
Triple-Negative Breast Cancer circRNAome Reveals Hsa_circ_0072309 as a Potential Risk Biomarker

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Consent to Participate
2.2. TNBC Sample Series, RNA-Seq Data, circRNA and mRNA Mapping, and Data Analysis
2.3. CircRNA-RBP Prediction
2.4. Functional Enrichment Analysis
2.5. Biological Samples
2.6. Total RNA Isolation and RT-qPCR
2.7. Statistical Analysis
3. Results
3.1. CircRNAs and Their Host Genes Are Predominantly Downregulated in TNBC
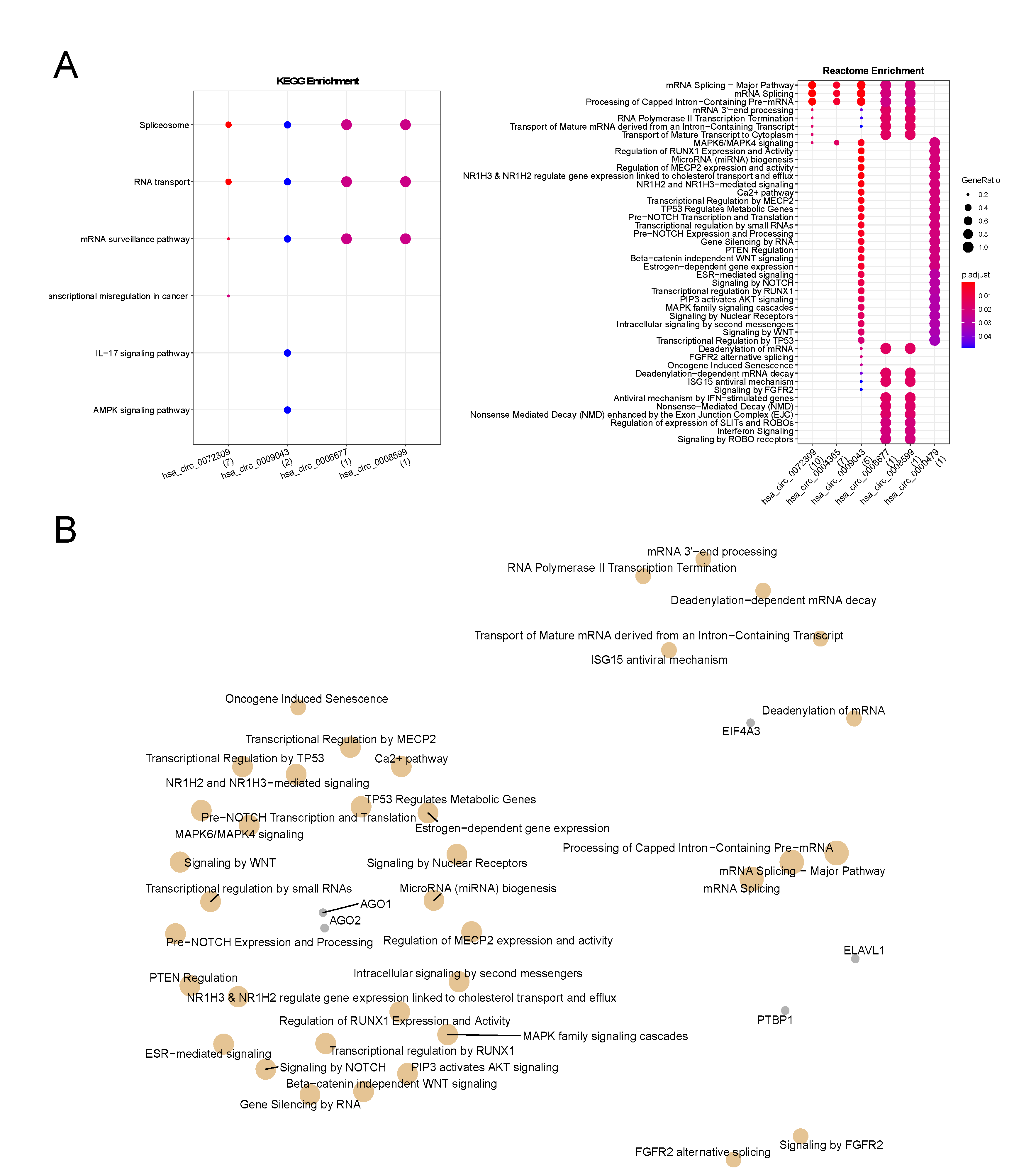
3.2. CircRNAs Interact with RNA Binding Proteins and Enrich to Cancer-Related Pathways
3.3. TNBC Patients with BRCA1 Mutations Have a Distinct circRNA Profile When Compared to Wildtype Ones
3.4. Hsa_circ_0072309 Is a Potential Biomarker of TNBC and May Be Involved in Cancer-Related Gene Regulation Signaling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids Are Single-Stranded Covalently Closed Circular RNA Molecules Existing as Highly Base-Paired Rod-like Structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Smid, M.; Wilting, S.M.; Uhr, K.; Rodríguez-González, F.G.; de Weerd, V.; Prager-Van der Smissen, W.J.C.; van der Vlugt-Daane, M.; van Galen, A.; Nik-Zainal, S.; Butler, A.; et al. The Circular RNome of Primary Breast Cancer. Genome Res. 2019, 29, 356–366. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sotiriou, C.; Pusztai, L. Gene-Expression Signatures in Breast Cancer. N. Engl. J. Med. 2009, 360, 790–800. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Aggarwal, R. An Overview of Triple-Negative Breast Cancer. Arch. Gynecol. Obstet. 2016, 293, 247–269. [Google Scholar] [CrossRef]
- Ju, H.-Q.; Zhao, Q.; Wang, F.; Lan, P.; Wang, Z.; Zuo, Z.-X.; Wu, Q.-N.; Fan, X.-J.; Mo, H.-Y.; Chen, L.; et al. A CircRNA Signature Predicts Postoperative Recurrence in Stage II/III Colon Cancer. EMBO Mol. Med. 2019, 11, e10168. [Google Scholar] [CrossRef]
- Yu, C.-Y.; Li, T.-C.; Wu, Y.-Y.; Yeh, C.-H.; Chiang, W.; Chuang, C.-Y.; Kuo, H.-C. The Circular RNA CircBIRC6 Participates in the Molecular Circuitry Controlling Human Pluripotency. Nat. Commun. 2017, 8, 1149. [Google Scholar] [CrossRef]
- Du, W.W.; Fang, L.; Yang, W.; Wu, N.; Awan, F.M.; Yang, Z.; Yang, B.B. Induction of Tumor Apoptosis through a Circular RNA Enhancing Foxo3 Activity. Cell Death Differ. 2017, 24, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Bachmayr-Heyda, A.; Reiner, A.T.; Auer, K.; Sukhbaatar, N.; Aust, S.; Bachleitner-Hofmann, T.; Mesteri, I.; Grunt, T.W.; Zeillinger, R.; Pils, D. Correlation of Circular RNA Abundance with Proliferation—Exemplified with Colorectal and Ovarian Cancer, Idiopathic Lung Fibrosis and Normal Human Tissues. Sci. Rep. 2015, 5, 8057. [Google Scholar] [CrossRef] [PubMed]
