
Phytochemicals for the Prevention and Treatment of Renal Cell Carcinoma: Preclinical and Clinical Evidence and Molecular Mechanisms

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Methodology for Literature Search and Study Selection
3. Phytocompounds in Renal Cancer Research: Preclinical Studies
3.1. Phenolics
3.1.1. Alpinumisoflavone
3.1.2. Angelicin
3.1.3. Apigenin
3.1.4. Chrysin
3.1.5. Coumarin
3.1.6. Curcumin
3.1.7. Cyanidin
3.1.8. Daphnetin
3.1.9. Dicoumarol
3.1.10. Esculetin
3.1.11. Eupafolin
3.1.12. Gambogic Acid
3.1.13. Genistein
3.1.14. Hispidulin
3.1.15. Honokiol
3.1.16. Hymecromone
3.1.17. Icaritin
3.1.18. Isoliquiritigenin
3.1.19. Luteolin
3.1.20. Osthole
3.1.21. Praeruptorin B
3.1.22. Pterostilbene
3.1.23. Resveratrol
3.1.24. Umbelliferone
3.2. Terpenes and Terpenoids
3.2.1. Anatolicin
3.2.2. Artemisinin
3.2.3. Artesunate
3.2.4. Auraptene
3.2.5. β-Elemene
3.2.6. Betulin
3.2.7. Betulinic Acid
3.2.8. Cafestol
3.2.9. Carsonic Acid
3.2.10. Corosolic Acid
3.2.11. Crocin
3.2.12. Cucurbitacins
3.2.13. Englerin A
3.2.14. Escin
3.2.15. Helenalin
3.2.16. 16-Hydroxyclerod-3,13-dien-15,16-olide (CD)
3.2.17. Kahweol
3.2.18. Lycopene
3.2.19. Nimbolide
3.2.20. Oridonin
3.2.21. Parthenolide
3.2.22. Silibinin
3.2.23. Sorghumol
3.2.24. Thymoquinone
3.2.25. Tonantzitlolone
3.2.26. Triptolide
3.2.27. Triptolidenol
3.2.28. Zerumbone
3.3. Alkaloids
3.3.1. (-)-Antofine
3.3.2. Berberine
3.3.3. Cepharanthine
3.3.4. Chelerythrine
3.3.5. Dauricine
3.3.6. Neferine
3.3.7. Neopapillarine
3.3.8. Oxymatrine
3.3.9. Piperlongumine
3.3.10. Tetrandrine
3.4. Sulfur-Containing Compounds
3.4.1. Allicin
3.4.2. Sulforaphane
4. Phytocompounds in Renal Cancer Research: Clinical Studies
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| ATF4 | activation of transcription factor-4 |
| ATG | autophagy related |
| Bax | Bcl-2 associated X-protein |
| Bcl-2 | B-cell lymphoma 2 |
| Bcl-xL | B-cell lymphoma-extra-large |
| BGPa | transmembrane carcinoembryonic antigen |
| Bim | Bcl-2-like protein 11 |
| BMK | baby mouse kidney |
| BTG3 | B-cell translocation gene 3 |
| ccRCC | clear cell renal cell carcinoma |
| CDK2 | cyclin-dependent kinase 2 |
| CDK4 | cyclin-dependent kinase 4 |
| CDK6 | cyclin-dependent kinase 6 |
| CD31 | cluster of differentiation 31 |
| CD44 | cluster of differentiation 44 |
| c-FLIP | cellular FLICE-like inhibitory protein |
| CHOP | CCSST enhancer-binding protein-homologous protein 1 |
| COX-2 | cyclooxygenase-2 |
| CRE | cAMP response element |
| CTSC | cathepsin C |
| CTSV | cathepsin V |
| CXCR4 | chemokine receptor 4 |
| CYP1B1 | dioxin-inducible cytochrome P450 |
| cyt. c | cytochrome c |
| DEN | N-nitrosodiethylamine |
| DFO | deferoxamine |
| Dkk2 | Dickkopf WNT signaling pathway inhibitor 2 |
| DMSO | dimethyl sulfoxide |
| DNMTase | DNA methyltransferase |
| DR | death receptor |
| EGFR | epidermal growth factor receptor |
| EGR1 | early growth response protein 1 |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| FADD | Fas-associated death domain |
| FDA | Food and Drug Administration |
| fIGF | c-FOS-induced growth factor |
| GADD45 | growth arrest and DNA-damage-inducible protein |
| GFRA2 | glial family receptor α 2 |
| GLUT1 | glucose transporter 1 |
| GPx | glutathione peroxidase |
| Grb2 | growth-factor receptor bound protein-2 |
| GRP78 | glucose-regulated protein 78 |
| GSH | glutathione |
| GSK-3β | glycogen synthase kinase 3β |
| GST | glutathione S-transferases |
| HCK | hemopoietic cell protein-tyrosine kinase |
| HDAC | histone deacetylase |
| HGF | hepatocyte growth factor |
| HGMB1 | high mobility group box 1 protein |
| HIF | hypoxia inducible factor |
| HO-1 | heme oxygenase-1 |
| HSF1 | heat shock factor 1 |
| HSP | heat shock protein |
| IAP | inhibitor of apoptosis |
| IFN-α | interferon-alpha |
| IKKβ | inhibitor of nuclear factor-κB kinase subunit β |
| IL-6 | interleukin 6 |
| i.p. | intraperitoneal |
| JAK | Janus kinase |
| LC3 | light chain 3 |
| LDH | lactate dehydrogenase |
| MAPK | mitogen activated protein kinase |
| Mcl-1 | myeloid cell leukemia-1 |
| MCM2 | minichromosome maintenance complex component 2 |
| Mdm2 | murine double minute 2 |
| MET | mesenchymal–epithelial transition |
| miR-101 | microRNA 101 |
| MLC | myosin light chain |
| MMP | matrix metalloproteinase |
| mTOR | mammalian target of rapamycin |
| MUC1 | mucin 1 |
| NAC | N-acetylcysteine |
| NEAA | nonessential amino acids |
| NFIB | nuclear factor I B |
| NF-κB | nuclear factor-κB |
| NLRP3 | NLR family pyrin domain containing 3 |
| Nrf2 | nuclear factor-like 2 |
| ODC | ornithine decarboxylase |
| PARP | poly ADP-ribose polymerase |
| PCNA | proliferating cell nuclear antigen |
| PDGF | platelet derived growth factor |
| PDK1 | phosphoinositide-dependent kinase-1 |
| PDT | photodynamic therapy |
| PGE2 | prostaglandin E2 |
| PI3K | phosphoinositide 3-kinase |
| PKCα | protein kinase C-α |
| PKCθ | protein kinase C-θ |
| PKM2 | pyruvate kinase muscle isozyme M2 |
| PLIN2 | perilipin 2 |
| PLK3 | polo like kinase 3 |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta-Analyses |
| PXR | human pregnane X |
| PUMA | p53 up-regulated modulator of apoptosis |
| p-ATM | phosphorylated-ataxia telangiectasia mutated |
| p-Cdc2 | phosphorylated Cdc2 on tyrosine 15 |
| p-Cdc25c | phosphorylated Cdc25 on serine 216 |
| p-ERK | phosphorylated extracellular signal-regulated kinase |
| p-JNK | phosphorylated c-Jun NH2-terminal kinase |
| p-MEK | phosphorylated mitogen activated protein kinase |
| p-STAT3 | phosphorylated signal transducer and activator of transcription 3 |
| p-S6 | phosphorylated ribosomal protein S6 |
| p-Akt | phosphorylated Akt |
| QR | quinone reductase |
| RAC1 | ras-related C3 botulinum toxin substrate 1 |
| RCC | renal cell carcinoma |
| REDD1 | regulation in development and DNA damage responses |
| RHAMM | receptor for hyaluronan-mediated motility |
| RIP-1 | receptor-interacting protein 1 |
| RIP-3 | receptor-interacting protein-3 |
| RLIP76 | Ral interacting protein of 76 kDa |
| ROCK | Rho-associated protein kinase |
| ROS | reactive oxygen species |
| SEPW1 | selenoprotein W |
| sFRP1 | secreted frizzled related protein 1 |
| SHP-1 | Src homology 2 domain-containing protein tyrosine phosphatase-1 |
| Smad4 | SMAD Family Member 4 |
| SphK1 | sphingosine kinase 1 |
| STAMBPL1 | STAM binding protein like 1 |
| TBARS | tissue thiobarbituric acid derivatives |
| TdT | terminal deoxynucleotide transferase |
| TGF-α | transforming growth factor-α |
| TERT | telomerase reverse transcriptase |
| TIMP | tissue inhibitor of metalloproteinase |
| TNF-α | tumor necrosis factor-α |
| TRAF-1 | tumor necrosis factor receptor associated factor 1 |
| TRAIL | tumor necrosis factor-related apoptosis-inducing ligand |
| TRPC4 | transient receptor potential cation channel 4 |
| TSC | tuberous sclerosis |
| ULK1 | Unc-51 like autophagy activating kinase |
| USP53 | ubiquitin-specific protease 53 |
| u-PA | urokinase plasminogen activator |
| VEGF | vascular endothelial growth factor |
| VDR | vitamin D receptor |
| VHL | von Hippel–Lindau |
| Wnt | wingless-related integration site |
| YAP | Yes-associated protein |
| ZEB2 | zinc finger E-box binding homeobox 2 |
| γH2AX | H2A histone family member X phosphorylated on serine 139 |
| 5-FU | 5-fluorouracil |
References
- Escudier, B.; Porta, C.; Schmidinger, M.; Rioux-Leclercq, N.; Bex, A.; Khoo, V.; Grünwald, V.; Gillessen, S.; Horwich, A.; ESMO Guidelines Committee. Renal cell carcinoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2019, 30, 706–720. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Safiri, S.; Kolahi, A.-A.; Mansournia, M.A.; Almasi-Hashiani, A.; Ashrafi-Asgarabad, A.; Sullman, M.J.M.; Bettampadi, D.; Qorbani, M.; Moradi-Lakeh, M.; Ardalan, M.; et al. The burden of kidney cancer and its attributable risk factors in 195 countries and territories, 1990–2017. Sci. Rep. 2020, 10, 13862. [Google Scholar] [CrossRef] [PubMed]
- Muglia, V.F.; Prando, A. Renal cell carcinoma: Histological classification and correlation with imaging findings. Radiol. Bras. 2015, 48, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Gray, R.E.; Harris, G.T. Renal Cell Carcinoma: Diagnosis and Management. Am. Fam. Physician 2019, 99, 179–184. [Google Scholar]
- Banumathy, G.; Cairns, P. Signaling pathways in renal cell carcinoma. Cancer Biol. Ther. 2010, 10, 658–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Cheng, L.; Zhang, H.; Ma, L.; Wang, Y.; Niu, W.; Liu, Z.; Song, Y.; Liang, P.; Zhao, G.; et al. Laparoscopic and Robotic-Assisted Partial Nephrectomy: An Overview of Hot Issues. Urol. Int. 2020, 104, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Atkins, M.B.; Tannir, N.M. Current and emerging therapies for first-line treatment of metastatic clear cell renal cell carcinoma. Cancer Treat. Rev. 2018, 70, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H. Predictive Biomarkers and Novel Targets in the Treatment of Metastatic Renal Cell Carcinoma. Curr. Med. Chem. 2021, 28, 5213–5227. [Google Scholar] [CrossRef]
- Secasan, C.C. Prognostic markers and new, innovative treatments in renal cell carcinoma. Saudi J. Kidney Dis. Transplant. 2009, 20, 355–361. [Google Scholar]
- Tenold, M.; Ravi, P.; Kumar, M.; Bowman, A.; Hammers, H.; Choueiri, T.K.; Lara, P.N. Current Approaches to the Treatment of Advanced or Metastatic Renal Cell Carcinoma. Am. Soc. Clin. Oncol. Educ. Book 2020, 40, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Tseng, M. Diet, cancer and public health nutrition. Public Health Nutr. 2009, 12, 737–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40–41, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, F.; Tabanca, N.; Kiss, R. Assessing the anticancer effects associated with food products and/or nutraceuticals using in vitro and in vivo preclinical development-related pharmacological tests. Semin. Cancer Biol. 2017, 46, 14–32. [Google Scholar] [CrossRef]
- Zubair, H.; Azim, S.; Ahmad, A.; Khan, M.A.; Patel, G.K.; Singh, S.; Singh, A.P. Cancer Chemoprevention by Phytochemicals: Nature’s Healing Touch. Molecules 2017, 22, 395. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med Princ. Pract. 2015, 25, 41–59. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, H.B.; Singh, A.; Naqvi, A.H.; Singh, B.R. Dietary phytochemicals alter epigenetic events and signaling pathways for inhibition of metastasis cascade: Phytoblockers of metastasis cascade. Cancer Metastasis Rev. 2014, 33, 41–85. [Google Scholar] [CrossRef]
- Das, B.; Sarkar, N.; Bishayee, A.; Sinha, D. Dietary phytochemicals in the regulation of epithelial to mesenchymal transition and associated enzymes: A promising anticancer therapeutic approach. Semin. Cancer Biol. 2018, 56, 196–218. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front. Pharmacol. 2019, 10, 1614, Corrigendum in Front. Pharmacol. 2020, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on natural compounds in chemoprevention and treatment of cancer: An update with new promising compounds. Eur. J. Cancer 2021, 149, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Shankar, M.G.; Swetha, M.; Keerthana, C.K.; Rayginia, T.P.; Anto, R.J. Cancer Chemoprevention: A Strategic Approach Using Phytochemicals. Front. Pharmacol. 2022, 12, 809308. [Google Scholar] [CrossRef]
- Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 9, 1451. [Google Scholar] [CrossRef]
- Gaikwad, S.; Srivastava, S.K. Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer. Antioxidants 2021, 10, 83. [Google Scholar] [CrossRef]
- Tewari, D.; Bawari, S.; Sharma, S.; DeLiberto, L.K.; Bishayee, A. Targeting the crosstalk between canonical Wnt/β-catenin and inflammatory signaling cascades: A novel strategy for cancer prevention and therapy. Pharmacol. Ther. 2021, 227, 107876. [Google Scholar] [CrossRef]
- Braicu, C.; Zanoaga, O.; Zimta, A.-A.; Tigu, A.B.; Kilpatrick, K.L.; Bishayee, A.; Nabavi, S.M.; Berindan-Neagoe, I. Natural compounds modulate the crosstalk between apoptosis- and autophagy-regulated signaling pathways: Controlling the uncontrolled expansion of tumor cells. Semin. Cancer Biol. 2022, 80, 218–236. [Google Scholar] [CrossRef]
- Ma, Z.; Xiang, X.; Li, S.; Xie, P.; Gong, Q.; Goh, B.-C.; Wang, L. Targeting hypoxia-inducible factor-1, for cancer treatment: Recent advances in developing small-molecule inhibitors from natural compounds. Semin. Cancer Biol. 2020, 80, 379–390. [Google Scholar] [CrossRef]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: A novel therapeutic strategy. Semin. Cancer Biol. 2022, 80, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Moloudizargari, M.; Asghari, M.H.; Nabavi, S.F.; Gulei, D.; Berindan-Neagoe, I.; Bishayee, A. Targeting Hippo signaling pathway by phytochemicals in cancer therapy. Semin. Cancer Biol. 2020, 80, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Moradi, S.Z.; Ash-Rafzadeh, A.; Bishayee, A. Targeting cellular senescence in cancer by plant secondary metabolites: A systematic review. Pharmacol. Res. 2021, 177, 105961. [Google Scholar] [CrossRef] [PubMed]
- Peiris, D.C. Natural Medicine as a Therapy for Renal Carcinoma: A Review. In Renal Cell Carcinoma, 1st ed.; SM Group: Dover, DE, USA, 2015. [Google Scholar]
- Haque, I.; Subramanian, A.; Huang, C.H.; Godwin, A.K.; Van Veldhuizen, P.J.; Banerjee, S.; Banerjee, S.K. The Role of Compounds Derived from Natural Supplement as Anticancer Agents in Renal Cell Carcinoma: A Review. Int. J. Mol. Sci. 2017, 19, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartogh, D.J.D.; Tsiani, E. Health Benefits of Resveratrol in Kidney Disease: Evidence from In Vitro and In Vivo Studies. Nutrient 2019, 11, 1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gotzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Healthcare Interventions: Explanation and Elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [Green Version]
- Ateba, S.B.; Mvondo, M.A.; Djiogue, S.; Zingue, S.; Krenn, L.; Njamen, D. A Pharmacological Overview of Alpinumisoflavone, a Natural Prenylated Isoflavonoid. Front. Pharmacol. 2019, 10, 952. [Google Scholar] [CrossRef]
- Wang, T.; Jiang, Y.; Chu, L.; Wu, T.; You, J. Alpinumisoflavone suppresses tumour growth and metastasis of clear-cell renal cell carcinoma. Am. J. Cancer Res. 2017, 7, 999–1015, Erratum in Am. J. Cancer Res. 2019, 9, 455–457. [Google Scholar]
- Min, K.-J.; Um, H.J.; Seo, S.U.; Woo, S.M.; Kim, S.; Park, J.-W.; Lee, H.-S.; Kim, S.H.; Choi, Y.H.; Lee, T.-J.; et al. Angelicin potentiates TRAIL-induced apoptosis in renal carcinoma Caki cells through activation of caspase 3 and down-regulation of c-FLIP expression. Drug Dev. Res. 2017, 79, 3–10. [Google Scholar] [CrossRef]
- Meng, S.; Zhu, Y.; Li, J.-F.; Wang, X.; Liang, Z.; Li, S.-Q.; Xu, X.; Chen, H.; Liu, B.; Zheng, X.-Y.; et al. Apigenin inhibits renal cell carcinoma cell proliferation. Oncotarget 2017, 8, 19834–19842. [Google Scholar] [CrossRef] [Green Version]
- Myers, R.B.; Parker, M.; Grizzle, W.E. The effects of coumarin and suramin on the growth of malignant renal and prostatic cell lines. J. Cancer Res. Clin. Oncol. 1994, 120, S11–S13. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.-H.; Kim, Y.-H.; Choi, Y.-J.; Kim, D.-G.; Lee, K.-S.; Bae, J.H.; Min, D.S.; Chang, J.-S.; Jeong, Y.-J.; Lee, Y.H.; et al. Molecular mechanisms of curcumin-induced cytotoxicity: Induction of apoptosis through generation of reactive oxygen species, down-regulation of Bcl-XL and IAP, the release of cytochrome c and inhibition of Akt. Carcinogenesis 2003, 24, 1199–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, E.M.; Lim, J.H.; Lee, T.J.; Park, J.-W.; Choi, K.S.; Kwon, T.K. Curcumin sensitizes tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through reactive oxygen species-mediated upregulation of death receptor 5 (DR5). Carcinogenesis 2005, 26, 1905–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, B.R.; Min, K.-J.; Cho, I.J.; Kim, S.C.; Kwon, T.K. Curcumin Significantly Enhances Dual PI3K/Akt and mTOR Inhibitor NVP-BEZ235-Induced Apoptosis in Human Renal Carcinoma Caki Cells through Down-Regulation of p53-Dependent Bcl-2 Expression and Inhibition of Mcl-1 Protein Stability. PLoS ONE 2014, 9, e95588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, W.; Li, B.; Zhang, K.; Wu, Y.; Xu, H.; Wang, J.; Zhang, J.; Fan, R.; Wei, J. Curcumin Promotes Cell Cycle Arrest and Inhibits Survival of Human Renal Cancer Cells by Negative Modulation of the PI3K/AKT Signaling Pathway. Cell Biophys. 2015, 73, 681–686. [Google Scholar] [CrossRef]
