
It Takes Two to Tango: A Review of Oncogenic Virus and Host Microbiome Associated Inflammation in Head and Neck Cancer

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
3. Results
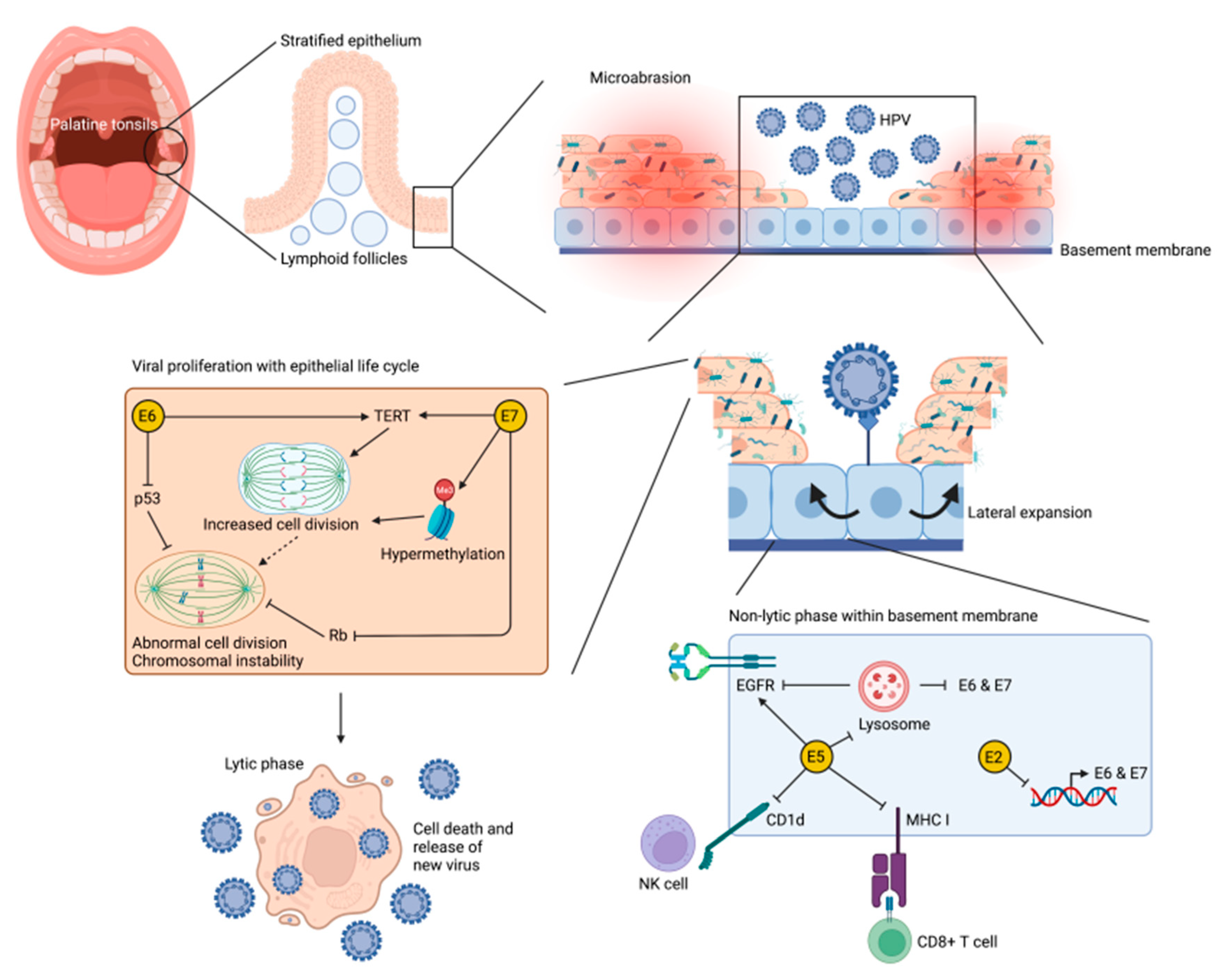
3.1. Human Papillomavirus and Oropharyngeal Carcinoma
3.2. Cellullar Mechanisims of HPV-Mediated Oropharyngeal Carcinogenesis
3.3. Role of the Immune System in HPV-Mediated Oropharyngeal Carcinogenesis
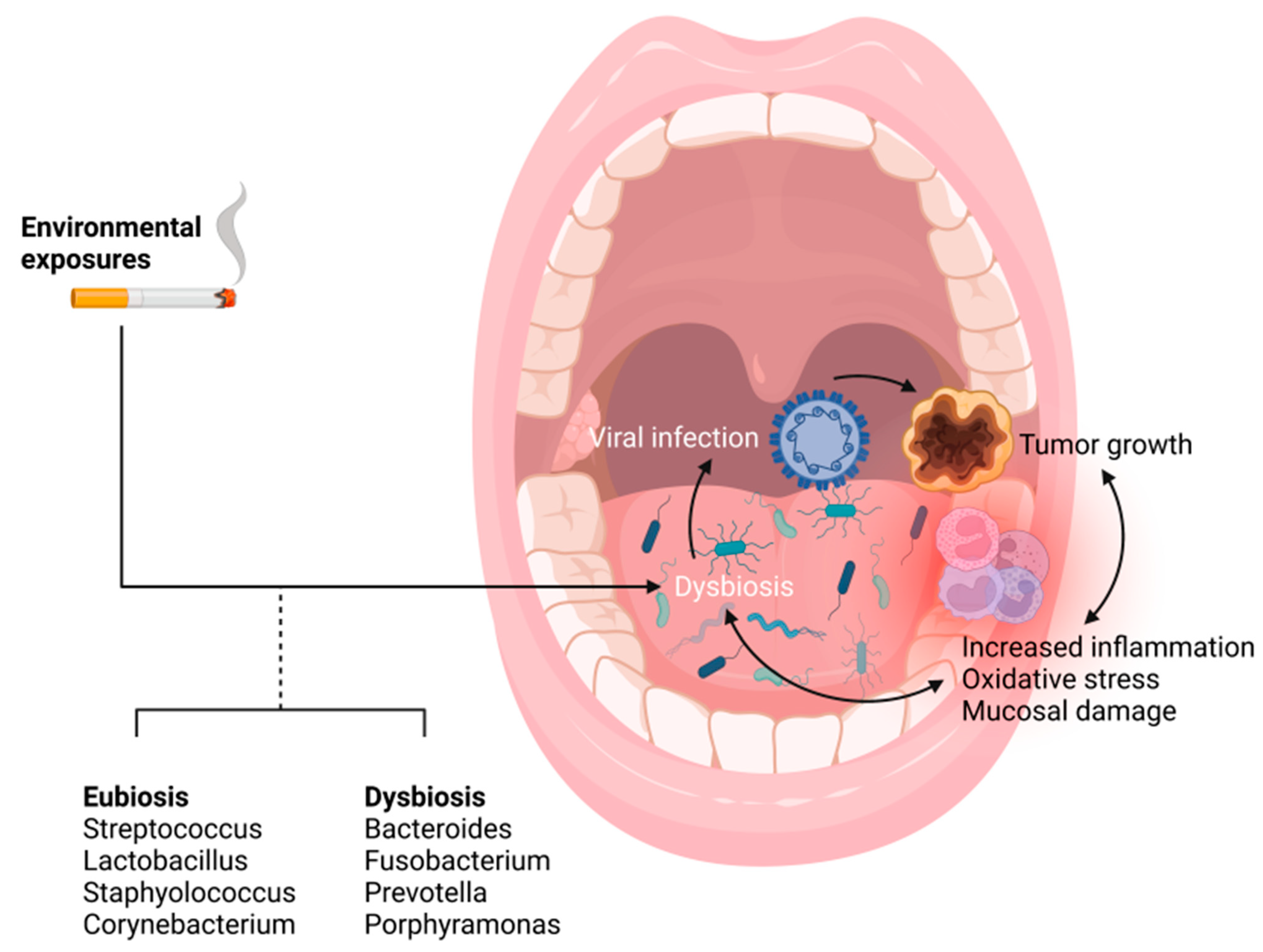
3.4. The Microbiome as a Primer for Inflammation and HPV Mediated Tumor Progression
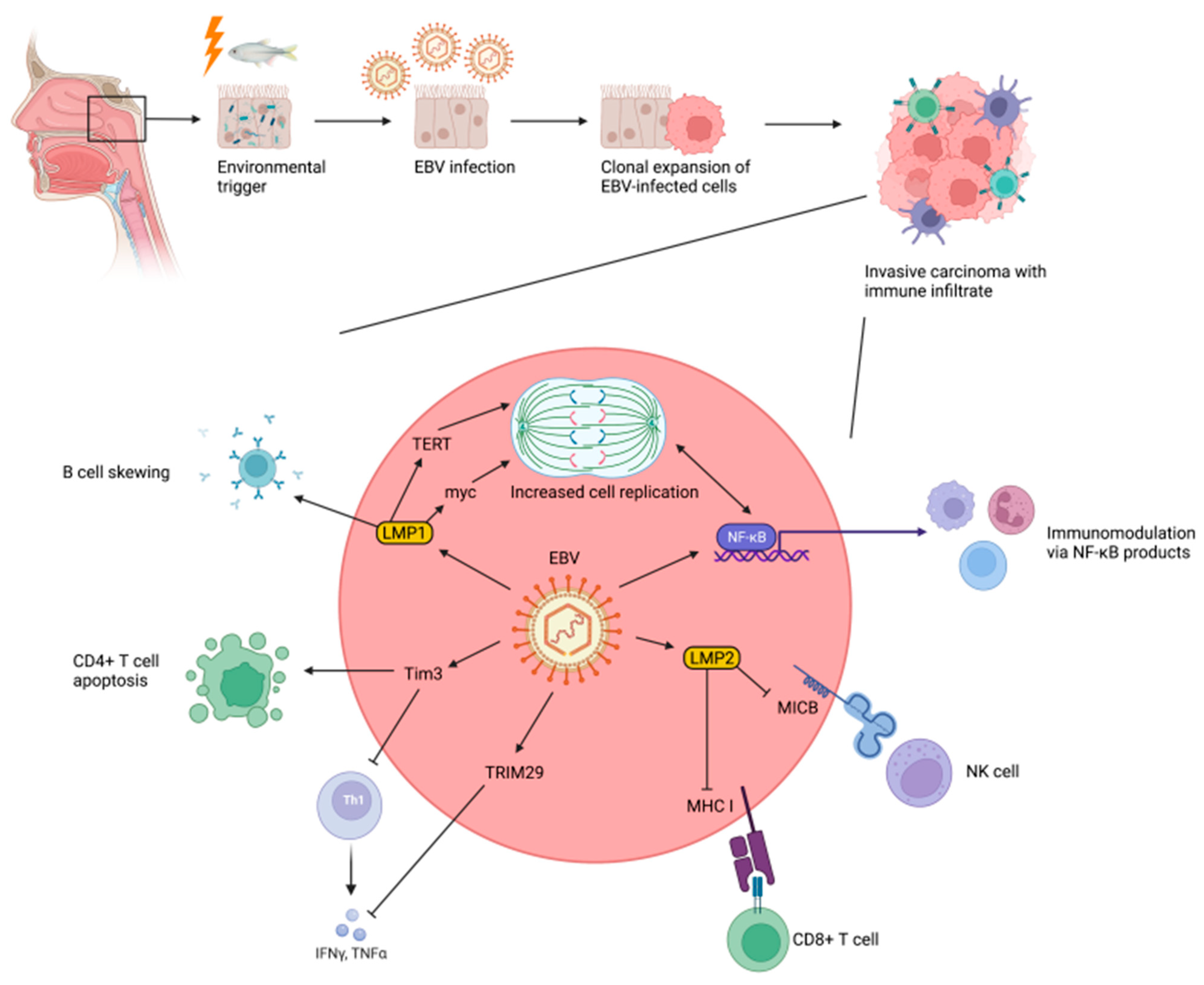
3.5. Epstein–Barr Virus and Nasopharyngeal Carcinogenesis
3.6. Cellular Mechanisms of EBV-Mediated Nasopharyngeal Carcinogenesis
3.7. Role of the Immune System in EBV-Mediated Nasopharyngeal Carcinogenesis
3.8. Does the Microbiome Play a Role in Nasopharyngeal Carcinogenesis?
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yamashita, Y.; Takeshita, T. The Oral Microbiome and Human Health. J. Oral Sci. 2017, 59, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.; Garg, P.K.; Dubey, A.K. Insights into the Human Oral Microbiome. Arch. Microbiol. 2018, 200, 525–540. [Google Scholar] [CrossRef]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [Green Version]
- Irfan, M.; Delgado, R.Z.R.; Frias-Lopez, J. The Oral Microbiome and Cancer. Front. Immunol. 2020, 11, 591088. [Google Scholar] [CrossRef]
- Corbella, S.; Veronesi, P.; Galimberti, V.; Weinstein, R.; Fabbro, M.D.; Francetti, L. Is Periodontitis a Risk Indicator for Cancer? A Meta-Analysis. PLoS ONE 2018, 13, e0195683. [Google Scholar] [CrossRef] [Green Version]
- Welch, J.L.M.; Ramírez-Puebla, S.T.; Borisy, G.G. Oral Microbiome Geography: Micron-Scale Habitat and Niche. Cell Host Microbe 2020, 28, 160–168. [Google Scholar] [CrossRef]
- Kumpitsch, C.; Koskinen, K.; Schöpf, V.; Moissl-Eichinger, C. The Microbiome of the Upper Respiratory Tract in Health and Disease. BMC Biol. 2019, 17, 87. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, A.K.; Engels, E.A.; Pfeiffer, R.M.; Hernandez, B.Y.; Xiao, W.; Kim, E.; Jiang, B.; Goodman, M.T.; Sibug-Saber, M.; Cozen, W.; et al. Human Papillomavirus and Rising Oropharyngeal Cancer Incidence in the United States. J. Clin. Oncol. 2011, 29, 4294–4301. [Google Scholar] [CrossRef]
- Klussmann, J.P.; Dinh, S.; Guntinas-Lichius, O.; Wittekindt, C.; Weissenborn, S.; Wieland, U.; Dienes, H.P.; Hoffmann, T.; Smith, E.; Turek, L.; et al. HPV-Associated Tonsillar Cancer. An Update. HNO 2004, 52, 208–218. [Google Scholar] [CrossRef]
- Marur, S.; D’Souza, G.; Westra, W.H.; Forastiere, A.A. HPV-Associated Head and Neck Cancer: A Virus-Related Cancer Epidemic. Lancet Oncol. 2010, 11, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Gillison, M.L. Human Papillomavirus-Related Diseases: Oropharynx Cancers and Potential Implications for Adolescent HPV Vaccination. J. Adolesc. Health 2008, 43, S52–S60. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, D.; Golz, A.; Netzer, A.; Rosenblatt, E.; Rachmiel, A.; Goldenberg, R.F.; Joachims, H.Z. Epstein-Barr Virus and Cancers of the Head and Neck. Am. J. Otolaryngol. 2001, 22, 197–205. [Google Scholar] [CrossRef]
- Tsang, C.M.; Deng, W.; Yip, Y.L.; Zeng, M.-S.; Lo, K.W.; Tsao, S.W. Epstein-Barr Virus Infection and Persistence in Nasopharyngeal Epithelial Cells. Chin. J. Cancer 2014, 33, 549–555. [Google Scholar] [CrossRef]
- Huang, W.B.; Chan, J.Y.W.; Liu, D.L. Human Papillomavirus and World Health Organization Type III Nasopharyngeal Carcinoma: Multicenter Study from an Endemic Area in Southern China. Cancer 2018, 124, 530–536. [Google Scholar] [CrossRef]
- Shukla, S.; Bharti, A.C.; Mahata, S.; Hussain, S.; Kumar, R.; Hedau, S.; Das, B.C. Infection of Human Papillomaviruses in Cancers of Different Human Organ Sites. Indian J. Med. Res. 2009, 130, 222–233. [Google Scholar]
- Hausen, H. zur Papillomaviruses and Cancer: From Basic Studies to Clinical Application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar] [CrossRef]
- Han, J.J.; Beltran, T.H.; Song, J.W.; Klaric, J.; Choi, Y.S. Prevalence of Genital Human Papillomavirus Infection and Human Papillomavirus Vaccination Rates Among US Adult Men: National Health and Nutrition Examination Survey (NHANES) 2013–2014. JAMA Oncol. 2017, 3, 810. [Google Scholar] [CrossRef]
- Dunmire, S.K.; Verghese, P.S.; Balfour, H.H. Primary Epstein-Barr Virus Infection. J. Clin. Virol. 2018, 102, 84–92. [Google Scholar] [CrossRef]
- Fernandes, J.V.; de Fernandes, T.A.A.M.; de Azevdeo, J.C.V.; Cobucci, R.N.O.; de Carvalho, M.G.F.; Andrade, V.S.; de Araujo, J.M.G. Link between Chronic Inflammation and Human Papillomavirus-Induced Carcinogenesis (Review). Oncol. Lett. 2015, 9, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Feller, L.; Altini, M.; Lemmer, J. Inflammation in the Context of Oral Cancer. Oral Oncol. 2013, 49, 887–892. [Google Scholar] [CrossRef]
- Heusinkveld, M.; Goedemans, R.; Briet, R.J.P.; Gelderblom, H.; Nortier, J.W.R.; Gorter, A.; Smit, V.T.H.B.M.; Langeveld, A.P.M.; Jansen, J.C.; Burg, S.H. van der Systemic and Local Human Papillomavirus 16-specific T-cell Immunity in Patients with Head and Neck Cancer. Int. J. Cancer 2012, 131, E74–E85. [Google Scholar] [CrossRef]
