
Serial Analysis of Gene Mutations and Gene Expression during First-Line Chemotherapy against Metastatic Colorectal Cancer: Identification of Potentially Actionable Targets within the Multicenter Prospective Biomarker Study REVEAL

 , , , , , ,
, , , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
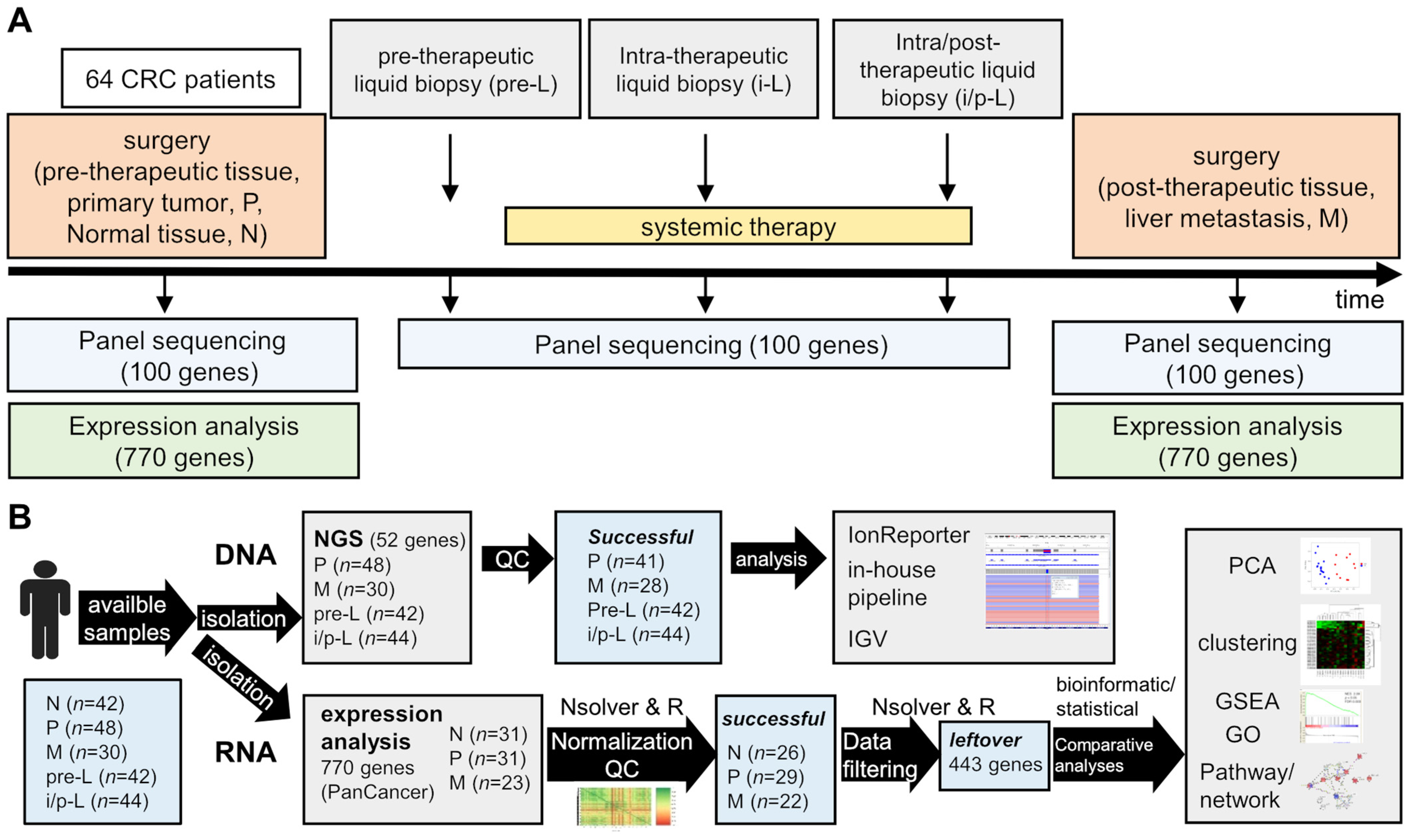
2.1. Study Design, Patients and Samples
2.2. Histopathological Samples
2.3. DNA Extraction and NGS Analyses
2.4. RNA Extraction and Expression Analysis (NanoString® nCounter Assay)
2.5. RNA Expression Quality Control (QC) and Filtering of the Data
2.6. Statistical Analysis of the Filtered RNA Expression Data
3. Results
3.1. Study Design and Population Demographics
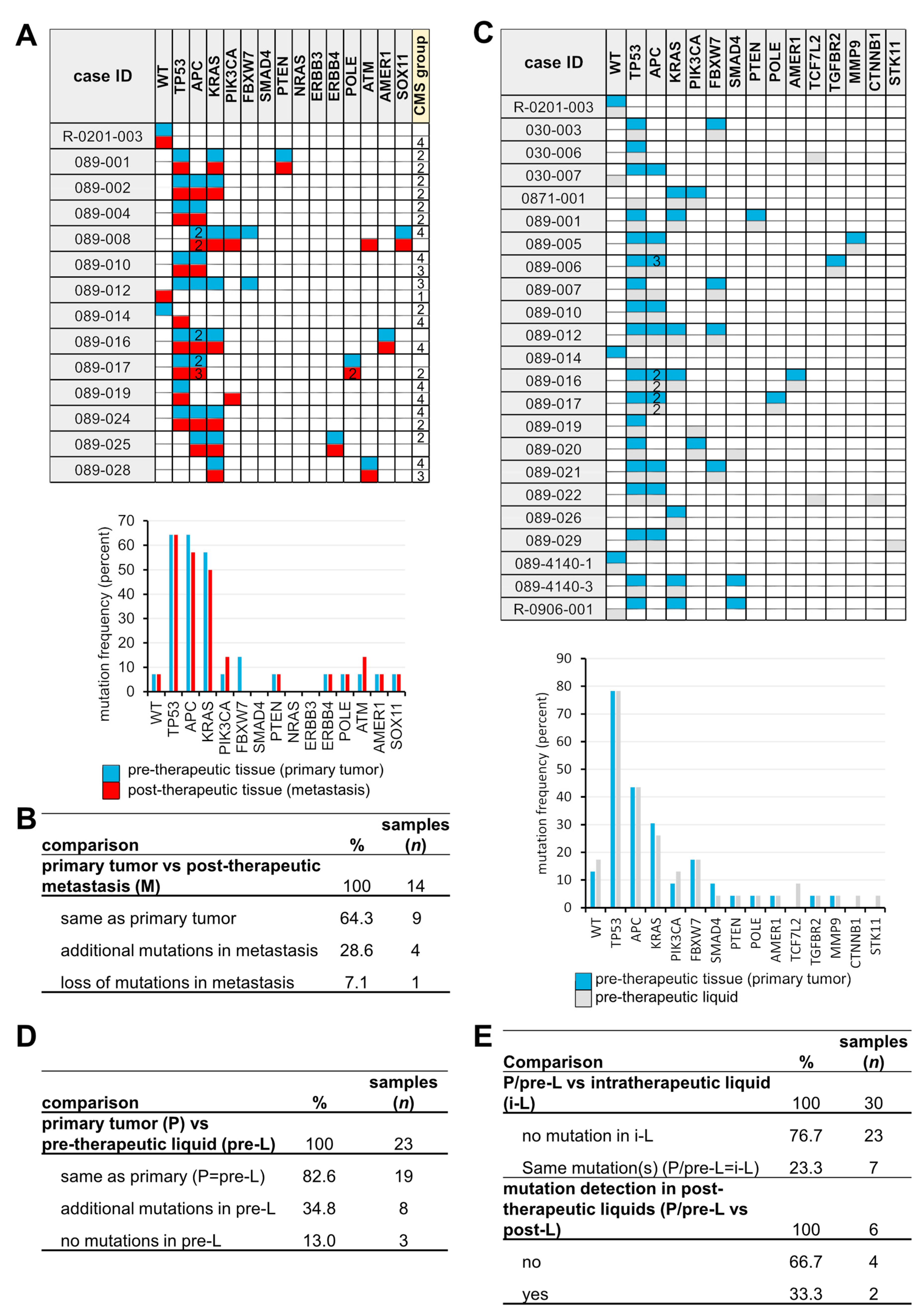
3.2. Sequential Mutation Screening
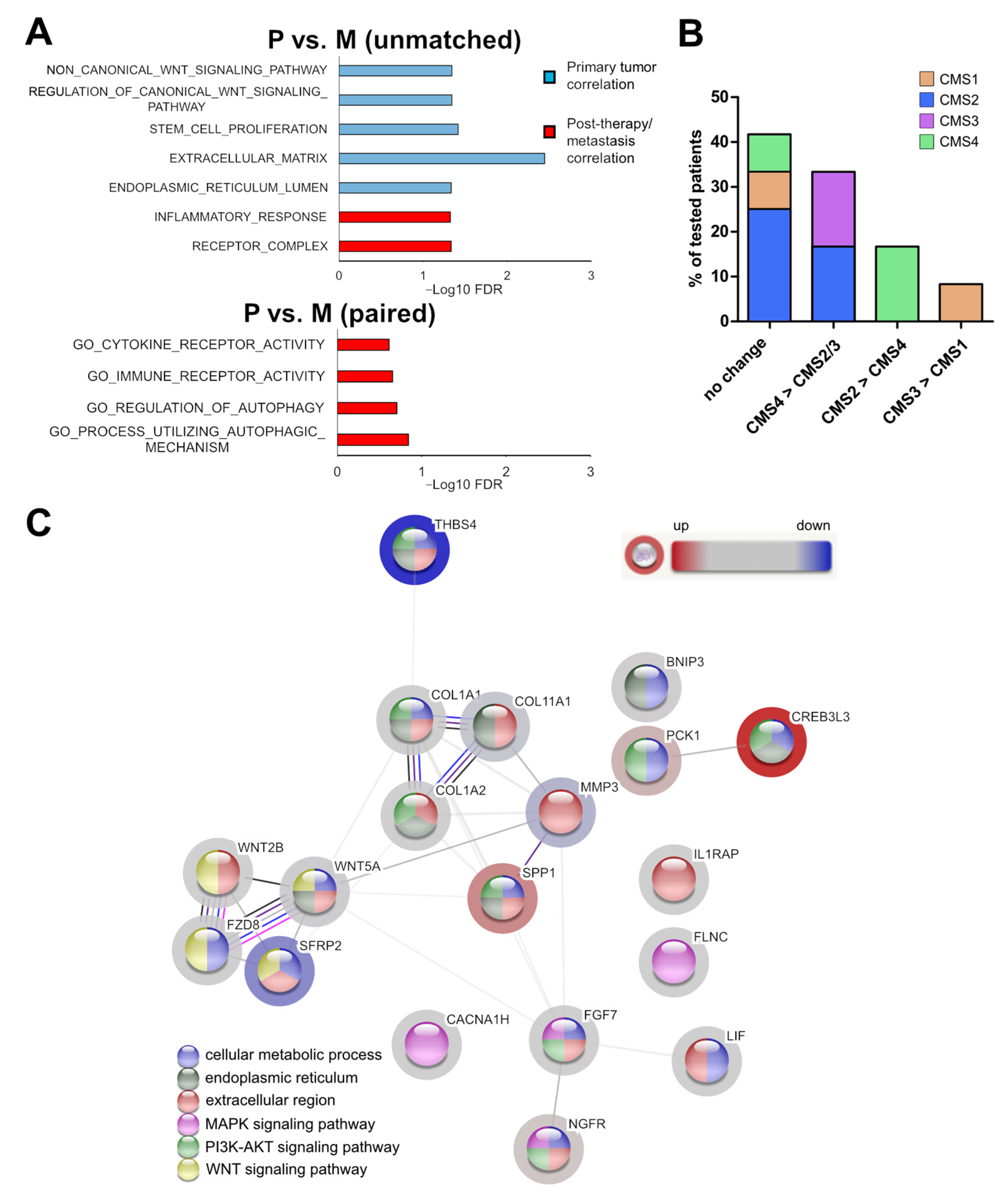
3.3. Identification of a Post-Therapeutic Liver Metastasis CRC Expression Signature
3.4. Association of the Expression Profile with Cellular Programs and Pathways
3.5. The Expression Pattern of the Post-Therapeutic Signature Genes Classifies Primary Tumor and Liver Metastasis of CRCs
3.6. Markers for the Sidedness of CRCs
3.7. Identification of Potential Biomarkers for Post-Therapeutic Metastatic CRCs
3.8. Clinical Association of the Identified Signature Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.; Renfro, L.A.; Allegra, C.J.; Andre, T.; de Gramont, A.; Schmoll, H.J.; Haller, D.G.; Alberts, S.R.; Yothers, G.; Sargent, D.J. Impact of Patient Factors on Recurrence Risk and Time Dependency of Oxaliplatin Benefit in Patients With Colon Cancer: Analysis From Modern-Era Adjuvant Studies in the Adjuvant Colon Cancer End Points (ACCENT) Database. J. Clin. Oncol. 2016, 34, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Parseghian, C.M.; Napolitano, S.; Loree, J.M.; Kopetz, S. Mechanisms of Innate and Acquired Resistance to Anti-EGFR Therapy: A Review of Current Knowledge with a Focus on Rechallenge Therapies. Clin. Cancer Res. 2019, 25, 6899–6908. [Google Scholar] [CrossRef] [PubMed]
- Stintzing, S.; Modest, D.P.; Rossius, L.; Lerch, M.M.; von Weikersthal, L.F.; Decker, T.; Kiani, A.; Vehling-Kaiser, U.; Al-Batran, S.E.; Heintges, T.; et al. FOLFIRI plus cetuximab versus FOLFIRI plus bevacizumab for metastatic colorectal cancer (FIRE-3): A post-hoc analysis of tumour dynamics in the final RAS wild-type subgroup of this randomised open-label phase 3 trial. Lancet Oncol. 2016, 17, 1426–1434. [Google Scholar] [CrossRef]