- Xu, S.; Yang, Z.; Fan, Y.; Guan, B.; Jia, J.; Gao, Y.; Wang, K.; Wu, K.; Wang, X.; Zheng, P.; et al. Curcumin enhances temsirolimus-induced apoptosis in human renal carcinoma cells through upregulation of YAP/p53. Oncol. Lett. 2016, 12, 4999–5006. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, D.; Hao, Y.; Liu, Q.; Wu, Y.; Liu, X.; Luo, J.; Zhou, T.; Sun, B.; Luo, X.; et al. Cyanidin Curtails Renal Cell Carcinoma Tumorigenesis. Cell. Physiol. Biochem. 2018, 46, 2517–2531. [Google Scholar] [CrossRef] [Green Version]
- Finn, G.J.; Creaven, B.S.; Egan, D.A. Daphnetin induced differentiation of human renal carcinoma cells and its mediation by p38 mitogen-activated protein kinase. Biochem. Pharmacol. 2004, 67, 1779–1788. [Google Scholar] [CrossRef]
- Park, E.J.; Min, K.-J.; Choi, K.S.; Kwon, T.K. Dicoumarol sensitizes renal cell carcinoma Caki cells to TRAIL-induced apoptosis through down-regulation of Bcl-2, Mcl-1 and c-FLIP in a NQO1-independent manner. Exp. Cell Res. 2014, 323, 144–154. [Google Scholar] [CrossRef]
- Duan, J.; Shi, J.; Ma, X.; Xuan, Y.; Li, P.; Wang, H.; Fan, Y.; Gong, H.; Wang, L.; Pang, Y.; et al. Esculetin inhibits proliferation, migration, and invasion of clear cell renal cell carcinoma cells. Biomed. Pharmacother. 2020, 125, 110031. [Google Scholar] [CrossRef]
- Han, M.A.; Min, K.-J.; Woo, S.M.; Seo, B.R.; Kwon, T.K. Eupafolin enhances TRAIL-mediated apoptosis through cathepsin S-induced down-regulation of Mcl-1 expression and AMPK-mediated Bim up-regulation in renal carcinoma Caki cells. Oncotarget 2016, 7, 65707–65720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.-L.; Zhang, Y.; Luo, C.-L.; Wu, X.-H. Targeting renal cell carcinoma with gambogic acid in combination with sunitinib in vitro and in vivo. Asian Pac. J. Cancer Prev. 2012, 13, 6463–6468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasamura, H.; Takahashi, A.; Yuan, J.; Kitamura, H.; Masumori, N.; Miyao, N.; Itoh, N.; Tsukamoto, T. Antiproliferative and antiangiogenic activities of genistein in human renal cell carcinoma. Urology 2004, 64, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Majid, S.; Dar, A.A.; Ahmad, A.E.; Hirata, H.; Kawakami, K.; Shahryari, V.; Saini, S.; Tanaka, Y.; Dahiya, A.V.; Khatri, G.; et al. BTG3 tumor suppressor gene promoter demethylation, histone modification and cell cycle arrest by genistein in renal cancer. Carcinogenesis 2009, 30, 662–670. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Ueno, K.; Nakajima, K.; Tabatabai, Z.L.; Hinoda, Y.; Ishii, N.; Dahiya, R. Genistein downregulates onco-miR-1260b and inhibits Wnt-signalling in renal cancer cells. Br. J. Cancer 2013, 108, 2070–2078. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Jiang, Q.; Han, Y.; Peng, J.; Wang, C. Hispidulin Potentiates the Antitumor Effect of Sunitinib against Human Renal Cell Carcinoma in Laboratory Models. Cell Biophys. 2014, 71, 757–764. [Google Scholar] [CrossRef]
- Gao, H.; Gao, M.-Q.; Peng, J.-J.; Han, M.; Liu, K.-L.; Han, Y.-T. RETRACTED ARTICLE: Hispidulin mediates apoptosis in human renal cell carcinoma by inducing ceramide accumulation. Acta Pharmacol. Sin. 2017, 38, 1618–1631. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, Q.; Su, Q.; Ma, D.; An, C.; Ma, L.; Liang, A.H. Honokiol Suppresses Renal Cancer Cells’ Metastasis via Dual-Blocking Epithelial-Mesenchymal Transition and Cancer Stem Cell Properties through Modulating miR-141/ZEB2 Signaling. Mol. Cells 2014, 37, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Castillo, V.; Welty, M.; Eliaz, I.; Sliva, D. Honokiol inhibits migration of renal cell carcinoma through activation of RhoA/ROCK/MLC signaling pathway. Int. J. Oncol. 2016, 49, 1525–1530. [Google Scholar] [CrossRef] [Green Version]
- Benitez, A.; Yates, T.J.; Shamaldevi, N.; Bowen, T.; Lokeshwar, V.B. Dietary Supplement Hymecromone and Sorafenib: A Novel Combination for the Control of Renal Cell Carcinoma. J. Urol. 2013, 190, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Priceman, S.J.; Xin, H.; Zhang, W.; Deng, J.; Liu, Y.; Huang, J.; Zhu, W.; Chen, M.; Hu, W.; et al. Icaritin Inhibits JAK/STAT3 Signaling and Growth of Renal Cell Carcinoma. PLoS ONE 2013, 8, e81657. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H.; Park, J.E.; Chae, I.G.; Park, G.; Lee, S.; Chun, K.-S. Isoliquiritigenin inhibits the proliferation of human renal carcinoma Caki cells through the ROS-mediated regulation of the Jak2/STAT3 pathway. Oncol. Rep. 2017, 38, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, Y.-C.; Kuan, Y.-H.; Li, J.-R.; Raung, S.-L.; Wang, C.-C.; Hung, Y.-Y.; Chen, C.-J. Induction of Apoptosis by Luteolin Involving Akt Inactivation in Human 786-O Renal Cell Carcinoma Cells. Evid.-Based Complementary Altern. Med. 2013, 2013, 109105. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.-C.; Li, J.-R.; Kuan, Y.-H.; Raung, S.-L.; Wang, C.-C.; Hung, Y.-Y.; Pan, P.-H.; Lu, H.-C.; Chen, C.-J. Luteolin sensitizes human 786-O renal cell carcinoma cells to TRAIL-induced apoptosis. Life Sci. 2014, 100, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Mao, J.; Wang, Q.; Zhang, Z.; Wu, G.; Tang, Q.; Zhao, B.; Li, L.; Li, Q. In vitro anticancer activities of osthole against renal cell carcinoma cells. Biomed. Pharmacother. 2017, 94, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Min, K.-J.; Han, M.A.; Kim, S.; Park, J.-W.; Kwon, T.K. Osthole enhances TRAIL-mediated apoptosis through downregulation of c-FLIP expression in renal carcinoma Caki cells. Oncol. Rep. 2017, 37, 2348–2354. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Li, J.; Zhu, W.; Leng, T.; Shu, M.; Huang, Y.; Xu, D.; Qiu, P.; Su, X. Triptolide-induced cell cycle arrest and apoptosis in human renal cell carcinoma cells. Oncol. Rep. 2011, 25, 979–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Ye, D.; Luo, Q.; Li, J.; Liu, J. Pterostilbene Inhibits Human Renal Cell Carcinoma Cells Growth and Induces DNA Damage. Biol. Pharm. Bull. 2020, 43, 258–265. [Google Scholar] [CrossRef]
- Shi, T.; Liou, L.S.; Sadhukhan, P.; Duan, Z.-H.; Novick, A.C.; Hissong, J.G.; Almasan, A.; DiDonato, J.A. Effects of resveratrol on gene expression in renal cell carcinoma. Cancer Biol. Ther. 2004, 3, 882–888. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yang, R.; Zhu, L. Inhibitory effect of resveratrol on the expression of the VEGF gene and proliferation in renal cancer cells. Mol. Med. Rep. 2011, 4, 981–983. [Google Scholar] [CrossRef]
- Dai, L.; Chen, L.; Wang, W.; Lin, P. Resveratrol inhibits ACHN cells via regulation of histone acetylation. Pharm. Biol. 2020, 58, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fang, Q.; Ji, S.; Han, Z.; Cheng, W.; Zhang, H. Resveratrol-mediated apoptosis in renal cell carcinoma via the p53/AMP-activated protein kinase/mammalian target of rapamycin autophagy signaling pathway. Mol. Med. Rep. 2017, 17, 502–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Tang, H.; Zeng, X.; Ye, D.; Liu, J. Resveratrol inhibits proliferation, migration and invasion via Akt and ERK1/2 signaling pathways in renal cell carcinoma cells. Biomed. Pharmacother. 2018, 98, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zhang, S.; Zhang, Q.; Kang, L.; Ma, C.; Feng, L.; Li, S.; Li, J.; Yang, L.; Liu, J.; et al. Resveratrol inhibits tumor progression by down-regulation of NLRP3 in renal cell carcinoma. J. Nutr. Biochem. 2020, 85, 108489. [Google Scholar] [CrossRef]
- Min, Z.; Ying-Jie, K.; Ling-Wei, C.; Jia-Ping, G.; Ying, W.; Dan, Z.; Sen, Z. Resveratrol enhances chemosensitivity of renal cell carcinoma to paclitaxel. Front. Biosci. 2019, 24, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Baek, S.H.; Um, J.-Y.; Shim, B.S.; Ahn, K.S. Resveratrol attenuates constitutive STAT3 and STAT5 activation through induction of PTPε and SHP-2 tyrosine phosphatases and potentiates sorafenib-induced apoptosis in renal cell carcinoma. BMC Nephrol. 2016, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Huang, S.; Xin, X.; Ren, Y.; Weng, G.; Wang, P. The antitumor activity of umbelliferone in human renal cell carcinoma via regulation of the p110γ catalytic subunit of PI3Kγ. Acta Pharm. 2018, 69, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Tosun, F.; Beutler, J.A.; Ransom, T.T.; Miski, M. Anatolicin, a Highly Potent and Selective Cytotoxic Sesquiterpene Coumarin from the Root Extract of Heptaptera anatolica. Molecules 2019, 24, 1153. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Sun, P.; Zhou, Y.; Shen, B.; Zhou, M.; Wu, L.; Kong, M. Inhibition of AKT enhances the anti-cancer effects of Artemisinin in clear cell renal cell carcinoma. Biomed. Pharmacother. 2019, 118, 109383. [Google Scholar] [CrossRef]
- Chauhan, A.K.; Min, K.-J.; Kwon, T.K. RIP1-dependent reactive oxygen species production executes artesunate-induced cell death in renal carcinoma Caki cells. Mol. Cell. Biochem. 2017, 435, 15–24. [Google Scholar] [CrossRef]
- Jang, Y.; Han, J.; Kim, S.J.; Kim, J.; Lee, M.J.; Jeong, S.; Ryu, M.J.; Seo, K.-S.; Choi, S.-Y.; Shong, M.; et al. Suppression of mitochondrial respiration with auraptene inhibits the progression of renal cell carcinoma: Involvement of HIF-1α degradation. Oncotarget 2015, 6, 38127–38138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.-H.; Liu, J.; Qu, X.-J.; Hou, K.-Z.; Wang, K.-F.; Liu, Y.-P.; Wu, B. β-Elemene induces apoptosis in human renal-cell carcinoma 786-O cells through inhibition of MAPK/ERK and PI3K/Akt/ mTOR signalling pathways. Asian Pac. J. Cancer Prev. 2012, 13, 2739–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; Ji, S.; Zhang, H.; Han, Z.; Liu, Q.; Wang, J.; Ping, H. mTOR activation is critical for betulin treatment in renal cell carcinoma cells. Biochem. Biophys. Res. Commun. 2017, 482, 1030–1036. [Google Scholar] [CrossRef]
- Yang, C.; Li, Y.; Fu, L.; Jiang, T.; Meng, F. Betulinic acid induces apoptosis and inhibits metastasis of human renal carcinoma cells in vitro and in vivo. J. Cell. Biochem. 2018, 119, 8611–8622. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Park, E.J.; Oh, J.H.; Min, K.-J.; Yang, E.S.; Kim, Y.H.; Lee, T.J.; Kim, S.H.; Choi, Y.H.; Park, J.-W.; et al. Cafestol, a coffee-specific diterpene, induces apoptosis in renal carcinoma Caki cells through down-regulation of anti-apoptotic proteins and Akt phosphorylation. Chem. Interact. 2011, 190, 102–108. [Google Scholar] [CrossRef]
- Woo, S.M.; Min, K.-J.; Seo, B.R.; Nam, J.-O.; Choi, K.S.; Yoo, Y.H.; Kwon, T.K. Cafestol overcomes ABT-737 resistance in Mcl-1-overexpressed renal carcinoma Caki cells through downregulation of Mcl-1 expression and upregulation of Bim expression. Cell Death Dis. 2014, 5, e1514. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.-J.; Min, K.-J.; Bae, J.H.; Kwon, T.K. Carnosic acid sensitized TRAIL-mediated apoptosis through down-regulation of c-FLIP and Bcl-2 expression at the post translational levels and CHOP-dependent up-regulation of DR5, Bim, and PUMA expression in human carcinoma caki cells. Oncotarget 2015, 6, 1556–1568. [Google Scholar] [CrossRef]
- Park, J.E.; Park, B.; Chae, I.G.; Kim, D.-H.; Kundu, J.; Kundu, J.K.; Chun, K.-S. Carnosic acid induces apoptosis through inactivation of Src/STAT3 signaling pathway in human renal carcinoma Caki cells. Oncol. Rep. 2016, 35, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.M.; Seo, S.U.; Min, K.-J.; Im, S.-S.; Nam, J.-O.; Chang, J.-S.; Kim, S.; Park, J.-W.; Kwon, T.K. Corosolic Acid Induces Non-Apoptotic Cell Death through Generation of Lipid Reactive Oxygen Species Production in Human Renal Carcinoma Caki Cells. Int. J. Mol. Sci. 2018, 19, 1309. [Google Scholar] [CrossRef] [Green Version]
- Niu, L.; Wang, X.F.; Wang, X.R. Crocin suppresses cell proliferation and migration by regulating miR-577/NFIB in renal cell carcinoma. J. Biol. Regul. Homeost. Agents 2020, 34, 1523–1527. [Google Scholar] [CrossRef]
- Henrich, C.J.; Thomas, C.L.; Brooks, A.D.; Booth, N.L.; Lowery, E.M.; Pompei, R.J.; McMahon, J.B.; Sayers, T.J. Effects of cucurbitacins on cell morphology are associated with sensitization of renal carcinoma cells to TRAIL-induced apoptosis. Apoptosis 2011, 17, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnayake, R.; Covell, D.; Ransom, T.T.; Gustafson, K.R.; Beutler, J.A. Englerin A, a Selective Inhibitor of Renal Cancer Cell Growth, from Phyllanthus engleri. Org. Lett. 2008, 11, 57–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulzmaier, F.J.; Li, Z.; Nakashige, M.L.; Fash, D.M.; Chain, W.J.; Ramos, J.W. Englerin A Selectively Induces Necrosis in Human Renal Cancer Cells. PLoS ONE 2012, 7, e48032. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.T.; Yu, A.L.; Diccianni, M.B.; Theodorakis, E.A.; Batova, A. Renal cancer-selective Englerin A induces multiple mechanisms of cell death and autophagy. J. Exp. Clin. Cancer Res. 2013, 32, 57. [Google Scholar] [CrossRef] [Green Version]
- Carson, C.; Raman, P.; Tullai, J.; Xu, L.; Hénault, M.; Thomas, E.; Yeola, S.; Lao, J.; McPate, M.; Verkuyl, J.M.; et al. Englerin A Agonizes the TRPC4/C5 Cation Channels to Inhibit Tumor Cell Line Proliferation. PLoS ONE 2015, 10, e0127498. [Google Scholar] [CrossRef]
- Batova, A.; Altomare, D.; Creek, K.E.; Naviaux, R.K.; Wang, L.; Li, K.; Green, E.; Williams, R.; Naviaux, J.C.; Diccianni, M.; et al. Englerin A induces an acute inflammatory response and reveals lipid metabolism and ER stress as targetable vulnerabilities in renal cell carcinoma. PLoS ONE 2017, 12, e0172632. [Google Scholar] [CrossRef]
- Yuan, S.-Y.; Cheng, C.-L.; Wang, S.-S.; Ho, H.-C.; Chiu, K.-Y.; Chen, C.-S.; Chen, C.-C.; Shiau, M.-Y.; Ou, Y.-C. Escin induces apoptosis in human renal cancer cells through G2/M arrest and reactive oxygen species-modulated mitochondrial pathways. Oncol. Rep. 2017, 37, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.H.; Iqbal, T.; Min, K.-J.; Kim, S.; Park, J.-W.; Son, E.-I.; Lee, T.-J.; Kwon, T.K. Helenalin-induced apoptosis is dependent on production of reactive oxygen species and independent of induction of endoplasmic reticulum stress in renal cell carcinoma. Toxicol. In Vitro 2013, 27, 588–596. [Google Scholar] [CrossRef]
- Liu, C.; Lee, W.-C.; Huang, B.-M.; Chia, Y.-C.; Chen, Y.-C.; Chen, Y.-C. 16-Hydroxycleroda-3, 13-dien-15, 16-olide inhibits the proliferation and induces mitochondrial-dependent apoptosis through Akt, mTOR, and MEK-ERK pathways in human renal carcinoma cells. Phytomedicine 2017, 36, 95–107. [Google Scholar] [CrossRef]
- Um, H.J.; Oh, J.H.; Kim, Y.-N.; Choi, Y.H.; Kim, S.H.; Park, J.-W.; Kwon, T.K. The coffee diterpene kahweol sensitizes TRAIL-induced apoptosis in renal carcinoma Caki cells through down-regulation of Bcl-2 and c-FLIP. Chem. Interact. 2010, 186, 36–42. [Google Scholar] [CrossRef]
- Min, K.-J.; Um, H.J.; Kim, J.I.; Kwon, T.K. The coffee diterpene kahweol enhances sensitivity to sorafenib in human renal carcinoma Caki cells through down-regulation of Mcl-1 and c-FLIP expression. Oncotarget 2017, 8, 83195–83206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, Y.-H.; Lee, C.-H.; Chen, H.-Y.; Hsieh, S.-C.; Lin, C.-L.; Tsai, J.-P. Induction of cell cycle arrest, DNA damage, and apoptosis by nimbolide in human renal cell carcinoma cells. Tumor Biol. 2015, 36, 7539–7547. [Google Scholar] [CrossRef]