- Bruce, J.P.; To, K.-F.; Lui, V.W.Y.; Chung, G.T.Y.; Chan, Y.-Y.; Tsang, C.M.; Yip, K.Y.; Ma, B.B.Y.; Woo, J.K.S.; Hui, E.P.; et al. Whole-Genome Profiling of Nasopharyngeal Carcinoma Reveals Viral-Host Co-Operation in Inflammatory NF-ΚB Activation and Immune Escape. Nat. Commun. 2021, 12, 4193. [Google Scholar] [CrossRef]
- Lun, S.W.-M.; Cheung, S.-T.; Lo, K.-W. Cancer Stem-like Cells in Epstein-Barr Virus-Associated Nasopharyngeal Carcinoma. Chin. J. Cancer 2014, 33, 529–538. [Google Scholar] [CrossRef]
- Andersen, A.S.; Sølling, A.S.K.; Ovesen, T.; Rusan, M. The Interplay between HPV and Host Immunity in Head and Neck Squamous Cell Carcinoma. Int. J. Cancer 2014, 134, 2755–2763. [Google Scholar] [CrossRef]
- Colman, G.; Beighton, D.; Chalk, A.J.; Wake, S. Cigarette Smoking and the Microbial Flora of the Mouth. Aust. Dent. J. 1976, 21, 111–118. [Google Scholar] [CrossRef]
- Morris, A.; Beck, J.M.; Schloss, P.D.; Campbell, T.B.; Crothers, K.; Curtis, J.L.; Flores, S.C.; Fontenot, A.P.; Ghedin, E.; Huang, L.; et al. Comparison of the Respiratory Microbiome in Healthy Nonsmokers and Smokers. Am. J. Respir. Crit. Care 2013, 187, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Mason, M.R.; Preshaw, P.M.; Nagaraja, H.N.; Dabdoub, S.M.; Rahman, A.; Kumar, P.S. The Subgingival Microbiome of Clinically Healthy Current and Never Smokers. ISME J. 2015, 9, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Sampaio-Maia, B.; Caldas, I.M.; Pereira, M.L.; Pérez-Mongiovi, D.; Araujo, R. Chapter Four The Oral Microbiome in Health and Its Implication in Oral and Systemic Diseases. Adv. Appl. Microbiol. 2016, 97, 171–210. [Google Scholar] [CrossRef]
- Lim, Y.; Totsika, M.; Morrison, M.; Punyadeera, C. Oral Microbiome: A New Biomarker Reservoir for Oral and Oropharyngeal Cancers. Theranostics 2017, 7, 4313–4321. [Google Scholar] [CrossRef]
- Tada, A.; Senpuku, H. The Impact of Oral Health on Respiratory Viral Infection. Dent. J. 2021, 9, 43. [Google Scholar] [CrossRef]
- Shetty, S.S.; Padam, K.S.R.; Hunter, K.D.; Kudva, A.; Radhakrishnan, R. Biological Implications of the Immune Factors in the Tumour Microenvironment of Oral Cancer. Arch. Oral Biol. 2021, 133, 105294. [Google Scholar] [CrossRef]
- Zong, Y.; Zhou, Y.; Liao, B.; Liao, M.; Shi, Y.; Wei, Y.; Huang, Y.; Zhou, X.; Cheng, L.; Ren, B. The Interaction Between the Microbiome and Tumors. Front. Cell Infect. Microbiol. 2021, 11, 673724. [Google Scholar] [CrossRef]
- Kurago, Z.; Loveless, J. Microbial Colonization and Inflammation as Potential Contributors to the Lack of Therapeutic Success in Oral Squamous Cell Carcinoma. Front. Oral Health 2021, 2, 739499. [Google Scholar] [CrossRef]
- Petca, A.; Borislavschi, A.; Zvanca, M.E.; Petca, R.-C.; Sandru, F.; Dumitrascu, M.C. Non-Sexual HPV Transmission and Role of Vaccination for a Better Future (Review). Exp. Ther. Med. 2020, 20, 186. [Google Scholar] [CrossRef]
- Pytynia, K.B.; Dahlstrom, K.R.; Sturgis, E.M. Epidemiology of HPV-Associated Oropharyngeal Cancer. Oral Oncol. 2014, 50, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Bohr, A.; Grønhøj, C.; Lajer, C.; Gerstoft, J.; von Buchwald, C. Transmission of and Infection with Human Papillomavirus in the Oropharynx. Ugeskr. Laeg. 2017, 179, V10160700. [Google Scholar]
- Szymonowicz, K.A.; Chen, J. Biological and Clinical Aspects of HPV-Related Cancers. Cancer Biol. Med. 2020, 17, 864–878. [Google Scholar] [CrossRef]
- El-Mofty, S.K. Human Papillomavirus (HPV) Related Carcinomas of the Upper Aerodigestive Tract. Head Neck Pathol. 2007, 1, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.O.; da Silva, S.P.; Costa, N.R.; da Costa, R.M.G.; Medeiros, R. The Role of MicroRNAs in the Metastatic Process of High-Risk HPV-Induced Cancers. Cancers 2018, 10, 493. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Kumar, P.; Das, B.C. HPV: Molecular Pathways and Targets. Curr. Prob. Cancer 2018, 42, 161–174. [Google Scholar] [CrossRef]
- Ilmarinen, T.; Munne, P.; Hagström, J.; Haglund, C.; Auvinen, E.; Virtanen, E.I.; Haesevoets, A.; Speel, E.J.M.; Aaltonen, L.-M. Prevalence of High-Risk Human Papillomavirus Infection and Cancer Gene Mutations in Nonmalignant Tonsils. Oral Oncol. 2017, 73, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herberhold, S.; Hellmich, M.; Panning, M.; Bartok, E.; Silling, S.; Akgül, B.; Wieland, U. Human Polyomavirus and Human Papillomavirus Prevalence and Viral Load in Non-Malignant Tonsillar Tissue and Tonsillar Carcinoma. Med. Microbiol. Immun. 2016, 206, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Venuti, A.; Paolini, F.; Nasir, L.; Corteggio, A.; Roperto, S.; Campo, M.S.; Borzacchiello, G. Papillomavirus E5: The Smallest Oncoprotein with Many Functions. Mol. Cancer 2011, 10, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crusius, K.; Auvinen, E.; Steuer, B.; Gaissert, H.; Alonso, A. The Human Papillomavirus Type 16 E5-Protein Modulates Ligand-Dependent Activation of the EGF Receptor Family in the Human Epithelial Cell Line HaCaT. Exp. Cell Res. 1998, 241, 76–83. [Google Scholar] [CrossRef]
- Miura, S.; Kawana, K.; Schust, D.J.; Fujii, T.; Yokoyama, T.; Iwasawa, Y.; Nagamatsu, T.; Adachi, K.; Tomio, A.; Tomio, K.; et al. CD1d, a Sentinel Molecule Bridging Innate and Adaptive Immunity, Is Downregulated by the Human Papillomavirus (HPV) E5 Protein: A Possible Mechanism for Immune Evasion by HPV. J. Virol. 2010, 84, 11614–11623. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, B.; Ashrafi, G.H.; Tsirimonaki, E.; O’Brien, P.M.; Campo, M.S. The Bovine Papillomavirus Oncoprotein E5 Retains MHC Class I Molecules in the Golgi Apparatus and Prevents Their Transport to the Cell Surface. Oncogene 2002, 21, 7808–7816. [Google Scholar] [CrossRef] [Green Version]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Briones-Herrera, A.; Pedraza-Chaverri, J. Regulation of Autophagy by High- and Low-risk Human Papillomaviruses. Rev. Med. Virol. 2021, 31, e2169. [Google Scholar] [CrossRef]