- Douillard, J.-Y.; Oliner, K.S.; Siena, S.; Tabernero, J.; Burkes, R.; Barugel, M.; Humblet, Y.; Bodoky, G.; Cunningham, D.; Jassem, J.; et al. Panitumumab–FOLFOX4 Treatment and RAS Mutations in Colorectal Cancer. N. Engl. J. Med. 2013, 369, 1023. [Google Scholar] [CrossRef] [Green Version]
- Loupakis, F.; Cremolini, C.; Masi, G.; Lonardi, S.; Zagonel, V.; Salvatore, L.; Cortesi, E.; Tomasello, G.; Ronzoni, M.; Spadi, R.; et al. Initial therapy with FOLFOXIRI and bevacizumab for metastatic colorectal cancer. N. Engl. J. Med. 2014, 371, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas, N. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Testa, U.; Castelli, G.; Pelosi, E. Genetic Alterations of Metastatic Colorectal Cancer. Biomedicines 2020, 8, 414. [Google Scholar] [CrossRef]
- Kamal, Y.; Schmit, S.L.; Hoehn, H.J.; Amos, C.I.; Frost, H.R. Transcriptomic Differences between Primary Colorectal Adenocarcinomas and Distant Metastases Reveal Metastatic Colorectal Cancer Subtypes. Cancer Res. 2019, 79, 4227–4241. [Google Scholar] [CrossRef] [Green Version]
- Sayagues, J.M.; Corchete, L.A.; Gutierrez, M.L.; Sarasquete, M.E.; Del Mar Abad, M.; Bengoechea, O.; Ferminan, E.; Anduaga, M.F.; Del Carmen, S.; Iglesias, M.; et al. Genomic characterization of liver metastases from colorectal cancer patients. Oncotarget 2016, 7, 72908–72922. [Google Scholar] [CrossRef] [Green Version]
- Bratman, S.V.; Yang, S.Y.C.; Iafolla, M.A.J.; Liu, Z.; Hansen, A.R.; Bedard, P.L.; Lheureux, S.; Spreafico, A.; Razak, A.A.; Shchegrova, S.; et al. Personalized circulating tumor DNA analysis as a predictive biomarker in solid tumor patients treated with pembrolizumab. Nat. Cancer 2020, 1, 873–881. [Google Scholar] [CrossRef]
- Goldberg, S.B.; Narayan, A.; Kole, A.J.; Decker, R.H.; Teysir, J.; Carriero, N.J.; Lee, A.; Nemati, R.; Nath, S.K.; Mane, S.M.; et al. Early Assessment of Lung Cancer Immunotherapy Response via Circulating Tumor DNA. Clin. Cancer Res. 2018, 24, 1872–1880. [Google Scholar] [CrossRef] [Green Version]
- Kruger, S.; Heinemann, V.; Ross, C.; Diehl, F.; Nagel, D.; Ormanns, S.; Liebmann, S.; Prinz-Bravin, I.; Westphalen, C.B.; Haas, M.; et al. Repeated mutKRAS ctDNA measurements represent a novel and promising tool for early response prediction and therapy monitoring in advanced pancreatic cancer. Ann. Oncol. 2018, 29, 2348–2355. [Google Scholar] [CrossRef]
