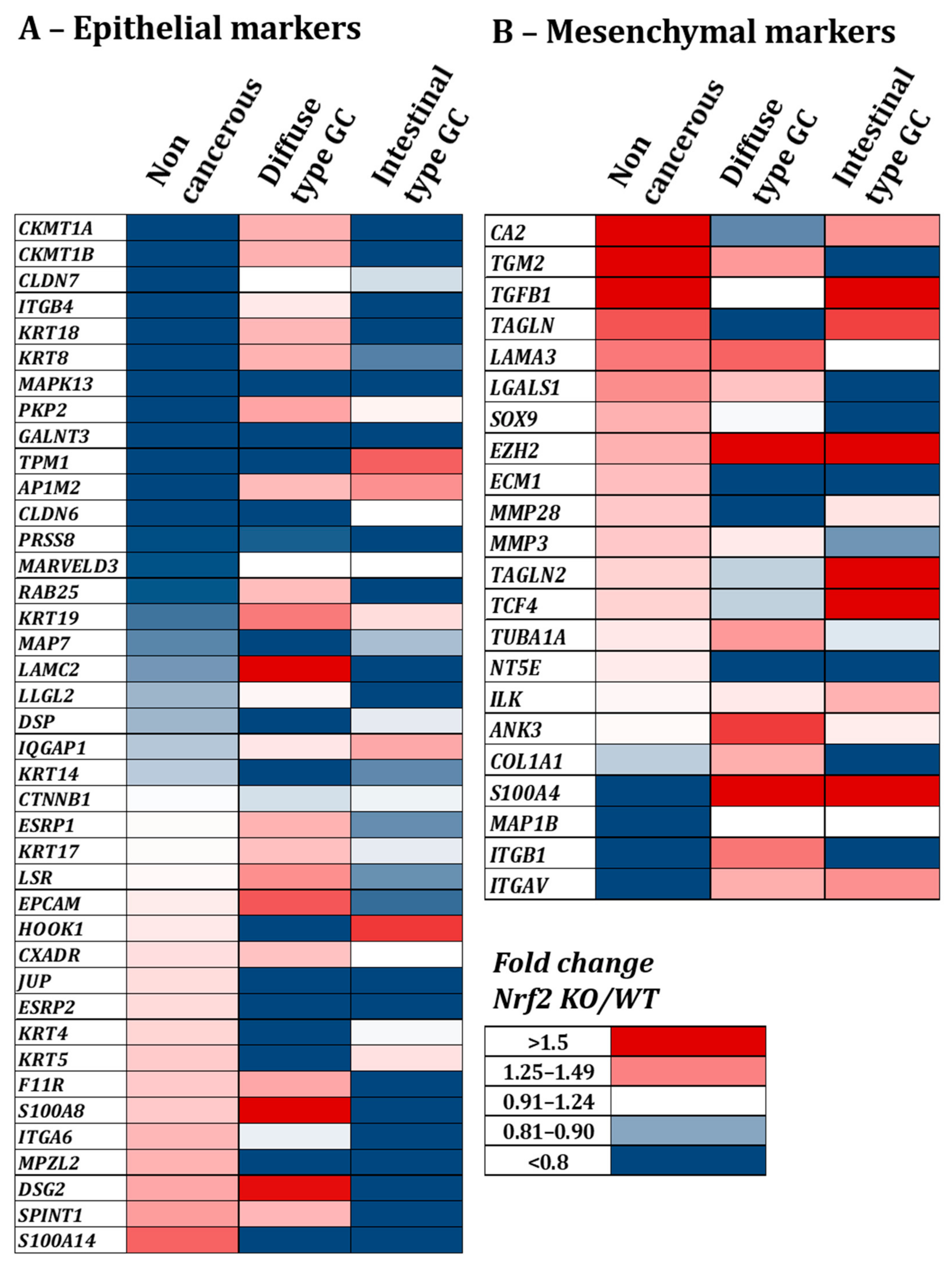
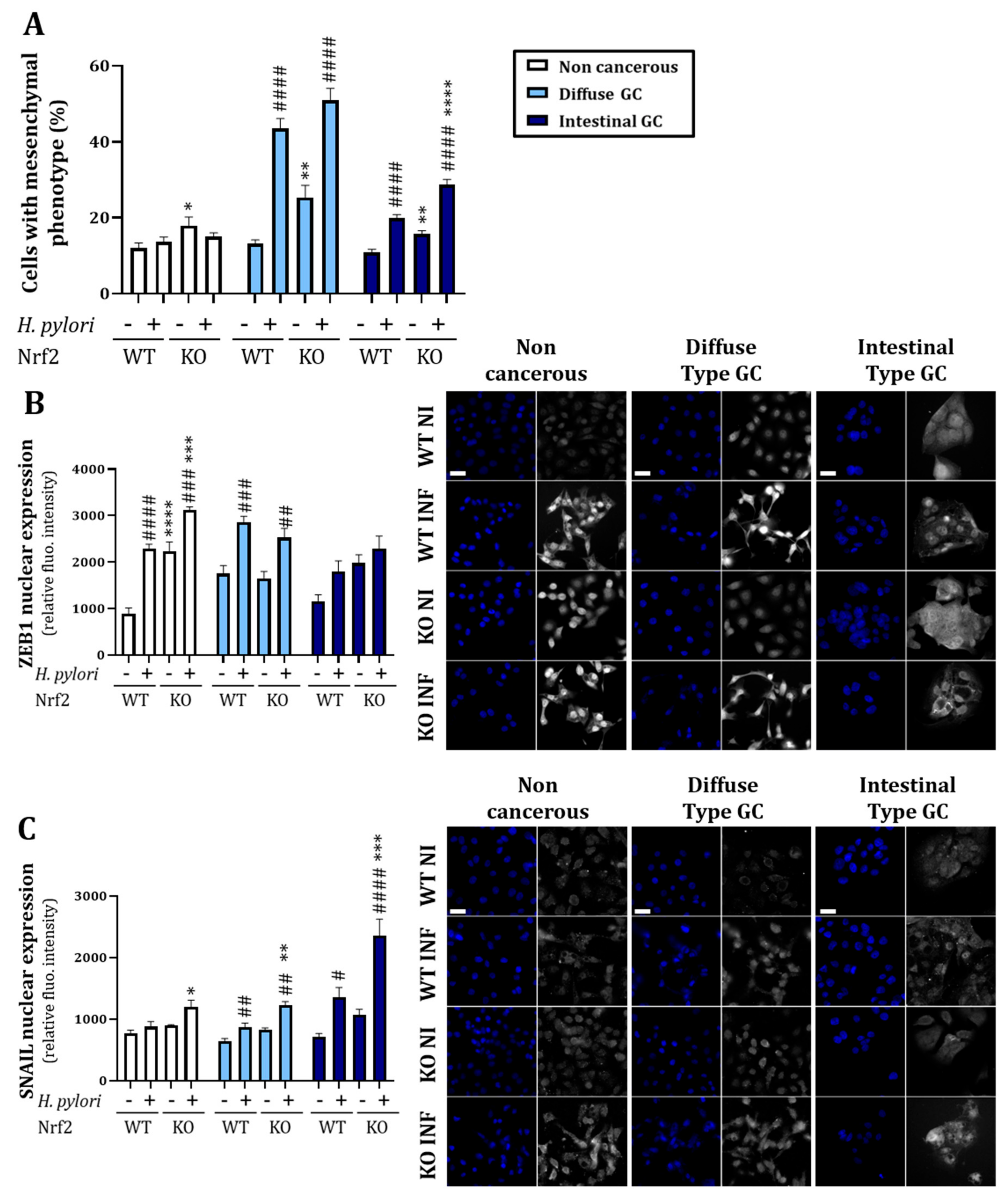
3.2.1. Knock-Out of Nrf2 Favors Decrease in Epithelial Proteins Expression and Increase in Mesenchymal Ones
3.2.2. Knock-Out of Nrf2 Favors H. pylori-Induced Mesenchymal Phenotype Acquisition and EMT Transcription Factor Nuclear Expression
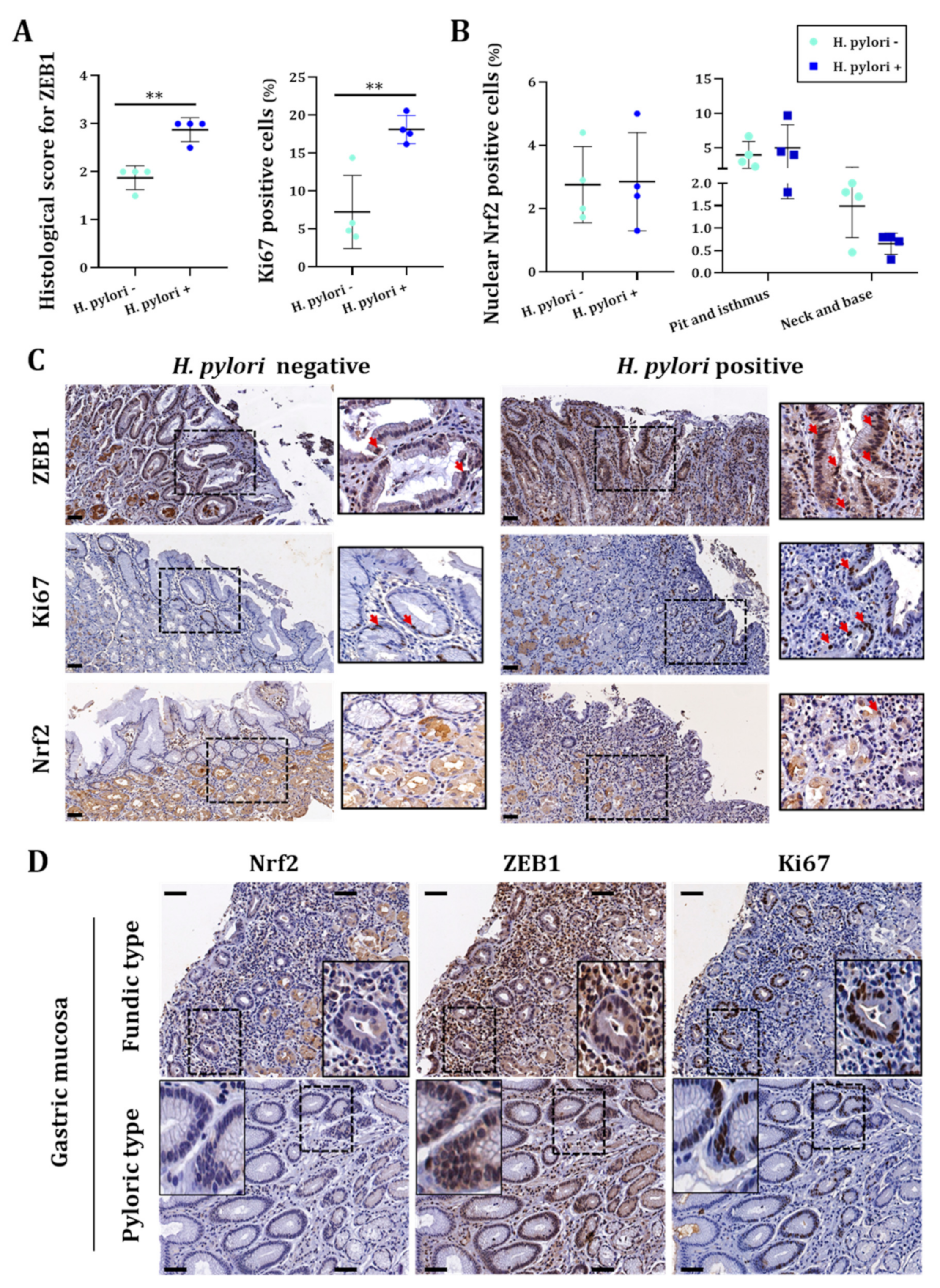
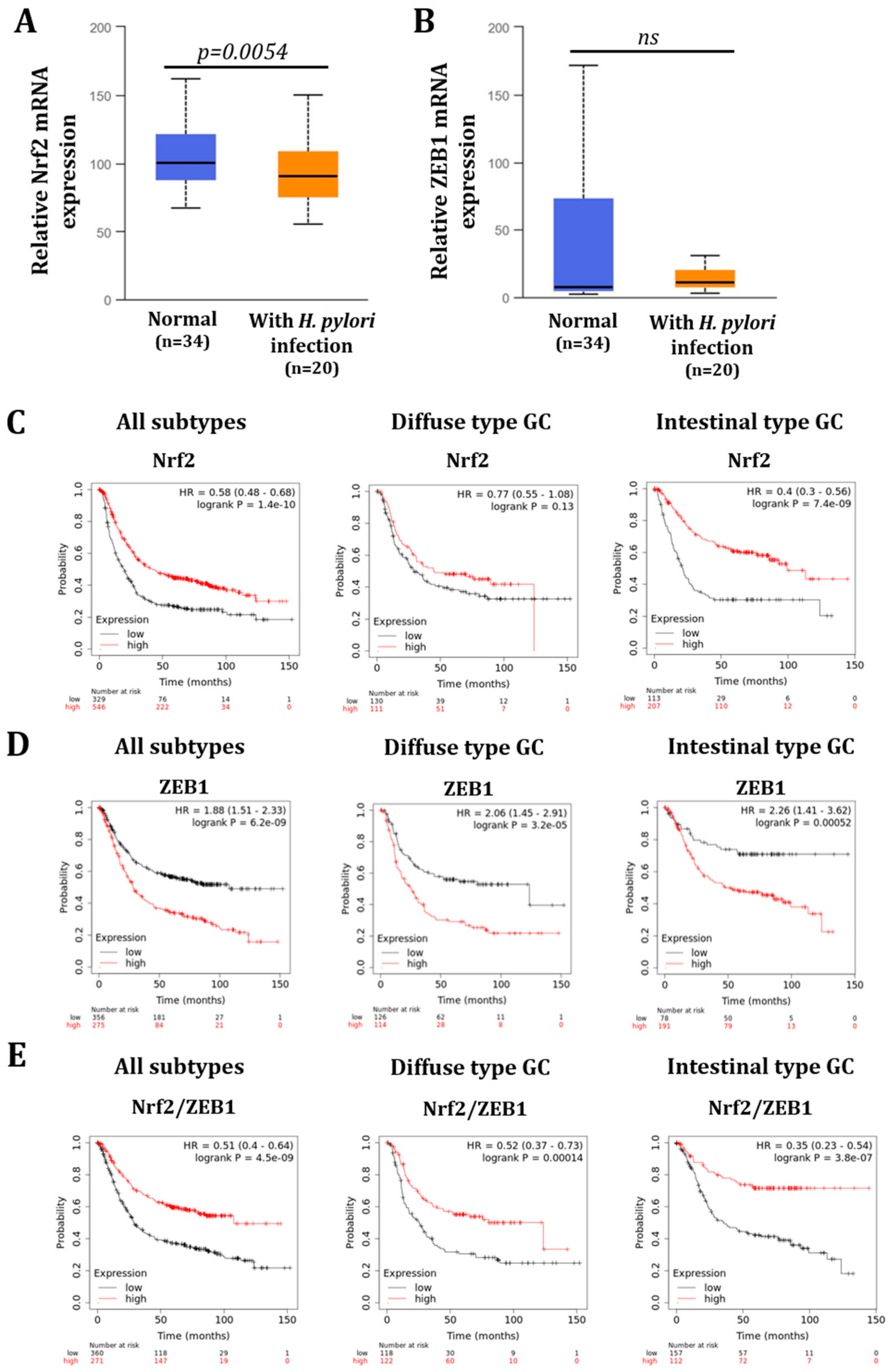
3.3. H. pylori-Infected Patients Have Low Expression of NRF2 Which Is Inversely Correlated to ZEB1 Expression and Related to Poor Overall Survival Probability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding

Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ilic, M.; Ilic, I. Epidemiology of Stomach Cancer. World J. Gastroenterol. 2022, 28, 1187–1203. [Google Scholar] [CrossRef] [PubMed]
- Seeneevassen, L.; Bessède, E.; Mégraud, F.; Lehours, P.; Dubus, P.; Varon, C. Gastric Cancer: Advances in Carcinogenesis Research and New Therapeutic Strategies. Int. J. Mol. Sci. 2021, 22, 3418. [Google Scholar] [CrossRef] [PubMed]
- Zullo, A.; Francesco, V.D.; Gatta, L. Helicobacter pylori Culture: From Bench to Bedside. Ann. Gastroenterol. 2022, 35, 243–248. [Google Scholar] [CrossRef]
- Sharndama, H.C.; Mba, I.E. Helicobacter pylori: An up-to-Date Overview on the Virulence and Pathogenesis Mechanisms. Braz. J. Microbiol. 2022, 53, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Kanemitsu, A.; Knight, C.T.; Hatakeyama, M. Molecular Anatomy and Pathogenic Actions of Helicobacter pylori CagA that Underpin Gastric Carcinogenesis. Cell. Mol. Immunol. 2020, 17, 50–63. [Google Scholar] [CrossRef]
- Román-Román, A.; Martínez-Carrillo, D.N.; Atrisco-Morales, J.; Azúcar-Heziquio, J.C.; Cuevas-Caballero, A.S.; Castañón-Sánchez, C.A.; Reyes-Ríos, R.; Betancourt-Linares, R.; Reyes-Navarrete, S.; Cruz-del Carmen, I.; et al. Helicobacter pylori VacA S1m1 Genotype but Not CagA or BabA2 Increase the Risk of Ulcer and Gastric Cancer in Patients from Southern Mexico. Gut. Pathog. 2017, 9, 18. [Google Scholar] [CrossRef]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-Mesenchymal Transition in Cancer: Complexity and Opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef]
