
Unraveling the Role of Guanylate-Binding Proteins (GBPs) in Breast Cancer: A Comprehensive Literature Review and New Data on Prognosis in Breast Cancer Subtypes



Abstract
:Simple Summary
Abstract
1. Introduction
Interferons and Breast Cancer
2. Materials and Methods
Gene Expression Profiling and Data Processing
3. Results
3.1. GBP-1
3.2. GBP-5
3.3. GBP-2
4. Discussion
4.1. GBP-1
4.2. GBP-5
4.3. GBP-2
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esquivel-Velázquez, M.; Ostoa-Saloma, P.; Palacios-Arreola, M.I.; Nava-Castro, K.E.; Castro, J.I.; Morales-Montor, J. The role of cytokines in breast cancer development and progression. J. Interferon Cytokine Res. 2015, 35, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromberg, J. Signal transducers and activators of transcription as regulators of growth, apoptosis and breast development. Breast Cancer Res. 2000, 2, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, D.H.; Shankaran, V.; Dighe, A.S.; Stockert, E.; Aguet, M.; Old, L.J.; Schreiber, R.D. Demonstration of an interferon gamma-dependent tumor surveillance system in immunocompetent mice. Proc. Natl. Acad. Sci. USA 1998, 95, 7556–7561. [Google Scholar] [CrossRef] [Green Version]
- Cheon, H.; Borden, E.C.; Stark, G.R. Interferons and their stimulated genes in the tumor microenvironment. Semin Oncol. 2014, 41, 156–173. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, S.; Takayama, Y.; Ogawa, A.; Tamura, K.; Okada, M. Transmembrane phosphoprotein Cbp positively regulates the activity of the Carboxyl-terminal Src kinase, Csk. J. Biol. Chem. 2000, 275, 29183–29186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.R.; Vermi, W.; Luo, J.; Lucini, L.; Rickert, C.; Fowler, A.M.; Lonardi, S.; Arthur, C.; Young, L.J.T.; Levy, D.E.; et al. STAT1-deficient mice spontaneously develop estrogen receptor a-positive luminal mammary carcinomas. Breast Cancer Res. 2012, 14, R16. [Google Scholar] [CrossRef] [Green Version]
- Varikuti, S.; Oghumu, S.; Elbaz, M.; Volpedo, G.; Ahirwar, D.K.; Alarcon, P.C.; Sperling, R.H.; Moretti, E.; Pioso, M.S.; Kimble, J.; et al. STAT1 gene deficient mice develop accelerated breast cancer growth and metastasis which is reduced by IL-17 blockade. Oncoimmunology 2017, 6, e1361088. [Google Scholar] [CrossRef] [Green Version]
- Guenzi, E.; Topolt, K.; Cornali, E.; Lubeseder-Martellato, C.; Jorg, A.; Matzen, K.; Zietz, C.; Kremmer, E.; Nappi, F.; Schwemmle, M.; et al. The helical domain of GBP-1 mediates the inhibition of endothelial cell proliferation by inflammatory cytokines. EMBO J. 2001, 20, 5568–5577. [Google Scholar] [CrossRef] [Green Version]
- Guenzi, E.; Topolt, K.; Lubeseder-Martellato, C.; Jorg, A.; Naschberger, E.; Benelli, R.; Albini, A.; Sturzl, M. The guanylate binding protein-1 GTPase controls the invasive and angiogenic capabiliy of endothelial cells through inhibition of MMP-1 expression. EMBO J. 2003, 22, 3772–3782. [Google Scholar] [CrossRef]
- Harvat, B.L.; Jetten, A.M. γ-interferon induces an irreversible growth arrest in mid-G1 in mammary epithelial cells which correlates with a block in hyperphosphorylation of retinoblastoma. Cell. Growth Diff 1996, 7, 289–300. [Google Scholar] [PubMed]
- Barrat, F.J.; Crow, M.K.; Ivashkiv, L.B. Interferon target-gene expression and epigenomic signatures in health and disease. Nat. Immunol. 2019, 20, 1574–1583. [Google Scholar] [CrossRef]
- Ascierto, M.L.; Idowu, M.O.; Zhao, Y.; Khalak, H.; Payne, K.K.; Wang, X.-Y.; Dumur, C.; Bedognetti, D.; Tomei, S.; Ascierto, P.A.; et al. Molecular signatures mostly associated with NK cells are predictive of relapse free survival in breast cancer patients. J. Transl. Med. 2013, 11, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascierto, M.L.; Kmieciak, M.; Idowu, M.O.; Manjii, R.; Zhao, Y.; Grimes, M.; Dumur, C.; Wang, E.; Ramakrishnan, V.; Wang, X.-Y.; et al. A signature of immune function genes associated with recurrence-free survival in breast cancer patients. Breast Cancer Res Treat. 2012, 131, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Lecchi, M.; Verderio, P.; Cappelletti, V.; De Santis, F.; Paolini, B.; Monica, M.; Sangaletti, S.; Pupa, S.M.; Iorio, M.V.; Bianchi, G.; et al. A combination of extracellular matrix- and interferon-associated signatures identifies high-grade breast cancers with poor prognosis. Mol. Oncol. 2021, 15, 1345–1357. [Google Scholar] [CrossRef]
