
STEAP1–4 (Six-Transmembrane Epithelial Antigen of the Prostate 1–4) and Their Clinical Implications for Prostate Cancer

 ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
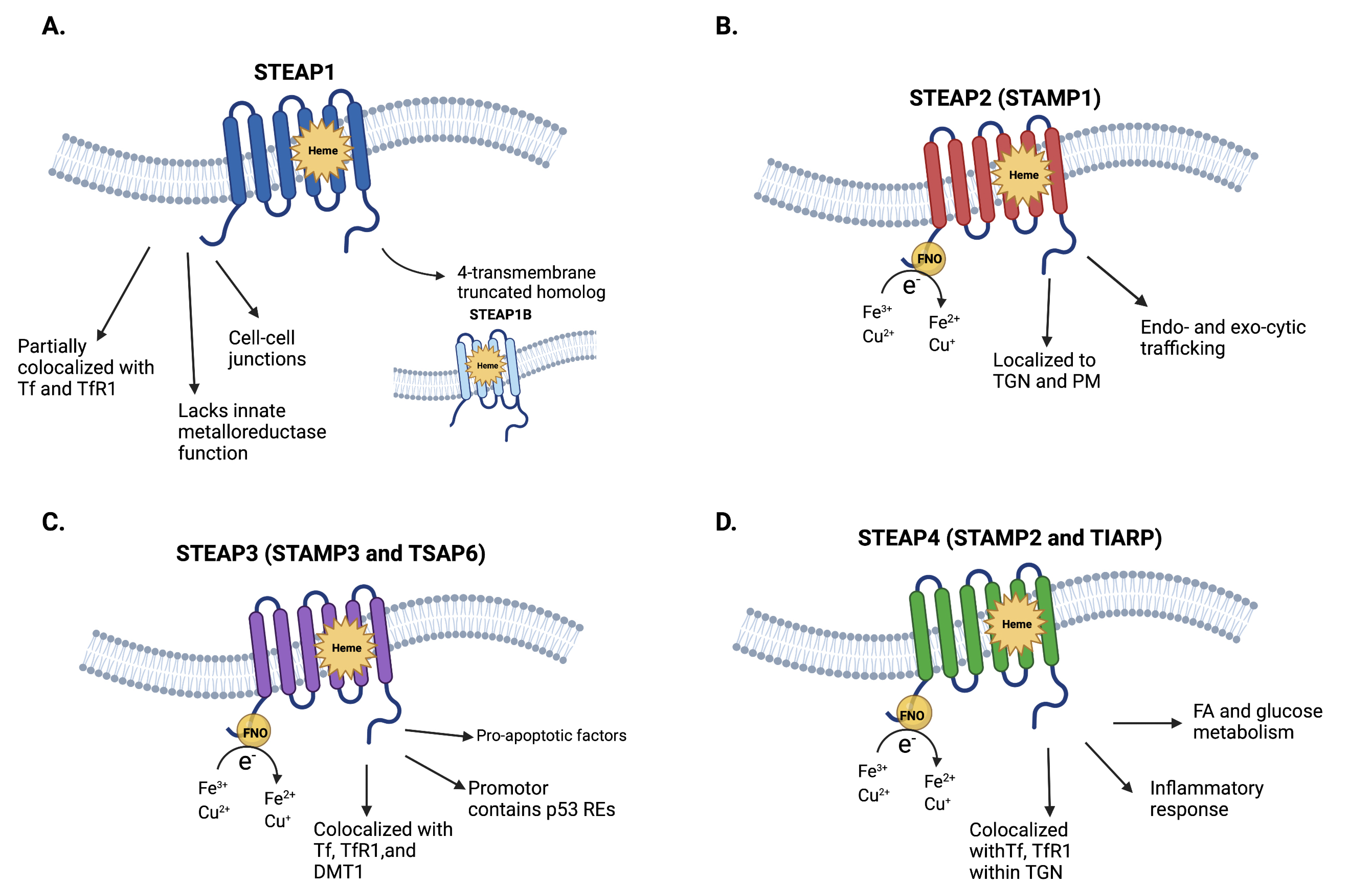
2. Structure and Physiology of STEAP1–4
2.1. STEAP1 and STEAP1B
2.2. STEAP2-4
3. Pathophysiologic Roles of STEAP1–4 in Prostate Cancer
3.1. STEAP1 in Prostate Cancer
3.2. STEAP2-4 in Prostate Cancer
4. STEAP1–4 as Biomarkers and Therapeutic Targets for Prostate Cancer
4.1. STEAP1, STEAP2, and STEAP4 as Biomarkers for Prostate Cancer
4.2. STEAP1 as a Therapeutic Target for Prostate Cancer
4.3. Other Considerations and Perspectives on the Role of STEAP1–4 in Prostate Cancer
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.J. The Diagnosis and Treatment of Prostate Cancer: A Review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef]
- Quigley, D.A.; Dang, H.X.; Zhao, S.G.; Lloyd, P.; Aggarwal, R.; Alumkal, J.J.; Foye, A.; Kothari, V.; Perry, M.D.; Bailey, A.M.; et al. Genomic Hallmarks and Structural Variation in Metastatic Prostate Cancer. Cell 2018, 174, 758–769.e9. [Google Scholar] [CrossRef] [Green Version]
- Ohgami, R.S.; Campagna, D.R.; McDonald, A.; Fleming, M.D. The Steap proteins are metalloreductases. Blood 2006, 108, 1388–1394. [Google Scholar] [CrossRef]
- Korkmaz, K.S.; Elbi, C.; Korkmaz, C.G.; Loda, M.; Hager, G.L.; Saatcioglu, F. Molecular cloning and characterization of STAMP1, a highly prostate-specific six transmembrane protein that is overexpressed in prostate cancer. J. Biol. Chem. 2002, 277, 36689–36696. [Google Scholar] [CrossRef] [Green Version]
- Passer, B.J.; Nancy-Portebois, V.; Amzallag, N.; Prieur, S.; Cans, C.; Roborel de Climens, A.; Fiucci, G.; Bouvard, V.; Tuynder, M.; Susini, L.; et al. The p53-inducible TSAP6 gene product regulates apoptosis and the cell cycle and interacts with Nix and the Myt1 kinase. Proc. Natl. Acad. Sci. USA 2003, 100, 2284–2289. [Google Scholar] [CrossRef] [Green Version]
- Korkmaz, C.G.; Korkmaz, K.S.; Kurys, P.; Elbi, C.; Wang, L.; Klokk, T.I.; Hammarstrom, C.; Troen, G.; Svindland, A.; Hager, G.L.; et al. Molecular cloning and characterization of STAMP2, an androgen-regulated six transmembrane protein that is overexpressed in prostate cancer. Oncogene 2005, 24, 4934–4945. [Google Scholar] [CrossRef] [Green Version]
- Gomes, I.M.; Santos, C.R.; Maia, C.J. Expression of STEAP1 and STEAP1B in prostate cell lines, and the putative regulation of STEAP1 by post-transcriptional and post-translational mechanisms. Genes. Cancer 2014, 5, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Hubert, R.S.; Vivanco, I.; Chen, E.; Rastegar, S.; Leong, K.; Mitchell, S.C.; Madraswala, R.; Zhou, Y.; Kuo, J.; Raitano, A.B.; et al. STEAP: A prostate-specific cell-surface antigen highly expressed in human prostate tumors. Proc. Natl. Acad. Sci. USA 1999, 96, 14523–14528. [Google Scholar] [CrossRef] [Green Version]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef]