- Reinert, T.; Henriksen, T.V.; Christensen, E.; Sharma, S.; Salari, R.; Sethi, H.; Knudsen, M.; Nordentoft, I.; Wu, H.T.; Tin, A.S.; et al. Analysis of Plasma Cell-Free DNA by Ultradeep Sequencing in Patients With Stages I to III Colorectal Cancer. JAMA Oncol. 2019, 5, 1124–1131. [Google Scholar] [CrossRef] [Green Version]
- Thierry, A.R.; Pastor, B.; Jiang, Z.Q.; Katsiampoura, A.D.; Parseghian, C.; Loree, J.M.; Overman, M.J.; Sanchez, C.; Messaoudi, S.E.; Ychou, M.; et al. Circulating DNA Demonstrates Convergent Evolution and Common Resistance Mechanisms during Treatment of Colorectal Cancer. Clin. Cancer Res. 2017, 23, 4578–4591. [Google Scholar] [CrossRef] [Green Version]
- Tie, J.; Wang, Y.; Tomasetti, C.; Li, L.; Springer, S.; Kinde, I.; Silliman, N.; Tacey, M.; Wong, H.L.; Christie, M.; et al. Circulating tumor DNA analysis detects minimal residual disease and predicts recurrence in patients with stage II colon cancer. Sci. Transl. Med. 2016, 8, 346ra392. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, T.V.; Tarazona, N.; Frydendahl, A.; Reinert, T.; Gimeno-Valiente, F.; Carbonell-Asins, J.A.; Sharma, S.; Renner, D.; Hafez, D.; Roda, D.; et al. Circulating Tumor DNA in Stage III Colorectal Cancer, beyond Minimal Residual Disease Detection, toward Assessment of Adjuvant Therapy Efficacy and Clinical Behavior of Recurrences. Clin. Cancer Res. 2022, 28, 507–517. [Google Scholar] [CrossRef]
- Schmiegel, W.; Scott, R.J.; Dooley, S.; Lewis, W.; Meldrum, C.J.; Pockney, P.; Draganic, B.; Smith, S.; Hewitt, C.; Philimore, H.; et al. Blood-based detection of RAS mutations to guide anti-EGFR therapy in colorectal cancer patients: Concordance of results from circulating tumor DNA and tissue-based RAS testing. Mol. Oncol. 2017, 11, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Mamlouk, S.; Childs, L.H.; Aust, D.; Heim, D.; Melching, F.; Oliveira, C.; Wolf, T.; Durek, P.; Schumacher, D.; Blaker, H.; et al. DNA copy number changes define spatial patterns of heterogeneity in colorectal cancer. Nat. Commun. 2017, 8, 14093. [Google Scholar] [CrossRef]
- von Baumgarten, L.; Kumbrink, J.; Jung, A.; Reischer, A.; Flach, M.; Liebmann, S.; Metzeler, K.H.; Holch, J.W.; Niyazi, M.; Thon, N.; et al. Therapeutic management of neuro-oncologic patients-potential relevance of CSF liquid biopsy. Theranostics 2020, 10, 856–866. [Google Scholar] [CrossRef]