- Hoppe-Seyler, K.; Bossler, F.; Braun, J.A.; Herrmann, A.L.; Hoppe-Seyler, F. The HPV E6/E7 Oncogenes: Key Factors for Viral Carcinogenesis and Therapeutic Targets. Trends Microbiol. 2018, 26, 158–168. [Google Scholar] [CrossRef]
- Doorslaer, K.V.; Burk, R.D. Association between HTERT Activation by HPV E6 Proteins and Oncogenic Risk. Virology 2012, 433, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Burgers, W.A.; Blanchon, L.; Pradhan, S.; de Launoit, Y.; Kouzarides, T.; Fuks, F. Viral Oncoproteins Target the DNA Methyltransferases. Oncogene 2007, 26, 1650–1655. [Google Scholar] [CrossRef] [Green Version]
- Chi, A.C.; Day, T.A.; Neville, B.W. Oral Cavity and Oropharyngeal Squamous Cell Carcinoma—An Update. CA Cancer J. Clin. 2015, 65, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Gandini, S.; Botteri, E.; Iodice, S.; Boniol, M.; Lowenfels, A.B.; Maisonneuve, P.; Boyle, P. Tobacco Smoking and Cancer: A Meta-analysis. Int. J. Cancer 2008, 122, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.B.; Edge, S.B.; Greene, F.L.; Byrd, D.R.; Brookland, R.K.; Washington, M.K.; Gershenwald, J.E.; Compton, C.C.; Hess, K.R.; Sullivan, D.C.; et al. (Eds.) AJCC Cancer Staging Manual, 8th ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-319-40617-6. [Google Scholar]
- NCCN. NCCN Guidelines Version 1. 2022 Head and Neck Cancers; NCCN: Plymouth Meeting, PA, USA, 2022. [Google Scholar]
- Cohan, D.M.; Popat, S.; Kaplan, S.E.; Rigual, N.; Loree, T.; Hicks, W.L. Oropharyngeal Cancer: Current Understanding and Management. Curr. Opin. Otolaryngol. 2009, 17, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernster, J.A.; Sciotto, C.G.; O’Brien, M.M.; Robinson, L.J.; Willson, T. Prevalence of Oncogenic Human Papillomavirus 16 and 18 in the Palatine Tonsils of the General Adult Population. Arch. Otolaryngol. Head Neck Surg. 2009, 135, 554–557. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.I.; Alawi, F. Human Papillomavirus and Oropharyngeal Cancer. Dent. Clin. N. Am. 2018, 62, 111–120. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, G.; Kreimer, A.R.; Viscidi, R.; Pawlita, M.; Fakhry, C.; Koch, W.M.; Westra, W.H.; Gillison, M.L. Case–Control Study of Human Papillomavirus and Oropharyngeal Cancer. N. Engl. J. Med. 2007, 356, 1944–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreimer, A.R.; Bhatia, R.K.; Messeguer, A.L.; González, P.; Herrero, R.; Giuliano, A.R. Oral Human Papillomavirus in Healthy Individuals; A Systematic Review of the Literature. Sex. Transm. Dis. 2010, 37, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Lee, M.; Cho, K.-J.; Kim, S.B.; Roh, J.-L.; Choi, S.-H.; Nam, S.Y.; Kim, S.Y.; Song, J.S. Association Between Fibroblast Growth Factor Receptor 1 Gene Amplification and Human Papillomavirus Prevalence in Tonsillar Squamous Cell Carcinoma With Clinicopathologic Analysis. J. Histochem. Cytochem. 2018, 66, 511–522. [Google Scholar] [CrossRef]
- Ilahi, N.E.; Bhatti, A. Impact of HPV E5 on Viral Life Cycle via EGFR Signaling. Microb. Pathog. 2019, 139, 103923. [Google Scholar] [CrossRef]
- Gutierrez-Xicotencatl, L.; Pedroza-Saavedra, A.; Chihu-Amparan, L.; Salazar-Piña, A.; Maldonado-Gama, M.; Esquivel-Guadarrama, F. Cellular Functions of HPV16 E5 Oncoprotein during Oncogenic Transformation. Mol. Cancer Res. 2021, 19, 167–179. [Google Scholar] [CrossRef]
- Tornesello, M.L.; Annunziata, C.; Tornesello, A.L.; Buonaguro, L.; Buonaguro, F.M. Human Oncoviruses and P53 Tumor Suppressor Pathway Deregulation at the Origin of Human Cancers. Cancers 2018, 10, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustafa, A.-E.A.; Foulkes, W.D.; Benlimame, N.; Wong, A.; Yen, L.; Bergeron, J.; Batist, G.; Alpert, L.; Alaoui-Jamali, M.A. E6/E7 Proteins of HPV Type 16 and ErbB-2 Cooperate to Induce Neoplastic Transformation of Primary Normal Oral Epithelial Cells. Oncogene 2004, 23, 350–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtig, H.; Gilboa, D.A.; Jackman, A.; Gonen, P.; Levav-Cohen, Y.; Haupt, Y.; Sherman, L. HPV16 E6 Augments Wnt Signaling in an E6AP-Dependent Manner. Virology 2010, 396, 47–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Shimane, T.; Kawano, S.; Alshaikh, A.; Kim, S.Y.; Chung, S.H.; Kim, R.H.; Shin, K.H.; Walentin, K.; Park, N.H.; et al. Human Papillomavirus 16 E6 Induces FoxM1B in Oral Keratinocytes through GRHL2. J. Dent. Res. 2018, 97, 795–802. [Google Scholar] [CrossRef]
- Spanos, W.C.; Geiger, J.; Anderson, M.E.; Harris, G.F.; Bossler, A.D.; Smith, R.B.; Klingelhutz, A.J.; Lee, J.H. Deletion of the PDZ Motif of HPV16 E6 Preventing Immortalization and Anchorage-Independent Growth in Human Tonsil Epithelial Cells. Head Neck 2008, 30, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Songock, W.K.; Kim, S.; Bodily, J.M. The Human Papillomavirus E7 Oncoprotein as a Regulator of Transcription. Virus Res. 2017, 231, 56–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, A.; Munger, K. The Papillomavirus E7 Proteins. Virology 2013, 445, 138–168. [Google Scholar] [CrossRef] [Green Version]