- Lee, P.L.; Gelbart, T.; West, C.; Halloran, C.; Beutler, E. The human Nramp2 gene: Characterization of the gene structure, alternative splicing, promoter region and polymorphisms. Blood Cells Mol. Dis. 1998, 24, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.D. Iron-copper interactions: Some new revelations. Nutr. Rev. 1994, 52, 311–315. [Google Scholar] [CrossRef]
- Guo, Q.; Li, L.; Hou, S.; Yuan, Z.; Li, C.; Zhang, W.; Zheng, L.; Li, X. The Role of Iron in Cancer Progression. Front. Oncol. 2021, 11, 778492. [Google Scholar] [CrossRef]
- Knutson, M.D. Steap proteins: Implications for iron and copper metabolism. Nutr. Rev. 2007, 65, 335–340. [Google Scholar] [CrossRef]
- Gomes, I.M.; Rocha, S.M.; Gaspar, C.; Alvelos, M.I.; Santos, C.R.; Socorro, S.; Maia, C.J. Knockdown of STEAP1 inhibits cell growth and induces apoptosis in LNCaP prostate cancer cells counteracting the effect of androgens. Med. Oncol. 2018, 35, 40. [Google Scholar] [CrossRef]
- Kim, K.; Mitra, S.; Wu, G.; Berka, V.; Song, J.; Yu, Y.; Poget, S.; Wang, D.N.; Tsai, A.L.; Zhou, M. Six-Transmembrane Epithelial Antigen of Prostate 1 (STEAP1) Has a Single b Heme and Is Capable of Reducing Metal Ion Complexes and Oxygen. Biochemistry 2016, 55, 6673–6684. [Google Scholar] [CrossRef]
- Oosterheert, W.; van Bezouwen, L.S.; Rodenburg, R.N.P.; Granneman, J.; Forster, F.; Mattevi, A.; Gros, P. Cryo-EM structures of human STEAP4 reveal mechanism of iron(III) reduction. Nat. Commun. 2018, 9, 4337. [Google Scholar] [CrossRef]
- Oosterheert, W.; Gros, P. Cryo-electron microscopy structure and potential enzymatic function of human six-transmembrane epithelial antigen of the prostate 1 (STEAP1). J. Biol. Chem. 2020, 295, 9502–9512. [Google Scholar] [CrossRef]
- Sikkeland, J.; Sheng, X.; Jin, Y.; Saatcioglu, F. STAMPing at the crossroads of normal physiology and disease states. Mol. Cell. Endocrinol. 2016, 425, 26–36. [Google Scholar] [CrossRef]
- Oosterheert, W.; Reis, J.; Gros, P.; Mattevi, A. An Elegant Four-Helical Fold in NOX and STEAP Enzymes Facilitates Electron Transport across Biomembranes-Similar Vehicle, Different Destination. Acc. Chem. Res. 2020, 53, 1969–1980. [Google Scholar] [CrossRef]
- Porkka, K.P.; Helenius, M.A.; Visakorpi, T. Cloning and characterization of a novel six-transmembrane protein STEAP2, expressed in normal and malignant prostate. Lab. Investig. 2002, 82, 1573–1582. [Google Scholar] [CrossRef] [Green Version]
- Ohgami, R.S.; Campagna, D.R.; Greer, E.L.; Antiochos, B.; McDonald, A.; Chen, J.; Sharp, J.J.; Fujiwara, Y.; Barker, J.E.; Fleming, M.D. Identification of a ferrireductase required for efficient transferrin-dependent iron uptake in erythroid cells. Nat. Genet. 2005, 37, 1264–1269. [Google Scholar] [CrossRef] [Green Version]