- Burstein, M.D.; Tsimelzon, A.; Poage, G.M.; Covington, K.R.; Contreras, A.; Fuqua, S.A.W.; Savage, M.I.; Osborne, C.K.; Hilsenbeck, S.G.; Chang, J.C.; et al. Comprehensive genomic analysis identifies novel subtypes and targets of triple-negative breast cancer. Clin. Cancer Res. 2015, 21, 1688–1698. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Baggerly, K.A.; Wang, Y.; Zhang, Y.; Gonzalez-Angulo, A.M.; Meric-Bernstam, F.; Valero, V.; Lehmann, B.D.; Pietenpol, J.A.; Hortobogyi, G.N.; et al. Differential response to neoadjuvant chemotherapy among 7 triple-negative breast cancer molecular subtypes. Clin. Cancer Res. 2013, 19, 5533–5540. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, U.; Guethlein, L.; Klamp, T.; Ozbek, K.; Schaub, A.; Futterer, A.; Pleffer, K.; Howard, J.C. Two families of GTPases dominate the complex cellular response to IFN-γ. J. Immunol. 1998, 161, 6715–6723. [Google Scholar]
- Quintero, M.; Adamoski, D.; Menezes dos Reis, L.; Ascencao, C.F.R.; Sousa de Oliveira, K.R.; Almaide Goncalves, K.; Dias, M.M.; Carazzolle, M.F.; Dias, S.M.G. Guanylate-binding protein-1 is a potential new therapeutic target for triple-negative breast cancer. BMC Cancer 2017, 17, 727. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, M.H.; Hood, B.L.; Beck, H.C.; Conrads, T.P.; Ditzel, H.J.; Leth-Larsen, R. Downregulation of antigen presentation-associated pathway proteins is linked to poor outcome in triple-negative breast cancer patient tumors. Oncoimmunology 2017, 6, e1305531. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.; Cadenas, C.; Hellwig, B.; Marchan, R.; Stewart, J.; Reif, R.; Lohr, M.; Gehrmann, M.; Rahnenfuhrer, J.; Schmidt, M.; et al. Interferon-inducible guanylate binding protein (GBP2) is associated with better prognosis in breast cancer and indicates an efficient T cell response. Breast Cancer 2014, 21, 491–499. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Wu, W.; Wang, F.; Liu, X.; Shui, G.; Nie, C. Guanylate-binding protein 2 regulates Drp1-mediated mitochondrial fission to suppress breast cancer cell invasion. Cell. Death Dis. 2017, 8, e3151. [Google Scholar] [CrossRef] [PubMed]
- Nyabuto, G.O.; Wilson, J.P.; Heilman, S.A.; Kalb, R.C.; Kopacz, J.P.; Abnave, A.V.; Vestal, D.J. The Large GTPase, GBP-2, Regulates Rho Family GTPases to Inhibit Migration and Invadosome Formation in Breast Cancer Cells. Cancers 2021, 13, 5632. [Google Scholar] [CrossRef]
- Rahvar, F.; Salimi, M.; Mozdarani, H. Plasma GBP2 promoter methylation is associated with advanced stages in breast cancer. Genet. Mol. Biol. 2020, 43, e20190230. [Google Scholar] [CrossRef]
- Cheng, S.W.; Chen, P.C.; Ger, T.R.; Chiu, H.W.; Lin, Y.F. GBP5 Serves as a Potential Marker to Predict a Favorable Response in Triple-Negative Breast Cancer Patients Receiving a Taxane-Based Chemotherapy. J. Pers. Med. 2021, 11, 197. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.W.; Chen, P.C.; Lin, M.H.; Ger, T.R.; Chiu, H.W.; Lin, Y.F. GBP5 Repression Suppresses the Metastatic Potential and PD-L1 Expression in Triple-Negative Breast Cancer. Biomedicines 2021, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Cimas, F.J.; Manzano, A.; Baliu-Piqué, M.; García-Gil, E.; Pérez-Segura, P.; Nagy, Á.; Pandiella, A.; Győrffy, B.; Ocana, A. Genomic Mapping Identifies Mutations in RYR2 and AHNAK as Associated with Favorable Outcome in Basal-Like Breast Tumors Expressing PD1/PD-L1. Cancers 2020, 12, 2243. [Google Scholar] [CrossRef]
- Hachim, M.Y.; Hachim, I.Y.; Talaat, I.M.; Yakout, N.M.; Hamoudi, R. M1 Polarization Markers Are Upregulated in Basal-Like Breast Cancer Molecular Subtype and Associated With Favorable Patient Outcome. Front. Immunol. 2020, 11, 560074. [Google Scholar] [CrossRef] [PubMed]