- Camuzi, D.; de Simão, T.A.; Dias, F.; Pinto, L.F.R.; Soares-Lima, S.C. Head and Neck Cancers Are Not Alike When Tarred with the Same Brush: An Epigenetic Perspective from the Cancerization Field to Prognosis. Cancers 2021, 13, 5630. [Google Scholar] [CrossRef]
- Basukala, O.; Banks, L. The Not-So-Good, the Bad and the Ugly: HPV E5, E6 and E7 Oncoproteins in the Orchestration of Carcinogenesis. Viruses 2021, 13, 1892. [Google Scholar] [CrossRef]
- Hoover, A.C.; Spanos, W.C.; Harris, G.F.; Anderson, M.E.; Klingelhutz, A.J.; Lee, J.H. The Role of Human Papillomavirus 16 E6 in Anchorage-Independent and Invasive Growth of Mouse Tonsil Epithelium. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Einstein, M.H.; Schiller, J.T.; Viscidi, R.P.; Strickler, H.D.; Coursaget, P.; Tan, T.; Halsey, N.; Jenkins, D. Clinician’s Guide to Human Papillomavirus Immunology: Knowns and Unknowns. Lancet Infect. Dis. 2009, 9, 347–356. [Google Scholar] [CrossRef]
- Rosenberger, S.; Arce, J.D.-C.; Langbein, L.; Steenbergen, R.D.M.; Rösl, F. Alternative Splicing of Human Papillomavirus Type-16 E6/E6* Early MRNA Is Coupled to EGF Signaling via Erk1/2 Activation. Proc. Natl. Acad. Sci. USA 2010, 107, 7006–7011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S. Papillomavirus Transcripts and Posttranscriptional Regulation. Virology 2013, 445, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, N. Human Papillomavirus-16 E5 Protein: Oncogenic Role and Therapeutic Value. Cell Oncol. 2012, 35, 67–76. [Google Scholar] [CrossRef] [PubMed]
- McBride, A.A. Human Papillomaviruses: Diversity, Infection and Host Interactions. Nat. Rev. Microbiol. 2022, 20, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Egawa, N.; Shiraz, A.; Katakuse, M.; Okamura, M.; Griffin, H.M.; Doorbar, J. The Reservoir of Persistent Human Papillomavirus Infection; Strategies for Elimination Using Anti-Viral Therapies. Viruses 2022, 14, 214. [Google Scholar] [CrossRef]
- Doorbar, J.; Zheng, K.; Aiyenuro, A.; Yin, W.; Walker, C.M.; Chen, Y.; Egawa, N.; Griffin, H.M. Principles of Epithelial Homeostasis Control during Persistent Human Papillomavirus Infection and Its Deregulation at the Cervical Transformation Zone. Curr. Opin. Virol. 2021, 51, 96–105. [Google Scholar] [CrossRef]
- Barros, M.R.; de Oliveira, T.H.A.; de Melo, C.M.L.; Venuti, A.; de Freitas, A.C. Viral Modulation of TLRs and Cytokines and the Related Immunotherapies for HPV-Associated Cancers. J. Immunol. Res. 2018, 2018, 2912671. [Google Scholar] [CrossRef]
- Jouhi, L.; Datta, N.; Renkonen, S.; Atula, T.; Mäkitie, A.; Haglund, C.; Ahmed, A.; Syrjänen, S.; Grénman, R.; Auvinen, E.; et al. Expression of Toll-like Receptors in HPV-Positive and HPV-Negative Oropharyngeal Squamous Cell Carcinoma—An in Vivo and in Vitro Study. Tumour Biol. 2015, 36, 7755–7764. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Toll-like Receptor Control of the Adaptive Immune Responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.S.; Thorstad, W.L.; Chernock, R.D.; Haughey, B.H.; Yip, J.H.; Zhang, Q.; El-Mofty, S.K. P16 Positive Oropharyngeal Squamous Cell Carcinoma: An Entity with a Favorable Prognosis Regardless of Tumor HPV Status. Am. J. Surg. Pathol. 2010, 34, 1088–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephen, J.K.; Divine, G.; Chen, K.M.; Chitale, D.; Havard, S.; Worsham, M.J. Significance of P16 in Site-Specific HPV Positive and HPV Negative Head and Neck Squamous Cell Carcinoma. Cancer Clin. Oncol. 2013, 2, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luginbuhl, A.; Sanders, M.; Spiro, J.D. Prevalence, Morphology, and Prognosis of Human Papillomavirus in Tonsillar Cancer. Ann. Otol. Rhinol. Laryngol. 2009, 118, 742–749. [Google Scholar] [CrossRef]
- Oguejiofor, K.; Hall, J.; Slater, C.; Betts, G.; Hall, G.; Slevin, N.; Dovedi, S.; Stern, P.L.; West, C.M.L. Stromal Infiltration of CD8 T Cells Is Associated with Improved Clinical Outcome in HPV-Positive Oropharyngeal Squamous Carcinoma. Brit. J. Cancer 2015, 113, 886–893. [Google Scholar] [CrossRef]
- Wilczynski, S.P.; Lin, B.T.; Xie, Y.; Paz, I.B. Detection of Human Papillomavirus DNA and Oncoprotein Overexpression Are Associated with Distinct Morphological Patterns of Tonsillar Squamous Cell Carcinoma. Am. J. Pathol. 1998, 152, 145–156. [Google Scholar]
- Begum, S.; Cao, D.; Gillison, M.; Zahurak, M.; Westra, W.H. Tissue Distribution of Human Papillomavirus 16 DNA Integration in Patients with Tonsillar Carcinoma. Clin. Cancer Res. 2005, 11, 5694–5699. [Google Scholar] [CrossRef] [Green Version]
- Hibbert, J.; Halec, G.; Baaken, D.; Waterboer, T.; Brenner, N. Sensitivity and Specificity of Human Papillomavirus (HPV) 16 Early Antigen Serology for HPV-Driven Oropharyngeal Cancer: A Systematic Literature Review and Meta-Analysis. Cancers 2021, 13, 3010. [Google Scholar] [CrossRef]
- Hladíková, K.; Koucký, V.; Bouček, J.; Laco, J.; Grega, M.; Hodek, M.; Zábrodský, M.; Vošmik, M.; Rozkošová, K.; Vošmiková, H.; et al. Tumor-Infiltrating B Cells Affect the Progression of Oropharyngeal Squamous Cell Carcinoma via Cell-to-Cell Interactions with CD8+ T Cells. J. Immunother. Cancer 2019, 7, 261. [Google Scholar] [CrossRef]