- Moldes, M.; Lasnier, F.; Gauthereau, X.; Klein, C.; Pairault, J.; Feve, B.; Chambaut-Guerin, A.M. Tumor necrosis factor-alpha-induced adipose-related protein (TIARP), a cell-surface protein that is highly induced by tumor necrosis factor-alpha and adipose conversion. J. Biol. Chem. 2001, 276, 33938–33946. [Google Scholar] [CrossRef] [Green Version]
- Scarl, R.T.; Lawrence, C.M.; Gordon, H.M.; Nunemaker, C.S. STEAP4: Its emerging role in metabolism and homeostasis of cellular iron and copper. J. Endocrinol. 2017, 234, R123–R134. [Google Scholar] [CrossRef]
- Challita-Eid, P.M.; Morrison, K.; Etessami, S.; An, Z.; Morrison, K.J.; Perez-Villar, J.J.; Raitano, A.B.; Jia, X.C.; Gudas, J.M.; Kanner, S.B.; et al. Monoclonal antibodies to six-transmembrane epithelial antigen of the prostate-1 inhibit intercellular communication In Vitro and growth of human tumor xenografts In Vivo. Cancer Res. 2007, 67, 5798–5805. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Tamura, Y.; Kobayashi, J.; Kamiguchi, K.; Hirohashi, Y.; Miyazaki, A.; Torigoe, T.; Asanuma, H.; Hiratsuka, H.; Sato, N. Six-transmembrane epithelial antigen of the prostate-1 plays a role for In Vivo tumor growth via intercellular communication. Exp. Cell. Res. 2013, 319, 2617–2626. [Google Scholar] [CrossRef]
- Fierro, F.A.; Sierralta, W.D.; Epunan, M.J.; Minguell, J.J. Marrow-derived mesenchymal stem cells: Role in epithelial tumor cell determination. Clin. Exp. Metastasis 2004, 21, 313–319. [Google Scholar] [CrossRef]
- Vaghjiani, R.J.; Talma, S.; Murphy, C.L. Six-transmembrane epithelial antigen of the prostate (STEAP1 and STEAP2)-differentially expressed by murine and human mesenchymal stem cells. Tissue Eng. Part A 2009, 15, 2073–2083. [Google Scholar] [CrossRef]
- Whiteland, H.; Spencer-Harty, S.; Thomas, D.H.; Davies, C.; Morgan, C.; Kynaston, H.; Bose, P.; Fenn, N.; Lewis, P.D.; Bodger, O.; et al. Putative prognostic epithelial-to-mesenchymal transition biomarkers for aggressive prostate cancer. Exp. Mol. Pathol. 2013, 95, 220–226. [Google Scholar] [CrossRef]
- Jiao, Z.; Huang, L.; Sun, J.; Xie, J.; Wang, T.; Yin, X.; Zhang, H.; Chen, J. Six-transmembrane epithelial antigen of the prostate 1 expression promotes ovarian cancer metastasis by aiding progression of epithelial-to-mesenchymal transition. Histochem. Cell Biol. 2020, 154, 215–230. [Google Scholar] [CrossRef]
- Epstein, J.I.; Allsbrook, W.C., Jr.; Amin, M.B.; Egevad, L.L.; Committee, I.G. The 2005 International Society of Urological Pathology (ISUP) Consensus Conference on Gleason Grading of Prostatic Carcinoma. Am. J. Surg. Pathol. 2005, 29, 1228–1242. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, P.A. Gleason grading and prognostic factors in carcinoma of the prostate. Mod. Pathol. 2004, 17, 292–306. [Google Scholar] [CrossRef]
- Wang, L.; Jin, Y.; Arnoldussen, Y.J.; Jonson, I.; Qu, S.; Maelandsmo, G.M.; Kristian, A.; Risberg, B.; Waehre, H.; Danielsen, H.E.; et al. STAMP1 is both a proliferative and an antiapoptotic factor in prostate cancer. Cancer Res. 2010, 70, 5818–5828. [Google Scholar] [CrossRef] [Green Version]