- Childs, L.H.; Mamlouk, S.; Brandt, J.; Sers, C.; Leser, U. SoFIA: A data integration framework for annotating high-throughput datasets. Bioinformatics 2016, 32, 2590–2597. [Google Scholar] [CrossRef] [Green Version]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.R.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Jang, W.; et al. ClinVar: Improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 2018, 46, D1062–D1067. [Google Scholar] [CrossRef] [Green Version]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [Green Version]
- Kopanos, C.; Tsiolkas, V.; Kouris, A.; Chapple, C.E.; Albarca Aguilera, M.; Meyer, R.; Massouras, A. VarSome: The human genomic variant search engine. Bioinformatics 2019, 35, 1978–1980. [Google Scholar] [CrossRef]
- Kohl, M. MKmisc: Miscellaneous Functions from M. Kohl; R Package Version 1.6. 2019. Available online: https://cran.r-project.org/package=MKmisc (accessed on 24 May 2022).
- RStudio Team. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 1 March 2020).
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Ru, B.; Tong, Y.; Zhang, J. MR4Cancer: A web server prioritizing master regulators for cancer. Bioinformatics 2019, 35, 636–642. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Statist. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eide, P.W.; Bruun, J.; Lothe, R.A.; Sveen, A. CMScaller: An R package for consensus molecular subtyping of colorectal cancer pre-clinical models. Sci. Rep. 2017, 7, 16618. [Google Scholar] [CrossRef] [Green Version]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Holch, J.W.; Ricard, I.; Stintzing, S.; Modest, D.P.; Heinemann, V. The relevance of primary tumour location in patients with metastatic colorectal cancer: A meta-analysis of first-line clinical trials. Eur. J. Cancer 2017, 70, 87–98. [Google Scholar] [CrossRef]
- Stintzing, S.; Tejpar, S.; Gibbs, P.; Thiebach, L.; Lenz, H.J. Understanding the role of primary tumour localisation in colorectal cancer treatment and outcomes. Eur. J. Cancer 2017, 84, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Meno, C.; Shimono, A.; Saijoh, Y.; Yashiro, K.; Mochida, K.; Ohishi, S.; Noji, S.; Kondoh, H.; Hamada, H. lefty-1 is required for left-right determination as a regulator of lefty-2 and nodal. Cell 1998, 94, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Paschke, S.; Jafarov, S.; Staib, L.; Kreuser, E.D.; Maulbecker-Armstrong, C.; Roitman, M.; Holm, T.; Harris, C.C.; Link, K.H.; Kornmann, M. Are Colon and Rectal Cancer Two Different Tumor Entities? A Proposal to Abandon the Term Colorectal Cancer. Int. J. Mol. Sci. 2018, 19, 2577. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337.e10. [Google Scholar] [CrossRef] [Green Version]