- Machtay, M.; Moughan, J.; Trotti, A.; Garden, A.S.; Weber, R.S.; Cooper, J.S.; Forastiere, A.; Ang, K.K. Factors Associated with Severe Late Toxicity After Concurrent Chemoradiation for Locally Advanced Head and Neck Cancer: An RTOG Analysis. J. Clin. Oncol. 2008, 26, 3582–3589. [Google Scholar] [CrossRef]
- Hanasoge, S.; Magliocca, K.R.; Switchenko, J.M.; Saba, N.F.; Wadsworth, J.T.; El-Deiry, M.W.; Shin, D.M.; Khuri, F.; Beitler, J.J.; Higgins, K.A. Clinical Outcomes in Elderly Patients with Human Papillomavirus–Positive Squamous Cell Carcinoma of the Oropharynx Treated with Definitive Chemoradiation Therapy. Head Neck 2016, 38, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Castle, P.E.; Einstein, M.H.; Sahasrabuddhe, V.V. Cervical Cancer Prevention and Control in Women Living with Human Immunodeficiency Virus. CA Cancer J. Clin. 2021, 71, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-C.J.; Palefsky, J.M. HIV/AIDS-Associated Viral Oncogenesis. Cancer Treat. 2018, 177, 183–209. [Google Scholar] [CrossRef]
- Beachler, D.C.; D’Souza, G. Oral Human Papillomavirus Infection and Head and Neck Cancers in HIV-Infected Individuals. Curr. Opin. Oncol. 2013, 25, 503–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.Y.; Massa, S.; Mazul, A.L.; Kallogjeri, D.; Yaeger, L.; Jackson, R.S.; Zevallos, J.; Pipkorn, P. The Association of Smoking and Outcomes in HPV-Positive Oropharyngeal Cancer: A Systematic Review. Am. J. Otolaryngol. 2020, 41, 102592. [Google Scholar] [CrossRef]
- Villepelet, A.; Hugonin, S.; Atallah, S.; Job, B.; Baujat, B.; Guily, J.L.S.; Lacave, R. Effects of Tobacco Abuse on Major Chromosomal Instability in Human Papilloma Virus 16-Positive Oropharyngeal Squamous Cell Carcinoma. Int. J. Oncol. 2019, 55, 527–535. [Google Scholar] [CrossRef]
- Lechner, M.; Liu, J.; Masterson, L.; Fenton, T.R. HPV-Associated Oropharyngeal Cancer: Epidemiology, Molecular Biology and Clinical Management. Nat. Rev. Clin. Oncol. 2022, 19, 306–327. [Google Scholar] [CrossRef]
- Elinav, E.; Garrett, W.S.; Trinchieri, G.; Wargo, J. The Cancer Microbiome. Nat. Rev. Cancer 2019, 19, 371–376. [Google Scholar] [CrossRef]
- Cullin, N.; Antunes, C.A.; Straussman, R.; Stein-Thoeringer, C.K.; Elinav, E. Microbiome and Cancer. Cancer Cell 2021, 39, 1317–1341. [Google Scholar] [CrossRef]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; von Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the Microbiome and TP53 in Human Lung Cancer. Genome Biol. 2018, 19, 123. [Google Scholar] [CrossRef]
- Kumar, M.; Singh, P.; Murugesan, S.; Vetizou, M.; McCulloch, J.; Badger, J.H.; Trinchieri, G.; Khodor, S.A. Microbiome as an Immunological Modifier. Methods Mol. Biol. Clifton N. J. 2020, 2055, 595–638. [Google Scholar] [CrossRef]
- Kumar, A.; Smith, C.; Jobin, C.; Trinchieri, G.; Howcroft, T.K.; Seifried, H.; Espey, M.G.; Flores, R.; Kim, Y.S.; Daschner, P.J. Workshop Report: Modulation of Antitumor Immune Responses by Dietary and Microbial Metabolites. J. Natl. Cancer Inst. 2017, 109, djx040. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.K.; Trinchieri, G. The Interplay between Neutrophils and Microbiota in Cancer. J. Leukoc. Biol. 2018, 104, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Dzutsev, A.; Badger, J.H.; Perez-Chanona, E.; Roy, S.; Salcedo, R.; Smith, C.K.; Trinchieri, G. Microbes and Cancer. Annu. Rev. Immunol. 2016, 35, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.A.; Garrett, W.S. Fusobacterium Nucleatum—Symbiont, Opportunist and Oncobacterium. Nat. Rev. Microbiol. 2019, 17, 156–166. [Google Scholar] [CrossRef]
- Healy, C.M.; Moran, G.P. The Microbiome and Oral Cancer: More Questions than Answers. Oral Oncol. 2019, 89, 30–33. [Google Scholar] [CrossRef]
- Shillitoe, E.J. The Microbiome of Oral Cancer. Crit. Rev. Oncog. 2018, 23, 153–160. [Google Scholar] [CrossRef]
- Vanhoecke, B.; Stringer, A. Host–Microbe Cross Talk in Cancer Therapy. Curr. Opin. Support. Palliat. Care 2015, 9, 174–181. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kitamura, N.; Yoshida, K.; Yamamoto, T.; Ozaki, K.; Kudo, Y. Involvement of Fusobacterium Species in Oral Cancer Progression: A Literature Review Including Other Types of Cancer. Int. J. Mol. Sci. 2020, 21, 6207. [Google Scholar] [CrossRef]
- Guerrero-Preston, R.; White, J.R.; Godoy-Vitorino, F.; Hilario, A.R.; Navarro, K.; González, H.; Michailidi, C.; Jedlicka, A.; Canapp, S.; Bondy, J.; et al. High-Resolution Microbiome Profiling Uncovers Fusobacterium Nucleatum, Lactobacillus Gasseri/Johnsonii, and Lactobacillus Vaginalis Associated to Oral and Oropharyngeal Cancer in Saliva from HPV Positive and HPV Negative Patients Trea. Oncotarget 2014, 5, 110931–110948. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.M.; Luo, T.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Microbial Communities Associated with Primary and Metastatic Head and Neck Squamous Cell Carcinoma–A High Fusobacterial and Low Streptococcal Signature. Sci. Rep. 2017, 7, 9934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stashenko, P.; Yost, S.; Choi, Y.; Danciu, T.; Chen, T.; Yoganathan, S.; Kressirer, C.; Ruiz-Tourrella, M.; Das, B.; Kokaras, A.; et al. The Oral Mouse Microbiome Promotes Tumorigenesis in Oral Squamous Cell Carcinoma. Msystems 2019, 4, e00323-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, J.-R.; Chang, C.-C.; Lee, W.-T.; Huang, C.-C.; Ou, C.-Y.; Tsai, S.-T.; Chen, K.-C.; Huang, J.-S.; Wong, T.-Y.; Lai, Y.