- Lipnik, K.; Naschberger, E.; Gonin-Laurent, N.; Kodajova, P.; Petznek, H.; Rungaldier, S.; Astigiano, S.; Ferrini, S.; Sturzl, M.; Hohenadl, C. Interferon gamma-induced human guanylate binding protein 1 inhibits mammary tumor growth in mice. Mol. Med. 2010, 16, 177–187. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Beeman, N.; Hilgarth, R.S.; Nava, P.; Louis, N.A.; Naschberger, E.; Sturzl, M.; Parkos, C.A.; Nusrat, A. IFN-gamma and TNF-alpha-induced GBP-1 inhibits epithelial cell proliferation through suppression of beta-catenin/TCF signaling. Mucosal Immunol 2012, 5, 681–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unterer, B.; Wiesmann, V.; Gunasekaran, M.; Sticht, H.; Tenkerian, C.; Behrens, J.; Leone, M.; Engel, F.B.; Britzen-Laurent, N.; Naschberger, E.; et al. IFN-gamma-response mediator GBP-1 represses human cell proliferation by inhibiting the Hippo signaling transcription factor TEAD. Biochem. J. 2018, 475, 2955–2967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forster, F.; Paster, W.; Supper, V.; Schatzlmaier, P.; Sunzenauer, S.; Ostler, N.; Saliba, A.; Eckerstorfer, P.; Britzen-Laurent, N.; Schutz, G.; et al. Guanylate binding protein 1-mediated interaction of T cell antigen receptor signaling with the cytoskeleton. J. Immunol 2014, 192, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Ostler, N.; Britzen-Laurent, N.; Leible, A.; Naschberger, E.; Lochnit, G.; Ostler, M.; Forster, F.; Kunzelmann, P.; Ince, S.; Supper, V.; et al. Gamma interferon-induced Guanylate-Binding Protein 1 is a novel actin cytoskeleton remodeling factor. Mol. Cell. Biol. 2014, 34, 196–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinlander, K.; Naschberger, E.; Lehmann, M.H.; Tripal, P.; Paster, W.; Stockinger, H.; Hohenadl, C.; Sturzl, M. Guanylate binding protein-1 inhibits spreading and migration of endothelial cells through induction of integrin α4 expression. FASEB J. 2008, 22, 4168–4178. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, J.; Li, L.; Zhang, H.; Zhang, H.; Li, J.; Zhong, H.; Lei, T.; Jin, Y.; Xu, B.; et al. Guanylate-Binding Protein 1 as a Potential Predictor of Immunotherapy: A Pan-Cancer Analysis. Front. Genet. 2022, 13, 820135. [Google Scholar] [CrossRef]
- Fukumoto, M.; Amanuma, T.; Kuwahara, Y.; Shimura, T.; Suzuki, M.; Mori, S.; Kumamoto, H.; Saito, Y.; Ohkubo, Y.; Duan, Z.; et al. Guanine nucleotide-binding protein 1 is one of the key molecules contributing to cancer cell radioresistance. Cancer Sci. 2014, 105, 1351–1359. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, D.A.M.; Pedrosa, R.; Smid, M.; van der Weiden, M.; de Weerd, V.; Nigg, A.L.; Berrevoets, C.; Zeneyedpour, L.; Priego, N.; Valiente, M.; et al. T lymphocytes facilitate brain metastasis of breast cancer by inducing Guanylate-Binding Protein 1 expression. Acta Neuropathol. 2018, 135, 581–599. [Google Scholar] [CrossRef] [Green Version]
- Fekete, J.T.; Győrffy, B. ROCplot.org: Validating predictive biomarkers of chemotherapy/hormonal therapy/anti-HER2 therapy using transcriptomic data of 3,104 breast cancer patients. Int. J. Cancer 2019, 145, 3140–3151. [Google Scholar] [CrossRef]
- Messmer-Blust, A.F.; Balasubramanian, S.; Gorbacheva, V.Y.; Jeyaratnam, J.A.; Vestal, D.J. The interferon-γ-induced murine Guanylate-Binding Protein-2 (mGBP-2) inhibits Rac activation during cell spreading on fibronectin and after platelet-derived growth factor (PDGF) treatment: Role for Phosphatidylinositol 3-Kinase. Mol. Biol. Cell. 2010, 15, 2514–2528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, S.; Fan, M.; Messmer-Blust, A.F.; Yang, C.H.; Trendel, J.A.; Jeyaratnam, J.A.; Pfeffer, L.M.; Vestal, D.J. The interferon-γ-induced GTPase, mGBP-2, inhibits tumor necrosis factor α (TNF-α) induction of matrix metalloproteinase-9 (MMP-9) by inhibiting NF-κB and Rac protein. J. Biol. Chem. 2011, 286, 20054–20064. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, D.P.; Oliveira, I.M.; de Moraes, E.; Paiva, G.R.; Souza, D.M.; Barnas, C.; Olmedo, D.B.; Pinto, C.E.; Faria, P.A.; De Moura Gallo, C.V.; et al. Interferon-inducible guanylate binding protein (GBP)-2: A novel p53-regulated tumor marker in esophageal squamous cell carcinomas. Int. J. Cancer 2009, 124, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yu, X.; Sun, L.; Zheng, Y.; Chen, L.; Xu, H.; Jin, J.; Lan, Q.; Chen, C.C.; Li, M. GBP2 enhances glioblastoma invasion through Stat3/fibronectin pathway. Oncogene 2020, 39, 5042–5055. [Google Scholar] [CrossRef] [PubMed]
- Aslakson, C.J.; Miller, F.R. Selective events in the metastatic process defined by analysis of the sequential dissemination of subpopulations of a mouse mammary tumor. Cancer Res. 1992, 52, 1399–1405. [Google Scholar] [PubMed]