- Zheng, W.; Zhou, C.-Y.; Zhu, X.-Q.; Wang, X.-J.; Li, Z.-Y.; Chen, X.-C.; Chen, F.; Che, X.-Y.; Xie, X. Oridonin enhances the cytotoxicity of 5-FU in renal carcinoma cells by inducting necroptotic death. Biomed. Pharmacother. 2018, 106, 175–182. [Google Scholar] [CrossRef]
- Oka, D.; Nishimura, K.; Shiba, M.; Nakai, Y.; Arai, Y.; Nakayama, M.; Takayama, H.; Inoue, H.; Okuyama, A.; Nonomura, N. Sesquiterpene lactone parthenolide suppresses tumor growth in a xenograft model of renal cell carcinoma by inhibiting the activation of NF-κB. Int. J. Cancer 2007, 120, 2576–2581. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-R.; Chen, P.-N.; Yang, S.-F.; Sun, Y.-S.; Wu, S.-W.; Hung, T.-W.; Lian, J.-D.; Chu, S.-C.; Hsieh, Y.-S. Silibinin inhibits the invasion and migration of renal carcinoma 786-O cells in vitro, inhibits the growth of xenografts in vivo and enhances chemosensitivity to 5-fluorouracil and paclitaxel. Mol. Carcinog. 2011, 50, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dan, C.; Gong, X.; Zeng, W.; Yao, Q.; Yang, Y.; Gong, X.; Wang, L.; Chen, C.; Huang, L. Sorghumol triterpene inhibits the growth of circulating renal cancer cells by promoting cell apoptosis, G2/M cell cycle arrest and downregulating m-TOR/PI3K/AKT signalling pathway. J. BUON 2019, 24, 310–314. [Google Scholar] [PubMed]
- Park, E.J.; Chauhan, A.K.; Min, K.-J.; Park, D.C.; Kwon, T.K. Thymoquinone induces apoptosis through downregulation of c-FLIP and Bcl-2 in renal carcinoma Caki cells. Oncol. Rep. 2016, 36, 2261–2267. [Google Scholar] [CrossRef] [Green Version]
- Liou, Y.-F.; Hsieh, Y.-S.; Hung, T.-W.; Chen, P.-N.; Chang, Y.-Z.; Kao, S.-H.; Lin, S.-W.; Chang, H.-R. Thymoquinone inhibits metastasis of renal cell carcinoma cell 786-O-SI3 associating with downregulation of MMP-2 and u-PA and suppression of PI3K/Src signaling. Int. J. Med Sci. 2019, 16, 686–695. [Google Scholar] [CrossRef] [Green Version]
- Sourbier, C.; Scroggins, B.T.; Mannes, P.Z.; Liao, P.-J.; Siems, K.; Wolf, D.; Beutler, J.A.; Linehan, W.M.; Neckers, L. Tonantzitlolone cytotoxicity toward renal cancer cells is PKCθ- and HSF1-dependent. Oncotarget 2015, 6, 29963–29974. [Google Scholar] [CrossRef] [Green Version]
- Rubaiy, H.N.; Ludlow, M.J.; Siems, K.; Norman, K.; Foster, R.; Wolf, D.; Beutler, J.A.; Beech, D.J. Tonantzitlolone is a nanomolar potency activator of transient receptor potential canonical 1/4/5 channels. J. Cereb. Blood Flow Metab. 2018, 175, 3361–3368. [Google Scholar] [CrossRef] [Green Version]
- Brincks, E.L.; Kucaba, T.A.; James, B.R.; Murphy, K.; Schwertfeger, K.L.; Sangwan, V.; Banerjee, S.; Saluja, A.K.; Griffith, T.S. Triptolide enhances the tumoricidal activity of TRAIL against renal cell carcinoma. FEBS J. 2015, 282, 4747–4765. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhou, M.; Wang, X.; Liu, M.; Huang, H.; Yan, F.; Yu, Z.; Shu, X.; Huo, X.; Feng, L.; et al. Triptolidenol, isolated from Tripterygium wilfordii, disrupted NF-κB/COX-2 pathway by targeting ATP-binding sites of IKKβ in clear cell renal cell carcinoma. Fitoterapia 2020, 148, 104779. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sheng, Q.; Cheng, Y.; Xu, Y.; Han, Y.; Wang, J.; Shi, L.; Zhao, H.; Du, C. Zerumbone induces apoptosis in human renal cell carcinoma via Gli-1/Bcl-2 pathway. Die Pharmazie 2013, 68, 141–145. [Google Scholar] [PubMed]
- Shanmugam, M.K.; Rajendran, P.; Li, F.; Kim, C.; Sikka, S.; Siveen, K.S.; Kumar, A.P.; Ahn, K.S.; Sethi, G. Abrogation of STAT3 signaling cascade by zerumbone inhibits proliferation and induces apoptosis in renal cell carcinoma xenograft mouse model. Mol. Carcinog. 2014, 54, 971–985. [Google Scholar] [CrossRef]
- Song, J.; Kwon, Y.; Kim, S.; Lee, S.K. Antitumor Activity of Phenanthroindolizidine Alkaloids Is Associated with Negative Regulation of Met Endosomal Signaling in Renal Cancer Cells. Chem. Biol. 2015, 22, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Noh, H.-J.; Sung, E.-G.; Song, I.-H.; Kim, J.-Y.; Kwon, T.K. Berberine sensitizes TRAIL-induced apoptosis through proteasome-mediated downregulation of c-FLIP and Mcl-1 proteins. Int. J. Oncol. 2011, 38, 485–492. [Google Scholar] [CrossRef]
- Lopes, T.Z.; de Moraes, F.R.; Tedesco, A.C.; Arni, R.K.; Rahal, P.; Calmon, M.F. Berberine associated photodynamic therapy promotes autophagy and apoptosis via ROS generation in renal carcinoma cells. Biomed. Pharmacother. 2019, 123, 109794. [Google Scholar] [CrossRef]
- Shahriyar, S.A.; Woo, S.M.; Seo, S.U.; Min, K.-J.; Kwon, T.K. Cepharanthine Enhances TRAIL-Mediated Apoptosis through STAMBPL1-Mediated Downregulation of Survivin Expression in Renal Carcinoma Cells. Int. J. Mol. Sci. 2018, 19, 3280. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Zhuo, R.; Dai, J.; Wang, X.; Huang, X.; Wang, H.; Xu, D. Chelerythrine induces apoptosis via ROS-mediated endoplasmic reticulum stress and STAT3 pathways in human renal cell carcinoma. J. Cell. Mol. Med. 2019, 24, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Ren, Y.; Qiu, J. Dauricine inhibits viability and induces cell cycle arrest and apoptosis via inhibiting the PI3K/Akt signaling pathway in renal cell carcinoma cells. Mol. Med. Rep. 2018, 17, 7403–7408. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-A.; Sung, E.-G.; Song, I.-H.; Kim, J.-Y.; Sung, H.-J.; Sohn, H.-Y.; Park, J.-Y.; Lee, T.-J. Neferine-induced apoptosis is dependent on the suppression of Bcl-2 expression via downregulation of p65 in renal cancer cells. Acta Biochim. Biophys. Sin. 2019, 51, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Tosun, F.; Mıhoğlugil, F.; Beutler, J.A.A.; Özkan, E.E.; Miski, M. Neopapillarine, an Unusual Coumarino-Alkaloid from the Root Extract of Neocryptodiscus papillaris with Cytotoxic Activity on Renal Cancer Cells. Molecules 2020, 25, 3040. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, J.; Liu, Y.; Liu, Y.; Guo, G.; Yu, S.; An, R. Oxymatrine Inhibits Renal Cell Carcinoma Progression by Suppressing β-Catenin Expression. Front. Pharmacol. 2020, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Makhov, P.; Golovine, K.; Teper, E.; Kutikov, A.; Mehrazin, R.; Corcoran, A.T.; Tulin, A.V.; Uzzo, R.G.; Kolenko, V.M. Piperlongumine promotes autophagy via inhibition of Akt/mTOR signalling and mediates cancer cell death. Br. J. Cancer 2014, 110, 899–907. [Google Scholar] [CrossRef] [Green Version]
- Golovine, K.; Makhov, P.; Naito, S.; Raiyani, H.; Tomaszewski, J.; Mehrazin, R.; Tulin, A.; Kutikov, A.; Uzzo, R.G.; Kolenko, V.M. Piperlongumine and its analogs down-regulate expression of c-Met in renal cell carcinoma. Cancer Biol. Ther. 2015, 16, 743–749. [Google Scholar] [CrossRef]
- Chen, S.; Liu, W.; Wang, K.; Fan, Y.; Chen, J.; Ma, J.; Wang, X.; He, D.; Zeng, J.; Li, L. Tetrandrine inhibits migration and invasion of human renal cell carcinoma by regulating Akt/NF-κB/MMP-9 signaling. PLoS ONE 2017, 12, e0173725. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Shu, Y.; Cui, T.; Fu, P. Allicin inhibits human renal clear cell carcinoma progression via suppressing HIF pathway. Int. J. Clin. Exp. Med. 2015, 8, 20573–20580. [Google Scholar]
- Juengel, E.; Maxeiner, S.; Rutz, J.; Justin, S.; Roos, F.; Khoder, W.; Tsaur, I.; Nelson, K.; Bechstein, W.; Haferkamp, A.; et al. Sulforaphane inhibits proliferation and invasive activity of everolimus-resistant kidney cancer cells in vitro. Oncotarget 2016, 7, 85208–85219. [Google Scholar] [CrossRef] [Green Version]
- Juengel, E.; Euler, S.; Maxeiner, S.; Rutz, J.; Justin, S.; Roos, F.; Khoder, W.; Nelson, K.; Bechstein, W.; Blaheta, R.A. Sulforaphane as an adjunctive to everolimus counteracts everolimus resistance in renal cancer cell lines. Phytomedicine 2017, 27, 1–7. [Google Scholar] [CrossRef]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Lateef, A.; Hamiza, O.O.; Qamar, W.; Ali, F.; Sultana, S. Chrysin suppresses renal carcinogenesis via amelioration of hyperproliferation, oxidative stress and inflammation: Plausible role of NF-κB. Toxicol. Lett. 2012, 216, 146–158. [Google Scholar] [CrossRef]
- Ruiz-Marcial, C.; Reyes-Chilpa, R.; Estrada, E.; Esparza, J.A.R.; Fariña, G.G.; Fragoso, L.R. Antiproliferative, cytotoxic and antitumour activity of coumarins isolated from Calophyllum brasiliense. J. Pharm. Pharmacol. 2007, 59, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, S.; Liao, W.; Xiong, Y. Modification of Antitumor Immunity and Tumor Microenvironment by Resveratrol in Mouse Renal Tumor Model. Cell Biochem. Biophys. 2015, 72, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Kabel, A.M.; Atef, A.; Estfanous, R.S. Ameliorative potential of sitagliptin and/or resveratrol on experimentally-induced clear cell renal cell carcinoma. Biomed. Pharmacother. 2018, 97, 667–674. [Google Scholar] [CrossRef]
- Sahin, K.; Cross, B.; Sahin, N.; Ciccone, K.; Suleiman, S.; Osunkoya, A.O.; Master, V.; Harris, W.; Carthon, B.; Mohammad, R.; et al. Lycopene in the prevention of renal cell cancer in the TSC2 mutant Eker rat model. Arch. Biochem. Biophys. 2015, 572, 36–39. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Khan, H.; Aschner, M.; Mirzae, H.; Akkol, E.K.; Capasso, R. Anticancer Potential of Furanocoumarins: Mechanistic and Therapeutic Aspects. Int. J. Mol. Sci. 2020, 21, 5622. [Google Scholar] [CrossRef]
- Mahendra, C.K.; Tan, L.T.H.; Lee, W.L.; Yap, W.H.; Pusparajah, P.; Low, L.E.; Tang, S.Y.; Chan, K.G.; Lee, L.H.; Goh, B.H. Angelicin—A Furocoumarin Compound with Vast Biological Potential. Front. Pharmacol. 2020, 11, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.A.; Parama, D.; Daimari, E.; Girisa, S.; Banik, K.; Harsha, C.; Dutta, U.; Kunnumakkara, A.B. Rationalizing the therapeutic potential of apigenin against cancer. Life Sci. 2020, 267, 118814. [Google Scholar] [CrossRef] [PubMed]
- Javed, Z.; Sadia, H.; Iqbal, M.J.; Shamas, S.; Malik, K.; Ahmed, R.; Raza, S.; Butnariu, M.; Cruz-Martins, N.; Sharifi-Rad, J. Apigenin role as cell-signaling pathways modulator: Implications in cancer prevention and treatment. Cancer Cell Int. 2021, 21, 189. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, E.R.; Ang, H.L.; Asnaf, S.E.; Zabolian, A.; Saleki, H.; Yavari, M.; Esmaeili, H.; Zarrabi, A.; Ashrafizadeh, M.; Kumar, A.P. Broad-Spectrum Preclinical Antitumor Activity of Chrysin: Current Trends and Future Perspectives. Biomolecules 2020, 10, 1374. [Google Scholar] [CrossRef]
- Talebi, M.; Talebi, M.; Farkhondeh, T.; Simal-Gandara, J.; Kopustinskiene, D.M.; Bernatoniene, J.; Samarghandian, S. Emerging cellular and molecular mechanisms underlying anticancer indications of chrysin. Cancer Cell Int. 2021, 21, 214. [Google Scholar] [CrossRef]
- Annunziata, F.; Pinna, C.; Dallavalle, S.; Tamborini, L.; Pinto, A. An Overview of Coumarin as a Versatile and Readily Accessible Scaffold with Broad-Ranging Biological Activities. Int. J. Mol. Sci. 2020, 21, 4618. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xu, J.; Liu, Y.; Zeng, Y.; Wu, G. A Review on Anti-Tumor Mechanisms of Coumarins. Front. Oncol. 2020, 10, 592853. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Sundaram, C.; Jhurani, S.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin and cancer: An “old-age” disease with an “age-old” solution. Cancer Lett. 2008, 267, 133–164. [Google Scholar] [CrossRef]
- Sinha, D.; Biswas, J.; Sung, B.; Aggarwal, B.B.; Bishayee, A. Chemopreventive and Chemotherapeutic Potential of Curcumin in Breast Cancer. Curr. Drug Targets 2012, 13, 1799–1819. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Tommonaro, G. Curcumin and Cancer. Nutrients 2019, 11, 2376. [Google Scholar] [CrossRef] [Green Version]
- Kasi, P.D.; Tamilselvam, R.; Skalicka-Woźniak, K.; Nabavi, S.M.; Daglia, M.; Bishayee, A.; Pazoki-Toroudi, H. Molecular targets of curcumin for cancer therapy: An updated review. Tumor Biol. 2016, 37, 13017–13028. [Google Scholar] [CrossRef]
- Thomasset, S.; Teller, N.; Cai, H.; Marko, D.; Berry, D.P.; Steward, W.P.; Gescher, A.J. Do anthocyanins and anthocyanidins, cancer chemopreventive pigments in the diet, merit development as potential drugs? Cancer Chemother. Pharmacol. 2009, 64, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, B.; Sun, J.; Jiang, X.; Bai, W. Anthocyanin supplement as a dietary strategy in cancer prevention and management: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2021, 1–13. [Google Scholar] [CrossRef]
- Pinto, D.C.; Silva, A.M. Anticancer Natural Coumarins as Lead Compounds for the Discovery of New Drugs. Curr. Top. Med. Chem. 2018, 17, 1. [Google Scholar] [CrossRef]
- Timson, D. Dicoumarol: A Drug which Hits at Least Two Very Different Targets in Vitamin K Metabolism. Curr. Drug Targets 2017, 18, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhao, W.; Wang, X.; Sun, Y.; Chen, X. A pharmacological review of dicoumarol: An old natural anticoagulant agent. Pharmacol. Res. 2020, 160, 105193. [Google Scholar] [CrossRef] [PubMed]
- Kadakol, A.; Sharma, N.; Kulkarni, Y.A.; Gaikwad, A.B. Esculetin: A phytochemical endeavor fortifying effect against non-communicable diseases. Biomed. Pharmacother. 2016, 84, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Park, C.; Chen, H.; Hwang, J.; Thimmegowda, N.; Bae, E.Y.; Lee, K.W.; Kim, H.-G.; Liu, H.; Soung, N.K.; et al. Eupafolin suppresses prostate cancer by targeting phosphatidylinositol 3-kinase-mediated Akt signaling. Mol. Carcinog. 2014, 54, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Tao, J.; Cai, X.; Fredimoses, M.; Wu, J.; Jiang, Z.; Zhang, X.; Li, S. Eupafolin Suppresses Esophagus Cancer Growth by Targeting T-LAK Cell-Originated Protein Kinase. Front. Pharmacol. 2019, 10, 1248. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, X.; Pan, H.; He, S.; Yuan, B.; Liu, Q.; Zhang, J.; Ding, Y. Eupafolin inhibits breast cancer cell proliferation and induces apoptosis by inhibiting the PI3K/Akt/mTOR pathway. Oncol. Lett. 2021, 21, 332. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Y.; Lin, L.; Li, H. Gambogic Acid as a Candidate for Cancer Therapy: A Review. Int. J. Nanomed. 2020, 15, 10385–10399. [Google Scholar] [CrossRef] [PubMed]
- Hatami, E.; Jaggi, M.; Chauhan, S.C.; Yallapu, M.M. Gambogic acid: A shining natural compound to nanomedicine for cancer therapeutics. Biochim. Biophys. Acta 2020, 1874, 188381. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, C.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Sureda, A.; Nabavi, S.F.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; et al. Genistein and Cancer: Current Status, Challenges, and Future Directions. Adv. Nutr. 2015, 6, 408–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [Green Version]
- Ashaq, A.; Maqbool, M.F.; Maryam, A.; Khan, M.; Shakir, H.A.; Irfan, M.; Qazi, J.I.; Li, Y.; Ma, T. Hispidulin: A novel natural compound with therapeutic potential against human cancers. Phytother. Res. 2020, 35, 771–789. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, F.; Yan, J.; Xia, Z.; Jiang, D.; Ma, P. Hispidulin: A promising flavonoid with diverse anti-cancer properties. Life Sci. 2020, 259, 118395. [Google Scholar] [CrossRef] [PubMed]
- Banik, K.; Ranaware, A.M.; Deshpande, V.; Nalawade, S.P.; Padmavathi, G.; Bordoloi, D.; Sailo, B.L.; Shanmugam, M.K.; Fan, L.; Arfuso, F.; et al. Honokiol for cancer therapeutics: A traditional medicine that can modulate multiple oncogenic targets. Pharmacol. Res. 2019, 144, 192–209. [Google Scholar] [CrossRef]
- Ong, C.P.; Lee, W.L.; Tang, Y.Q.; Yap, W.H. Honokiol: A Review of Its Anticancer Potential and Mechanisms. Cancers 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5280567, Hymecromone. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Hymecromone (accessed on 3 April 2022).