-H.; et al. The Interplay between Oral Microbiome, Lifestyle Factors and Genetic Polymorphisms in the Risk of Oral Squamous Cell Carcinoma. Carcinogenesis 2018, 39, 778–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yin, G.; Guo, Y.; Zhao, Y.; Zhao, M.; Lai, Y.; Sui, P.; Shi, T.; Guo, W.; Huang, Z. Variations in Oral Microbiota Composition Are Associated with a Risk of Throat Cancer. Front. Cell Infect. Microbiol. 2019, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; MacIntyre, D.A.; Marchesi, J.R.; Lee, Y.S.; Bennett, P.R.; Kyrgiou, M. The Vaginal Microbiota, Human Papillomavirus Infection and Cervical Intraepithelial Neoplasia: What Do We Know and Where Are We Going Next? Microbiome 2016, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; D’Souza, G.; Fakhry, C.; Bigelow, E.O.; Usyk, M.; Burk, R.D.; Zhao, N. Oral HPV Associated with Differences in Oral Microbiota Beta Diversity and Microbiota Abundance. J. Infect. Dis. 2022. [Google Scholar] [CrossRef]
- Hayes, R.B.; Ahn, J.; Fan, X.; Peters, B.A.; Ma, Y.; Yang, L.; Agalliu, I.; Burk, R.D.; Ganly, I.; Purdue, M.P.; et al. Association of Oral Microbiome with Risk for Incident Head and Neck Squamous Cell Cancer. JAMA Oncol. 2018, 4, 358. [Google Scholar] [CrossRef]
- Shigeishi, H.; Sugiyama, M.; Ohta, K. Relationship between the Prevalence of Oral Human Papillomavirus DNA and Periodontal Disease (Review). Biomed. Rep. 2021, 14, 40. [Google Scholar] [CrossRef]
- Zakrzewski, M.; Gannon, O.M.; Panizza, B.J.; Saunders, N.A.; Antonsson, A. Human Papillomavirus Infection and Tumor Microenvironment Are Associated with the Microbiota in Patients with Oropharyngeal Cancers—Pilot Study. Head Neck 2021, 43, 3324–3330. [Google Scholar] [CrossRef]
- Astl, J.; Holy, R.; Maute, E.; Rotnágl, J.; Kalfeřt, D.; Drnková, B.; Younus, T.; Pavlík, E. Genome of Helicobacter Pylori and Serotype of HPV Detected in Oropharyngeal and Laryngeal Cancer and Chronic Inflammation Patients. Int. J. Environ. Res. Public Health 2021, 18, 9545. [Google Scholar] [CrossRef]
- Bouland, C.; Dequanter, D.; Lechien, J.R.; Hanssens, C.; Aubain, N.D.S.; Digonnet, A.; Javadian, R.; Yanni, A.; Rodriguez, A.; Loeb, I.; et al. Prognostic Significance of a Scoring System Combining P16, Smoking, and Drinking Status in a Series of 131 Patients with Oropharyngeal Cancers. Int. J. Otolaryngol. 2021, 2021, 8020826. [Google Scholar] [CrossRef] [PubMed]
- Ference, R.; Liao, D.; Gao, Q.; Mehta, V. Impact of Smoking on Survival Outcomes in HPV-Related Oropharyngeal Carcinoma: A Meta-Analysis. Otolaryngol. Head Neck Surg. 2020, 163, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Börnigen, D.; Ren, B.; Pickard, R.; Li, J.; Ozer, E.; Hartmann, E.M.; Xiao, W.; Tickle, T.; Rider, J.; Gevers, D.; et al. Alterations in Oral Bacterial Communities Are Associated with Risk Factors for Oral and Oropharyngeal Cancer. Sci. Rep. 2017, 7, 17686. [Google Scholar] [CrossRef] [PubMed]
- Perera, M.; Al-hebshi, N.N.; Perera, I.; Ipe, D.; Ulett, G.C.; Speicher, D.J.; Chen, T.; Johnson, N.W. Inflammatory Bacteriome and Oral Squamous Cell Carcinoma. J. Dent. Res. 2018, 97, 725–732. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-Y.; Yeh, Y.-M.; Yu, H.-Y.; Chin, C.-Y.; Hsu, C.-W.; Liu, H.; Huang, P.-J.; Hu, S.-N.; Liao, C.-T.; Chang, K.-P.; et al. Oral Microbiota Community Dynamics Associated with Oral Squamous Cell Carcinoma Staging. Front. Microbiol. 2018, 9, 862. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.; Jasinski-Bergner, S.; Mandelboim, O.; Wickenhauser, C.; Seliger, B. Epstein–Barr Virus—Associated Malignancies and Immune Escape: The Role of the Tumor Microenvironment and Tumor Cell Evasion Strategies. Cancers 2021, 13, 5189. [Google Scholar] [CrossRef]
- Epstein, M.A.; Achong, B.G.; Barr, Y.M. Virus Particles in Cultured Lymphoblasts from Burkitt’s Lymphoma. Lancet 1964, 283, 702–703. [Google Scholar] [CrossRef]
- Hsu, J.L.; Glaser, S.L. Epstein–Barr Virus-Associated Malignancies: Epidemiologic Patterns and Etiologic Implications. Crit. Rev. Oncol. 2000, 34, 27–53. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Chan, A.T.C.; Le, Q.-T.; Blanchard, P.; Sun, Y.; Ma, J. Nasopharyngeal Carcinoma. Lancet 2019, 394, 64–80. [Google Scholar] [CrossRef]
- Chua, M.L.K.; Wee, J.T.S.; Hui, E.P.; Chan, A.T.C. Nasopharyngeal Carcinoma. Lancet 2016, 387, 1012–1024. [Google Scholar] [CrossRef]
- Stan, D.J.; Niculet, E.; Lungu, M.; Onisor, C.; Rebegea, L.; Vesa, D.; Bezman, L.; Bujoreanu, F.C.; Sarbu, M.I.; Mihailov, R.; et al. Nasopharyngeal Carcinoma: A New Synthesis of Literature Data (Review). Exp. Ther. Med. 2022, 23, 136. [Google Scholar] [CrossRef]
- Lee, A.W.M.; Ng, W.T.; Chan, L.L.K.; Hung, W.M.; Chan, C.C.C.; Sze, H.C.K.; Chan, O.S.H.; Chang, A.T.Y.; Yeung, R.M.W. Evolution of Treatment for Nasopharyngeal Cancer–Success and Setback in the Intensity-Modulated Radiotherapy Era. Radiother. Oncol. 2014, 110, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Vasef, M.A.; Ferlito, A.; Weiss, L.M. Nasopharyngeal Carcinoma, with Emphasis on Its Relationship to Epstein-Barr Virus. Ann. Otol. Rhinol. Laryngol. 1997, 106, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.C.W.; Hui, E.P.; Lo, K.-W.; Lam, W.K.J.; Johnson, D.; Li, L.; Tao, Q.; Chan, K.C.A.; To, K.-F.; King, A.D.; et al. Nasopharyngeal Carcinoma: An Evolving Paradigm. Nat. Rev. Clin. Oncol. 2021, 18, 679–695. [Google Scholar] [CrossRef]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal Analysis Reveals High Prevalence of Epstein-Barr Virus Associated with Multiple Sclerosis. Science 2022, 375, 296–301. [Google Scholar] [CrossRef]