- Baj, J.; Korona-Głowniak, I.; Forma, A.; Maani, A.; Sitarz, E.; Rahnama-Hezavah, M.; Radzikowska, E.; Portincasa, P. Mechanisms of the Epithelial–Mesenchymal Transition and Tumor Microenvironment in Helicobacter pylori-Induced Gastric Cancer. Cells 2020, 9, 1055. [Google Scholar] [CrossRef]
- Bessède, E.; Staedel, C.; Acuña Amador, L.A.; Nguyen, P.H.; Chambonnier, L.; Hatakeyama, M.; Belleannée, G.; Mégraud, F.; Varon, C. Helicobacter pylori Generates Cells with Cancer Stem Cell Properties via Epithelial–Mesenchymal Transition-like Changes. Oncogene 2014, 33, 4123–4131. [Google Scholar] [CrossRef]
- Li, N.; Feng, Y.; Hu, Y.; He, C.; Xie, C.; Ouyang, Y.; Artim, S.C.; Huang, D.; Zhu, Y.; Luo, Z.; et al. Helicobacter Pylori CagA Promotes Epithelial Mesenchymal Transition in Gastric Carcinogenesis via Triggering Oncogenic YAP Pathway. J. Exp. Clin. Cancer Res. 2018, 37, 280. [Google Scholar] [CrossRef] [Green Version]
- Tiffon, C.; Giraud, J.; Molina-Castro, S.E.; Peru, S.; Seeneevassen, L.; Sifré, E.; Staedel, C.; Bessède, E.; Dubus, P.; Mégraud, F.; et al. TAZ Controls Helicobacter pylori-Induced Epithelial-Mesenchymal Transition and Cancer Stem Cell-Like Invasive and Tumorigenic Properties. Cells 2020, 9, 1462. [Google Scholar] [CrossRef] [PubMed]
- Molina-Castro, S.E.; Tiffon, C.; Giraud, J.; Boeuf, H.; Sifre, E.; Giese, A.; Belleannée, G.; Lehours, P.; Bessède, E.; Mégraud, F.; et al. The Hippo Kinase LATS2 Controls Helicobacter Pylori-Induced Epithelial-Mesenchymal Transition and Intestinal Metaplasia in Gastric Mucosa. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 257–276. [Google Scholar] [CrossRef] [PubMed]
- Butcher, L.D.; den Hartog, G.; Ernst, P.B.; Crowe, S.E. Oxidative Stress Resulting From Helicobacter Pylori Infection Contributes to Gastric Carcinogenesis. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Shu, X.; Wang, J. Helicobacter Pylori-Mediated Oxidative Stress and Gastric Diseases: A Review. Front. Microbiol. 2022, 13, 811258. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Suzuki, H. Chapter 5—Oxidative Stress in Stomach Cancer. In Cancer, 2nd ed.; Preedy, V.R., Patel, V.B., Eds.; Academic Press: San Diego, CA, USA, 2021; pp. 49–54. ISBN 978-0-12-819547-5. [Google Scholar]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 Regulatory Network Provides an Interface between Redox and Intermediary Metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Li, R.; Jia, Z.; Zhu, H. Regulation of Nrf2 Signaling. React. Oxyg. Species 2019, 8, 312–322. [Google Scholar] [CrossRef]
- Yanaka, A. Role of NRF2 in Protection of the Gastrointestinal Tract against Oxidative Stress. J. Clin. Biochem. Nutr. 2018, 63, 18–25. [Google Scholar] [CrossRef]
- Kawami, M.; Honda, M.; Hara, T.; Yumoto, R.; Takano, M. Role of Nrf2 in Methotrexate-Induced Epithelial-Mesenchymal Transition in Alveolar A549 Cells. Biol. Pharm. Bull. 2022, 45, 1069–1076. [Google Scholar] [CrossRef]
- Zhang, C.-Y.; Guan, X.-X.; Song, Z.-H.; Jiang, H.-L.; Liu, Y.-B.; Chen, P.; Duan, J.-X.; Zhou, Y. COX-2/SEH Dual Inhibitor PTUPB Attenuates Epithelial-Mesenchymal Transformation of Alveolar Epithelial Cells via Nrf2-Mediated Inhibition of TGF-Β1/Smad Signaling. Oxid. Med. Cell. Longev. 2022, 2022, 5759626. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, Y.; Liu, S.; Yin, H.; Duan, J.; Fan, L.; Zhao, X.; Jiang, B. Implications of Withaferin A for the Metastatic Potential and Drug Resistance in Hepatocellular Carcinoma Cells via Nrf2-Mediated EMT and Ferroptosis. Toxicol. Mech. Methods 2022, 1–9. [Google Scholar] [CrossRef]
- Barinda, A.J.; Arozal, W.; Sandhiutami, N.M.D.; Louisa, M.; Arfian, N.; Sandora, N.; Yusuf, M. Curcumin Prevents Epithelial-to Mesenchymal Transition-Mediated Ovarian Cancer Progression through NRF2/ETBR/ET-1 Axis and Preserves Mitochondria Biogenesis in Kidney after Cisplatin Administration. Adv. Pharm. Bull. 2022, 12, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Vilchez Mercedes, S.A.; Bocci, F.; Ahmed, M.; Eder, I.; Zhu, N.; Levine, H.; Onuchic, J.N.; Jolly, M.K.; Wong, P.K. Nrf2 Modulates the Hybrid Epithelial/Mesenchymal Phenotype and Notch Signaling During Collective Cancer Migration. Front. Mol. Biosci. 2022, 9, 807324. [Google Scholar] [CrossRef] [PubMed]