- Stintzing, S.; Wirapati, P.; Lenz, H.J.; Neureiter, D.; Fischer von Weikersthal, L.; Decker, T.; Kiani, A.; Kaiser, F.; Al-Batran, S.; Heintges, T.; et al. Consensus molecular subgroups (CMS) of colorectal cancer (CRC) and first-line efficacy of FOLFIRI plus cetuximab or bevacizumab in the FIRE3 (AIO KRK-0306) trial. Ann. Oncol. 2019, 30, 1796–1803. [Google Scholar] [CrossRef] [Green Version]
- Hao, C.; Cui, Y.; Owen, S.; Li, W.; Cheng, S.; Jiang, W.G. Human osteopontin: Potential clinical applications in cancer (Review). Int. J. Mol. Med. 2017, 39, 1327–1337. [Google Scholar] [CrossRef] [Green Version]
- Vatandoost, N.; Ghanbari, J.; Mojaver, M.; Avan, A.; Ghayour-Mobarhan, M.; Nedaeinia, R.; Salehi, R. Early detection of colorectal cancer: From conventional methods to novel biomarkers. J. Cancer Res. Clin. Oncol. 2016, 142, 341–351. [Google Scholar] [CrossRef]
- Jung, A.; Kirchner, T. Liquid Biopsy in Tumor Genetic Diagnosis. Dtsch. Arztebl. Int. 2018, 115, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Brodt, P. Role of the Microenvironment in Liver Metastasis: From Pre- to Prometastatic Niches. Clin. Cancer Res. 2016, 22, 5971–5982. [Google Scholar] [CrossRef] [Green Version]
- Woolston, A.; Khan, K.; Spain, G.; Barber, L.J.; Griffiths, B.; Gonzalez-Exposito, R.; Hornsteiner, L.; Punta, M.; Patil, Y.; Newey, A.; et al. Genomic and Transcriptomic Determinants of Therapy Resistance and Immune Landscape Evolution during Anti-EGFR Treatment in Colorectal Cancer. Cancer Cell 2019, 36, 35–50.e9. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Fritzmann, J.; Morkel, M.; Besser, D.; Budczies, J.; Kosel, F.; Brembeck, F.H.; Stein, U.; Fichtner, I.; Schlag, P.M.; Birchmeier, W. A colorectal cancer expression profile that includes transforming growth factor beta inhibitor BAMBI predicts metastatic potential. Gastroenterology 2009, 137, 165–175. [Google Scholar] [CrossRef]
- Mooi, J.K.; Wirapati, P.; Asher, R.; Lee, C.K.; Savas, P.; Price, T.J.; Townsend, A.; Hardingham, J.; Buchanan, D.; Williams, D.; et al. The prognostic impact of consensus molecular subtypes (CMS) and its predictive effects for bevacizumab benefit in metastatic colorectal cancer: Molecular analysis of the AGITG MAX clinical trial. Ann. Oncol. 2018, 29, 2240–2246. [Google Scholar] [CrossRef]