- Zhao, J.; Zhang, J.; Yu, M.; Xie, Y.; Huang, Y.; Wolff, D.W.; Abel, P.W.; Tu, Y. Mitochondrial dynamics regulates migration and invasion of breast cancer cells. Oncogene 2013, 32, 4814–4824. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22, 277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Gyorffy, B.; Lanczky, A.; Szallasi, Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data of 1287 patients. Endocr. -Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Jin, J.; Zheng, Y.; Zhu, H.; Xu, H.; Ma, J.; Lan, Q.; Zhuang, Z.; Chen, C.C.; Li, M. GBP5 drives malignancy of glioblastoma via the Src/ERK1/2/MMP3 pathway. Cell. Death Dis. 2021, 12, 203. [Google Scholar] [CrossRef]
- Ji, G.; Luo, B.; Chen, L.; Shen, G.; Tian, T. GBP2 Is a Favorable Prognostic Marker of Skin Cutaneous Melanoma and Affects Its Progression via the Wnt/β-catenin Pathway. Ann. Clin. Lab Sci. 2021, 51, 772–782. [Google Scholar]
- Honkala, A.T.; Tailor, D.; Malhotra, S.V. Guanylate-Binding Protein 1: An Emerging Target in Inflammation and Cancer. Front. Immunol 2019, 10, 3139. [Google Scholar] [CrossRef]
- Bai, S.; Chen, T.; Deng, X. Guanylate-Binding Protein 1 Promotes Migration and Invasion of Human Periodontal Ligament Stem Cells. Stem Cells Int. 2018, 2018, 6082956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Zhu, H.; Dai, X.; Xi, Y.; Sheng, Y.; Gao, C.; Liu, H.; Xue, Y.; Liu, J.; Shi, J.; et al. Overexpression of GBP1 predicts poor prognosis and promotes tumor growth in human glioblastoma multiforme. Cancer Biomark 2019, 25, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Wadi, S.; Tipton, A.R.; Trendel, J.A.; Khuder, S.A.; Vestal, D.J. hGBP-1 Expression Predicts Shorter Progression-Free Survival in Ovarian Cancers, While Contributing to Paclitaxel Resistance. J. Cancer Ther. 2016, 7, 994–1007. [Google Scholar] [CrossRef] [Green Version]
- Tipton, A.R.; Nyabuto, G.O.; Trendel, J.A.; Mazur, T.M.; Wilson, J.P.; Wadi, S.; Justinger, J.S.; Moore, G.L.; Nguyen, P.T.; Vestal, D.J. Guanylate-Binding Protein-1 protects ovarian cancer cell lines but not breast cancer cell lines from killing by paclitaxel. Biochem. Biophys. Res. Commun 2016, 478, 1617–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Q.; Wang, A.; Cheng, Y.; Mukasa, A.; Ma, J.; Hong, L.; Yu, S.; Sun, L.; Huang, Q.; Purow, B.; et al. Guanylate binding protein-1 mediates EGFRvIII and promotes glioblastoma growth in vivo but not in vitro. Oncotarget 2016, 7, 9680–9691. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ma, G.; Jing, C.; Liu, Z. Guanylate-binding protein 1 (GBP1) promotes lymph node metastasis in human esophageal squamous cell carcinoma. Discov. Med. 2015, 20, 369–378. [Google Scholar]
- Britzen-Laurent, N.; Lipnik, K.; Ocker, M.; Naschberger, E.; Schellerer, V.S.; Croner, R.S.; Vieth, M.; Waldner, M.; Steinberg, P.; Hohenadl, C.; et al. GBP-1 acts as a tumor suppressor in colorectal cancer cells. Carcinogenesis 2013, 34, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, H.; Del Rosario, A.M.; Bryson, B.D.; Schroeder, M.A.; Sarkaria, J.N.; White, F.M. Molecular characterization of EGFR and EGFRvIII signaling networks in human glioblastoma tumor xenografts. Mol. Cell. Proteom. MCP 2012, 11, 1724–1740. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Mukasa, A.; Inda, M.M.; Zhang, J.; Chin, L.; Cavenee, W.; Furnari, F. Guanylate binding protein 1 is a novel effector of EGFR-driven invasion in glioblastoma. J. Exp. Med. 2011, 208, 2657–2673. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.J.; Chang, K.P.; Chang, Y.J.; Hsu, C.W.; Liang, Y.; Yu, J.S.; Chi, L.M.; Chang, Y.S.; Wu, C.C. Identification of guanylate-binding protein 1 as a potential oral cancer marker involved in cell invasion using omics-based analysis. J. Proteome Res. 2011, 10, 3778–3788. [Google Scholar] [CrossRef]
- De Donato, M.; Mariani, M.; Petrella, L.; Martinelli, E.; Zanninum, G.F.; Vellone, V.; Ferrandina, G.; Shahabi, S.; Scambia, G.; Ferlini, C. Class III b-tubulin and the cytoskeletal gateway for drug resistance in ovarian cancer. J. Cell. Physiol. 2011, 227, 1034–1041. [Google Scholar] [CrossRef]