- Marlink, K.L.; Bacon, K.D.; Sheppard, B.C.; Ashktorab, H.; Smoot, D.T.; Cover, T.L.; Deveney, C.W.; Rutten, M.J. Effects of Helicobacter pylori on Intracellular Ca2+ Signaling in Normal Human Gastric Mucous Epithelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G163–G176. [Google Scholar] [CrossRef] [PubMed]
- Ferrand, J.; Lehours, P.; Schmid-Alliana, A. Helicobacter pylori Infection of Gastrointestinal Epithelial Cells in Vitro Induces Mesenchymal Stem Cell Migration through an NF-KB-Dependent Pathway. PLoS ONE 2011, 6, e29007. [Google Scholar] [CrossRef]
- Haeussler, M.; Schönig, K.; Eckert, H.; Eschstruth, A.; Mianné, J.; Renaud, J.-B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of Off-Target and on-Target Scoring Algorithms and Integration into the Guide RNA Selection Tool CRISPOR. Genome Biol. 2016, 17, 148. [Google Scholar] [CrossRef]
- Bloh, K.; Kanchana, R.; Bialk, P.; Banas, K.; Zhang, Z.; Yoo, B.-C.; Kmiec, E.B. Deconvolution of Complex DNA Repair (DECODR): Establishing a Novel Deconvolution Algorithm for Comprehensive Analysis of CRISPR-Edited Sanger Sequencing Data. CRISPR J. 2021, 4, 120–131. [Google Scholar] [CrossRef]
- Conant, D.; Hsiau, T.; Rossi, N.; Oki, J.; Maures, T.; Waite, K.; Yang, J.; Joshi, S.; Kelso, R.; Holden, K.; et al. Inference of CRISPR Edits from Sanger Trace Data. CRISPR J. 2022, 5, 123–130. [Google Scholar] [CrossRef]
- Seeneevassen, L.; Giraud, J.; Molina-Castro, S.; Sifré, E.; Tiffon, C.; Beauvoit, C.; Staedel, C.; Mégraud, F.; Lehours, P.; Martin, O.C.B.; et al. Leukaemia Inhibitory Factor (LIF) Inhibits Cancer Stem Cells Tumorigenic Properties through Hippo Kinases Activation in Gastric Cancer. Cancers 2020, 12, 2011. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Matsubara, D.; Kishaba, Y.; Yoshimoto, T.; Sakuma, Y.; Sakatani, T.; Tamura, T.; Endo, S.; Sugiyama, Y.; Murakami, Y.; Niki, T. Immunohistochemical Analysis of the Expression of E-Cadherin and ZEB1 in Non-Small Cell Lung Cancer. Pathol. Int. 2014, 64, 560–568. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet. Res. 2021, 23, e27633. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.D.; Cha, J.; Lo, J.; Falkow, S.; Tompkins, L.S. Altered States: Involvement of Phosphorylated CagA in the Induction of Host Cellular Growth Changes by Helicobacter pylori. Proc. Natl. Acad. Sci. USA 1999, 96, 14559–14564. [Google Scholar] [CrossRef] [PubMed]