- Pathmanathan, R.; Prasad, U.; Sadler, R.; Flynn, K.; Raab-Traub, N. Clonal Proliferations of Cells Infected with Epstein-Barr Virus in Preinvasive Lesions Related to Nasopharyngeal Carcinoma. N. Engl. J. Med. 1995, 333, 693–698. [Google Scholar] [CrossRef]
- Biggi, A.F.B.; de Oliveira, D.E. The Epstein-Barr Virus Hacks Immune Checkpoints: Evidence and Consequences for Lymphoproliferative Disorders and Cancers. Biomolecules 2022, 12, 397. [Google Scholar] [CrossRef]
- Van Senten, J.R.; Fan, T.S.; Siderius, M.; Smit, M.J. Viral G Protein-Coupled Receptors as Modulators of Cancer Hallmarks. Pharmacol. Res. 2020, 156, 104804. [Google Scholar] [CrossRef]
- Wang, L.; Ning, S. New Look of EBV LMP1 Signaling Landscape. Cancers 2021, 13, 5451. [Google Scholar] [CrossRef]
- Cen, O.; Longnecker, R. Epstein Barr Virus Volume 2, One Herpes Virus: Many Diseases. Curr. Top Microbiol. 2015, 391, 151–180. [Google Scholar] [CrossRef]
- Chen, J.; Fu, L.; Zhang, L.-Y.; Kwong, D.L.; Yan, L.; Guan, X.-Y. Tumor Suppressor Genes on Frequently Deleted Chromosome 3p in Nasopharyngeal Carcinoma. Chin. J. Cancer 2012, 31, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.T.; Ye, W.; Zeng, Y.-X.; Adami, H.-O. The Evolving Epidemiology of Nasopharyngeal Carcinoma. Cancer Epidemiol. Prev. Biomark. 2021, 30, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.M.; Yip, Y.L.; Lo, K.W.; Deng, W.; To, K.F.; Hau, P.M.; Lau, V.M.Y.; Takada, K.; Lui, V.W.Y.; Lung, M.L.; et al. Cyclin D1 Overexpression Supports Stable EBV Infection in Nasopharyngeal Epithelial Cells. Proc. Natl. Acad. Sci. USA 2012, 109, E3473–E3482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Zhang, W.; Ye, Q.; Zhou, Y.; Xiong, W.; He, W.; Deng, M.; Zhou, M.; Guo, X.; Chen, P.; et al. Inhibition of Epstein-Barr Virus Infection by Lactoferrin. J. Innate Immun. 2012, 4, 387–398. [Google Scholar] [CrossRef]
- Roberts, M.L.; Cooper, N.R. Activation of a Ras–MAPK-Dependent Pathway by Epstein–Barr Virus Latent Membrane Protein 1 Is Essential for Cellular Transformation. Virology 1998, 240, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Endo, K.; Shackelford, J.; Aga, M.; Yoshizaki, T.; Pagano, J.S. Upregulation of Special AT-Rich-Binding Protein 1 by Epstein–Barr Virus Latent Membrane Protein 1 in Human Nasopharyngeal Cells and Nasopharyngeal Cancer. J. Gen. Virol. 2013, 94, 507–513. [Google Scholar] [CrossRef]
- Lo, A.K.F.; Liu, Y.; Wang, X.H.; Huang, D.P.; Yuen, P.W.; Wong, Y.C.; Tsao, G.S.W. Alterations of Biologic Properties and Gene Expression in Nasopharyngeal Epithelial Cells by the Epstein-Barr Virus–Encoded Latent Membrane Protein 1. Lab. Investig. 2003, 83, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Deng, X.; Deng, L.; Gu, H.; Fan, W.; Cao, Y. Telomerase Activation by Epstein-Barr Virus Latent Membrane Protein 1 Is Associated with c-Myc Expression in Human Nasopharyngeal Epithelial Cells. J. Exp. Clin. Cancer Res. 2004, 23, 495–506. [Google Scholar]
- Yip, Y.; Tsang, C.; Deng, W.; Cheung, P.; Jin, Y.; Cheung, A.L.; Lung, M.L.; Tsao, S. Expression of Epstein-Barr Virus-encoded LMP1 and HTERT Extends the Life Span and Immortalizes Primary Cultures of Nasopharyngeal Epithelial Cells. J. Med. Virol. 2010, 82, 1711–1723. [Google Scholar] [CrossRef]
- Yang, J.; Deng, X.Y.; Gu, H.H.; Xia, L.Q.; Yi, W.; Cao, Y. Epstein-Barr Virus Latent Membrane Protein 1 Induces the Telomerase Activity in Human Nasopharyngeal Epithelial Cells. Shi Yan Sheng Wu Xue Bao 2001, 34, 207–211. [Google Scholar]
- Mordasini, V.; Ueda, S.; Aslandogmus, R.; Berger, C.; Gysin, C.; Hühn, D.; Sartori, A.A.; Bernasconi, M.; Nadal, D. Activation of ATR-Chk1 Pathway Facilitates EBV-Mediated Transformation of Primary Tonsillar B-Cells. Oncotarget 2016, 8, 6461–6474. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.; Lo, K.; Ko, C.; Young, L.S.; Dawson, C.W. Inhibition of the LKB1–AMPK Pathway by the Epstein–Barr Virus-encoded LMP1 Promotes Proliferation and Transformation of Human Nasopharyngeal Epithelial Cells. J. Pathol. 2013, 230, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Glatzel-Plucińska, N.; Piotrowska, A.; Dzięgiel, P.; Podhorska-Okołów, M. The Role of SATB1 in Tumour Progression and Metastasis. Int. J. Mol. Sci. 2019, 20, 4156. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, M.M.; Schulz, T.F. Regulation of Intracellular Signalling by the Terminal Membrane Proteins of Members of the Gammaherpesvirinae. J. Gen. Virol. 2006, 87, 1047–1074. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.-F.; Dawson, C.W.; Young, L.S.; Ko, C.-W.; Hau, P.-M.; Lo, K.-W. Activation of the FGFR1 Signalling Pathway by the Epstein-Barr Virus-Encoded LMP1 Promotes Aerobic Glycolysis and Transformation of Human Nasopharyngeal Epithelial Cells. J. Pathol. 2015, 237, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegtel, D.M.; Subramanian, A.; Sheen, T.-S.; Tsai, C.-H.; Golub, T.R.; Thorley-Lawson, D.A. Epstein-Barr-Virus-Encoded LMP2A Induces Primary Epithelial Cell Migration and Invasion: Possible Role in Nasopharyngeal Carcinoma Metastasis. J. Virol. 2005, 79, 15430–15442. [Google Scholar] [CrossRef] [Green Version]