- Duan, Z.; Foster, R.; Brakora, K.A.; Yusuf, R.Z.; Seiden, M.V. GBP1 overexpression is associated with a paclitaxel resistance phenotype. Cancer Chemother Pharm. 2005, 57, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Lamendola, D.E.; Duan, Y.; Yusaf, R.; Seiden, M.V. Description of paclitaxel resistance-associated genes in ovarian and breast cancer cell lines. Cancer Chemother Pharm. 2005, 55, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Carbotti, G.; Petretto, A.; Naschberger, E.; Stürzl, M.; Martini, S.; Mingari, M.C.; Filaci, G.; Ferrini, S.; Fabbi, M. Cytokine-Induced Guanylate Binding Protein 1 (GBP1) Release from Human Ovarian Cancer Cells. Cancers 2020, 12, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Gou, L.; Wei, T.; Zhang, J. GBP1 promotes erlotinib resistance via PGK1-activated EMT signaling in non-small cell lung cancer. Int. J. Oncol. 2020, 57, 858–870. [Google Scholar] [CrossRef]
- Luo, Y.; Jin, H.; Kim, J.H.; Bae, J. Guanylate-binding proteins induce apoptosis of leukemia cells by regulating MCL-1 and BAK. Oncogenesis 2021, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Qu, J.; Li, L.; Gao, F. Guanylate-binding protein 1 correlates with advanced tumor features, and serves as a prognostic biomarker for worse survival in lung adenocarcinoma patients. J. Clin. Lab Anal. 2021, 35, e23610. [Google Scholar] [CrossRef]
- Yamakita, I.; Mimae, T.; Tsutani, Y.; Miyata, Y.; Ito, A.; Okada, M. Guanylate binding protein 1 (GBP-1) promotes cell motility and invasiveness of lung adenocarcinoma. Biochem. Biophys. Res. Commun. 2019, 518, 266–272. [Google Scholar] [CrossRef]
- Zhao, J.; Li, X.; Liu, L.; Cao, J.; Goscinski, M.A.; Fan, H.; Li, H.; Suo, Z. Oncogenic Role of Guanylate Binding Protein 1 in Human Prostate Cancer. Front. Oncol. 2019, 9, 1494. [Google Scholar] [CrossRef]
- Grenz, S.; Naschberger, E.; Merkel, S.; Britzen-Laurent, N.; Schaal, U.; Konrad, A.; Aigner, M.; Rau, T.T.; Hartmann, A.; Croner, R.S.; et al. IFN-gamma-driven intratumoral microenvironment exhibits superior prognostic effect compared with an IFN-alpha-driven microenvironment in patients with colon carcinoma. Am. J. Pathol. 2013, 183, 1897–1909. [Google Scholar] [CrossRef]
- Kominsky, S.; Johnson, H.M.; Bryan, G.; Tanabe, T.; Hobeika, A.C.; Subramaniam, P.S.; Torres, B. IFNg inhibition of cell growth in glioblastomas correlates with increased levels of the cyclin dependent kinase inhibitor p21WAF1/CIP1. Oncogene 1998, 17, 2973–2979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubeseder-Martellato, C.; Guenzi, E.; Jorg, A.; Topolt, D.; Naschberger, E.; Kremmer, E.; Zietz, C.; Tschachler, E.; Hutzer, P.; Schwemmle, M.; et al. Guanylate-binding protein-1 expression is selectively induced by inflammatory cytokines and is an activation marker of endothelial cells during inflammatory diseases. Am. J. Pathol. 2002, 161, 1749–1759. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, A.R.; Wellington, D.A.; Kumar, P.; Kassa, H.; Booth, C.J.; Cresswell, P.; MacMicking, J.D. GBP5 promotes NLRP3 inflammasome assembly and immunity in mammals. Science 2012, 336, 481–485. [Google Scholar] [CrossRef]
- Fellenberg, F.; Hartmann, T.B.; Dummer, R.; Usener, D.; Schadendorf, D.; Eichmüller, S. GBP-5 splicing variants: New guanylate-binding proteins with tumor-associated expression and antigenicity. J. Investig. Derm. 2004, 122, 1510–1517. [Google Scholar] [CrossRef] [Green Version]
- Wehner, M.; Herrmann, C. Biochemical properties of the human guanylate binding protein 5 and a tumor-specific truncated splice variant. FEBS J. 2010, 277, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.; Praefcke, G.J.K.; Renault, L.; Wittinghofer, A.; Herrmann, C. Structure of human guanylate-binding protein 1 representing a unique class of GTP-binding proteins. Nature 2000, 403, 567–571. [Google Scholar] [CrossRef]
- Prakash, B.; Renault, L.; Praefcke, G.J.K.; Herrmann, C.; Wittinghofer, A. Triphosphate structure of guanylate-binding protein 1 and implications for nucleotide binding and GTPase mechanism. EMBO J. 2000, 19, 4555–4564. [Google Scholar] [CrossRef] [Green Version]
- Praefcke, G.J.K.; McMahon, H.T. The dynamin superfamily: Universal membrane tubulation and fission molecules. Nat. Rev. Mol. Cell. Biol. 2004, 5, 133–147. [Google Scholar] [CrossRef]