- Vasaikar, S.V.; Deshmukh, A.P.; den Hollander, P.; Addanki, S.; Kuburich, N.A.; Kudaravalli, S.; Joseph, R.; Chang, J.T.; Soundararajan, R.; Mani, S.A. EMTome: A Resource for Pan-Cancer Analysis of Epithelial-Mesenchymal Transition Genes and Signatures. Br. J. Cancer 2021, 124, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Cloer, E.W.; Goldfarb, D.; Schrank, T.P.; Weissman, B.E.; Major, M.B. NRF2 Activation in Cancer: From DNA to Protein. Cancer Res. 2019, 79, 889–898. [Google Scholar] [CrossRef]
- Liou, G.-Y.; Storz, P. Reactive Oxygen Species in Cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef]
- Kennedy, L.; Sandhu, J.K.; Harper, M.-E.; Cuperlovic-Culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef]
- Abdullah, M.; Greenfield, L.K.; Bronte-Tinkew, D.; Capurro, M.I.; Rizzuti, D.; Jones, N.L. VacA Promotes CagA Accumulation in Gastric Epithelial Cells during Helicobacter pylori Infection. Sci. Rep. 2019, 9, 38. [Google Scholar] [CrossRef]
- Matsuoka, K.; Nishiumi, S.; Yoshida, M.; Kodama, Y. Effects of Helicobacter pylori on the Glutathione-Related Pathway in Gastric Epithelial Cells. Biochem. Biophys. Res. Commun. 2020, 526, 1118–1124. [Google Scholar] [CrossRef]
- Shirin, H.; Pinto, J.T.; Liu, L.U.; Merzianu, M.; Sordillo, E.M.; Moss, S.F. Helicobacter pylori Decreases Gastric Mucosal Glutathione. Cancer Lett. 2001, 164, 127–133. [Google Scholar] [CrossRef]
- Jung, H.K.; Lee, K.E.; Chu, S.H.; Yi, S.Y. Reactive Oxygen Species Activity, Mucosal Lipoperoxidation and Glutathione in Helicobacter pylori-Infected Gastric Mucosa. J. Gastroenterol. Hepatol. 2001, 16, 1336–1340. [Google Scholar] [CrossRef]