- Lenz, H.J.; Ou, F.S.; Venook, A.P.; Hochster, H.S.; Niedzwiecki, D.; Goldberg, R.M.; Mayer, R.J.; Bertagnolli, M.M.; Blanke, C.D.; Zemla, T.; et al. Impact of Consensus Molecular Subtype on Survival in Patients With Metastatic Colorectal Cancer: Results From CALGB/SWOG 80405 (Alliance). J. Clin. Oncol. 2019, 37, 1876–1885. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reynies, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- De Smedt, L.; Palmans, S.; Andel, D.; Govaere, O.; Boeckx, B.; Smeets, D.; Galle, E.; Wouters, J.; Barras, D.; Suffiotti, M.; et al. Expression profiling of budding cells in colorectal cancer reveals an EMT-like phenotype and molecular subtype switching. Br. J. Cancer 2017, 116, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Trumpi, K.; Ubink, I.; Trinh, A.; Djafarihamedani, M.; Jongen, J.M.; Govaert, K.M.; Elias, S.G.; van Hooff, S.R.; Medema, J.P.; Lacle, M.M.; et al. Neoadjuvant chemotherapy affects molecular classification of colorectal tumors. Oncogenesis 2017, 6, e357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Loon, K.; Huijbers, E.J.M.; Griffioen, A.W. Secreted frizzled-related protein 2: A key player in noncanonical Wnt signaling and tumor angiogenesis. Cancer Metastasis Rev. 2020, 40, 191–203. [Google Scholar] [CrossRef]
- Vincent, K.M.; Postovit, L.M. A pan-cancer analysis of secreted Frizzled-related proteins: Re-examining their proposed tumour suppressive function. Sci. Rep. 2017, 7, 42719. [Google Scholar] [CrossRef]
- Ng, L.; Wan, T.M.; Lam, C.S.; Chow, A.K.; Wong, S.K.; Man, J.H.; Li, H.S.; Cheng, N.S.; Pak, R.C.; Cheung, A.H.; et al. Post-operative plasma osteopontin predicts distant metastasis in human colorectal cancer. PLoS ONE 2015, 10, e0126219. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Gong, K.; Zhang, X.; Wu, S.; Cui, Y.; Qian, B.Z. Osteopontin as a multifaceted driver of bone metastasis and drug resistance. Pharmacol. Res. 2019, 144, 235–244. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | N | % |
|---|---|---|
| Age | ||
| Age-median | 62 (range 20–87) | |
| Sex | ||
| Male | 38 | 59.4 |
| Female | 26 | 40.6 |
| Performance status | ||
| ECOG 0–1 | 58 | 90.6 |
| ECOG 2–3 | 2 | 3.1 |
| NA | 4 | 6.3 |
| Primary tumor sidedness | ||
| Right-sided | 13 | 20.3 |
| Left-sided | 49 | 76.6 |