- Lü, L.; Sun, J.; Shi, P.; Kong, W.; Xu, K.; He, B.; Zhang, S.; Wang, J. Identification of Circular RNAs as a Promising New Class of Diagnostic Biomarkers for Human Breast Cancer. Oncotarget 2017, 8, 44096–44107. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA Profiling Reveals an Abundant CircHIPK3 That Regulates Cell Growth by Sponging Multiple MiRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA Circles Function as Efficient MicroRNA Sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Liu, P.; Xie, X.; Zhou, Y.; Liao, Q.; Xiong, W.; Li, X.; Li, G.; Zeng, Z.; Tang, H. CircGFRA1 and GFRA1 Act as CeRNAs in Triple Negative Breast Cancer by Regulating MiR-34a. J. Exp. Clin. Cancer Res. 2017, 36, 145. [Google Scholar] [CrossRef] [Green Version]
- Zeng, K.; He, B.; Yang, B.B.; Xu, T.; Chen, X.; Xu, M.; Liu, X.; Sun, H.; Pan, Y.; Wang, S. The Pro-Metastasis Effect of CircANKS1B in Breast Cancer. Mol. Cancer 2018, 17, 160. [Google Scholar] [CrossRef]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded Identification and Characterization of Mammalian Circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Denzler, R.; Agarwal, V.; Stefano, J.; Bartel, D.P.; Stoffel, M. Assessing the CeRNA Hypothesis with Quantitative Measurements of MiRNA and Target Abundance. Mol. Cell 2014, 54, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Abdelmohsen, K.; Panda, A.C.; Munk, R.; Grammatikakis, I.; Dudekula, D.B.; De, S.; Kim, J.; Noh, J.H.; Kim, K.M.; Martindale, J.L.; et al. Identification of HuR Target Circular RNAs Uncovers Suppression of PABPN1 Translation by CircPABPN1. RNA Biol. 2017, 14, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okholm, T.L.H.; Sathe, S.; Park, S.S.; Kamstrup, A.B.; Rasmussen, A.M.; Shankar, A.; Chua, Z.M.; Fristrup, N.; Nielsen, M.M.; Vang, S.; et al. Transcriptome-Wide Profiles of Circular RNA and RNA-Binding Protein Interactions Reveal Effects on Circular RNA Biogenesis and Cancer Pathway Expression. Genome Med. 2020, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Brianese, R.C.; de Mello Nakamura, K.D.; Dos Santos Ramos de Almeida , F.G.; Ramalho, R.F.; de Figueiredo Barros, B.D.; E Ferreira, E.N.; da Cruz Formiga , M.N.; de Andrade, V.P.; Cordeiro de Lima, V.C.; Carraro, D.M. BRCA1 Deficiency Is a Recurrent Event in Early-Onset Triple-Negative Breast Cancer: A Comprehensive Analysis of Germline Mutations and Somatic Promoter Methylation. Breast Cancer Res. Treat. 2018, 167, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-O.; Dong, R.; Zhang, Y.; Zhang, J.-L.; Luo, Z.; Zhang, J.; Chen, L.-L.; Yang, L. Diverse Alternative Back-Splicing and Alternative Splicing Landscape of Circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An Efficient and Unbiased Algorithm for de Novo Circular RNA Identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Metge, F.; Dieterich, C. Specific Identification and Quantification of Circular RNAs from Sequencing Data. Bioinformatics 2016, 32, 1094–1096. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon Provides Fast and Bias-Aware Quantification of Transcript Expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A Web Tool for Exploring Circular RNAs and Their Interacting Proteins and MicroRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. Omics A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; He, Q.-Y. ReactomePA: An R/Bioconductor Package for Reactome Pathway Analysis and Visualization. Mol. Biosyst. 2016, 12, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Janowski, B.A.; Huffman, K.E.; Schwartz, J.C.; Ram, R.; Nordsell, R.; Shames, D.S.; Minna, J.D.; Corey, D.R. Involvement of AGO1 and AGO2 in Mammalian Transcriptional Silencing. Nat. Struct. Mol. Biol. 2006, 13, 787–792. [Google Scholar] [CrossRef]
- Zhan, X.; Yan, C.; Zhang, X.; Lei, J.; Shi, Y. Structure of a Human Catalytic Step I Spliceosome. Science 2018, 359, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Doller, A.; Akool, E.-S.; Huwiler, A.; Müller, R.; Radeke, H.H.; Pfeilschifter, J.; Eberhardt, W. Posttranslational Modification of the AU-Rich Element Binding Protein HuR by Protein Kinase Cδ Elicits Angiotensin II-Induced Stabilization and Nuclear Export of Cyclooxygenase 2 MRNA. Mol. Cell. Biol. 2008, 28, 2608–2625. [Google Scholar] [CrossRef] [Green Version]
- Rossow, K.L.; Janknecht, R. The Ewing’s Sarcoma Gene Product Functions as a Transcriptional Activator. Cancer Res 2001, 61, 2690–2695. [Google Scholar]
- Erkizan, H.V.; Uversky, V.N.; Toretsky, J.A. Oncogenic Partnerships: EWS-FLI1 Protein Interactions Initiate Key Pathways of Ewing’s Sarcoma. Clin. Cancer Res. 2010, 16, 4077–4083. [Google Scholar] [CrossRef] [Green Version]
- Kenny, P.J.; Zhou, H.; Kim, M.; Skariah, G.; Khetani, R.S.; Drnevich, J.; Arcila, M.L.; Kosik, K.S.; Ceman, S. MOV10 and FMRP Regulate AGO2 Association with MicroRNA Recognition Elements. Cell Rep. 2014, 9, 1729–1741. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Bellini, M.; Ceman, S. Fragile X Mental Retardation Protein FMRP Binds MRNAs in the Nucleus. Mol. Cell. Biol. 2009, 29, 214–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Reed, R. FUS Functions in Coupling Transcription to Splicing by Mediating an Interaction between RNAP II and U1 SnRNP. Proc. Natl. Acad. Sci. USA 2015, 112, 8608–8613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noubissi, F.K.; Elcheva, I.; Bhatia, N.; Shakoori, A.; Ougolkov, A.; Liu, J.; Minamoto, T.; Ross, J.; Fuchs, S.Y.; Spiegelman, V.S. CRD-BP Mediates Stabilization of ΒTrCP1 and c-Myc MRNA in Response to β-Catenin Signalling. Nature 2006, 441, 898–901. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-Methyladenosine by IGF2BP Proteins Enhances MRNA Stability and Translation. Nat. Cell. Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J. Sequential Dimerization of Human Zipcode-Binding Protein IMP1 on RNA: A Cooperative Mechanism Providing RNP Stability. Nucleic Acids Res. 2004, 32, 4368–4376. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Jing, W.; Lei, X.-X.; Feng, C.; Peng, S.; Boris-Lawrie, K.; Huang, Y. Evidence That Lin28 Stimulates Translation by Recruiting RNA Helicase A to Polysomes. Nucleic Acids Res. 2011, 39, 3724–3734. [Google Scholar] [CrossRef]
- Heo, I.; Joo, C.; Kim, Y.-K.; Ha, M.; Yoon, M.-J.; Cho, J.; Yeom, K.-H.; Han, J.; Kim, V.N. TUT4 in Concert with Lin28 Suppresses MicroRNA Biogenesis through Pre-MicroRNA Uridylation. Cell 2009, 138, 696–708. [Google Scholar] [CrossRef] [Green Version]
- Clerte, C.; Hall, K.B. Characterization of Multimeric Complexes Formed by the Human PTB1 Protein on RNA. RNA 2006, 12, 457–475. [Google Scholar] [CrossRef] [Green Version]
- Kafasla, P.; Lin, H.; Curry, S.; Jackson, R.J. Activation of Picornaviral IRESs by PTB Shows Differential Dependence on Each PTB RNA-Binding Domain. RNA 2011, 17, 1120–1131. [Google Scholar] [CrossRef] [Green Version]
- David, C.J.; Boyne, A.R.; Millhouse, S.R.; Manley, J.L. The RNA Polymerase II C-Terminal Domain Promotes Splicing Activation through Recruitment of a U2AF65–Prp19 Complex. Genes Dev. 2011, 25, 972–983. [Google Scholar] [CrossRef] [Green Version]
- Millevoi, S.; Loulergue, C.; Dettwiler, S.; Karaa, S.Z.; Keller, W.; Antoniou, M.; Vagner, S. An Interaction between U2AF 65 and CF Im Links the Splicing and 3′ End Processing Machineries. EMBO J. 2006, 25, 4854–4864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dube, U.; Del-Aguila, J.L.; Li, Z.; Budde, J.P.; Jiang, S.; Hsu, S.; Ibanez, L.; Fernandez, M.V.; Farias, F.; Norton, J.; et al. An Atlas of Cortical Circular RNA Expression in Alzheimer Disease Brains Demonstrates Clinical and Pathological Associations. Nat. Neurosci. 2019, 22, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Elia, L.; Quintavalle, M.; Condorelli, G. Circular RNAs and Heart Failure: New Players for an Old Disease. Cardiovasc. Res. 2017, 113, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Stoll, L.; Sobel, J.; Rodriguez-Trejo, A.; Guay, C.; Lee, K.; Venø, M.T.; Kjems, J.; Laybutt, D.R.; Regazzi, R. Circular RNAs as Novel Regulators of β-Cell Functions in Normal and Disease Conditions. Mol. Metab. 2018, 9, 69–83. [Google Scholar] [CrossRef]
- Vidal, A.F.; Ribeiro-dos-Santos, A.M.; Vinasco-Sandoval, T.; Magalhães, L.; Pinto, P.; Anaissi, A.K.M.; Demachki, S.; de Assumpção, P.P.; dos Santos, S.E.B.; Ribeiro-dos-Santos, Â. The Comprehensive Expression Analysis of Circular RNAs in Gastric Cancer and Its Association with Field Cancerization. Sci. Rep. 2017, 7, 14551. [Google Scholar] [CrossRef] [Green Version]