- Verhulst, M.L.; van Oijen, A.H.; Roelofs, H.M.; Peters, W.H.; Jansen, J.B. Antral Glutathione Concentration and Glutathione S-Transferase Activity in Patients with and without Helicobacter pylori. Dig. Dis. Sci. 2000, 45, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Oijen, A.H.; Verhulst, M.L.; Roelofs, H.M.; Peters, W.H.; de Boer, W.A.; Jansen, J.B. Eradication of Helicobacter pylori Restores Glutathione S-Transferase Activity and Glutathione Levels in Antral Mucosa. Jpn. J. Cancer Res. 2001, 92, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K.; Matsuno, Y.; Yoshida, K.; Sherpa, M.; Nakajima, M.; Matsuyama, M.; Kiwamoto, T.; Morishima, Y.; Ishii, Y.; Hizawa, N. ROS-Nrf2 Pathway Mediates the Development of TGF-Β1-Induced Epithelial-Mesenchymal Transition through the Activation of Notch Signaling. Eur. J. Cell Biol. 2021, 100, 151181. [Google Scholar] [CrossRef]
- Čipak Gašparović, A.; Milković, L.; Dandachi, N.; Stanzer, S.; Pezdirc, I.; Vrančić, J.; Šitić, S.; Suppan, C.; Balic, M. Chronic Oxidative Stress Promotes Molecular Changes Associated with Epithelial Mesenchymal Transition, NRF2, and Breast Cancer Stem Cell Phenotype. Antioxidants 2019, 8, 633. [Google Scholar] [CrossRef]
- Weitzenböck, H.P.; Gschwendtner, A.; Wiesner, C.; Depke, M.; Schmidt, F.; Trautinger, F.; Hengstschläger, M.; Hundsberger, H.; Mikula, M. Proteome Analysis of NRF2 Inhibition in Melanoma Reveals CD44 Up-Regulation and Increased Apoptosis Resistance upon Vemurafenib Treatment. Cancer Med. 2022, 11, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, H.; Huang, L.; Zhu, X.; Sha, J.; Li, G.; Ma, G.; Zhang, W.; Gu, M.; Guo, Y. Nrf2 Signaling Attenuates Epithelial-to-Mesenchymal Transition and Renal Interstitial Fibrosis via PI3K/Akt Signaling Pathways. Exp. Mol. Pathol. 2019, 111, 104296. [Google Scholar] [CrossRef] [PubMed]
- Kanlaya, R.; Subkod, C.; Nanthawuttiphan, S.; Thongboonkerd, V. Caffeine Prevents Oxalate-Induced Epithelial-Mesenchymal Transition of Renal Tubular Cells by Its Anti-Oxidative Property through Activation of Nrf2 Signaling and Suppression of Snail1 Transcription Factor. Biomed. Pharmacother. 2021, 141, 111870. [Google Scholar] [CrossRef]
- Wu, Z.; Hui, J. Crocin Reverses 1-Methyl-3-Nitroso-1-Nitroguanidine (MNNG)-Induced Malignant Transformation in GES-1 Cells through the Nrf2/Hippo Signaling Pathway. J. Gastrointest. Oncol. 2020, 11, 1242–1252. [Google Scholar] [CrossRef]