- Burnell, S.E.A.; Spencer-Harty, S.; Howarth, S.; Bodger, O.; Kynaston, H.; Morgan, C.; Doak, S.H. STEAP2 Knockdown Reduces the Invasive Potential of Prostate Cancer Cells. Sci. Rep. 2018, 8, 6252. [Google Scholar] [CrossRef]
- Whiteland, H.; Spencer-Harty, S.; Morgan, C.; Kynaston, H.; Thomas, D.H.; Bose, P.; Fenn, N.; Lewis, P.; Jenkins, S.; Doak, S.H. A role for STEAP2 in prostate cancer progression. Clin. Exp. Metastasis 2014, 31, 909–920. [Google Scholar] [CrossRef]
- Jin, Y.; Wang, L.; Qu, S.; Sheng, X.; Kristian, A.; Maelandsmo, G.M.; Pallmann, N.; Yuca, E.; Tekedereli, I.; Gorgulu, K.; et al. STAMP2 increases oxidative stress and is critical for prostate cancer. EMBO Mol. Med. 2015, 7, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Bredell, B.X.; Anderson, E.R.; Martin, A.; Mays, C.; Nagao-Kitamoto, H.; Huang, S.; Gyorffy, B.; Greenson, J.K.; Hardiman, K.; et al. Quantitative proteomics identifies STEAP4 as a critical regulator of mitochondrial dysfunction linking inflammation and colon cancer. Proc. Natl. Acad. Sci. USA 2017, 114, E9608–E9617. [Google Scholar] [CrossRef] [Green Version]
- Lespagnol, A.; Duflaut, D.; Beekman, C.; Blanc, L.; Fiucci, G.; Marine, J.C.; Vidal, M.; Amson, R.; Telerman, A. Exosome secretion, including the DNA damage-induced p53-dependent secretory pathway, is severely compromised in TSAP6/Steap3-null mice. Cell Death Differ. 2008, 15, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- Porkka, K.P.; Nupponen, N.N.; Tammela, T.L.; Vessella, R.L.; Visakorpi, T. Human pHyde is not a classical tumor suppressor gene in prostate cancer. Int. J. Cancer 2003, 106, 729–735. [Google Scholar] [CrossRef]
- Isobe, T.; Baba, E.; Arita, S.; Komoda, M.; Tamura, S.; Shirakawa, T.; Ariyama, H.; Takaishi, S.; Kusaba, H.; Ueki, T.; et al. Human STEAP3 maintains tumor growth under hypoferric condition. Exp. Cell Res. 2011, 317, 2582–2591. [Google Scholar] [CrossRef]
- Han, M.; Xu, R.; Wang, S.; Yang, N.; Ni, S.; Zhang, Q.; Xu, Y.; Zhang, X.; Zhang, C.; Wei, Y.; et al. Six-Transmembrane Epithelial Antigen of Prostate 3 Predicts Poor Prognosis and Promotes Glioblastoma Growth and Invasion. Neoplasia 2018, 20, 543–554. [Google Scholar] [CrossRef]
- Rutkowski, D.T.; Kaufman, R.J. All roads lead to ATF4. Dev. Cell 2003, 4, 442–444. [Google Scholar] [CrossRef] [Green Version]
- Lewerenz, J.; Maher, P. Control of redox state and redox signaling by neural antioxidant systems. Antioxid. Redox Signal. 2011, 14, 1449–1465. [Google Scholar] [CrossRef]
- Li, W.; Yin, X.; Yan, Y.; Liu, C.; Li, G. STEAP4 knockdown inhibits the proliferation of prostate cancer cells by activating the cGMP-PKG pathway under lipopolysaccharide-induced inflammatory microenvironment. Int. Immunopharmacol. 2021, 101, 108311. [Google Scholar] [CrossRef]