| NA | 2 | 3.1 |
| T-stage of primary | ||
| T1-2 | 13 | 20.3 |
| T3-4 | 39 | 60.9 |
| NA | 12 | 18.8 |
| N-stage of primary | ||
| N0 | 7 | 10.9 |
| N1 | 9 | 14.1 |
| N2 | 11 | 17.2 |
| NA | 22 | 57.8 |
| Grading of primary | ||
| G1-2 | 44 | 68.8 |
| G3 | 8 | 12.5 |
| NA | 12 | 18.8 |
| Metastasis | ||
| synchronous | 48 | 75 |
| metachronous | 16 | 25 |
| Number of metastatic sites | ||
| 1 site | 25 | 39.1 |
| ≥2 sites | 39 | 60.9 |
| Chemotherapy | ||
| FOLFOXIRI | 6 | 9.4 |
| plus Bevacizumab | 3 | 4.7 |
| plus Panitumumab | 3 | 4.7 |
| FOLFOX | 22 | 34.4 |
| plus Panitumumab | 2 | 3.1 |
| plus Bevacizumab | 11 | 17.2 |
| plus Cetuximab | 2 | 3.1 |
| FOLFIRI | 21 | 32.8 |
| plus Bevacizumab | 7 | 10.9 |
| plus Cetuximab | 9 | 14.1 |
| Capecitabine | 4 | 6.3 |
| plus Irinotecan & Bevacizumab | 1 | 1.6 |
| plus Bevacizumab | 2 | 3.1 |
| Cetuximab mono | 1 | 1.6 |
| RAS mutation | ||
| no | 37 | 57.8 |
| yes | 24 | 37.5 |
| NA | 3 | 4.7 |
| BRAF mutation | ||
| no | 54 | 84.4 |
| yes | 3 | 4.7 |
| NA | 7 | 10.9 |
| total | 64 | 100 |
| Gene | DEG | Log2 FC | Avg Expr | p Value | Padj | FC | %Change | Program/Pathway/Function |
|---|---|---|---|---|---|---|---|---|
| SFRP2 | A | −3.46 | 5.81 | 1.77 × 10−5 | 0.0016 | 0.09 | −90.91 | EMT/MET/WNT |
| B | −4.33 | 4.94 | 5.85 × 10−6 | 0.00065 | 0.05 | −95.03 | ||
| THBS4 | A | −3.44 | 4.55 | 3.98 × 10−7 | 8.81 × 10−5 | 0.09 | −90.79 | ER stress, tumor suppressor CRC |
| B | −4.52 | 4.05 | 7.26 × 10−7 | 0.00016 | 0.04 | −95.64 | ||
| MMP3 | A | −2.94 | 4.24 | 6.56 × 10−7 | 9.69 × 10−5 | 0.13 | −86.97 | ECM modulating/related |
| B | −4.11 | 3.76 | 1.99 × 10−6 | 0.00029 | 0.06 | −94.21 | ||
| COL11A1 | A | −1.84 | 5.89 | 0.00012 | 0.0065 | 0.28 | −72.07 | ECM modulating/related |
| B | −2.17 | 5.73 | 2.21 × 10−5 | 0.002 | 0.22 | −77.78 | ||
| WNT5A | A | −1.53 | 5.66 | 1.28 × 10−5 | 0.0014 | 0.35 | −65.37 | EMT/MET/WNT |
| B | −1.97 | 5.41 | 0.00022 | 0.0098 | 0.26 | −74.47 | ||
| FLNC | A | −1.33 | 5.90 | 9.01 × 10−5 | 0.0065 | 0.40 | −60.22 | ECM modulating/related |
| B | −1.85 | 5.73 | 5.75 × 10−5 | 0.0042 | 0.28 | −72.26 | ||
| WNT2B | A | −1.17 | 3.93 | 0.00022 | 0.011 | 0.44 | −55.56 | EMT/MET/WNT |
| B | −1.24 | 3.61 | 7.2 × 10−5 | 0.0046 | 0.42 | −57.66 | ||
| CACNA1H | A | −1.05 | 5.80 | 0.00011 | 0.0065 | 0.48 | −51.70 | ER stress, inh. of proliferation |
| B | −1.32 | 5.66 | 0.0013 | 0.038 | 0.40 | −59.95 | ||