- Wilusz, J.E. A 360° View of Circular RNAs: From Biogenesis to Functions. Wiley Interdiscip. Rev. RNA 2018, 9, e1478. [Google Scholar] [CrossRef] [Green Version]
- Nair, A.A.; Niu, N.; Tang, X.; Thompson, K.J.; Wang, L.; Kocher, J.-P.; Subramanian, S.; Kalari, K.R. Circular RNAs and Their Associations with Breast Cancer Subtypes. Oncotarget 2016, 7, 80967–80979. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Wang, H.; Ye, L.; Shi, X.; Yan, K.; Lin, K.; Huang, Q.; Li, B.; Lin, Q.; Zhu, L.; et al. Hsa_circ_0000479 as a Novel Diagnostic Biomarker of Systemic Lupus Erythematosus. Front. Immunol. 2019, 10, 2281. [Google Scholar] [CrossRef] [Green Version]
- Ye, F.; Tang, Q.L.; Ma, F.; Cai, L.; Chen, M.; Ran, X.X.; Wang, X.Y.; Jiang, X.F. Analysis of the Circular RNA Transcriptome in the Grade 3 Endometrial Cancer. Cancer Manag. Res. 2019, 11, 6215–6227. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Huang, Q.-Y.; Sun, Y.; Wu, S. High-Throughput Data Reveals Novel Circular RNAs via Competitive Endogenous RNA Networks Associated with Human Intracranial Aneurysms. Med. Sci. Monit. 2019, 25, 4819–4830. [Google Scholar] [CrossRef]
- Chen, T.; Shao, S.; Li, W.; Liu, Y.; Cao, Y. The Circular RNA Hsa-Circ-0072309 Plays Anti-Tumour Roles by Sponging MiR-100 through the Deactivation of PI3K/AKT and MTOR Pathways in the Renal Carcinoma Cell Lines. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3638–3648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Zheng, M.; Wang, H. Circular RNA Hsa_circ_0072309 Inhibits Proliferation and Invasion of Breast Cancer Cells via Targeting MiR-492. Cancer Manag. Res. 2019, 11, 1033–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive Analysis of Normal Adjacent to Tumor Transcriptomes. Nat. Commun. 2017, 8, 1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.; Zhao, W.; Wang, J.; Seifer, B.J.; Ye, C.; Chen, Y.; Jia, Y.; Chen, C.; Shen, J.; Wang, L.; et al. Oncogenic Mechanisms of Lin28 in Breast Cancer: New Functions and Therapeutic Opportunities. Oncotarget 2017, 8, 25721–25735. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Wildtype | Wildtype, BRCA Hypermethylated | BRCA1 Mutation | |||
|---|---|---|---|---|---|---|
| n = 18 | n = 10 | p-Value * | n = 9 | p-Value * | ||
| Age of onset, mean (SD) | 41.8 (6.1) | 38.5 (7.6) | 0.3 | 36.5 (5.6) | 0.16 | |
| nº (%) | nº (%) | nº (%) | ||||
| TNM classification, T | T1 | 3 (16.7) | - | 0.11 | 1 (11.1) | 0.94 |
| T2 | 8 (44.4) | 9 (90) | 5 (55.6) | |||
| T3 | 3 (16.7) | 1 (10) | 2 (22.2) | |||
| T4 | 4 (22.2) | - | 1 (11.1) | |||
| TNM classification, N | N0 | 5 (27.8) | 8 (80) | 0.054 | 5 (55.6) | 0.59 |
| N1 | 7 (38.9) | 2 (20) | 3 (33.3) | |||
| N2 | 4 (22.2) | 1 (11.1) | ||||
| N3 | 2 (11.1) | - | ||||
| TNM classification, M | M0 | 11 (61.1) | 8 (80) | 0.41 | 4 (44.4) | 0.06 |
| M1 | 7 (38.9) | 2 (20) | 2 (22.2) | |||
| Mx | - | - | 3 (33.3) | |||