- Tamura, T.; Chiba, J. STEAP4 regulates focal adhesion kinase activation and CpG motifs within STEAP4 promoter region are frequently methylated in DU145, human androgen-independent prostate cancer cells. Int. J. Mol. Med. 2009, 24, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Andriole, G.L.; Crawford, E.D.; Grubb, R.L. 3rd ed.; Buys, S.S.; Chia, D.; Church, T.R.; Fouad, M.N.; Gelmann, E.P.; Kvale, P.A.; Reding, D.J.; et al. Mortality results from a randomized prostate-cancer screening trial. N. Engl. J. Med. 2009, 360, 1310–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, F.H.; Hugosson, J.; Roobol, M.J.; Tammela, T.L.; Ciatto, S.; Nelen, V.; Kwiatkowski, M.; Lujan, M.; Lilja, H.; Zappa, M.; et al. Screening and prostate-cancer mortality in a randomized European study. N. Engl. J. Med. 2009, 360, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Thompson, I.M.; Pauler, D.K.; Goodman, P.J.; Tangen, C.M.; Lucia, M.S.; Parnes, H.L.; Minasian, L.M.; Ford, L.G.; Lippman, S.M.; Crawford, E.D.; et al. Prevalence of prostate cancer among men with a prostate-specific antigen level < or = 4.0 ng per milliliter. N. Engl. J. Med. 2004, 350, 2239–2246. [Google Scholar] [CrossRef] [Green Version]
- Moyer, V.A.; Force, U.S.P.S.T. Screening for prostate cancer: U.S. Preventive Services Task Force recommendation statement. Ann. Intern. Med. 2012, 157, 120–134. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xu, C.; Yu, Q.; Zhong, C.; Peng, Y.; Chen, J.; Chen, G. Comprehensive landscape of STEAP family functions and prognostic prediction value in glioblastoma. J. Cell. Physiol. 2021, 236, 2988–3000. [Google Scholar] [CrossRef]
- Wu, H.T.; Chen, W.J.; Xu, Y.; Shen, J.X.; Chen, W.T.; Liu, J. The Tumor Suppressive Roles and Prognostic Values of STEAP Family Members in Breast Cancer. Biomed. Res. Int. 2020, 2020, 9578484. [Google Scholar] [CrossRef]
- Cheung, I.Y.; Feng, Y.; Danis, K.; Shukla, N.; Meyers, P.; Ladanyi, M.; Cheung, N.K. Novel markers of subclinical disease for Ewing family tumors from gene expression profiling. Clin. Cancer Res. 2007, 13, 6978–6983. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Niu, X.; Li, Y.; Xu, Z.; Chen, J.; Xu, G. Expression and prognostic analyses of the significance of STEAP1 and STEAP2 in lung cancer. World J. Surg. Oncol. 2022, 20, 96. [Google Scholar] [CrossRef]
- Burnell, S.E.A.; Spencer-Harty, S.; Howarth, S.; Bodger, O.; Kynaston, H.; Morgan, C.; Doak, S.H. Utilisation of the STEAP protein family in a diagnostic setting may provide a more comprehensive prognosis of prostate cancer. PLoS ONE 2019, 14, e0220456. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.; Salmond, N.; Lynn, K.S.; Leong, H.S.; Williams, K.C. Clinical significance of STEAP1 extracellular vesicles in prostate cancer. Prostate Cancer Prostatic Dis. 2021, 24, 802–811. [Google Scholar] [CrossRef]
- Mariscal, J.; Vagner, T.; Kim, M.; Zhou, B.; Chin, A.; Zandian, M.; Freeman, M.R.; You, S.; Zijlstra, A.; Yang, W.; et al. Comprehensive palmitoyl-proteomic analysis identifies distinct protein signatures for large and small cancer-derived extracellular vesicles. J. Extracell. Vesicles 2020, 9, 1764192. [Google Scholar] [CrossRef]