- Nagy, N.; Kuipers, H.F.; Frymoyer, A.R.; Ishak, H.D.; Bollyky, J.B.; Wight, T.N.; Bollyky, P.L. 4-Methylumbelliferone Treatment and Hyaluronan Inhibition as a Therapeutic Strategy in Inflammation, Autoimmunity, and Cancer. Front. Immunol. 2015, 6, 123. [Google Scholar] [CrossRef] [Green Version]
- Abdulaziz, N.T.; Mustafa, Y.F. Anticancer properties of hymecromone-derived compounds: A review. Int. Pharm. Res. 2020, 13, 2163–2174. [Google Scholar] [CrossRef]
- Zhang, C.; Sui, X.; Jiang, Y.; Wang, X.; Wang, S. Antitumor effects of icaritin and the molecular mechanisms. Discov. Med. 2020, 29, 7–18. [Google Scholar]
- Zhao, T.-T.; Xu, Y.-Q.; Hu, H.-M.; Gong, H.-B.; Zhu, H.-L. Isoliquiritigenin (ISL) and its Formulations: Potential Antitumor Agents. Curr. Med. Chem. 2019, 26, 6786–6796. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a flavonoid, as an anticancer agent: A review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer activity of the plant flavonoid luteolin against preclinical models of various cancers and insights on different signalling mechanisms modulated. Phytother. Res. 2021, 35, 3509–3532. [Google Scholar] [CrossRef]
- Zhang, Z.-R.; Leung, W.N.; Cheung, H.Y.; Chan, C.W. Osthole: A Review on Its Bioactivities, Pharmacological Properties, and Potential as Alternative Medicine. Evid.-Based Complementary Altern. Med. 2015, 2015, 919616. [Google Scholar] [CrossRef] [Green Version]
- Shokoohinia, Y.; Jafari, F.; Mohammadi, Z.; Bazvandi, L.; Hosseinzadeh, L.; Chow, N.; Bhattacharyya, P.; Farzaei, M.H.; Farooqi, A.A.; Nabavi, S.M.; et al. Potential Anticancer Properties of Osthol: A Comprehensive Mechanistic Review. Nutrients 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Ashrafizadeh, M.; Mohammadinejad, R.; Samarghandian, S.; Yaribeygi, H.; Johnston, T.P.; Sahebkar, A. Anti-Tumor Effects of Osthole on Different Malignant Tissues: A Review of Molecular Mechanisms. Anti-Cancer Agents Med. Chem. 2020, 20, 918–931. [Google Scholar] [CrossRef]
- Zafar, S.; Sarfraz, I.; Rasul, A.; Shah, M.A.; Hussain, G.; Zahoor, M.K.; Shafiq, N.; Riaz, A.; Selamoglu, Z.; Sarker, S.D. Osthole: A Multifunctional Natural Compound with Potential Anticancer, Antioxidant and Anti-inflammatory Activities. Mini-Rev. Med. Chem. 2021, 21, 2747–2763. [Google Scholar] [CrossRef] [PubMed]
- Sarkhail, P.; Shafiee, A.; Sarkheil, P. Biological Activities and Pharmacokinetics of Praeruptorins from Peucedanum Species: A Systematic Review. BioMed Res. Int. 2013, 2013, 343808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.-L.; Hung, T.-W.; Ying, T.-H.; Lin, C.-J.; Hsieh, Y.-H.; Chen, C.-M. Praeruptorin B Mitigates the Metastatic Ability of Human Renal Carcinoma Cells through Targeting CTSC and CTSV Expression. Int. J. Mol. Sci. 2020, 21, 2919. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhang, X.; Xu, L.; Liu, D.; Di, S.; Li, W.; Zhang, J.; Zhang, H.; Li, X.; Han, J.; et al. Pterostilbene: Mechanisms of its action as oncostatic agent in cell models and in vivo studies. Pharmacol. Res. 2019, 145, 104265. [Google Scholar] [CrossRef]
- Lin, W.-S.; Leland, J.V.; Ho, C.-T.; Pan, M.-H. Occurrence, Bioavailability, Anti-inflammatory, and Anticancer Effects of Pterostilbene. J. Agric. Food Chem. 2020, 68, 12788–12799. [Google Scholar] [CrossRef]
- Obrador, E.; Salvador-Palmer, R.; Jihad-Jebbar, A.; López-Blanch, R.; Dellinger, T.; Dellinger, R.; Estrela, J. Pterostilbene in Cancer Therapy. Antioxidants 2021, 10, 492. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A. Cancer Prevention and Treatment with Resveratrol: From Rodent Studies to Clinical Trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Li, M.; Tang, C.; Huang, Z.; Najafi, M. Targeting of cancer cell death mechanisms by resveratrol: A review. Apoptosis 2021, 26, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Mazimba, O. Umbelliferone: Sources, chemistry and bioactivities review. Bull. Fac. Pharm. Cairo Univ. 2017, 55, 223–232. [Google Scholar] [CrossRef]
- Garg, S.S.; Gupta, J.; Sharma, S.; Sahu, D. An insight into the therapeutic applications of coumarin compounds and their mechanisms of action. Eur. J. Pharm. Sci. 2020, 152, 105424. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T. From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 2017, 46, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, J.; Zhao, X. Mechanisms and Molecular Targets of Artemisinin in Cancer Treatment. Cancer Investig. 2021, 39, 675–684. [Google Scholar] [CrossRef]
- Khanal, P. Antimalarial and anticancer properties of artesunate and other artemisinins: Current development. Mon. Chem. 2021, 152, 387–400. [Google Scholar] [CrossRef]
- Yang, X.; Zheng, Y.; Liu, L.; Huang, J.; Wang, F.; Zhang, J. Progress on the study of the anticancer effects of artesunate (Review). Oncol. Lett. 2021, 22, 750. [Google Scholar] [CrossRef]
- Bibak, B.; Shakeri, F.; Barreto, G.E.; Keshavarzi, Z.; Sathyapalan, T.; Sahebkar, A. A review of the pharmacological and therapeutic effects of auraptene. BioFactors 2019, 45, 867–879. [Google Scholar] [CrossRef]
- Tayarani-Najaran, Z.; Tayarani-Najaran, N.; Eghbali, S. A Review of Auraptene as an Anticancer Agent. Front. Pharmacol. 2021, 12, 698352. [Google Scholar] [CrossRef]
- Zhai, B.; Zhang, N.; Han, X.; Li, Q.; Zhang, M.; Chen, X.; Li, G.; Zhang, R.; Chen, P.; Wang, W.; et al. Molecular targets of β-elemene, a herbal extract used in traditional Chinese medicine, and its potential role in cancer therapy: A review. Biomed. Pharmacother. 2019, 114, 108812. [Google Scholar] [CrossRef]
- Bai, Z.; Yao, C.; Zhu, J.; Xie, Y.; Ye, X.-Y.; Bai, R.; Xie, T. Anti-Tumor Drug Discovery Based on Natural Product β-Elemene: Anti-Tumor Mechanisms and Structural Modification. Molecules 2021, 26, 1499. [Google Scholar] [CrossRef] [PubMed]
- Amiri, S.; Dastghaib, S.; Ahmadi, M.; Mehrbod, P.; Khadem, F.; Behrouj, H.; Aghanoori, M.R.; Machaj, F.; Ghamsari, M.; Rosik, J.; et al. Betulin and its derivatives as novel compounds with different pharmacological effects. Biotechnol. Adv. 2020, 38, 107409. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Li, X.; Dong, S.; Zhou, W. Betulinic acid in the treatment of tumour diseases: Application and research progress. Biomed. Pharmacother. 2021, 142, 111990. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sak, K.; Gupta, D.S.; Kaur, G.; Aggarwal, D.; Parashar, N.C.; Choudhary, R.; Yerer, M.B.; Kaur, J.; Kumar, M.; et al. Anti-Inflammatory and Anticancer Properties of Birch Bark-Derived Betulin: Recent Developments. Plants 2021, 10, 2663. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Patel, D. Ocimum Sanctum: An All-Round Treatment for Cancer? Altern. Health Med. 2021; Online ahead of print. [Google Scholar]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Grant, S. Targeting Multiple Arms of the Apoptotic Regulatory Machinery. Cancer Res. 2007, 67, 2908–2911. [Google Scholar] [CrossRef] [Green Version]
- Petiwala, S.M.; Johnson, J.J. Diterpenes from rosemary (Rosmarinus officinalis): Defining their potential for anti-cancer activity. Cancer Lett. 2015, 367, 93–102. [Google Scholar] [CrossRef]
- Kumar, P.; Bhadauria, A.S.; Singh, A.K.; Saha, S. Betulinic acid as apoptosis activator: Molecular mechanisms, mathematical modeling and chemical modifications. Life Sci. 2018, 209, 24–33. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, H.; An, Y.; Shen, K.; Yu, L. Biological effects of corosolic acid as an anti-inflammatory, anti-metabolic syndrome and anti-neoplasic natural compound (Review). Oncol. Lett. 2020, 21, 84. [Google Scholar] [CrossRef]
- Arzi, L.; Hoshyar, R. Saffron anti-metastatic properties, ancient spice novel application. Crit. Rev. Food Sci. Nutr. 2021, 62, 3939–3950. [Google Scholar] [CrossRef]
- Veisi, A.; Akbari, G.; Mard, A.; Badfar, G.; Zarezade, V.; Mirshekar, M.A. Role of crocin in several cancer cell lines: An updated review. Iran J. Basic Med. Sci. 2020, 23, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Lambrianidou, A.; Koutsougianni, F.; Papapostolou, I.; Dimas, K. Recent Advances on the Anticancer Properties of Saffron (Crocus sativus L.) and Its Major Constituents. Molecules 2020, 26, 86. [Google Scholar] [CrossRef]
- Cai, Y.; Fang, X.; He, C.; Li, P.; Xiao, F.; Wang, Y.; Chen, M. Cucurbitacins: A Systematic Review of the Phytochemistry and Anticancer Activity. Am. J. Chin. Med. 2015, 43, 1331–1350. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Green, I.R.; Saleem, M.; Khattak, K.F.; Irshad, M.; Ali, M. Cucurbitacins as Anticancer Agents: A Patent Review. Recent Pat. Anti-Cancer Drug Discov. 2019, 14, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhao, S.; Fash, D.M.; Li, Z.; Chain, W.J.; Beutler, J.A. Englerins: A Comprehensive Review. J. Nat. Prod. 2017, 80, 771–781. [Google Scholar] [CrossRef]
- Cheong, D.H.; Arfuso, F.; Sethi, G.; Wang, L.; Hui, K.M.; Kumar, A.P.; Tran, T. Molecular targets and anti-cancer potential of escin. Cancer Lett. 2018, 422, 1–8. [Google Scholar] [CrossRef]
- Kriplani, P. Recent Patents on Anti-Cancer Potential of Helenalin. Recent Pat. Anti-Cancer Drug Discov. 2020, 15, 132–142. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, W.; Yu, X.; Wang, F.; Li, Y.; Zhang, Y.; Yang, Y. Helenalin Facilitates Reactive Oxygen Species-Mediated Apoptosis and Cell Cycle Arrest by Targeting Thioredoxin Reductase-1 in Human Prostate Cancer Cells. Med. Sci. Monit. 2021, 27, e930083. [Google Scholar] [CrossRef]
- Drogosz, J.; Janecka, A. Helenalin—A Sesquiterpene Lactone with Multidirectional Activity. Curr. Drug Targets 2019, 20, 444–452. [Google Scholar] [CrossRef]
- Chang, F.-R.; Hwang, T.-L.; Yang, Y.-L.; Li, C.-E.; Wu, C.-C.; Issa, H.; Hsieh, W.-B.; Wu, Y.-C. Anti-Inflammatory and Cytotoxic Diterpenes from Formosan Polyalthia longifolia var. pendula. Planta Med. 2006, 72, 1344–1347. [Google Scholar] [CrossRef]
- Thiyagarajan, V.; Lin, S.-H.; Chia, Y.-C.; Weng, C.-F. A novel inhibitor, 16-hydroxy-cleroda-3,13-dien-16,15-olide, blocks the autophosphorylation site of focal adhesion kinase (Y397) by molecular docking. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 4091–4101. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Hwang, Y.P.; Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Chung, Y.H.; Jeong, H.G. Kahweol inhibits proliferation and induces apoptosis by suppressing fatty acid synthase in HER2-overexpressing cancer cells. Food Chem. Toxicol. 2018, 121, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Rengasamy, K.R.; Mahomoodally, F.M.; Keum, Y.-S. Protective effects of lycopene in cancer, cardiovascular, and neurodegenerative diseases: An update on epidemiological and mechanistic perspectives. Pharmacol. Res. 2020, 155, 104730. [Google Scholar] [CrossRef] [PubMed]
- Marzocco, S.; Singla, R.K.; Capasso, A. Multifaceted Effects of Lycopene: A Boulevard to the Multitarget-Based Treatment for Cancer. Molecules 2021, 26, 5333. [Google Scholar] [CrossRef]
- Patel, S.M.; Venkata, K.C.N.; Bhattacharyya, P.; Sethi, G.; Bishayee, A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin. Cancer Biol. 2016, 40–41, 100–115. [Google Scholar] [CrossRef]
- Nagini, S.; Nivetha, R.; Palrasu, M.; Mishra, R. Nimbolide, a Neem Limonoid, Is a Promising Candidate for the Anticancer Drug Arsenal. J. Med. Chem. 2021, 64, 3560–3577. [Google Scholar] [CrossRef]
- Liu, X.; Xu, J.; Zhou, J.; Shen, Q. Oridonin and its derivatives for cancer treatment and overcoming therapeutic resistance. Genes Dis. 2020, 8, 448–462. [Google Scholar] [CrossRef]
- Kou, B.; Yang, Y.; Bai, Y.-E.; Shi, Y.-H.; Gao, R.-X.; Yang, F.-L.; Zhang, S.-Q.; Liu, W. Oridonin Induces Apoptosis of Laryngeal Carcinoma via Endoplasmic Reticulum Stress. Cancer Manag. Res. 2020, 12, 8387–8396. [Google Scholar] [CrossRef]
- Freund, R.R.A.; Gobrecht, P.; Fischer, D.; Arndt, H.-D. Advances in chemistry and bioactivity of parthenolide. Nat. Prod. Rep. 2019, 37, 541–565. [Google Scholar] [CrossRef]
- Sztiller-Sikorska, M.; Czyz, M. Parthenolide as Cooperating Agent for Anti-Cancer Treatment of Various Malignancies. Pharmaceuticals 2020, 13, 194. [Google Scholar] [CrossRef]
- Mastron, J.K.; Siveen, K.S.; Sethi, G.; Bishayee, A. Silymarin and hepatocellular carcinoma: A systematic, comprehensive, and critical review. Anti-Cancer Drugs 2015, 26, 475–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuli, H.S.; Mittal, S.; Aggarwal, D.; Parashar, G.; Parashar, N.C.; Upadhyay, S.K.; Barwal, T.S.; Jain, A.; Kaur, G.; Savla, R.; et al. Path of Silibinin from diet to medicine: A dietary polyphenolic flavonoid having potential anti-cancer therapeutic significance. Semin. Cancer Biol. 2020, 73, 196–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-N.; Cai, J.-Y.; Zhao, L.; Zhu, E.; Zhang, D.-Z. Study on chemical constituents from Anoectochilus chapaensis. Zhong yao cai = Zhongyaocai = J. Chin. Med. Mater. 2012, 35, 911–913. [Google Scholar]
- Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Wang, L.; Goh, B.C.; Ahn, K.S.; Bishayee, A.; Sethi, G. Modulation of diverse oncogenic transcription factors by thymoquinone, an essential oil compound isolated from the seeds of Nigella sativa Linn. Pharmacol. Res. 2018, 129, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Almajali, B.; Al-Jamal, H.; Taib, W.; Ismail, I.; Johan, M.; Doolaanea, A.; Ibrahim, W. Thymoquinone, as a Novel Therapeutic Candidate of Cancers. Pharmaceuticals 2021, 14, 369. [Google Scholar] [CrossRef] [PubMed]