| Tumor stage | I | 2 (11.1) | - | 0.07 | 1 (11.1) | 0.45 |
| II | 7 (38.9) | 9 (90) | 5 (55.6) | |||
| III | 7 (38.9) | 1 (10) | 1 (11.1) | |||
| IV | 2 (11.1) | - | - | |||
| Tumor grade | 2 | 4 (22.2) | - | 0.26 | - | 0.056 |
| 3 | 14 (77.8) | 10 (100) | 7 (77.8) | |||
| Chemotherapy | No | 11 (61.1) | 8 (80) | 0.5 | 6 (66.7) | 0.65 |
| Yes | 4 (22.2) | 2 (20) | 3 (33.3) | |||
| TP53 mutation | No | 4 (22.2) | - | 0.13 | 4 (44.4) | 1 |
| Yes | 12 (66.7) | 6 (60) | 5 (55.6) | |||
| Family history | No | 10 (55.6) | 5 (50) | 0.87 | 2 (22.2) | 0.001 |
| Yes | 2 (11.1) | 2 (20) | 7 (77.8) | |||
| Outcome | ||||||
| Relapse or metastasis | No | 11 (61.1) | 5 (50) | 0.69 | 7 (77.8) | 1 |
| Yes | 7 (38.9) | 5 (50) | 2 (22.2) | |||
| Death | No | 13 (72.2) | 7 (70) | 1 | 7 (77.8) | 1 |
| Yes | 5 (27.8) | 3 (30) | 2 (22.2) | |||
| Host Gene | Strand | circRNA | BaseMean | Log2FC | Padj |
|---|---|---|---|---|---|
| LIFR | − | hsa_circ_0072309 | 7.478 | −2.392 | 4.68 × 10−11 |
| SEMA3C | − | hsa_circ_0004365 | 5.284 | −2.347 | 6.61 × 10−10 |
| MIR31HG | − | hsa_circ_0008599 | 3.758 | −1.981 | 3.61 × 10−06 |
| EXOC6B | − | hsa_circ_0009043 | 10.768 | −1.955 | 2.74 × 10−07 |
| WDR78 | − | hsa_circ_0006677 | 8.343 | −1.931 | 1.29 × 10−07 |
| FAM126A | − | hsa_circ_0008951 | 4.761 | −1.925 | 2.74 × 10−07 |
| ACVR2A | + | hsa_circ_0001073 | 11.392 | −1.895 | 3.01 × 10−07 |
| RMST | + | hsa_circ_0099634 | 3.223 | −1.846 | 8.57 × 10−05 |
| N4BP2L2 | − | hsa_circ_0000471 | 24.928 | −1.825 | 3.61 × 10−06 |
| RHOBTB3 | + | hsa_circ_0007444 | 8.915 | −1.700 | 1.66 × 10−04 |
| SEMA3C | − | hsa_circ_0002714 | 2.767 | −1.693 | 2.00 × 10−04 |
| FGFR1 | − | hsa_circ_0008016 | 2.983 | −1.641 | 3.11 × 10−04 |
| RBM23 | − | hsa_circ_0000524 | 7.974 | −1.626 | 8.57 × 10−05 |
| PRDM5 | − | hsa_circ_0005654 | 2.864 | −1.584 | 5.28 × 10−04 |
| NFIB | − | hsa_circ_0086376 | 7.423 | −1.540 | 2.00 × 10−04 |
| TPTEP1 | + | chr22:17117929-17119630 | 5.560 | −1.522 | 2.76 × 10−03 |
| EPSTI1 | − | hsa_circ_0000479 * | 4.422 | 1.560 | 2.55 × 10−04 |
| circRNAs | RBP | Number of circRNAs Binding Sites * | RBP Function | Reference |
|---|---|---|---|---|
| hsa_circ_0009043 | AGO1 | 1 | miRNA-mediated gene regulation | [35] |
| hsa_circ_0072309, hsa_circ_0009043, hsa_circ_0004365, hsa_circ_0000479 | AGO2 | 3/1/6/1 | miRNA-mediated gene regulation | [35] |
| hsa_circ_0072309, hsa_circ_0009043, hsa_circ_0004365, hsa_circ_0008599, hsa_circ_0006677 | EIF4A3 | 6/4/6/1/2 | RNA splicing by acting as a core component of the spliceosome and splicing-dependent exon junction complex | [36] |
| hsa_circ_0072309, hsa_circ_0009043, hsa_circ_0004365 | ELAVL1 (HuR) | 2/2/2 | Increase mRNA stability through binding to their 3’-UTR | [37] |
| hsa_circ_0072309 | EWSR1 | 4 | Regulating transcription through interaction with CREB-binding protein; RNA splicing by cooperating with multiple splicing factors | [38,39] |