- Ihlaseh-Catalano, S.M.; Drigo, S.A.; de Jesus, C.M.; Domingues, M.A.; Trindade Filho, J.C.; de Camargo, J.L.; Rogatto, S.R. STEAP1 protein overexpression is an independent marker for biochemical recurrence in prostate carcinoma. Histopathology 2013, 63, 678–685. [Google Scholar] [CrossRef]
- Gomes, I.M.; Arinto, P.; Lopes, C.; Santos, C.R.; Maia, C.J. STEAP1 is overexpressed in prostate cancer and prostatic intraepithelial neoplasia lesions, and it is positively associated with Gleason score. Urol. Oncol. 2014, 32, 53.e23–53.e29. [Google Scholar] [CrossRef]
- O’Donoghue, J.A.; Danila, D.C.; Pandit-Taskar, N.; Beylergil, V.; Cheal, S.M.; Fleming, S.E.; Fox, J.J.; Ruan, S.; Zanzonico, P.B.; Ragupathi, G.; et al. Pharmacokinetics and Biodistribution of a [(89)Zr]Zr-DFO-MSTP2109A Anti-STEAP1 Antibody in Metastatic Castration-Resistant Prostate Cancer Patients. Mol. Pharm. 2019, 16, 3083–3090. [Google Scholar] [CrossRef]
- Carrasquillo, J.A.; Fine, B.M.; Pandit-Taskar, N.; Larson, S.M.; Fleming, S.E.; Fox, J.J.; Cheal, S.M.; O’Donoghue, J.A.; Ruan, S.; Ragupathi, G.; et al. Imaging Patients with Metastatic Castration-Resistant Prostate Cancer Using (89)Zr-DFO-MSTP2109A Anti-STEAP1 Antibody. J. Nucl. Med. 2019, 60, 1517–1523. [Google Scholar] [CrossRef] [Green Version]
- Moreaux, J.; Kassambara, A.; Hose, D.; Klein, B. STEAP1 is overexpressed in cancers: A promising therapeutic target. Biochem. Biophys. Res. Commun. 2012, 429, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Esmaeili, S.A.; Nejatollahi, F.; Sahebkar, A. Inhibition of Intercellular Communication between Prostate Cancer Cells by A Specific Anti-STEAP-1 Single Chain Antibody. Anticancer Agents Med. Chem. 2018, 18, 1674–1679. [Google Scholar] [CrossRef]
- Stenzl, A.; Feyerabend, S.; Syndikus, I.; Sarosiek, T.; Kübler, H.; Heidenreich, A.; Cathomas, R.; Grüllich, C.; Loriot, Y.; Perez Garcia, S.L.; et al. Results of the randomized, placebo-controlled phase I/IIB trial of CV9104, an mRNA based cancer immunotherapy, in patients with metastatic castration-resistant prostate cancer (mCRPC). J. Immunother. Cancer 2017, 28, V408–V409. [Google Scholar] [CrossRef]
- Guo, L.; Xie, H.; Zhang, Z.; Wang, Z.; Peng, S.; Niu, Y.; Shang, Z. Fusion Protein Vaccine Based on Ag85B and STEAP1 Induces a Protective Immune Response against Prostate Cancer. Vaccines 2021, 9, 786. [Google Scholar] [CrossRef]
- Allahbahshi, E. Vaccination Against Prostate Cancer; ISRCTN Registry; Ahvaz Jundishapur University of Medical Sciences: Ahvaz, Iran, 2017. [Google Scholar] [CrossRef]
- Marco, G.; Selina, K.; Alejandro, S.M.; Roland, Z.; Brent, R. PSMA and STEAP1 Vaccines and Their Uses. WO/2021/209897 A1, 21 October 2021. [Google Scholar]