- Busch, T.; Dräger, G.; Kunst, E.; Benson, H.; Sasse, F.; Siems, K.; Kirschning, A. Synthesis and antiproliferative activity of new tonantzitlolone-derived diterpene derivatives. Org. Biomol. Chem. 2016, 14, 9040–9045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Abrantes, R.A.; Batista, T.M.; Mangueira, V.M.; de Sousa, T.K.G.; Ferreira, R.C.; Moura, A.P.G.; Abreu, L.S.; Alves, A.F.; Velozo, E.S.; Batista, L.M.; et al. Antitumor and antiangiogenic effects of Tonantzitlolone B, an uncommon diterpene from Stillingia loranthacea. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 395, 267–274. [Google Scholar] [CrossRef]
- Sun, X.; Yan, P. Triptolide: A new star for treating human malignancies. J. Cancer Res. Ther. 2018, 14, S271–S275. [Google Scholar] [CrossRef]
- Noel, P.; Von Hoff, D.D.; Saluja, A.K.; Velagapudi, M.; Borazanci, E.; Han, H. Triptolide and Its Derivatives as Cancer Therapies. Trends Pharmacol. Sci. 2019, 40, 327–341. [Google Scholar] [CrossRef]
- Tong, L.; Zhao, Q.; Datan, E.; Lin, G.-Q.; Minn, I.; Pomper, M.G.; Yu, B.; Romo, D.; He, Q.-L.; Liu, J.O. Triptolide: Reflections on two decades of research and prospects for the future. Nat. Prod. Rep. 2020, 38, 843–860. [Google Scholar] [CrossRef] [PubMed]
- Girisa, S.; Shabnam, B.; Monisha, J.; Fan, L.; Halim, C.E.; Arfuso, F.; Ahn, K.S.; Sethi, G.; Kunnumakkara, A.B. Potential of Zerumbone as an Anti-Cancer Agent. Molecules 2019, 24, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechner, J.F.; Stoner, G.D. Gingers and Their Purified Components as Cancer Chemopreventative Agents. Molecules 2019, 24, 2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Kim, G.D.; Kim, S.; Lee, S.K. Antofine, a natural phenanthroindolizidine alkaloid, suppresses angiogenesis via regulation of AKT/mTOR and AMPK pathway in endothelial cells and endothelial progenitor cells derived from mouse embryonic stem cells. Food Chem. Toxicol. 2017, 107, 201–207. [Google Scholar] [CrossRef]
- Jia, X.-H.; Zhao, H.-X.; Du, C.-L.; Tang, W.-Z.; Wang, X.-J. Possible pharmaceutical applications can be developed from naturally occurring phenanthroindolizidine and phenanthroquinolizidine alkaloids. Phytochem. Rev. 2020, 20, 845–868. [Google Scholar] [CrossRef] [PubMed]
- Samadi, P.; Sarvarian, P.; Gholipour, E.; Asenjan, K.S.; Aghebati-Maleki, L.; Motavalli, R.; Hojjat-Farsangi, M.; Yousefi, M. Berberine: A novel therapeutic strategy for cancer. IUBMB Life 2020, 72, 2065–2079. [Google Scholar] [CrossRef]
- Rauf, A.; Abu-Izneid, T.; Khalil, A.A.; Imran, M.; Shah, Z.A.; Bin Emran, T.; Mitra, S.; Khan, Z.; Alhumaydhi, F.A.; Aljohani, A.S.M.; et al. Berberine as a Potential Anticancer Agent: A Comprehensive Review. Molecules 2021, 26, 7368. [Google Scholar] [CrossRef] [PubMed]
- Devarajan, N.; Jayaraman, S.; Mahendra, J.; Venkatratnam, P.; Rajagopal, P.; Palaniappan, H.; Ganesan, S.K. Berberine—A potent chemosensitizer and chemoprotector to conventional cancer therapies. Phytother. Res. 2021, 35, 3059–3077. [Google Scholar] [CrossRef]
- Agostinis, P.; Berg, K.; Cengel, K.A.; Foster, T.H.; Girotti, A.W.; Gollnick, S.O.; Hahn, S.M.; Hamblin, M.R.; Juzeniene, A.; Kessel, D.; et al. Photodynamic therapy of cancer: An update. CA Cancer J. Clin. 2011, 61, 250–281. [Google Scholar] [CrossRef]
- Osaki, T.; Yokoe, I.; Takahashi, K.; Inoue, K.; Ishizuka, M.; Tanaka, T.; Azuma, K.; Murahata, Y.; Tsuka, T.; Itoh, N.; et al. Metformin enhances the cytotoxicity of 5-aminolevulinic acid-mediated photodynamic therapy in vitro. Oncol. Lett. 2017, 14, 1049–1053. [Google Scholar] [CrossRef]
- Gülçin, I.; Elias, R.; Gepdiremen, A.; Chea, A.; Topal, F. Antioxidant activity of bisbenzylisoquinoline alkaloids from Stephania rotunda: Cepharanthine and fangchinoline. J. Enzym. Inhib. Med. Chem. 2009, 25, 44–53. [Google Scholar] [CrossRef]
- Rogosnitzky, M.; Danks, R. Therapeutic potential of the biscoclaurine alkaloid, cepharanthine, for a range of clinical conditions. Pharmacol. Rep. 2011, 63, 337–347. [Google Scholar] [CrossRef]
- Bailly, C. Cepharanthine: An update of its mode of action, pharmacological properties and medical applications. Phytomedicine 2019, 62, 152956. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.-H.; Cao, W.-X.; Wang, Z.-Y.; Lu, J.; Liu, B.; Chen, X.; Lu, J.-J. Induction of reactive oxygen species-stimulated distinctive autophagy by chelerythrine in non-small cell lung cancer cells. Redox Biol. 2017, 12, 367–376. [Google Scholar] [CrossRef]
- Wu, S.; Yang, Y.; Li, F.; Huang, L.; Han, Z.; Wang, G.; Yu, H.; Li, H. Chelerythrine induced cell death through ROS-dependent ER stress in human prostate cancer cells. OncoTargets Ther. 2018, 11, 2593–2601. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Huang, J.; Yuan, Z.; Feng, S.; Xie, Y.; Ma, W. Protein kinase C inhibitor chelerythrine selectively inhibits proliferation of triple-negative breast cancer cells. Sci. Rep. 2017, 7, 2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.-Y.; Jiang, S.-Q.; Zhang, L.; Liu, Q.-N.; Gong, P.-L. Inhibitory Effect of Dauricine on Inflammatory Process Following Focal Cerebral Ischemia/Reperfusion in Rats. Am. J. Chin. Med. 2007, 35, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.; Lee, E.; Lee, K.W. Therapeutic Implications of Autophagy Inducers in Immunological Disorders, Infection, and Cancer. Int. J. Mol. Sci. 2017, 18, 1959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fei, H.; Guo, J.; Zhang, X.; Wu, S.; Zhong, L. Dauricine suppresses the growth of pancreatic cancer in vivo by modulating the Hedgehog signaling pathway. Oncol. Lett. 2019, 18, 4403–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asokan, S.M.; Mariappan, R.; Muthusamy, S.; Velmurugan, B.K. Pharmacological benefits of neferine—A comprehensive review. Life Sci. 2018, 199, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Manogaran, P.; Beeraka, N.M.; Padma, V.V. The Cytoprotective and Anti-cancer Potential of Bisbenzylisoquinoline Alkaloids from Nelumbo nucifera. Curr. Top. Med. Chem. 2020, 19, 2940–2957. [Google Scholar] [CrossRef] [PubMed]
- Priya, L.B.; Huang, C.; Hu, R.; Balasubramanian, B.; Baskaran, R. An updated review on pharmacological properties of neferine—A bisbenzylisoquinoline alkaloid from Nelumbo nucifera. J. Food Biochem. 2021, 45, e13986. [Google Scholar] [CrossRef]
- Bishayee, A.; Patel, P.A.; Sharma, P.; Thoutireddy, S.; Das, N. Lotus (Nelumbo nucifera Gaertn.) and Its Bioactive Phytocompounds: A Tribute to Cancer Prevention and Intervention. Cancers 2022, 14, 529. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hu, Y.; Shen, Y.-F.; Wang, G.-X.; Zhu, B. Evaluation on antiviral activity of coumarin derivatives against spring viraemia of carp virus in epithelioma papulosum cyprini cells. Antivir. Res. 2017, 144, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Song, H.Y.; Jo, A.; Shin, J.; Lim, E.H.; Lee, Y.E.; Jeong, D.E. Anti-Inflammatory Activities of Isogosferol, a Furanocoumarin Isolated from Citrus junos Seed Shells through Bioactivity-Guided Fractionation. Molecules 2019, 24, 4088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Pandey, A.; Manvati, S. Coumarin: An emerging antiviral agent. Heliyon 2020, 6, e03217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong-Gang, C.; Shan, J.; Lei, L.; Jing-Quan, G.; Zhi-Ying, S.; Yan, L.; Yan, X.; Ming-Li, W.; Ye, W.; Chang-Qing, X.; et al. Antiarrhythmic effects and ionic mechanisms of oxymatrine from Sophora flavescens. Phytother. Res. 2010, 24, 1844–1849. [Google Scholar] [CrossRef]
- Cao, Z.; Yang, P.; Zhou, Q. Multiple biological functions and pharmacological effects of lycorine. Sci. China Ser. B Chem. 2013, 56, 1382–1391. [Google Scholar] [CrossRef]
- Halim, C.E.; Xinjing, S.L.; Fan, L.; Vitarbo, J.B.; Arfuso, F.; Tan, C.H.; Narula, A.S.; Kumar, A.P.; Sethi, G.; Ahn, K.S. Anti-cancer effects of oxymatrine are mediated through multiple molecular mechanism(s) in tumor models. Pharmacol. Res. 2019, 147, 104327. [Google Scholar] [CrossRef]
- Lan, X.; Zhao, J.; Zhang, Y.; Chen, Y.; Liu, Y.; Xu, F. Oxymatrine exerts organ- and tissue-protective effects by regulating inflammation, oxidative stress, apoptosis, and fibrosis: From bench to bedside. Pharmacol. Res. 2019, 151, 104541. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Attar, R.; Yaylim, I.; Qureshi, M.Z.; Todorovska, M.; Karatoprak, G.; Najafi, S.; Sabitaliyevich, U.Y.; Zhenisovna, T.G.; De Sousa, D.P.; et al. Piperlongumine as anticancer agent: The story so far about killing many birds with one stone. Cell. Mol. Biol. 2018, 64, 102–107. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Biswal, B.K. Piperlongumine, a potent anticancer phytotherapeutic: Perspectives on contemporary status and future possibilities as an anticancer agent. Pharmacol. Res. 2020, 156, 104772. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Qian, J.; Xu, Z.; Meng, C.; Zhu, W.; Ran, F.; Zhang, W.; Zhang, Y.; Ling, Y. Overview of piperlongumine analogues and their therapeutic potential. Eur. J. Med. Chem. 2021, 220, 113471. [Google Scholar] [CrossRef] [PubMed]
- Bhagya, N.; Chandrashekar, K.R. Tetrandrine and cancer—An overview on the molecular approach. Biomed. Pharmacother. 2017, 97, 624–632. [Google Scholar] [CrossRef]
- Luan, F.; He, X.; Zeng, N. Tetrandrine: A review of its anticancer potentials, clinical settings, pharmacokinetics and drug delivery systems. J. Pharm. Pharmacol. 2020, 72, 1491–1512. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Wong, S.K.; Chan, H.T. An overview on the chemistry, pharmacology and anticancer properties of tetrandrine and fangchinoline (alkaloids) from Stephania tetrandra roots. J. Integr. Med. 2021, 19, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Mondal, A.; Banerjee, S.; Bose, S.; Mazumder, S.; Haber, R.A.; Farzaei, M.H.; Bishayee, A. Garlic constituents for cancer prevention and therapy: From phytochemistry to novel formulations. Pharmacol. Res. 2021, 175, 105837. [Google Scholar] [CrossRef]
- Catanzaro, E.; Canistro, D.; Pellicioni, V.; Vivarelli, F.; Fimognari, C. Anticancer potential of allicin: A review. Pharmacol. Res. 2022, 177, 106118. [Google Scholar] [CrossRef]
- De Greef, D.; Barton, E.M.; Sandberg, E.N.; Croley, C.R.; Pumarol, J.; Wong, T.L.; Das, N.; Bishayee, A. Anticancer potential of garlic and its bioactive constituents: A systematic and comprehensive review. Semin. Cancer Biol. 2020, 73, 219–264. [Google Scholar] [CrossRef]
- Daniel, M.; Tollefsbol, T.O. Epigenetic linkage of aging, cancer and nutrition. J. Exp. Biol. 2015, 218, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Elkashty, O.A.; Tran, S.D. Sulforaphane as a Promising Natural Molecule for Cancer Prevention and Treatment. Curr. Med Sci. 2021, 41, 250–269. [Google Scholar] [CrossRef]
- Kaiser, A.E.; Baniasadi, M.; Giansiracusa, D.; Giansiracusa, M.; Garcia, M.; Fryda, Z.; Wong, T.L.; Bishayee, A. Sulforaphane: A Broccoli Bioactive Phytocompound with Cancer Preventive Potential. Cancers 2021, 13, 4796. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.M.; Gago-Dominguez, M.; Castelao, J.E.; Hankin, J.H.; Ross, R.K.; Yu, M.C. Cruciferous vegetables in relation to renal cell carcinoma. Int. J. Cancer 1998, 77, 211–216. [Google Scholar] [CrossRef]
- Lee, J.E.; Giovannucci, E.; Smith-Warner, S.A.; Spiegelman, D.; Willett, W.C.; Curhan, G.C. Intakes of Fruits, Vegetables, Vitamins A, C, and E, and Carotenoids and Risk of Renal Cell Cancer. Cancer Epidemiol. Biomark. Prev. 2006, 15, 2445–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.E.; Männistö, S.; Spiegelman, D.; Hunter, D.J.; Bernstein, L.; Brandt, P.A.V.D.; Buring, J.E.; Cho, E.; English, D.R.; Flood, A.; et al. Intakes of Fruit, Vegetables, and Carotenoids and Renal Cell Cancer Risk: A Pooled Analysis of 13 Prospective Studies. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1730–1739. [Google Scholar] [CrossRef] [Green Version]
- Bosetti, C.; Scotti, L.; Maso, L.D.; Talamini, R.; Montella, M.; Negri, E.; Ramazzotti, V.; Franceschi, S.; La Vecchia, C. Micronutrients and the risk of renal cell cancer: A case-control study from Italy. Int. J. Cancer 2006, 120, 892–896. [Google Scholar] [CrossRef]
- Bosetti, C.; Rossi, M.; McLaughlin, J.K.; Negri, E.; Talamini, R.; Lagiou, P.; Montella, M.; Ramazzotti, V.; Franceschi, S.; LaVecchia, C. Flavonoids and the Risk of Renal Cell Carcinoma. Cancer Epidemiol. Biomark. Prev. 2007, 16, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; The Canadian Cancer Registries Epidemiology Research Group; La Vecchia, C.; Negri, E.; Desmeules, M.; Mery, L. Dietary vitamin C, E, and carotenoid intake and risk of renal cell carcinoma. Cancer Causes Control 2009, 20, 1451–1458. [Google Scholar] [CrossRef]
- Brock, K.E.; Ke, L.; Gridley, G.; Chiu, B.C.-H.; Ershow, A.G.; Lynch, C.F.; Graubard, B.I.; Cantor, K.P. Fruit, vegetables, fibre and micronutrients and risk of US renal cell carcinoma. Br. J. Nutr. 2011, 108, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.H.; Ruterbusch, J.J.; Holowatyj, A.N.; Steck, S.E.; Van Dyke, A.L.; Ho, W.J.; Cote, M.L.; Hofmann, J.N.; Davis, F.; Graubard, B.I.; et al. Renal cell carcinoma risk associated with lower intake of micronutrients. Cancer Med. 2018, 7, 4087–4097. [Google Scholar] [CrossRef]