| hsa_circ_0072309 | FMR1 (FMRP) | 3 | Associates in an RNA-dependent manner with MOV10 and facilitates miRNA-mediated gene silencing; binds to mRNAs and mediates RNA transport from nucleus to cytoplasm | [40,41] |
| hsa_circ_0072309 | FUS | 1 | Mediates the binding of U1 snRNP and RNAPII, being required for splicing to occur during transcription | [42] |
| hsa_circ_0072309, hsa_circ_0004365 | IGF2BP1 | 1/1 | Binding to cytoplasmic mRNAs in order to prevent premature RNA decay; transport RNA in the cytoplasm and provide stability to bound mRNAs | [43,44,45] |
| hsa_circ_0072309, hsa_circ_0004365 | IGF2BP2 | 1/1 | Binding to cytoplasmic mRNAs in order to prevent premature RNA decay; transport RNA in the cytoplasm and provide stability to bound mRNAs | [43,44,45] |
| hsa_circ_0072309, hsa_circ_0004365 | IGF2BP3 | 1/2 | Binding to cytoplasmic mRNAs in order to prevent premature RNA decay; transport RNA in the cytoplasm and provide stability to bound mRNAs | [43,44,45] |
| hsa_circ_0072309 | LIN28A | 2 | Stimulates translation by actively recruiting RNA helicase A to polysomes; inhibiting miRNA biogenesis | [46,47] |
| hsa_circ_0072309 | LIN28B | 1 | Stimulates translation by actively recruiting RNA helicase A to polysomes; inhibiting miRNA biogenesis | [46,47] |
| hsa_circ_0072309, hsa_circ_0009043, hsa_circ_0004365 | PTBP1 | 1/1/3 | Exon exclusion during alternative splicing events and mRNA stabilization; stimulates translation at picorna virus internal ribosome entry sites (IRES) | [48,49] |
| hsa_circ_0072309 | U2AF2 (U2AF) | 1 | Activation of splicing and its coupling to transcription; 3’ end processing of vertebrates | [50,51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magalhães, L.; Ribeiro-dos-Santos, A.M.; Cruz, R.L.; Nakamura, K.D.d.M.; Brianese, R.; Burbano, R.; Ferreira, S.P.; Oliveira, E.L.F.d.; Anaissi, A.K.M.; Nahúm, M.C.d.S.; et al. Triple-Negative Breast Cancer circRNAome Reveals Hsa_circ_0072309 as a Potential Risk Biomarker. Cancers 2022, 14, 3280. https://doi.org/10.3390/cancers14133280
Magalhães L, Ribeiro-dos-Santos AM, Cruz RL, Nakamura KDdM, Brianese R, Burbano R, Ferreira SP, Oliveira ELFd, Anaissi AKM, Nahúm MCdS, et al. Triple-Negative Breast Cancer circRNAome Reveals Hsa_circ_0072309 as a Potential Risk Biomarker. Cancers. 2022; 14(13):3280. https://doi.org/10.3390/cancers14133280
Chicago/Turabian StyleMagalhães, Leandro, André M. Ribeiro-dos-Santos, Rebecca L. Cruz, Kivvi Duarte de Mello Nakamura, Rafael Brianese, Rommel Burbano, Sâmio Pimentel Ferreira, Ewaldo Lúcio Foro de Oliveira, Ana Karyssa Mendes Anaissi, Márcia Cristina de Sousa Nahúm, and et al. 2022. "Triple-Negative Breast Cancer circRNAome Reveals Hsa_circ_0072309 as a Potential Risk Biomarker" Cancers 14, no. 13: 3280. https://doi.org/10.3390/cancers14133280