- Port, R.J.; Pinheiro-Maia, S.; Hu, C.; Arrand, J.R.; Wei, W.; Young, L.S.; Dawson, C.W. Epstein–Barr Virus Induction of the Hedgehog Signalling Pathway Imposes a Stem Cell Phenotype on Human Epithelial Cells. J. Pathol. 2013, 231, 367–377. [Google Scholar] [CrossRef]
- Baizig, N.M.; Wided, B.A.; Amine, O.E.; Gritli, S.; ElMay, M. The Clinical Significance of IGF-1R and Relationship with Epstein–Barr Virus Markers: LMP1 and EBERs in Tunisian Patients with Nasopharyngeal Carcinoma. Ann. Otol. Rhinol. Laryngol. 2020, 129, 1011–1019. [Google Scholar] [CrossRef]
- Hu, J.; Li, Y.; Li, H.; Shi, F.; Xie, L.; Zhao, L.; Tang, M.; Luo, X.; Jia, W.; Fan, J.; et al. Targeting Epstein-Barr Virus Oncoprotein LMP1-Mediated High Oxidative Stress Suppresses EBV Lytic Reactivation and Sensitizes Tumors to Radiation Therapy. Theranostics 2020, 10, 11921–11937. [Google Scholar] [CrossRef]
- Li, H.M.; Zhuang, Z.H.; Wang, Q.; Pang, J.C.S.; Wang, X.H.; Wong, H.L.; Feng, H.C.; Jin, D.Y.; Ling, M.T.; Wong, Y.C.; et al. Epstein–Barr Virus Latent Membrane Protein 1 (LMP1) Upregulates Id1 Expression in Nasopharyngeal Epithelial Cells. Oncogene 2004, 23, 4488–4494. [Google Scholar] [CrossRef] [Green Version]
- Shi, F.; Zhou, M.; Shang, L.; Du, Q.; Li, Y.; Xie, L.; Liu, X.; Tang, M.; Luo, X.; Fan, J.; et al. EBV(LMP1)-Induced Metabolic Reprogramming Inhibits Necroptosis through the Hypermethylation of the RIP3 Promoter. Theranostics 2019, 9, 2424–2438. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.F.; Lo, K.W.; Tsao, S.W.; Wong, H.L.; Hui, J.W.Y.; To, K.F.; Hayward, S.D.; Chui, Y.L.; Lau, Y.L.; Takada, K.; et al. Epstein-Barr Virus Infection Alters Cellular Signal Cascades in Human Nasopharyngeal Epithelial Cells. Neoplasia 2006, 8, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Hu, C.; Zhu, Y.; Sun, R.; Fang, Y.; Fan, Y.; Xu, F. LMP1 Increases Expression of NADPH Oxidase (NOX) and Its Regulatory Subunit P22 in NP69 Nasopharyngeal Cells and Makes Them Sensitive to a Treatment by a NOX Inhibitor. PLoS ONE 2015, 10, e0134896. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.-W.; Chung, G.T.-Y.; To, K.-F. Deciphering the Molecular Genetic Basis of NPC through Molecular, Cytogenetic, and Epigenetic Approaches. Semin. Cancer Biol. 2012, 22, 79–86. [Google Scholar] [CrossRef]
- Iwakiri, D. Multifunctional Non-Coding Epstein–Barr Virus Encoded RNAs (EBERs) Contribute to Viral Pathogenesis. Virus Res. 2016, 212, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.K.; Tsai, K.; Allan, L.L.; Zheng, D.J.; Nie, J.C.; Biggs, C.M.; Hasan, M.R.; Kozak, F.K.; van den Elzen, P.; Priatel, J.J.; et al. Innate Immune Control of EBV-Infected B Cells by Invariant Natural Killer T Cells. Blood 2013, 122, 2600–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chang, E.T.; Liu, Q.; Cai, Y.; Zhang, Z.; Chen, G.; Huang, Q.; Xie, S.; Cao, S.; Jia, W.; et al. Occupational Exposures and Risk of Nasopharyngeal Carcinoma in a High-risk Area: A Population-based Case-control Study. Cancer 2021, 127, 2724–2735. [Google Scholar] [CrossRef]
- Wang, L.; Xu, D.; Huang, Q.; Yang, G.; Zhang, M.; Bi, J.; Shan, J.; Li, E.; He, S. Characterization of Tonsil Microbiota and Their Effect on Adenovirus Reactivation in Tonsillectomy Samples. Microbiol. Spectr. 2021, 9, e01246-21. [Google Scholar] [CrossRef]
- Zeidler, R.; Meissner, P.; Eissner, G.; Lazis, S.; Hammerschmidt, W. Rapid Proliferation of B Cells from Adenoids in Response to Epstein-Barr Virus Infection. Cancer Res. 1996, 56, 5610–5614. [Google Scholar]
- Jiang, R.; Ekshyyan, O.; Moore-Medlin, T.; Rong, X.; Nathan, S.; Gu, X.; Abreo, F.; Rosenthal, E.L.; Shi, M.; Guidry, J.T.; et al. Association between Human Papilloma Virus/Epstein-Barr Virus Coinfection and Oral Carcinogenesis. J. Oral Pathol. Med. 2014, 44, 28–36. [Google Scholar] [CrossRef]
- Lewis, J.S. Human Papillomavirus Testing in Head and Neck Squamous Cell Carcinoma in 2020: Where Are We Now and Where Are We Going? Head Neck Pathol. 2020, 14, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Phua, S.K.A.; Soong, Y.L.; Oon, L.L.E.; Chan, K.S.; Lucky, S.S.; Mong, J.; Tan, M.H.; Lim, C.M. Clinical Utility of Epstein-Barr Virus DNA and Other Liquid Biopsy Markers in Nasopharyngeal Carcinoma. Cancer Commun. 2020, 40, 564–585. [Google Scholar] [CrossRef] [PubMed]
- Mirghani, H.; Kuhs, K.A.L.; Waterboer, T. Biomarkers for Early Identification of Recurrences in HPV-Driven Oropharyngeal Cancer. Oral Oncol. 2018, 82, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Jud, A.; Kotur, M.; Berger, C.; Gysin, C.; Nadal, D.; Lünemann, A. Tonsillar CD56brightNKG2A+ NK Cells Restrict Primary Epstein-Barr Virus Infection in B Cells via IFN-γ. Oncotarget 2016, 8, 6130–6141. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKeon, M.G.; Gallant, J.-N.; Kim, Y.J.; Das, S.R. It Takes Two to Tango: A Review of Oncogenic Virus and Host Microbiome Associated Inflammation in Head and Neck Cancer. Cancers 2022, 14, 3120. https://doi.org/10.3390/cancers14133120
McKeon MG, Gallant J-N, Kim YJ, Das SR. It Takes Two to Tango: A Review of Oncogenic Virus and Host Microbiome Associated Inflammation in Head and Neck Cancer. Cancers. 2022; 14(13):3120. https://doi.org/10.3390/cancers14133120
Chicago/Turabian StyleMcKeon, Mallory G., Jean-Nicolas Gallant, Young J. Kim, and Suman R. Das. 2022. "It Takes Two to Tango: A Review of Oncogenic Virus and Host Microbiome Associated Inflammation in Head and Neck Cancer" Cancers 14, no. 13: 3120. https://doi.org/10.3390/cancers14133120