- Ho, W.J.; Simon, M.S.; Yildiz, V.O.; Shikany, J.M.; Kato, I.; Beebe-Dimmer, J.L.; Cetnar, J.P.; Bock, C.H. Antioxidant micronutrients and the risk of renal cell carcinoma in the Women’s Health Initiative cohort. Cancer 2014, 121, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Bertoia, M.; Albanes, D.; Mayne, S.T.; Männistö, S.; Virtamo, J.; Wright, M.E. No association between fruit, vegetables, antioxidant nutrients and risk of renal cell carcinoma. Int. J. Cancer 2009, 126, 1504–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, M.E.; Mendelsohn, L.; Butler, K.; Riley, L.; Cantrell, J.; Wiseman, C.; Taylor, R.; Macdonald, J.S. Treatment of metastatic renal cell carcinoma with coumarin (1,2-benzopyrone) and cimetidine: A pilot study. J. Clin. Oncol. 1987, 5, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, R.; Manegold, C.; Maurer, B.; Hennig, F.; Matthiessen, W. Short report: Phase II trial of coumarin and cimetidine in advanced renal cell carcinoma. Ann. Oncol. 1990, 1, 445–446. [Google Scholar] [CrossRef] [PubMed]
- Dexeus, F.H.; Logothetis, C.J.; Sella, A.; Fitz, K.; Amato, R.; Reuben, J.M.; Dozier, N. Phase II study of coumarin and cimetidine in patients with metastatic renal cell carcinoma. J. Clin. Oncol. 1990, 8, 325–329. [Google Scholar] [CrossRef]
- Kokron, O.; Maca, S.; Gasser, G.; Schmidt, P.R. Cimetidine and Coumarin Therapy of Renal Cell Carcinoma. Oncology 1991, 48, 102–106. [Google Scholar] [CrossRef]
- Sagaster, P.; Micksche, M.; Flamm, J.; Ludwig, H. Randomised study using IFN-α versus IFN-α plus coumarin and cimetidine for treatment of advanced renal cell cancer. Ann. Oncol. 1995, 6, 999–1003. [Google Scholar] [CrossRef]
- Marquardt, A.; Solimando, A.G.; Kerscher, A.; Bittrich, M.; Kalogirou, C.; Kübler, H.; Rosenwald, A.; Bargou, R.; Kollmannsberger, P.; Schilling, B.; et al. Subgroup-Independent Mapping of Renal Cell Carcinoma—Machine Learning Reveals Prognostic Mitochondrial Gene Signature Beyond Histopathologic Boundaries. Front. Oncol. 2021, 11, 621278. [Google Scholar] [CrossRef]
- Argentiero, A.; Solimando, A.G.; Krebs, M.; Leone, P.; Susca, N.; Brunetti, O.; Racanelli, V.; Vacca, A.; Silvestris, N. Anti-angiogenesis and Immunotherapy: Novel Paradigms to Envision Tailored Approaches in Renal Cell-Carcinoma. J. Clin. Med. 2020, 9, 1594. [Google Scholar] [CrossRef]
- Lagoa, R.; Silva, J.; Rodrigues, J.R.; Bishayee, A. Advances in phytochemical delivery systems for improved anticancer activity. Biotechnol. Adv. 2019, 38, 107382. [Google Scholar] [CrossRef]
- Ahmad, R.; Srivastava, S.; Ghosh, S.; Khare, S.K. Phytochemical delivery through nanocarriers: A review. Colloids Surf. B Biointerfaces 2020, 197, 111389. [Google Scholar] [CrossRef]
- Aiello, P.; Consalvi, S.; Poce, G.; Raguzzini, A.; Toti, E.; Palmery, M.; Biava, M.; Bernardi, M.; Kamal, M.A.; Perry, G.; et al. Dietary flavonoids: Nano delivery and nanoparticles for cancer therapy. Semin. Cancer Biol. 2019, 69, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Tuli, H.S.; Yerer, M.B.; Sharma, A.; Sak, K.; Srivastava, S.; Pandey, A.; Garg, V.K.; Sethi, G.; Bishayee, A. Natural product-based nanoformulations for cancer therapy: Opportunities and challenges. Semin. Cancer Biol. 2021, 69, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari-Movahed, M.; Mondal, A.; Farzaei, M.H.; Bishayee, A. Quercetin- and rutin-based nano-formulations for cancer treatment: A systematic review of improved efficacy and molecular mechanisms. Phytomedicine 2021, 97, 153909. [Google Scholar] [CrossRef] [PubMed]







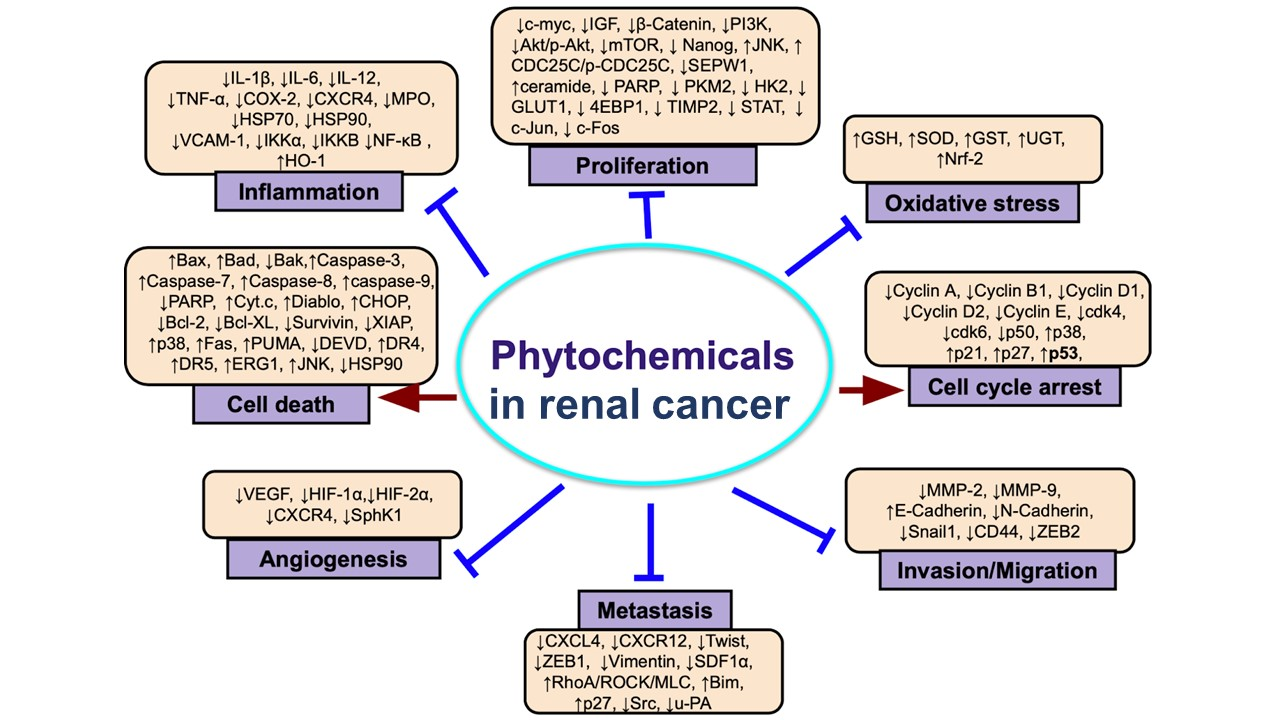{kind=link}
{kind=link}
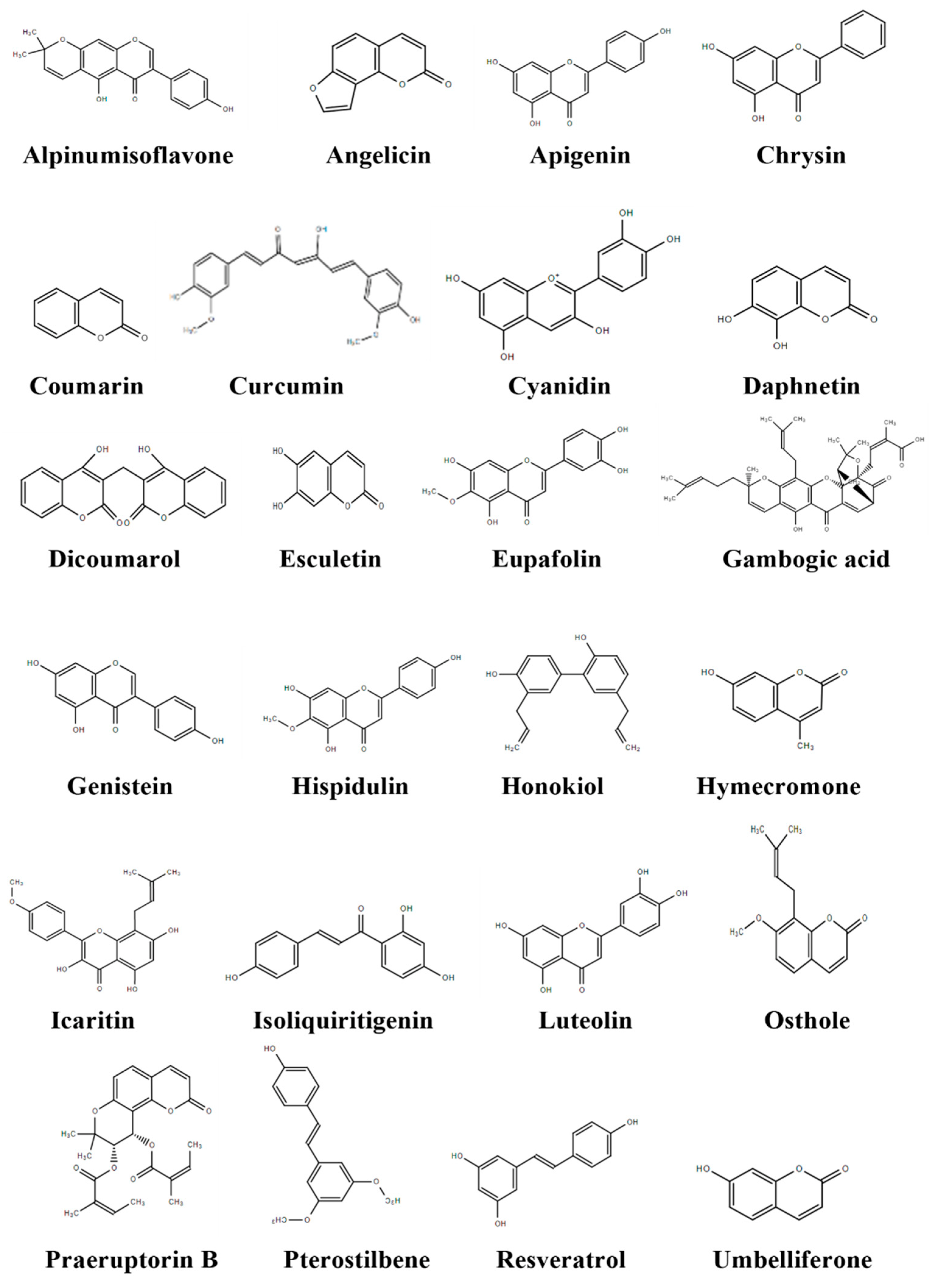{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Cell Lines Used | Concentrations (Duration) | Anticancer Effects | Mechanisms | References |
|---|---|---|---|---|---|
| Phenolics | |||||
| Alpinumisoflavone | 786-O (renal cell carcinoma); Caki-1 (clear cell renal cell carcinoma); RCC4 (clear cell renal cell carcinoma) | 2.5–10 μM (48 h) | Suppressed cell growth and invasion | ↑Apoptosis; ↑miR-101; ↓RLIP76; ↓p-Akt/Akt | Wang et al., 2017 [39] |
| Angelicin | Caki (renal cell carcinoma) | 50–100 μM (24 h) | No effects alone | Not reported | Min et al., 2018 [40] |
| Apigenin | ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma); Caki-1 (clear cell renal cell carcinoma) | 15.4–50.9 μM (24, 48 h) | Inhibited cell proliferation | ↑Apoptosis; ↑p53; ↑Bax; ↑caspase-3; ↑caspase-9; ↑DNA damage; ↑γH2AX; ↑G2/M phase arrest | Meng et al., 2017 [41] |
| Coumarin | 786-O (renal cell carcinoma); A-498 (renal cell carcinoma) | 10–500 μg/mL (5 days) | Inhibited cell proliferation | Not reported | Myers et al., 1994 [42] |
| Curcumin | Caki (renal cell carcinoma) | 25–100 μM (24 h) | Showed cytotoxic activity | ↑Apoptosis; ↓pro-caspase 3; ↑DEVD-pNA cleavage; ↓cIAP1; ↓XIAP; ↓Bcl-2; ↓IAP; ↑cyt c; ↓p-Akt | Woo et al., 2003 [43] |
| Curcumin | Caki (renal cell carcinoma) | 10–30 μM (24 h) | Enhanced therapeutic effects of TRAIL | ↑Apoptosis; ↑sub-G1; ↑DEVDase; ↑DR4; ↑DR5; ↑ROS | Jung et al., 2005 [44] |
| Curcumin | Caki (renal cell carcinoma); ACHN (papillary renal cell carcinoma); A-498 (renal cell carcinoma) | 10–30 μM (48 h) | No effects alone | Not reported | Seo et al., 2014 [45] |
| Curcumin | RCC-949 (renal cell carcinoma) | 10–100 μM (24–96 h) | Inhibited proliferation | ↑Apoptosis; ↓Bcl-2; ↑Bax; ↑G2/M cell cycle arrest; ↓cyclin B1; ↓PI3K/Akt | Zhang et al., 2015 [46] |
| Curcumin | Caki-1 (clear cell renal cell carcinoma); OS-RC-2 (renal cell carcinoma) | 10–15 μM (48 h) | Decreased proliferation | ↑Apoptosis; ↑YAP | Xu et al., 2016 [47] |
| Cyanidin | 786-O (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 0.5–100 μM (24–72 h) | Inhibited growth and migration | ↑Apoptosis, ↑G1/M phase arrest; ↑ERG1; ↓SEPW1; ↓HIF2A; ↓E-cadherin; ↓Bcl-2; ↓caspase-3; ↓ROS; ↓autophagy | Liu et al., 2018 [48] |
| Daphnetin | A-498 (renal cell carcinoma) | 5–500 μM (48–96 h) | Inhibited proliferation | ↓G1/S; ↑S phase; ↓p-ERK1/2; ↑p38 | Finn et al., 2004 [49] |
| Dicoumarol | ACHN (papillary renal cell carcinoma); A-498 (renal cell carcinoma); Caki (renal cell carcinoma) | 25–50 μM (24 h) | Decreased cell viability | ↑Apoptosis; ↓Bcl-2; ↓Mcl-1; ↓c-FLIP; ↓NF-κB; ↓CRE; ↑REDD; ↑ATF4; ↑CHOP; ↑GRP78 | Park et al., 2014 [50] |
| Esculetin | 786-O (renal cell carcinoma); SN12-PM6 (renal cell carcinoma) | 12.5–800 µg/mL (24, 48 h) | Reduced cell viability and inhibited invasion and migration | ↑Apoptosis; ↑G0/G1 arrest; ↑G2/M arrest; ↓S phase; ↓cyclin D1; ↓CDK4; ↓CDK6; ↓c-Myc; ↑E-cadherin; ↓N-cadherin; ↓vimentin | Duan et al., 2020 [51] |
| Eupafolin | Caki (renal cell carcinoma) | 20–30 μM (24 h) | No effect alone | None | Han et al., 2016 [52] |
| Gambogic acid | Caki-1 (clear cell renal cell carcinoma); 786-O (renal cell carcinoma) | 0.5–4 μM (48 h) | Enhanced therapeutic effects of sunitinib | ↑Sub-G1 population; ↑P21; ↓Bcl-2; ↓VEGF | Jiang et al., 2012 [53] |
| Genistein | SMKT-R1 (renal cell carcinoma); SMKT-R2 (clear cell renal cell carcinoma); SMKT-R3 (clear cell renal cell carcinoma); SMKT-R4 (renal cell carcinoma) | 12.5–100 μg/mL (48 h) | Inhibited proliferation | ↑Apoptosis | Sasamura et al., 2004 [54] |
| Genistein | A-498 (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 10–50 μmol/L (24–120 h) | Induced antiproliferative effects and reduced cell viability | ↑G2/M phase arrest; ↑BTG3; ↓DNMTase; ↓DNMT1; ↓DNMT 3b; ↓HDAC | Majid et al., 2009 [55] |
| Genistein | A-498 (renal cell carcinoma); 786-O (renal cell carcinoma); Caki-2 (papillary renal cell carcinoma) | 25 μM (24–72 h) | Inhibited proliferation and invasion | ↑Apoptosis; ↓miR-1260b; ↓TCF reporter activity; ↓sFRP1; ↓Dkk2; ↓Smad4 | Hirata et al., 2013 [56] |
| Hispidulin | Caki-1 (clear cell renal cell carcinoma); 786-O (renal cell carcinoma) | 12.5–50 μM (48 h) | Inhibited proliferation | ↑G0-G1 phase arrest; ↓G2 phase; ↑caspase-3; ↓p-STAT3; ↓Bcl-2; ↓survivin | Gao et al., 2015 [57] |
| Hispidulin | ACHN (papillary renal cell carcinoma); Caki-2 (clear cell renal cell carcinoma) | 10–20 μmol/L (24–72 h) | Decreased cell viability | ↑Apoptosis; ↑Fas/Fas ligand; ↑DR5; ↑ceramide; ↓SphK1; ↑ROS/JNK | Gao et al., 2017 [58] |
| Honokiol | A-498 (renal cell carcinoma) | 2.5–80 μmol (72 h) | Suppressed proliferation and migration, inhibited invasion | ↑E-cadherin; ↓fibronectin; ↓vimentin; ↓SP cells; ↑miR-141; ↓ZEB2; | Li et al., 2014 [59] |
| Honokiol | 786-O (renal cell carcinoma); A-498 (renal cell carcinoma) | 5–80 μM (24 h) | Inhibited migration, suppressed proliferation | ↑RhoA/ROCK/MLC | Cheng et al., 2016 [60] |
| Hymecromone | Caki-1 (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma); A-498 (renal cell carcinoma) | 16 μg/mL (18, 48 h) | Enhanced therapeutic effects of sorafenib | ↑Apoptosis; ↓CD44; ↓RHAMM; ↑caspase-3; ↑caspase-8; ↑caspase-9; ↑cleaved PARP; ↓Mcl-1; ↓p-EGFR; ↓p-Met | Benitez et al., 2013 [61] |
| Icaritin | 786-O (renal cell carcinoma) | 1–10 μM (24, 48 h) | Inhibited proliferation | ↓Cyclin E; ↓cyclin D1; ↓survivin; ↑caspase-3; ↑cleaved PAR; ↓Bcl-xL; ↓Mcl-1; ↓activated STAT3; ↓JAK2; ↓IL-6-induced JAK2/STAT3; ↓IL-6 induced p-Akt and p-MAPK | Li et al., 2013 [62] |
| Isoliquiritigenin | Caki (renal cell carcinoma) | 5–50 μM (24–72 h) | Decreased cell viability | ↑Apoptosis; ↑caspase-3; ↑caspase-7; ↑caspase-9; ↑cleaved PARP; ↑Bax; ↓Bcl-2; ↓Bcl-xL; ↑cyt. c; ↑p53; ↓Mdm2; ↑ROS; ↓STAT3; ↓cyclin D1; ↓cyclin D2; ↑JAK2 | Kim et al., 2017 [63] |
| Luteolin | ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma); A-498 (renal cell carcinoma) | 10–40 μM (24, 48 h) | Reduced cell viability | ↑Apoptosis; ↓p-Akt; ↑p-P38; ↑Ask1; ↑p-JNK; ↑p-ERK; ↑caspase-3; ↑PARP-1; ↓HSP90 | Ou et al., 2013 [64] |
| Luteolin | ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma); A-498 (renal cell carcinoma) | 10–40 μM (48 h) | Reduced cell viability | ↑Apoptosis; ↑caspase-3; ↑caspase-8; ↑caspase-9; ↓Mcl-1; ↓FLIP; ↓p-Akt; ↓p-STAT3, | Ou et al., 2014 [65] |
| Osthole | 786-O (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 20–240 μM (24, 48 h) | Suppressed proliferation, inhibited migration and invasion | ↑Apoptosis; ↑caspase-3; ↑Bax; ↓Bcl-2; ↓survivin; ↓MMP2; ↓MMP9; ↑E-cadherin; ↑Beta-catenin; ↓N-cadherin; ↓Smad-3; ↓Snail-1; ↓Twist-1 | Liu et al., 2017 [66] |
| Osthole | Caki (renal cell carcinoma) | 20–30 μM (24 h) | Decreased cell viability | ↑Apoptosis; ↑sub-G1 population; ↑nuclear condensation; ↑DNA fragmentation; ↑caspase-3; ↓c-FLIP; ↓MMP; ↑cyt. c | Min et al., 2017 [67] |
| Praeruptorin B | 786-O (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 10–50 μM (24 h) | Inhibited cell migration and invasion | ↓CTSC; ↓CTSV; ↓p-ERK; ↓p-EGFR; ↓p-MEK | Lin et al., 2020 [68] |
| Pterostilbene | A-498 (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 5–100 μM (24–72 h) | Displayed antiproliferative effects | ↑Apoptosis; ↑Bad; ↑Bax; ↑cyt c; ↑cleaved-caspase 3; ↑cleaved-caspase 9; ↑cleaved-PARP; ↓Bcl-2; ↑S phase cells; ↓p-Akt; ↓p-ERK1/2; ↑γH2AX; ↓Rad1; ↓PCNA | Zhao et al., 2020 [69] |
| Resveratrol | RCC5430 (renal cell carcinoma) | 25–50 μM (12–72 h) | Inhibited cell growth and induced cell death | ↑VDR; ↑TRAF1; ↑BGPa; ↑GADD45; ↓GFRA2; ↓HCK; ↓CYP1B1; ↓MUC1; ↓CXCR4 | Shi et al., 2004 [70] |
| Resveratrol | 786-O (renal cell carcinoma) | 10–40 μM (24–72 h) | Inhibited proliferation | ↓VEGF | Yang et al., 2011 [71] |
| Resveratrol | ACHN (papillary renal cell carcinoma) | 7.8125–62.5 µg/mL (12–48 h) | Reduced cell viability, suppressed migration and invasion | ↓MMP2; ↓MMP9; ↑acH3K9; ↑acH3K14; ↑acH4K12; ↑acH4K5; ↑acH4K16 | Dai et al., 2020 [72] |
| Resveratrol | Ketr-3 (renal cell carcinoma) | 12.5–100 μM (12–72 h) | Suppressed migration and viability | ↑Bax; ↓Bcl-2; ↑p53; ↑p-AMPK; ↓p-mTOR; ↑LC3; ↑ATG5; ↑ATG7 | Liu et al., 2018 [73] |
| Resveratrol | ACHN (papillary renal cell carcinoma); A-498 (renal cell carcinoma) | 10–200 μM (24–72 h) | Inhibited proliferation, migration and invasion | ↑E-cadherin; ↑TIMP-1; ↓N-cadherin; ↓vimentin; ↓snail; ↓MMP2; ↓MMP9; ↓p-Akt; ↓p-ERK1/2 | Zhao et al., 2018 [74] |
| Resveratrol | ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma) | 25–100 μM (12, 24 h) | Inhibited cell growth and viability | ↑Apoptosis; ↑Bax/Bcl-2 ratio; ↓NLRP3; ↓caspase-1; ↑caspase-3; ↓IL-6; ↑E-cadherin | Tian et al., 2020 [75] |
| Resveratrol | Caki-1 (clear cell renal cell carcinoma) | 10–50 μM (24 h) | Enhanced chemosensitivity to paclitaxel and inhibited cell growth | ↑Apoptosis; ↓survivin; ↓p-PI3K; ↓p-Akt | Ying-jie et al., 2019 [76] |
| Resveratrol | Caki-1 (clear cell renal cell carcinoma); 786-O (renal cell carcinoma) | 10–50 μM (6 h) | Inhibited proliferation | ↑Apoptosis; ↓STAT3; ↓STAT5; ↓JAK1; ↓JAK2; ↓Src kinase; ↑PTPε; ↑SHP-2; ↑S phase; ↓G1/G0; ↑p21; ↓cyclin D1; ↓cyclin E; ↑Bax; ↑p53 | Kim et al., 2016 [77] |
| Umbelliferone | ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma); OS-RC-2 (renal cell carcinoma) | 5–150 µmol/L (48, 72 h) | Decreased cell proliferation | ↑Apoptosis; ↑G1 cell cycle arrest; ↑Bax; ↓Ki-67; ↓MCM2; ↓Bcl-2; ↓CDK2; ↓Cyclin E1; ↓CDK4; ↓CyclinD1; ↓p110γ | Wang et al., 2019 [78] |
| Terpenoids | |||||
| Anatolicin | UO31 (RCC); A498 (RCC) | 0.017, 0.024 μM (IC50) (48 h) | Decreased cell viability | Not reported | Tosun et al., 2019 [79] |
| Artemisinin | UMRC-2 (clear cell renal cell carcinoma); CAKI-2 (clear cell renal cell carcinoma) | 25 μM (24 h) | Inhibited cell growth, inhibited cell invasion and migration | ↓c-Myc; ↓ cyclin D1; ↓PCNA, ↓N-cadherin; ↓Vimentin; ↓Snail; ↑E-cadherin | Yu et al. 2019 [80] |