- Britzen-Laurent, N.; Bauer, M.; Berton, V.; Fischer, N.; Syguda, A.; Reipschläger, S.; Naschberger, E.; Herrmann, C.; Stürzl, M. Intracellular trafficking of guanylate-binding proteins is regulated by heterodimerization in a hierarchical manner. PLoS ONE 2010, 5, e14246. [Google Scholar] [CrossRef] [Green Version]
- Degrandi, D.; Konermann, C.; Beuter-Gunia, C.; Kresse, A.; Wurthner, J.; Kurig, S.; Beer, S.; Pfeffer, K. Extensive characterization of IFN-induced GTPases mGTP1 to mGBP10 involved in host defense. J. Immunol 2007, 179, 7729–7740. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Min, H.; Hu, B.; Xue, X.; Liu, Y. Guanylate-binding protein-2 inhibits colorectal cancer cell growth and increases the sensitivity to paclitaxel of paclitaxel-resistant colorectal cancer cells by interfering Wnt signaling. J. Cell. Biochem. 2020, 121, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yang, B.; Guo, G.; Zhang, J.; Sun, Y.; Liu, D.; Guo, S.; Wu, Y.; Wang, X.; Wang, S.; et al. GBP2 facilitates the progression of glioma via regulation of KIF22/EGFR signaling. Cell. Death Discov. 2022, 8, 208. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Clinical Samples | Results | Reference |
|---|---|---|---|
| - | Breast cancer samples | GBP-1 and STAT1 are part of a 5 gene signature that correlates with improved RFS in all breast cancers. GBP-1 expression is highest in infiltrating cells but was also expressed in the tumor cells of recurrence-free patients. | Ascierto, 2013 [13] |
| - | TNBC tumor samples | Subtyped TNBCs into 4 subgroups. Two subgroups were of basal histology. Basal-like Immune Activated (BLIA) tumors had elevated expression for genes for T-cell, B-cell, and NK activation. Further, IFN-γ stimulated genes. GBP-5 was the 3rd–5th most robustly induced gene in BLIA tumors, and GBP-1 was the 11th. BLIA tumors are predicted to have greater than 85% RFS over 10 years, much better than other forms of TNBC. | Burstein, 2015 [16] |
| TS/A | - | Forced expression of GBP-1 in TS/A cells resulted in smaller tumors in immune competent mice. Not accompanied by decrease in infiltrating cells. Reduced Ki67. Reduced level of VEGF-A both in vitro and in vivo. | Lipnik, 2010 [30] |
| MCF-7, SKBR3, MDA-MB-468, MDA-MB-436, MDA-MB-231, BT549, T47D | Breast cancer samples | Upregulated genes in TNBCs were analyzed for druggability using ChRMBL Studel. GBP-1 was ranked second in the upregulated genes based on druggability. KD of GBP-1 inhibited proliferation in a subgroup of TNBC cell lines. EGFR can drive GBP-1 expression in breast cancer. | Quintero, 2017 [20] |
| SAS, HepG2, KB, MM102 cells | - | Cells made clinically cells radioresistant (CRR). All CRR cells expressed elevated GBP compared to parental cells. KD of GBP reduced radioresistance. | Fukumoto, 2014 [37] |
| MDA-MB-231, MDA-MB-231-BM, SUM159PT | Human primary T-cells | Co-culturing activated T-cells with breast cancer cells increased their crossing of artificial blood brain barrier (BBB). GBP-1 was upregulated in the tumor cells after incubation with activated T-cells. KD of GBP-1 in tumor cells reduced crossing of the BBB after incubation with activated T-cells. | Mustafa, 2018 [38] |
| Jurkat cells | Primary human T-cells | Silencing of GBP-1 increases T-cell spreading and surface expression of TCR/CD3 and CD45. Modulates early TCR signaling. | Forster, 2014 [33] |
| Cell Lines | Clinical Samples | Results | Reference |
|---|---|---|---|
| - | TNBCs | GBP-5 is 5th most robustly induced gene in BLIA TNBC tumors with gene signatures of IFN-γ, B-cell, T-cell, and NK cell activation. Significantly improved survival compared to other TNBCs, particularly other basal TNBCs. | Burstein, 2015 [16] |
| MDA-MB-231, Hs578T | TNBCs | High expression of GBP-5 correlated with improved RFS and PRS in TNBCs. GBP-5 not expressed in normal breast epithelial cells but expressed in 5/7 TNBC cell lines. Contributes to paclitaxel sensitivity. Suggest GBP-5 promotes TNBC protection by activating Akt/mTOR and inhibiting autophagy. | Cheng, 2021 [26] |
| MDA-MB-231, Hs578T | TNBCs | GBP-5 expression correlates with poor prognosis in TNBCs. KD of GBP-5 inhibited cell migration and activity from both GAS and NF-kB promoter elements. | Cheng, 2021 [27] |