- Guan, D.; Zhou, W.; Wei, H.; Wang, T.; Zheng, K.; Yang, C.; Feng, R.; Xu, R.; Fu, Y.; Li, C.; et al. Ferritinophagy-Mediated Ferroptosis and Activation of Keap1/Nrf2/HO-1 Pathway Were Conducive to EMT Inhibition of Gastric Cancer Cells in Action of 2,2′-Di-Pyridineketone Hydrazone Dithiocarbamate Butyric Acid Ester. Oxid. Med. Cell Longev. 2022, 2022, 3920664. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, Y.; Zhou, J.; Wu, R.; Yang, N.; Bao, Q.; Xu, X. Luteolin Alleviates Epithelial-Mesenchymal Transformation Induced by Oxidative Injury in ARPE-19 Cell via Nrf2 and AKT/GSK-3β Pathway. Oxid. Med. Cell Longev. 2022, 2022, 2265725. [Google Scholar] [CrossRef] [PubMed]
- Bocci, F.; Tripathi, S.C.; Vilchez Mercedes, S.A.; George, J.T.; Casabar, J.P.; Wong, P.K.; Hanash, S.M.; Levine, H.; Onuchic, J.N.; Jolly, M.K. NRF2 Activates a Partial Epithelial-Mesenchymal Transition and Is Maximally Present in a Hybrid Epithelial/Mesenchymal Phenotype. Integr. Biol. 2019, 11, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-Y.; Lee, D.-Y.; Kim, E.-H. Anti-Inflammatory and Anti-Oxidative Effect of Korean Propolis on Helicobacter pylori-Induced Gastric Damage in Vitro. J. Microbiol. 2020, 58, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lim, J.W.; Kim, H. Korean Red Ginseng Extract Inhibits IL-8 Expression via Nrf2 Activation in Helicobacter pylori-Infected Gastric Epithelial Cells. Nutrients 2022, 14, 1044. [Google Scholar] [CrossRef]
- Park, J.M.; Han, Y.M.; Park, Y.J.; Hahm, K.B. Dietary Intake of Walnut Prevented Helicobacter pylori-Associated Gastric Cancer through Rejuvenation of Chronic Atrophic Gastritis. J. Clin. Biochem. Nutr. 2021, 68, 37–50. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, J.; Dong, Y.; Shen, X.; Zhang, Z. Vicenin-2 Inhibits the Helicobacterium Pylori Infection Associated Gastric Carcinogenic Events through Modulation of PI3K/AKT and Nrf2 Signaling in GES-1 Cells. J. Biochem. Mol. Toxicol. 2021, 35, e22680. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, A.; Qi, B.; Ma, Z.; Xiong, Y.; Dou, J.; Wang, J. Resveratrol Protects against Helicobacter pylori-Associated Gastritis by Combating Oxidative Stress. Int. J. Mol. Sci. 2015, 16, 27757–27769. [Google Scholar] [CrossRef]