- Bordoloi, D.; Xiao, P.; Choi, H.; Ho, M.; Perales-Puchalt, A.; Khoshnejad, M.; Kim, J.J.; Humeau, L.; Srinivasan, A.; Weiner, D.B.; et al. Immunotherapy of prostate cancer using novel synthetic DNA vaccines targeting multiple tumor antigens. Genes Cancer 2021, 12, 51–64. [Google Scholar] [CrossRef]
- Lacher, M.D. Compositions and Methods for Preventing and Treating Prostate Cancer. WO/2014/165818 A2, 9 October 2014. [Google Scholar]
- Dennis, M.S.; Rubinfeld, B.; Polakis, P.; Jakobovits, A. Antibodies and Immunoconjugates and Uses Therefor. U.S. Patent 8436147 B2, 26 October 2007. [Google Scholar]
- Danila, D.C.; Szmulewitz, R.Z.; Vaishampayan, U.; Higano, C.S.; Baron, A.D.; Gilbert, H.N.; Brunstein, F.; Milojic-Blair, M.; Wang, B.; Kabbarah, O.; et al. Phase I Study of DSTP3086S, an Antibody-Drug Conjugate Targeting Six-Transmembrane Epithelial Antigen of Prostate 1, in Metastatic Castration-Resistant Prostate Cancer. J. Clin. Oncol. 2019, 37, 3518–3527. [Google Scholar] [CrossRef]
- Kelly, W.K.; Pook, D.W.; Appleman, L.J.; Waterhouse, D.M.; Horvath, L.; Edenfield, W.J.; Nobuaki, M.; Danila, D.C.; Aggarwal, R.R.; Petrylak, D.P.; et al. Phase I study of AMG 509, a STEAP1 x CD3 T-cell recruiting XmAb 2 + 1 immune therapy, in patients with metastatic castration-resistant prostate cancer (mCRPC). J. Clin. Oncol. 2021, 39, TPS183. [Google Scholar] [CrossRef]
- Lin, T.Y.; Park, J.A.; Long, A.; Guo, H.F.; Cheung, N.V. Novel potent anti-STEAP1 bispecific antibody to redirect T cells for cancer immunotherapy. J. Immunother. Cancer 2021, 9, e003114. [Google Scholar] [CrossRef]
- Lee, J.K. CAR-T Development in Subtypes of Prostate Cancer: How Mechanistic Biology Impacts Future Therapies, Fred Hutchinson Cancer Center; Fred Hutchinson Cancer Center: Seattle, WA, USA, 24 June 2017. [Google Scholar]
- Alves, P.M.; Faure, O.; Graff-Dubois, S.; Cornet, S.; Bolonakis, I.; Gross, D.A.; Miconnet, I.; Chouaib, S.; Fizazi, K.; Soria, J.C.; et al. STEAP, a prostate tumor antigen, is a target of human CD8+ T cells. Cancer Immunol. Immunother. 2006, 55, 1515–1523. [Google Scholar] [CrossRef]
- Rodeberg, D.A.; Nuss, R.A.; Elsawa, S.F.; Celis, E. Recognition of six-transmembrane epithelial antigen of the prostate-expressing tumor cells by peptide antigen-induced cytotoxic T lymphocytes. Clin. Cancer Res. 2005, 11, 4545–4552. [Google Scholar] [CrossRef] [Green Version]
- Luz Garcia-Hernandez, M.; Gray, A.; Hubby, B.; Kast, W.M. In Vivo effects of vaccination with six-transmembrane epithelial antigen of the prostate: A candidate antigen for treating prostate cancer. Cancer Res. 2007, 67, 1344–1351. [Google Scholar] [CrossRef] [Green Version]
- Rausch, S.; Schwentner, C.; Stenzl, A.; Bedke, J. mRNA vaccine CV9103 and CV9104 for the treatment of prostate cancer. Hum. Vaccin Immunother. 2014, 10, 3146–3152. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Fort, M.; Liang, L.; Moore, G.; Bernett, M.; Muchhal, U.; Osgood, T.; Yabut, R.; Kaliyaperumal, S.; Harrold, J.; et al. AMG 509, a STEAP1 x CD3 Bispecific XmAb® 2 + 1 Immune Therapy, Exhibits Avidity-Driven Binding and Preferential Killing of High STEAP1-Expressing Prostate and Ewing Sarcoma Cancer Cells. J. Immunother. Cancer 2020, 8, A430–A431. [Google Scholar] [CrossRef]