| - | Breast Cancer Samples | Mutations in tumors with high PD1 and PD-L1 were associated with GBP-5 expression and good prognosis. Also associated with immune infiltration of the tumors. | Cimas, 2020 [28] |
| Cell Lines | Clinical Samples | Results | Reference |
|---|---|---|---|
| 766 node negative breast cancers | - | Elevated GBP-2 correlated with longer time to distant metastasis in highly proliferating ER+ tumors with infiltrating T-cells (as judged by gene signature). | Godoy, 2014 [22] |
| MDA-MB-231 and MDA-MB-436 | - | GBP-2 interacts with Drp1 to both inhibit mitochondrial fission and cell migration. | Zhang, 2017 [23] |
| 4T1, 67NR murine breast cancer cells | - | GBP-2 inhibits cell migration by inhibiting Rac1 activation and activating CDC42 and RhoA. Inhibits invadosome formation. | Nyabuto, 2021 [24] |
| NIH 3T3 fibroblasts, B16 melanoma | - | GBP-2 inhibits cell spreading downstream of integrins, PDGF, and TNF-α treatment. Inhibits activation of Rac1 and PI3-K when cells plated on fibronectin. | Messmer-Blust, 2010 [40] |
| NIH3T3 cells | - | GBP-2 inhibits TNF-α induction of matrix metalloprotease-9 (MMP-9) by inhibiting the binding of NF-κB p65 to the MMP-9 promoter. | Balasubramanian, 2011 [41] |
| TE-1 squamous cell carcinoma cells | - | GBP-2 is a p53 responsive gene. | Guimaraes, 2009 [42] |
| - | Breast cancer and normal breast tissue, plasma | Levels of GBP-2 are reduced in breast tumors compared to normal breast tissue. GBP-2 is reduced in TNBC tumors, higher stages of breast cancers, and in node positive tumors compared to other breast tumors. The GBP-2 promoter in about 87% of breast cancers was methylated. GBP-2 was downregulated in 73% of breast cancers, 26% had normal GBP-2 expression, and none showed elevation of GBP-2. The methylation status of the GBP-2 promoter in tumors matched with the methylation status of cell free DNA isolated from the plasma. GBP-2 promoter was methylated in 100% of stage III or IV breast tumors. | Rahvar, 2020 [25] |
| GBP-1 | Affymetrix Probe Sets | ||
|---|---|---|---|
| 202269_x_at | 231577_s_at | 202270_at | |
| Recurrence-Free Survival | |||
| All Breast Cancers | HR = 1.26 (1.14–1.39), p = 9.1 × 10−6 | HR = 1.21 (1.04–1.41), p = 0.012 | HR = 1.19 (1.07–1.31), p = 0.00088 |
| ER+ | HR = 1.02 (0.87–1.19), p = 0.83 | HR = 1.18 (0.89–1.58), p = 0.25 | HR = 1 (0.86–1.17), p = 0.97 |
| ER+/HER2− | HR = 1.02 (0.86–1.2), p = 0.85 | HR = 1.1 (0.8–1.51), p = 0.55 | HR = 1.1 (0.93–1.3), p = 0.28 |
| HER2+ | HR = 0.87 (0.7–1.08), p = 0.19 | HR = 1 (0.74–1.35), p = 1 | HR = 0.69 (0.55–0.86), p = 00081 |
| ER− | HR = 0.59 (0.47–0.76), p = 2 × 10−5 | HR = 0.58 (0.4–0.85), p = 0.0042 | HR = 0.58 (0.46–0.74), p = 1 × 10−5 |
| TNBC | HR = 0.5 (0.35–0.73), p = 0.00025 | HR = 0.31 (0.17–0.59), p = 0.00016 | HR = 0.45 (0.31–0.66), p = 2.1 × 10−5 |
| Overall Survival | |||
| All Breast Cancers | HR = 0.96 (0.79–1.15), p = 0.63 | HR = 0.91 (0.7–119), p = 0.5 | HR = 1.03 (0.85–1.24), p = 0.75 |
| ER+ | HR = 1 (0.72–1.38), p = 0.99 | HR = 0.94 (0.46–1.96), p = 0.88 | HR = 1.08 (0.78–1.49), p = 0.64 |
| ER+/HER2− | HR = 1.05 (0.73–1.5), p = 0.81 | HR = 0.92 (0.39–2.17), p = 0.85 | HR = 0.66 (0.45–0.95), p = 0.023 |
| HER2+ | HR = 0.7 (0.49–1.01), p = 0.58 | HR = 0.9 (0.55–1.49), p = 0.69 | HR = 1.12 (0.78–1.62), p = 0.53 |
| ER− | HR = 0.55 (0.37–0.82), p = 0.0033 | HR = 0.59 (0.35–0.98), p = 0.039 | HR = 0.55 (0.37–0.82), p = 0.003 |
| TNBC | HR = 0.35 (0.17–0.72), p = 0.003 | HR = 0.36 (0.16–0.78), p = 0.0071 | HR = 0.49 (0.25–0.98), p = 0.039 |
| Distant Metastasis-Free Survival | |||
| All Breast Cancers | HR = 1.15 (0.99–1.34), p = 0.072 | HR = 1.07 (0.82–1.39), p = 0.63 | HR = 1.16 (1–1.36), p = 0.053 |
| ER+ | HR = 1.13 (0.86–1.49), p = 0.37 | HR = 2.14 (0.98–4.65), p = 0.05 | HR = 0.99 (0.76–1.31), p = 0.96 |
| ER+/HER2− | HR = 1.09 (0.81–1.47), p = 0.58 | HR = 1.91 (0.69–5.29), p = 0.2 | HR = 1 (0.74–1.35), p = 0.99 |
| HER2+ | HR = 0.64 (0.45–0.89), p = 0.0083 | HR = 0.86 (0.54–1.35), p = 0.51 | HR = 0.73 (0.53–1.03), p = 0.07 |
| ER− | HR = 0.58 (0.42–0.79), p = 0.00062 | HR = 0.61 (0.38–0.98), p = 0.04 | HR = 0.58 (0.42–0.79), p = 0.00059 |
| TNBC | HR = 0.58 (0.38–0.9), p = 0.015 | HR = 0.47 (0.25–0.88), p = 0.017 | HR = 0.52 (0.34–0.81), p = 0.003 |