| Artesunate | Caki (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma); A-498 (renal cell carcinoma) | 20–50 μM (12–24 h) | Reduced cell viability | ↑ROS; ↑RIP1-dependent cell death | Chauhan et al., 2017 [81] |
| Auraptene | RCC4 (clear cell renal cell carcinoma) | 25–100 μM (24 h) | Reduced motility and inhibited metabolism | ↓Mitochondrial complex 1; ↓GLUT1; ↓HK2; ↓PFK; ↓LDHA | Jang et al., 2015 [82] |
| β-elemene | 786-O (renal cell carcinoma) | 50, 100, 150, 200 μg/mL (24–72 h) | Inhibited cell viability | ↑PARP cleavage; ↓Bcl-2, ↓Survivin, ↑p-ERK; ↑p-JNK, ↓PI3K/Akt/mTOR | Zhan et al. 2012 [83] |
| Betulin | 786-O (renal cell carcinoma); Caki-2 (clear cell renal cell carcinoma) | 0.5–5 μM (48 h) | Inhibited cell proliferation and reduced cell viability | ↑mTOR activation; ↓aerobic glycolysis; ↓p-S6; ↓p-4EBP1; ↓PKM2; ↓HK2 | Cheng et al., 2017 [84] |
| Betulinic acid | 786-O (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 5, 10, 20 μg/mL (24–72 h) | Repressed migration and invasion of cancer cells, exerted cytotoxic and cytostatic effects | ↑ROS; ↑ MMP2; ↑MMP9; ↑vimentin; ↓TIMP2; ↓E-cadherin; ↑Bax; ↑cleaved caspase-3; ↓Bcl2 | Yang et al., 2018 [85] |
| Cafestol | Caki (clear cell renal cell carcinoma) | 10–40 μM (24 h) | Inhibited cell growth and decreased viability | ↑Apoptosis; ↑caspase-2; ↑caspase-3; ↑Bim; ↑Bax; ↓cFLIP; ↓Bcl-2; ↓Mcl-1; ↓Bcl-xL; ↑cyt. c; ↓MMP; ↑sub-G1 phase cells; ↓p-Akt; ↓p-STAT3 | Choi et al., 2011 [86] |
| Cafestol; Cafesol+ABT-737 | Caki (clear cell renal cell carcinoma) | 30 μM (24 h) | Induced cell death | ↓Mcl-1; ↑Bim; ↑proteasome activity; ↑sensitivity to ABT-737-mediated apoptosis | Woo et al., 2014 [87] |
| Carsonic acid | Caki (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma); A-498 (renal cell carcinoma) | 10–20 μM (24 h) | Inhibited cell viability | ↑Apoptosis; ↑sub-G1 population; ↑cleaved-PARP; ↑caspase-3; ↑caspase-8; ↑caspase-9; ↓MMP; ↑cyt c; ↑Bax; ↓c-FLIP; ↑Bcl-2; ↑DR5; ↑Bim; ↑PUMA; ↑ATF4; ↑CHOP; ↑Ca2+ | Jung et al., 2015 [88] |
| Carsonic acid | Caki (clear cell renal cell carcinoma) | 5–100 μM (24–72 h) | Decreased cell viability | ↑Caspase-9; ↑ caspase-7; ↑caspase-3; ↑PARP; ↓Bcl-2; ↓Bcl-xL; ↑Bax; ↑p53; ↓Mdm2; ↑ p27; ↓STAT DNA-binding activity; ↓phosphorylation Y705 and S727; ↓c-Myc; ↓survivin; ↓D-series of cyclins; ↓Src; ↑ROS | Park et al., 2016 [89] |
| Corosolic acid | Caki (clear cell renal cell carcinoma) | 2.5, 5, 10 μM (24 h) | Induces cell death | ↓a-tocopherol; ↑lipid peroxidation-dependent non-apoptotic cell death; ↓a-tocopherol; ↓ferrostatin-1; ↓DFO | Woo et al., 2018 [90] |
| Crocin | A498 (renal cell carcinoma); A704 (renal cell carcinoma) | 2.5–10 μM (24–72 h) | Suppressed cell proliferation and migration | ↑miR-577 inhibitors; ↓NFIB-wt | Niu et al., 2020 [91] |
| Cucurbitacins | Caki-1 (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 1–10 μM (1, 4 h) | Sensitized cells to programmed cell death | ↑Apoptosis; ↑caspase-8; ↑TRAIL sensitizing effect | Henrich et al., 2012 [92] |
| Englerin A | 786-O (hypertriploid RCC); A498 (RCC); ACHN (papillary renal cell carcinoma); CAKI-1 (ccRCC); RXF-393 (RCC); SN12C (RCC); TK-10 (ccRCC); UO-31 (RCC) | 1–87 nM | Decreased cell growth | Not reported | Ratnayake et al., 2009 [93] |
| Englerin A | A-498 (renal cell carcinoma); UO-31 (renal cell carcinoma) | 1 μM (48 h) | Reduced cell viability | ↑ROS; ↑Ca2+ | Sulzmaier et al., 2012 [94] |
| Englerin A | A498 (renal cell carcinoma) | 50, 100 nm (24, 48 h) | Increased cytotoxicity, blocks cell cycle | ↓Akt; ↓ERK; ↓G2/M transition; ↓NEAA; ↑cathepsins; ↑calpains; ↓PI3/Akt; ↑PKCθ; ↑HFS1 | Williams et al., 2013 [95] |
| Englerin A | A-498 (renal cell carcinoma) | 0.1–5000 nM 24 h | Inhibited cell proliferation | ↑intracellular Ca2+; ↑activation TRPC4/C5; ↓TRPA1; ↓TRPV3; ↓TRPV4; ↓TRPM8; ↑PXR | Carson et al., 2015 [96] |
| Englerin A | cc-RCC (clear cell renal cell carcinoma); A498 (renal cell carcinoma); UO-31 (renal cell carcinoma) | 100 nm (24–48 h) | Altered lipid metabolism, inducing ER stress and acute inflammatory response | ↑PLIN2; ↑autophagy; ↑PKCθ; ↑ IRF systolic pattern recognition receptors; ↑RIG-I/MDA5; ↑INF-α/ß | Batova et al., 2017 [97] |
| Escin | 786-O (renal cell carcinoma); Caki-1 (clear cell renal cell carcinoma) | 6, 12.5, 25, 50, 100 μM (24–72 h) | Exhibited cytotoxic effects, induced cell cycle arrest, ROS production | ↓cdc-2; ↑caspase-8; ↑ caspase-9; ↑caspase-3; ↑PARP; ↓ΔΨm; ↓Bcl-2; ↑Bax; ↓IAP proteins like ZIAP and survivin; ↑ ROS; ↓Fas death receptor; Fas-L; FADD | Yuan et al., 2017 [98] |
| Helenalin | Caki (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 2–6 μM (24 h) | Decreased cell viability | ↑Apoptosis; ↑sub-G1 population; ↑caspase-3; ↑ROS, ↑PARP; ↑ATF4; ↑REDD1; ↑CHOP | Jang et al., 2013 [99] |
| 16-hydroxyclerod-3,113-dien-15,16-olide | ccRCC (clear cell renal cell carcinoma); A-498 (renal cell carcinoma); 786-O (renal cell carcinoma) | 10–40 μM (24–72 h) | Inhibited cell proliferation and decreased cell viability | ↑ROS; ↓MMP; ↓mitochondrial cyt. c; ↑cytosolic cyt. c, ↓ Bcl-2; ↓hsp70; ↑caspase-3; ↑cleaved PARP; ↓p-Akt; ↓mTOR; ↓MEK1/2; ↓ERK1/2; ↓c-Myc; ↓HIF-2a; ↑FOXO2a | Liu et al., 2017 [100] |
| Kahweol | Caki (clear cell renal cell carcinoma) | 5–20 μM (24 h) | Decreased cell viability | ↑Apoptosis; ↓c-FLIP; ↓Bcl-2; ↑PARP cleavage; ↑caspase-8; ↓NF-kB p65 | Um et al., 2010 [101] |
| Kahweol | Caki (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma); A498 (renal cell carcinoma) | 20 μM (24 h) | Decreased cell viability | ↑sub-G1 population; ↑PARP cleavage; ↓c-FLIP; ↓Mcl-1 | Min et al., 2017 [102] |
| Nimbolide | 786-O (renal cell carcinoma); A-498 (renal cell carcinoma) | 1–6 μM (24 h) | Decreased cell viability | ↑Apoptosis; ↑G2/M cell cycle arrest; ↑p-p53; ↑p-cdc2; ↑p-cdc25c; ↓cyclin A; ↓cyclin B; ↓cdc2; ↓cdc25c; ↑DNA damage; ↑γ-H2AX; ↑cleaved-caspase-3; ↑cleaved-PARP; ↓pro-caspase-8; ↓Mcl-1; ↓Bcl-2; ↑Bax; ↑DR5; ↑CHOP | Hsiesh et al., 2015 [103] |
| Oridonin | 786-O (renal cell carcinoma) | 10, 20, 40 μM (24 h) | Increased cell cytotoxicity | ↑Apoptosis; ↑necroptosis; ↑cleaved caspase-3; ↑Bax; ↓Bcl-2; ↑RIP-1; ↑RIP-3; ↑LDH; ↑HGMB1; ↑Parp-1; ↓GSH; ↑p-JNK; ↑p-ERK; ↑p-P38 | Zheng et al., 2018 [104] |
| Parthenolide | OUR-10 (renal cell carcinoma) ACHN (papillary renal cell carcinoma) | 0.1–5 μM (24 h) | Decreased cell growth and proliferation | ↑NF-κB p65; ↑IkBα; ↑p-NF-kB | Oka et al., 2007 [105] |
| Silibinin | 786-O (renal cell carcinoma) | 10–50 μM (24 h) | Inhibits migration and invasion of cancer cells | ↓MMP-9; ↓MMP-2; ↓u-PA; ↑TIMP-2; ↑PAI-1; ↓ERK 1/2 phosphorylation; ↓p38; ↓NF-kB; ↓c-Jun; ↓c-Fos | Chang et al., 2011 [106] |
| Sorghumol | A-498 (renal cell carcinoma) | 10–40 μM (24 h) | Reduced cell proliferation | ↑Apoptosis; ↑arrest in G2/M; ↓p-mTOR; ↓p-PI3K; ↓p-Akt | Li et al., 2019 [107] |
| Thymoquinone | Caki (clear cell renal cell carcinoma); A498 (renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 25–75 μM (24 h) | Decreased cell viability | ↑PARP; ↓c-FLIP; ↓Bcl-2; ↓NF-κB; ↑ROS; ↑cyt c; ↓MMP | Park et al., 2016 [108] |
| Thymoquinone | 786-O-S13 (RCC) | 5–20 µM (24 h) | Decreased cell movement, repressed migration and invasion of cancer cells | ↓MMP-2; ↓u-PA; ↓p-phosphatidylinositol 3-kinase; ↓p-Akt; ↓p-Src; ↓p-Paxillin; ↓fibronectin; ↓N-cadherin; ↓Rho A; ↓TGF-β-promoted u-PA activity; ↑adhesion type I collagen; ↑adhesion type IV collagen | Liou et al., 2019 [109] |
| Tonantzitlolone | ccRCC (clear cell renal cell carcinoma); 786-O (renal cell carcinoma); A498 (renal cell carcinoma) | 1–5 µM (24 h) | Increased cell cytotoxicity | ↑PKCθ; ↑PKCα; ↑p-IRS1; ↓p-Akt; ↓glucose uptake; ↑PKCθ-dependent HSF1 phosphorylation | Sourbier et al., 2015 [110] |
| Tonantzitlolone | A498 (renal cell carcinoma); HEK293 (human embryonic kidney cell) | 10–1000 nM (24 h) | Increased cell cytotoxicity | ↑intracellular Ca2+; ↑TRPC5 channels | Rubaiy et al., 2018 [111] |
| Triptolide | 786-O (renal cell carcinoma); OS-RC-2 (renal cell carcinoma) | 12.5–200 nM (24, 48 h) | Decreased cell viability | ↑Apoptosis; ↑caspase-3; ↑cyt. c; ↑Bax; ↓Bcl-2; ↓Bcl-xL; ↑cell cycle arrest at S phase; ↓Rb; ↓A/CDK1; ↓CDK2; ↓B/CDK; Induced cell cycle arrest | Li et al., 2011 [68] |
| Triptolide | A498 (renal cell carcinoma); Caki-1 (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma); 786-O (renal cell carcinoma); 769-P (renal cell adenocarcinoma) | 10–100 nM (24 h) | Enhanced therapeutic effects of TRAIL | ↑TRAIL-R2; ↓HSP70; ↑Apoptosis | Brincks et al., 2015 [112] |
| Triptolidenol | 786-O (renal cell carcinoma); Caki-1 (clear cell renal cell carcinoma); ACHN (papillary renal cell carcinoma) | 30–500 nM (48 h) | Reduced cell proliferation | ↑Apoptosis; ↑Bax; ↓Bcl-2; ↑cyt. c; ↑cleaved-caspase-3; ↑cleaved-caspase-9; ↑cleaved-PARP; ↑S phase arrest; ↓cyclin A2; ↓cyclin D1; ↓cyclin E1; ↓CDK2; ↓CDK4; ↓COX-2; ↓NF-κB; ↓MMP | Jin et al., 2021 [113] |
| Zerumbone | 786-O (renal cell carcinoma); 769-P (renal cell adenocarcinoma) | 10–50 µM (24–72 h) | Decreased cell viability | ↑caspase-3; ↑caspase-9; ↓Gli-1; ↓Bcl-2 | Sun et al., 2013 [114] |
| Zerumbone | 786-O (renal cell carcinoma); Caki-1 (clear cell renal cell carcinoma) | 5–50 µM (24 h) | Inhibited colony formation | ↓Ki-67; ↓Bcl-2; ↓Bcl-xL; ↑caspase-3; ↑PARP; ↓cyclin D1; ↓SHP-1; ↓Mcl-1; ↓Survivin; ↓MMP-9; ↓VEGF; ↓CD31; ↓constitutive p-STAT3 activation; ↓STAT3-DNA binding activity; ↓phosphorylation of c-Src; ↓p-JAK1; ↓p-JAK2 | Shanmugam et al., 2015 [115] |
| Alkaloids | |||||
| (-)-Antofine | Caki-1 (ccRCC) | 25 nM (24 h) | Decreased cell growth | ↓Met; ↓STAT3; ↓Grb2; ↓RAC1; ↓Src; ↓ERK1 | Song et al., 2015 [116] |
| Berberine | Caki (clear cell RCC) | 60 µM (24 h) | Increased cell death | ↓c-FLIP; ↓Mcl-1 | Lee et al., 2011 [117] |
| Berberine | ACHN (papillary renal cell carcinoma); 786-O (hypertriploid RCC) | 5–320 µM (24, 48 h) | Decreased cell viability and increased cell death | ↑ROS; ↑caspase 3; ↑PLK3; ↓FIGF; ↓TERT; ↓formate; ↓lactate; ↑lysine | Lopes et al., 2020 [118] |
| Cepharanthine | Caki (clear cell RCC); ACHN (papillary renal cell carcinoma); A498 (RCC) | 10–15 µM (18 h) | Decreased cell viability | ↓Survivin; ↓cellular FLICE inhibitory protein (cFLIP); ↓STAMBPL1; ↑USP53; ↑DR5 | Shahriyar et al., 2018 [119] |
| Chelerythrine | Caki (clear cell RCC); 786-O (hypertriploid RCC) | 0.3–40 µM/L (24 h) | Anti-tumor activity of CHE against RCC cells and CHE induced cell cycle arrest and cell viability. | ↓STAT3; ↑ROS; ↑Cle–PARP; ↓Bcl-2; ↑Bax; ↑G2/M phase; ↓cyclin-dependent kinase 1 (CDC2); ↓cyclin B1; ↓MDM-2; ↑ROS; ↑p-elF2alpha; ↑ATF4; ↓Y705 residues of STAT3 | He et al., 2020 [120] |
| Dauricine | 786-O (hypertriploid RCC); A498 (RCC); ACHN (papillary renal cell carcinoma); Caki (clear cell RCC) | 5–20 µM (24 h) | Decreased cell growth and decreased cell viability and increased cell death | ↓Cyclin D1; ↓CDK4; ↓CDK2; ↓pro-caspase-9; ↓Bcl2; ↓MCL-1; ↑p21; ↑Bax | Zhang et al., 2018 [121] |
| Neferine | Caki-1 (ccRCC); ACHN (papillary renal cell carcinoma); A498 (RCC) | 5–25 µM (24 h) | Decreased cell viability | ↑Bcl-2; ↑p65; ↓NF-κB | Kim et al., 2019 [122] |
| Neopapillarine | A498 (RCC); UO31 (RCC) | 20, 67 µM | Decreased cell growth | Not reported | Tosun et al., 2020 [123] |
| Oxymatrine | A498 (RCC); SW839 (ccRCC) | 0.5–10 mg/mL (24–72 h) | Decreased cell viability | ↓Ki-67; ↓cyclin D1; ↓CDK6; ↑p27; ↑caspase-3; ↑cleaved PARP; ↓Vimetin; ↓MMP2; ↓MMP9; ↑E-cadherin; ↑GSK-3β; ↓β-catenin translocation | Jin et al., 2020 [124] |
| Piperlongumine | 786-O (RCC) | 2.5–20 µM (24, 72 h) | Decreased cell viability and cell growth | ↑Apoptosis; ↓LC3-II; ↓p-mTORC1; ↓GSK-3β; ↓TSC2; ↓Akt/mTORC1; ↑ROS; ↓p-Ser757 ULK1 | Makhov et al., 2014 [125] |
| Piperlongumine | RCC 786-O (hypertriploid RCC); PNX0010 (ccRCC) | 2.5–10 µM/L (24 h) | Inhibited cell growth and cell viability | ↓c-Met; ↓ERK; ↓MAPK; ↓STAT3; ↓NF-κB; ↓Akt; ↓mTOR | Golovine et al., 2015 [126] |
| Tetrandrine | RCC 786-O (hypertriploid RCC); 769-P (RCC) | 0.05–10 mg/mL (24 h) | Inhibited cell growth | ↓NF-κB; ↓MMP-9; ↓p-Akt; ↓p-PI3K; ↓p-PDK1 | Chen et al., 2017 [127] |
| Sulfur-containing compounds | |||||
| Allicin | RCC-9863 (RCC) | 0.016, 0.05, 0.1 mg/mL (48 h) | Decreased cell viability; decreased cell differentiation; decreased chemotactic ability | ↓Bcl-2; ↓VEGF; ↓HIF-1α; ↑Bax | Song et al., 2015 [128] |
| Sulforaphane | Caki-1res, KTCTL-26res, A498res, Caki-1par, KTCTL-26par, and A498par | 1.25 to 20 µM (24–72 h) | Decreased cell growth and migration | ↑G2/M; ↑S phase; ↓Cdk1; ↓pCdk2; ↓Cdk2; ↓cyclin A; ↓cyclin B; ↓p27; ↓p-Akt; ↑p19; ↓p-Rictor; ↓α5; ↓α6; ↓β1; ↓β4; ↓β3; ↑β3; ↓integrin α2 | Juengel et al., 2016 [129] |
| Sulforaphane | A498 (RCC); Caki-1 (ccRCC); KTCL-26 (RCC) | Short term: 5 µM (24 h) Long term: 5 µM (8 wk) | Decreased cell growth | Short term: ↑G2/M; ↑cdk1; ↑cdk2; ↑cyclin A; ↑cyclin B; ↓p-Akt; ↓p-Raptor Long term: ↑G0/G1; ↓cdk1; ↓cdk2; ↓cyclin A; ↓cyclin B; ↓p-Akt; ↓p-Raptor | Juengel et al., 2017 [130] |
| Phytochemicals | Animal Tumor Models | Anticancer Effects | Mechanisms | Dose (Route) | Duration | References |
|---|---|---|---|---|---|---|
| Phenolics | ||||||
| Alpinumisoflavone | BALB/c male nude mice xenografted with 786-O cells | Suppressed tumor growth and metastasis | ↑Apoptosis; ↑miR-101; ↓RLIP76; ↓p-Akt/t-Akt ratio | 40, 80 mg/kg (s.c.) | Not reported | Wang et al., 2017 [39] |