- de la Vega, M.R.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in Cancers: A Double-Edged Sword. Cancer Med. 2019, 8, 2252–2267. [Google Scholar] [CrossRef]
- Zimta, A.-A.; Cenariu, D.; Irimie, A.; Magdo, L.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. The Role of Nrf2 Activity in Cancer Development and Progression. Cancers 2019, 11, 1755. [Google Scholar] [CrossRef] [Green Version]
- Käll, L.; Canterbury, J.D.; Weston, J.; Noble, W.S.; MacCoss, M.J. Semi-Supervised Learning for Peptide Identification from Shotgun Proteomics Datasets. Nat. Methods 2007, 4, 923–925. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| NFE2L2 (Nrf2) | CACATCCAGTCAGAAACCAGTGG | GGAATGTCTGCGCCAAAGCTG |
| HMOX1 (HO1) | CCAGGCAGAGAATGCTGAGTTC | AAGACTGGGCTCTCCTTGTTGC |
| NQO1 | CCTGCCATTCTGAAAGGCTGGT | GTGGTGATGGAAAGCACTGCCT |
| HPRT1 | TGGTCAGGCAGTATAATCCA | GGTCCTTTTCACCAGCAAGCT |
| TBP | TGCACAGGAGCCAAGAGTGAA | CACATCACAGCTCCCCACCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacon, S.; Seeneevassen, L.; Fratacci, A.; Rose, F.; Tiffon, C.; Sifré, E.; Haykal, M.M.; Moubarak, M.M.; Ducournau, A.; Bruhl, L.; et al. Nrf2 Downregulation Contributes to Epithelial-to-Mesenchymal Transition in Helicobacter pylori-Infected Cells. Cancers 2022, 14, 4316. https://doi.org/10.3390/cancers14174316
Bacon S, Seeneevassen L, Fratacci A, Rose F, Tiffon C, Sifré E, Haykal MM, Moubarak MM, Ducournau A, Bruhl L, et al. Nrf2 Downregulation Contributes to Epithelial-to-Mesenchymal Transition in Helicobacter pylori-Infected Cells. Cancers. 2022; 14(17):4316. https://doi.org/10.3390/cancers14174316
Chicago/Turabian StyleBacon, Sarah, Lornella Seeneevassen, Alison Fratacci, Faustine Rose, Camille Tiffon, Elodie Sifré, Maria M. Haykal, Maya M. Moubarak, Astrid Ducournau, Lucie Bruhl, and et al. 2022. "Nrf2 Downregulation Contributes to Epithelial-to-Mesenchymal Transition in Helicobacter pylori-Infected Cells" Cancers 14, no. 17: 4316. https://doi.org/10.3390/cancers14174316