| GBP-5 | Affymetrix Probe Sets | |
|---|---|---|
| 229625_at | 23581_at | |
| Recurrence-Free Survival | ||
| All Breast Cancers | HR = 0.79 (0.68–0.92), p = 0.002 | HR = 0.78 (0.67–0.91), p = 0.0015 |
| ER+ | HR = 1.04 (0.78–1.38), p = 0.81 | HR = 1.05 (0.79–1.39), p = 0.76 |
| ER+/HER2− | HR = 1.1 (0.8–1.51), p = 0.55 | HR = 1.02 (0.75–1.4), p = 0.9 |
| HER2+ | HR = 0.58 (0.43–0.8), p = 0.00054 | HR = 0.59 (0.43–0.8), p = 0.00057 |
| ER− | HR = 0.56 (0.39–0.82), p = 0.0087 | HR = 0.56 (0.39–0.82), p = 0.0088 |
| TNBC | HR = 0.42 (0.23–0.76), p = 0.0032 | HR = 0.46 (0.26–0.83), p = 0.0088 |
| Overall Survival | ||
| All Breast Cancers | HR = 0.7 (0.53–0.92), p = 0.009 | HR = 0.72 (0.55–0.94), p = 0.017 |
| ER+ | HR = 0.59 (0.28–1.25), p = 0.17 | HR = 0.8 (0.39–1.67), p = 0.55 |
| ER+/HER2− | HR = 0.63 (0.26–1.49), p = 0.28 | HR = 0.72 (0.3–1.72), p = 0.46 |
| HER2+ | HR = 0.68 (0.41–1.13), p = 0.14 | HR = 0.6 (0.36–0.99), p = 0.045 |
| ER− | HR = 0.52 (0.31–0.88), p = 0.012 | HR = 0.4 (0.23–0.68), p = 0.00043 |
| TNBC | HR = 0.4 (0.19–0.86), p = 0.016 | HR = 0.41 (0.19–0.87), p = 0.017 |
| Distant Metastasis-Free Survival | ||
| All Breast Cancers | HR = 0.91 (0.7–1.19), p = 0.5 | HR = 0.96 (0.73–1.24), p = 0.74 |
| ER+ | HR = 1.32 (0.62–2.79), p = 0.47 | HR = 1.37 (0.65–2.9), p = 0.41 |
| ER+/HER2− | HR = 1.17 (0.43–3.15), p = 0.76 | HR = 1.06 (0.4–2.82), p = 0.91 |
| HER2+ | HR = 0.74 (0.47–1.17), p = 0.2 | HR = 0.74 (0.47–1.17), p = 0.19 |
| ER− | HR = 0.53 (0.33–0.86), p = 0.0082 | HR = 0.61 (0.38–0.98), p = 0.09 |
| TNBC | HR = 0.48 (0.26–0.9), p = 0.02 | HR = 0.61 (0.33–1.13), p = 0.11 |
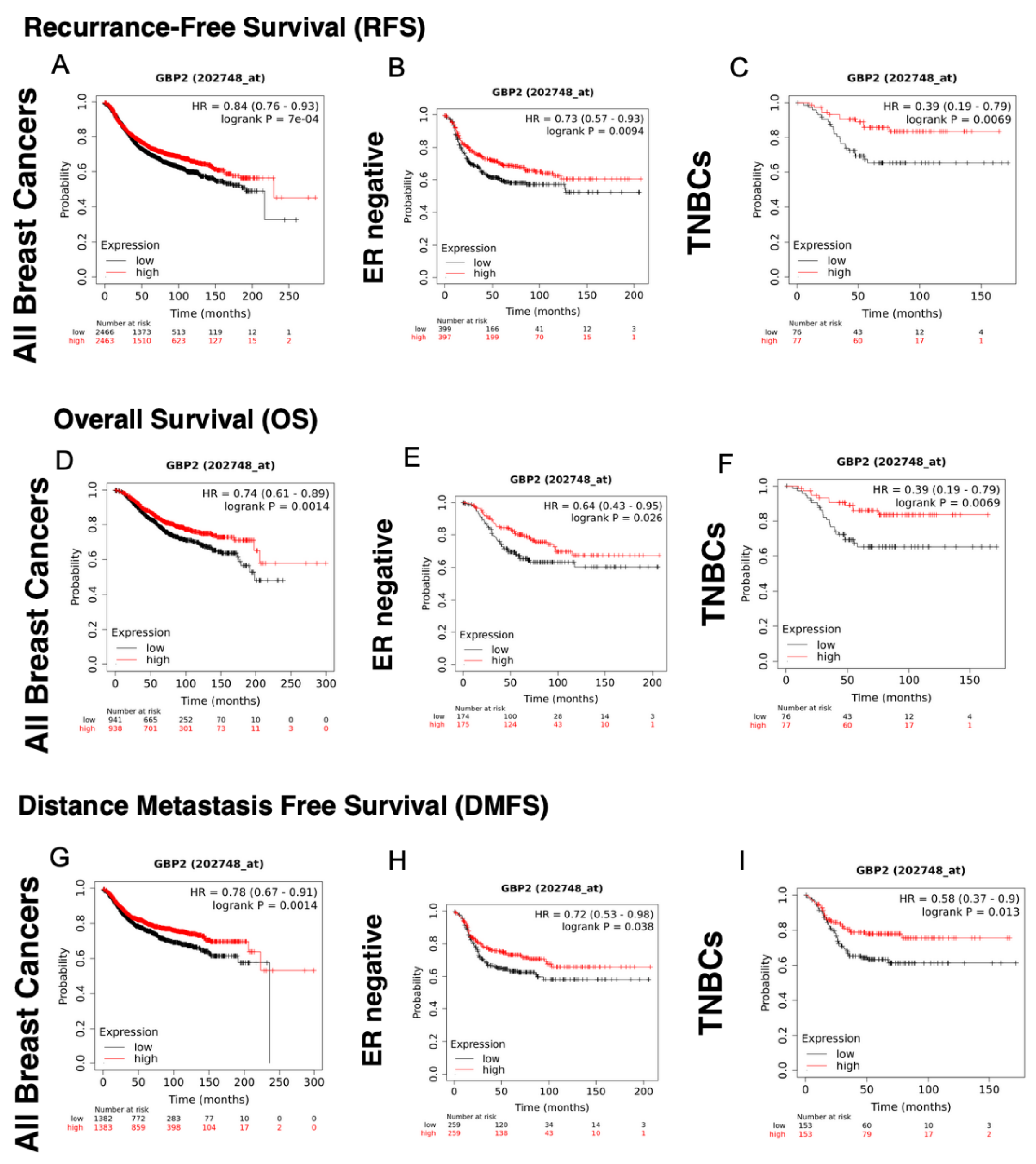
| GBP-2 | Affymetrix Probe Sets | |
|---|---|---|
| 202748_at | 242907_at | |
| Recurrence-Free Survival | ||
| All Breast Cancers | HR = 0.84 (0.76–0.93), p = 7 × 10−4 | HR = 0.72 (0.62–0.84), p = 2.8 × 10−5 |
| ER+ | HR = 0.86 (0.74–1.01), p = 0.061 | HR = 0.97 (0.72–1.29), p = 0.81 |
| ER+/HER2− | HR = 0.83 (0.7–0.98), p = 0.031 | HR = 0.88 (0.65–1.21), p = 0.45 |
| HER2+ | HR = 0.77 (0.62–0.96), p = 0.022 | HR = 0.85 (0.63–1.15), p = 0.3 |
| ER− | HR = 0.73 (0.57–0.93), p = 0.0094 | HR = 0.73 (0.57–0.93), p = 0.0095 |
| TNBC | HR = 0.59 (0.41–0.86), p = 0.0048 | HR = 0.34 (0.18–0.64), p = 0.00045 |
| Overall Survival | ||
| All Breast Cancers | HR = 0.74 (0.61–0.898), p = 0.0014 | HR = 0.6 (0.46–0.79), p = 0.00019 |
| ER+ | HR = 0.75 (0.55–1.04), p = 0.085 | HR = 0.49 (0.23–1.05), p = 0.061 |
| ER+/HER2− | HR = 0.72 (0.5–1.03), p = 0.071 | HR = 0.44 (0.18–1.09). p = 0.068 |
| HER2+ | HR = 0.82 (0.57–1.17), p = 0.27 | HR = 0.89 (0.54–1.47), p = 0.64 |
| ER− | HR = 0.64 (0.43–0.95), p = 0.026 | HR = 0.64 (0.43–0.95), p = 0.027 |
| TNBC | HR = 0.39 (0.19–0.79), p = 0.0069 | HR = 0.34 (0.16–0.75), p = 0.0052 |
| Distant Metastasis-Free Survival | ||
| All Breast Cancers | HR = 0.78 (0.67–0.91), p = 0.0014 | HR = 0.81 (0.62–1.06), p = 0.13 |
| ER+ | HR = 0.65 (0.49–0.86), p = 0.0022 | HR = 0.84 (0.39–1.78), p = 0.64 |
| ER+/HER2− | HR = 0.66 (0.49–0.9), p = 0.0079 | HR = 0.66 (0.49–0.9), p = 0.0079 |
| HER2+ | HR = 0.73 (0.52–1.02), p = 0.065 | HR = 0.9 (0.57–1.42), p = 0.65 |
| ER− | HR = 0.72 (0.53–0.98), p = 0.038 | HR = 0.75 (0.47–1.2), p = 0.23 |
| TNBC | HR = 0.58 (0.37–0.9), p = 0.013 | HR = 0.42 (0.22–0.8), p = 0.0064 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunt, E.N.; Kopacz, J.P.; Vestal, D.J. Unraveling the Role of Guanylate-Binding Proteins (GBPs) in Breast Cancer: A Comprehensive Literature Review and New Data on Prognosis in Breast Cancer Subtypes. Cancers 2022, 14, 2794. https://doi.org/10.3390/cancers14112794
Hunt EN, Kopacz JP, Vestal DJ. Unraveling the Role of Guanylate-Binding Proteins (GBPs) in Breast Cancer: A Comprehensive Literature Review and New Data on Prognosis in Breast Cancer Subtypes. Cancers. 2022; 14(11):2794. https://doi.org/10.3390/cancers14112794
Chicago/Turabian StyleHunt, Erin N., Jonathan P. Kopacz, and Deborah J. Vestal. 2022. "Unraveling the Role of Guanylate-Binding Proteins (GBPs) in Breast Cancer: A Comprehensive Literature Review and New Data on Prognosis in Breast Cancer Subtypes" Cancers 14, no. 11: 2794. https://doi.org/10.3390/cancers14112794