| Apigenin | BALB/c male nude mice xenografted with ACHN cells | Reduced tumor growth and volume | ↓Ki-67 | 30 mg/kg (i.p.) | 21 days | Meng et al., 2017 [41] |
| Chrysin | Male Wistar rats induced by Fe-NTA and DEN | Suppressed renal carcinogenesis | ↑GSH; ↑GR; ↑GPx; ↑catalase; ↑SOD; ↓ODC; ↓proliferation; ↓IL-6; ↓TNFα; ↓PGE2; ↓NF-κBp65; ↓COX-2; ↓iNOS | 20, 40 mg/kg (p.o.) | 16 weeks | Rehman et al., 2013 [131] |
| Coumarin | BALB/c female mice injected with tumor inducing BMK cells | Suppressed tumor growth and metastasis | Not reported | 20 mg/kg (i.p.) | 3 weeks | Ruiz-Marcial, 2007 [132] |
| Cyanidin | BALB/c nude male mice xenografted with ACHN cells | Inhibited tumor growth | ↓Ki-67; | 6 mg/kg (i.p.) | 4 weeks | Liu et al., 2018 [48] |
| Eupafolin | BALB/c male nude mice xenografted with Caki cells | Suppressed tumor growth | ↑Apoptosis | 10 mg/kg (i.p.) | 3 weeks | Han et al., 2016 [52] |
| Gambogic acid | BALB/c male mice xenografted with Caki-1 cells | Decreased tumor size | ↓Ki-67; ↓CD31; ↓p65 | 5 mg/kg (p.o.) | 4 weeks | Jiang et al., 2012 [53] |
| Genistein | C57 BL 6 female mice injected with SMKT R-1 cells | Suppressed angiogenesis | Not reported | 100 μg/mL (dorsal air sac model) | 12 h | Sasamura et al., 2004 [54] |
| Hispidulin | BALB/c nu/nu male mice xenografted with Caki-1 cells | Decreased tumor size | Not reported | 20 mg/kg (i.p.) | 4 weeks | Gao et al., 2015 [57] |
| Hispidulin | BALB/c nu/nu male mice xenografted with Caki-2 cells | Suppressed tumor growth | ↑Apoptosis; ↑cleaved caspase-3; ↑ceramide; ↑p-JNK; ↓SphK1 | 20, 40 mg/kg (i.p.) | 27 days | Gao et al., 2017 [58] |
| Honokiol | BALB/c nude mice xenografted with A-498 cells | Delayed tumor growth | ↑E-cadherin; ↓ZEB2; ↓vimentin; ↓fibronectin | 20 μmol (s.c.) | 21 days | Li et al., 2014 [59] |
| Icaritin | BALB/c female mice xenografted with RENCA cells | Inhibited tumor growth | ↓STAT3; ↓Bcl-xL; ↓cyclin E; ↓VEGF; ↓tumor vasculature | 10 mg/kg (peritumoral injection) | 7 days | Li et al., 2013 [62] |
| Resveratrol | BALB/c female mice xenografted with Renca cells | Inhibited tumor growth | ↑CD8+ T cells; ↑perforin; ↑granzyme B; ↑FasL; ↑Fas; ↓Tregs; ↑IFN-ℽ; ↓IL-6; ↓IL-10; ↓VEGF | 1–5 mg/kg (i.p.) | 22 days | Chen et al., 2015 [133] |
| Resveratrol | Male nude mice xenografted with ACHN cells | Reduced tumor volume | ↑Apoptosis; ↑Bax/Bcl-2 ratio; ↓NLRP3 | 60 mg/kg (gavage injection) | 40 days | Tian et al., 2020 [75] |
| Resveratrol | Male Wistar rats induced with clear cell renal cell carcinoma | Displayed antioxidant and anti-inflammatory properties | ↑Nrf2; ↑HO-1; ↑CAT; ↑SOD; ↑GPx; ↓TBARS; ↓xanthine oxidase; ↓LDH; ↓TGF-β1; ↓IL-6; ↓TNF-α; ↓STAT3; ↑caspase-3; ↓NF-κB | 30 mg/kg (p.o.) | 24 weeks | Kabel et al., 2018 [134] |
| Terpenoids | ||||||
| Artemisinin | Male nude mice subcutaneously injected with UMRC-2 cells | Reduced tumor growth and volume | ↓c-Myc; ↓cyclin D1; ↓PCNA; ↓N-cadherin; ↓Vimetin; ↓Snail; ↑E-cadherin; ↓p-Akt | 20 mg/kg (p.o.) | 2 weeks | Yu et al., 2019 [80] |
| Auraptene | Nude male mice xenografted with RCC4 cells | Reduced tumor growth | ↓Vegf-a; ↓angiogenesis | 100 μM (s.c.) | 25 days | Jang et al., 2015 [82] |
| Betulinic acid | Female BALB/c nude mice injected with 786-O cells | Impedes tumor growth | ↓Ki-67-positive; ↓MMP9 | 5, 10 mg/kg (p.o.) | 15 days | Yang et al., 2018 [85] |
| Cafestol; Cafestol + ABT-737 | Male BALB/c-nude mice injected with Mcl-1-overexpressing Caki cells in combination with ABT-737 | Suppressed tumor growth and increased cell death | ↑Apoptosis; ↑caspase 3 | 75 mg/kg (i.p.) | 2 weeks | Woo et al., 2014 [87] |
| Lycopene | Female Eker rats that carry the Eker TSC2 mutation | Reduced number and size of renal carcinomas | Not reported | 100, 200 mg (p.o.) | 18 months | Sahin et al., 2015 [135] |
| Oridonin | Nude mice injected subcutaneously with 786-O cells | Decrease in tumor weight | ↑cleaved caspase-3; ↑Bax, ↓Bcl-2; ↑RIP-1 and RIP-3; ↑LDH; ↑HGMB1; ↑Parp-1; ↓GSH; ↑p-JN; p-ERK; p-P38 | 20 mg/kg (i.p.) | 32 days | Zheng et al., 2018 [104] |
| Parthenolide | Nude male mice inoculated with OUR-10 cells | Reduction in tumor volume | ↓NF-κB; ↓NF-κB p65; ↓p-NF-κB; ↑IκBα; ↓Bcl-xL; ↓COX-2; ↓MMP-9 | 10 mg/kg (p.o.) 3 μg/mouse (s.c.) | 6 weeks | Oka et al., 2007 [105] |
| Silibinin | Nude mice with subcutaneous inoculation of RCC 786-O cells | Reduction in tumor volume | Not reported | 200 mg/kg (p.o.) | 44 days | Chang et al., 2011 [106] |
| Thymoquinone | Five-week-old male C57BL/6 mice | Inhibited RCC spread to lungs | ↓MMP-2; ↓u-PA; ↓p-phosphatidylinositol 3-kinase; ↓p-Akt; ↓p-Src; ↓p-Paxillin; ↓fibronectin; ↓N-cadherin; ↓Rho A; ↓TGF-β-promoted u-PA activity; ↑adhesion type I collagen; ↑adhesion type IV collagen | 10 mg/kg or 20 mg/kg per day (p.o.) | 42 days | Liou et al. 2019 [109] |
| Zerumbone | Athymic nu/nu female mice inoculated with 786-O cells | Decreased tumor growth | ↓Ki-67; ↓Bcl-2; ↑caspase-3; ↓CD31; ↑SHP-1; ↓p-STAT2 | 50 mg/kg (i.p.) | 6 weeks | Shanmugam et al., 2015 [115] |
| Alkaloids | ||||||
| (-)-Antofine | Athymic mice (BALB/c-nu) injected with A Caki-1 cells | Decreased tumor growth | ↓Met-mediated STAT | 5 mg/kg, five times a week (p.o.) | 42 days | Song et al., 2015 [116] |
| Oxymatrine | BALB/c nude mice xenografted with A498 or SW839 cells | Suppressed tumor cell growth | ↑Apoptosis; ↓β-catenin | 50 mg/kg (i.p.) | 28 days | Jin et al., 2020 [124] |
| Piperlongumine | C.B17/Icr-scid mice inoculated with PNX0010 cells | Decreased tumor growth | ↓c-Met | Treated with PL or PL-Di (20 mg/kg, i.p., 3 times/week) | 18 days | Golovine et al., 2015 [126] |
| Agents Tested | Study Subjects | Study Type | Study Population | No. of Patients/Control Subjects | Intervention | Main Findings/Objectives | References |
|---|---|---|---|---|---|---|---|
| Fruits/vegetables and micronutrients | |||||||
| Cruciferous and dark-green vegetables (α-carotene, β-carotene, lutein, β-cryptoxanthin) | 1276 cases of confirmed RCC among residents of Los Angeles, CA | Case–control | USA | 1276/1204 | Dietary intake of cruciferous vegetables was determined via a validated food frequency questionnaire | Dietary intake of cruciferous vegetables was associated with a decreased risk that may be explained by a variety of carotenoids. | Yuan et al., 1998 [275] |
| Carotenoids, vitamin A, and C | 88,759 women in the Nurses’ Health Study and 47,828 men in the Health Professionals Follow-up Study with 248 developing RCC | Prospective cohort study | USA | 136,587 | Dietary intake of fruit and vegetables was determined via a validated semiquantitative food frequency questionnaire implemented every 2–4 years | Dietary intake of fruits and vegetables was associated with a decreased risk of RCC in men, but not in women. Intake of vitamin A and C and carotenoids from food was inversely associated with a decreased risk of RCC in men. | Lee et al., 2006 [276] |
| Carotenoids including α-carotene and lutein/zeaxanthin | 1478 cases of RCC among 530,469 women and 244,483 men | A pooled analysis of 13 prospective studies | USA | 774,952 | Dietary intake of fruit and vegetables was determined via a validated food frequency questionnaire implemented | Dietary intake of fruits and vegetables was associated with a decreased risk of RCC and may be due to certain carotenoids contained within the diet. | Lee et al., 2009 [277] |
| Vitamin E and vitamin C | Hospitalized patients with confirmed RCC | Case–control | Italy | 767/1534 | Dietary habits were determined by food frequency questionnaire | A significant inverse relationship between dietary vitamin E intake and borderline significant inverse relationship of vitamin C intake with associated risk of RCC. | Bosetti et al., 2006 [278] |
| Flavonoids | Patients with RCC | Case–control | Italy | 767/1534 | Intake of flavonoids was determined via diet questionnaire | Inverse association between flavonoid intake and the risk of developing RCC. | Bosetti et al., 2007 [279] |
| Carotenoids, vitamin C, and E | 1138 cases of confirmed RCC | Case-control | Canada | 1138/5039 | Dietary intake was determined via a 69-item food questionnaire | Dietary intake of β-carotene and lutein/zeaxanthin was inversely associated with RCC. This association was more pronounced in women as well as overweight and obese subjects. There was no significant association between vitamin C, vitamin E, β-cryptoxanthin, and lycopene with RCC. | Hu et al., 2009 [280] |
| β-cryptoxanthin and vegetable fiber | Residents of Iowa between the ages of 40–85 with RCC | Case–control | USA | 323/1827 | Dietary habits were determined by food frequency questionnaire | Dietary intake of vegetable fiber was associated with a decreased risk of RCC, but this was not observed for fruit or grain fiber. Additionally, β-cryptoxanthin was the only micronutrient associated with a significant decreased risk of RCC. | Brock et al., 2012 [281] |
| α-carotene, β-carotene, lutein zeaxanthin, lycopene, vitamin A, folate, thiamin, vitamin C, α-tocopherol, β-tocopherol, γ-tocopherol, selenium, and β-cryptoxanthin | European, American, and African American men and women with newly diagnosed RCC | Population-based case–control | USA | 1142/1154 | Dietary micronutrient intake was derived from an interviewer-administered diet history questionnaire (DHQ) and queried frequency of intake for foods, beverages, multivitamin, vitamin C, and vitamin E supplements, for a total of 80 line items. | Inverse associations with RCC risk were observed for α-carotene, β-carotene, lutein zeaxanthin, lycopene, vitamin A, folate, thiamin, vitamin C, α-tocopherol, β-tocopherol, γ-tocopherol, and selenium. A trend for β-cryptoxanthin was suggested among EA but not AA or the total sample (P-interaction = 0.04). | Bock et al., 2018 [282] |
| Lycopene | Postmenopausal women | Prospective cohort study | USA | 96,196 | Dietary micronutrient intake was estimated from the baseline WHI food frequency questionnaire, and data on supplement use were collected using an interview-based inventory procedure. Study conducted over 11–12 years | Inverse relationship with lycopene intake and risk of developing RCC. No associations with intake of α-carotene, β-carotene, β-cryptoxanthin, lutein plus zeaxanthin, vitamin C, and vitamin E. | Ho et al., 2015 [283] |
| β-carotene, α-tocopherol, and retinol | Male smokers | Prospective cohort study | Finland | 27,062 | 276-item dietary questionnaire with pre-randomization serum β-carotene, retinol, and α-tocopherol levels. Additionally daily supplementation with β-carotene (20 mg) and/or vitamin E (50 mg) during up to 19 years follow-up | Overall, no association between fruit, vegetables, antioxidant nutrients and risk of renal cell carcinoma. | Bertoia et al., 2010 [284] |
| Phytochemicals with other agents | |||||||
| Coumarin + cimetidine | Patients with metastatic RCC | Pilot study | Not specified | 45 | 100 mg coumarin p.o. daily followed by 300 mg cimetidine four times a day on day 15 | Three patients with complete remission, eleven patients with partial remission. Twelve patients experienced stabilization of disease. | Marshall et al., 1987 [285] |
| Coumarin + cimetidine | Patients with RCC | Phase II trial | Germany | 31 | 100 mg coumarin p.o. q.d. followed by 400 mg cimetidine four times a day on day 15 | Two patients achieved partial responses. One mixed response, and five with stable disease. | Hermann et al., 1990 [286] |
| Coumarin + Cimetidine | Patients with metastatic RCC | Pilot | USA | 50 | 100 mg coumarin p.o daily starting on day 1 and cimetidine 300 mg p.o four times a day starting on day 15. When disease progressed, coumarin was escalated to 100 mg orally four times a day | Three patients achieved a partial response, one of those after dose escalation. In addition, one patient had a minor response, then progressing disease, and again had a minor response after dose escalation. | Dexeus et al., 1990 [287] |
| Coumarin + cimetidine | Patients with metastatic RCC | Pilot study | Austria | 39 | 400 mg cimetidine p.o. daily followed by 100 mg coumarin p.o. daily after 1 week | Complete remission and partial remission of 5 patients. | Kokron et al., 1991 [288] |
| Coumarin + IFN-α | Patients with RCC | Randomized prospective multicenter phase III trial | Austria | 148 | 100 mg coumarin p.o. daily, IFN-a-2b (5 MU) 5 × weekly s.c. | No significant increase in response rate or prolongation of survival times. | Sagaster et al., 1995 [289] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajalia, E.M.; Azzouz, F.B.; Chism, D.A.; Giansiracusa, D.M.; Wong, C.G.; Plaskett, K.N.; Bishayee, A. Phytochemicals for the Prevention and Treatment of Renal Cell Carcinoma: Preclinical and Clinical Evidence and Molecular Mechanisms. Cancers 2022, 14, 3278. https://doi.org/10.3390/cancers14133278
Bajalia EM, Azzouz FB, Chism DA, Giansiracusa DM, Wong CG, Plaskett KN, Bishayee A. Phytochemicals for the Prevention and Treatment of Renal Cell Carcinoma: Preclinical and Clinical Evidence and Molecular Mechanisms. Cancers. 2022; 14(13):3278. https://doi.org/10.3390/cancers14133278
Chicago/Turabian StyleBajalia, Essa M., Farah B. Azzouz, Danielle A. Chism, Derrek M. Giansiracusa, Carina G. Wong, Kristina N. Plaskett, and Anupam Bishayee. 2022. "Phytochemicals for the Prevention and Treatment of Renal Cell Carcinoma: Preclinical and Clinical Evidence and Molecular Mechanisms" Cancers 14, no. 13: 3278. https://doi.org/10.3390/cancers14133278