- Azumi, M.; Kobayashi, H.; Aoki, N.; Sato, K.; Kimura, S.; Kakizaki, H.; Tateno, M. Six-transmembrane epithelial antigen of the prostate as an immunotherapeutic target for renal cell and bladder cancer. J. Urol. 2010, 183, 2036–2044. [Google Scholar] [CrossRef]
- Machlenkin, A.; Paz, A.; Bar Haim, E.; Goldberger, O.; Finkel, E.; Tirosh, B.; Volovitz, I.; Vadai, E.; Lugassy, G.; Cytron, S.; et al. Human CTL epitopes prostatic acid phosphatase-3 and six-transmembrane epithelial antigen of prostate-3 as candidates for prostate cancer immunotherapy. Cancer Res. 2005, 65, 6435–6442. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Molecule(s) | Mechanism of Action | Company or Institute/ Trial or Patent | Citation |
|---|---|---|---|
| CV9104 | mRNA vaccine | CureVac AG/NCT01817738 | [63] |
| Ag85B-3 × STEAP1186-193 | Fusion protein vaccine | Tianjin University/N/A | [64] |
| Unnamed | RNA–lipoplex vaccine | Ahvaz Jundishapur University of Medical Sciences/N/A | [65] |
| Various, e.g., Ad26.hSTEAP1, MVA.PSMA.hSTEAP1 | Recombinant viral vaccine | Janssen/WO2021209897 | [66] |
| PCaA-SEV | DNA vaccine | Inovio/N/A | [67] |
| Various, e.g., TCT001, TCT002 | STEAP1 x CD3, anti-STEAP1 antibodies | Roche/WO2014165818A2 | [68] |
| MSTP2109A | Anti-STEAP1 antibody | Genentech/N/A | [69] |
| 89Zr-DFO-MSTP2109A | Antibody–radionuclide conjugate | MSKCC, Genentech/NCT01774071 | [59,60] |
| DSTP3086S (vandortuzumab vedotin) | Antibody–drug conjugate | Genentech/NCT01283373 | [70] |
| AMG 509 | Bispecific T-cell-engaging antibody | Amgen/NCT04221542 | [71] |
| BC261 | Bispecific T-cell-engaging antibody | MSKCC/N/A | [72] |
| Unnamed | STEAP1 CAR-T | FHCC/N/A | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Evans, L.; Bizzaro, C.L.; Quaglia, F.; Verrillo, C.E.; Li, L.; Stieglmaier, J.; Schiewer, M.J.; Languino, L.R.; Kelly, W.K. STEAP1–4 (Six-Transmembrane Epithelial Antigen of the Prostate 1–4) and Their Clinical Implications for Prostate Cancer. Cancers 2022, 14, 4034. https://doi.org/10.3390/cancers14164034
Xu M, Evans L, Bizzaro CL, Quaglia F, Verrillo CE, Li L, Stieglmaier J, Schiewer MJ, Languino LR, Kelly WK. STEAP1–4 (Six-Transmembrane Epithelial Antigen of the Prostate 1–4) and Their Clinical Implications for Prostate Cancer. Cancers. 2022; 14(16):4034. https://doi.org/10.3390/cancers14164034
Chicago/Turabian StyleXu, Michael, Latese Evans, Candice L. Bizzaro, Fabio Quaglia, Cecilia E. Verrillo, Li Li, Julia Stieglmaier, Matthew J. Schiewer, Lucia R. Languino, and William K. Kelly. 2022. "STEAP1–4 (Six-Transmembrane Epithelial Antigen of the Prostate 1–4) and Their Clinical Implications for Prostate Cancer" Cancers 14, no. 16: 4034. https://doi.org/10.3390/cancers14164034