| FZD8 | A | −0.98 | 4.53 | 0.00071 | 0.031 | 0.51 | −49.30 | EMT/MET/WNT |
| B | −1.62 | 4.30 | 0.00022 | 0.0098 | 0.33 | −67.47 | ||
| FGF7 | B | −1.65 | 4.00 | 0.00067 | 0.025 | 0.32 | −68.14 | ECM modulating/related |
| COL1A1 | B | −1.07 | 11.31 | 0.00093 | 0.032 | 0.48 | −52.37 | ECM modulating/related |
| COL1A2 | B | −1.04 | 8.57 | 0.0017 | 0.047 | 0.49 | −51.37 | ECM modulating/related |
| LIF | B | −0.75 | 6.01 | 0.0013 | 0.038 | 0.59 | −40.54 | NOTCH inihibition |
| IL1RAP | A | 0.61 | 4.78 | 0.0012 | 0.042 | 1.53 | 52.63 | Oncogenic signaling |
| BNIP3 | B | 1.41 | 5.32 | 0.00038 | 0.015 | 2.66 | 165.74 | ER stress, apoptosis, autophagy |
| NGFR | A | 1.45 | 4.98 | 0.0012 | 0.042 | 2.73 | 173.21 | Ambivalent, tumor suppressor CRC |
| PCK1 | A | 1.6 | 4.37 | 0.00092 | 0.037 | 3.03 | 203.14 | NOTCH, metabolism |
| SPP1/OPN | B | 1.76 | 9.70 | 0.00021 | 0.0098 | 3.39 | 238.70 | EMT/MET/WNT |
| CREB3L3 | A | 2.16 | 2.13 | 3.33 × 10−8 | 1.48 × 10−5 | 4.47 | 346.91 | ER stress, transcription factor |
| B | 2.78 | 2.54 | 5.23 × 10−10 | 2.32 × 10−7 | 6.87 | 586.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumbrink, J.; Bohlmann, L.; Mamlouk, S.; Redmer, T.; Peilstöcker, D.; Li, P.; Lorenzen, S.; Algül, H.; Kasper, S.; Hempel, D.; et al. Serial Analysis of Gene Mutations and Gene Expression during First-Line Chemotherapy against Metastatic Colorectal Cancer: Identification of Potentially Actionable Targets within the Multicenter Prospective Biomarker Study REVEAL. Cancers 2022, 14, 3631. https://doi.org/10.3390/cancers14153631
Kumbrink J, Bohlmann L, Mamlouk S, Redmer T, Peilstöcker D, Li P, Lorenzen S, Algül H, Kasper S, Hempel D, et al. Serial Analysis of Gene Mutations and Gene Expression during First-Line Chemotherapy against Metastatic Colorectal Cancer: Identification of Potentially Actionable Targets within the Multicenter Prospective Biomarker Study REVEAL. Cancers. 2022; 14(15):3631. https://doi.org/10.3390/cancers14153631
Chicago/Turabian StyleKumbrink, Jörg, Lisa Bohlmann, Soulafa Mamlouk, Torben Redmer, Daniela Peilstöcker, Pan Li, Sylvie Lorenzen, Hana Algül, Stefan Kasper, Dirk Hempel, and et al. 2022. "Serial Analysis of Gene Mutations and Gene Expression during First-Line Chemotherapy against Metastatic Colorectal Cancer: Identification of Potentially Actionable Targets within the Multicenter Prospective Biomarker Study REVEAL" Cancers 14, no. 15: 3631. https://doi.org/10.3390/cancers14153631