
PRKCA Overexpression Is Frequent in Young Oral Tongue Squamous Cell Carcinoma Patients and Is Associated with Poor Prognosis

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Patients and Methods
2.1. Study Design and Setting
2.2. TCGA Data Retrieval and Selection
2.3. Austrian Patient Sample
2.4. Smoking and Alcohol Consumption Status in the Patient Samples
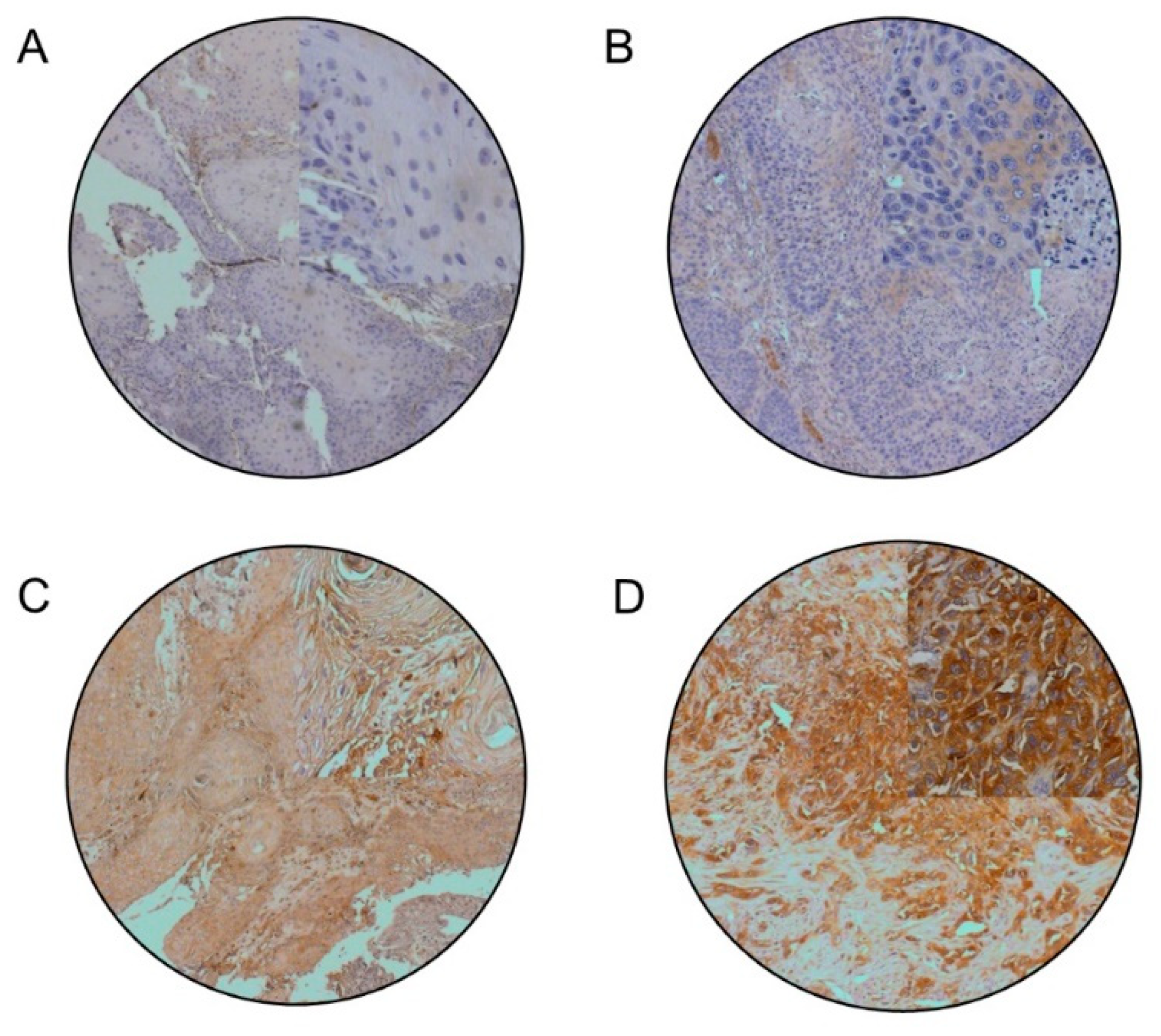
2.5. Immunohistochemistry
2.6. Protein Expression Quantification
2.7. Statistical Analysis
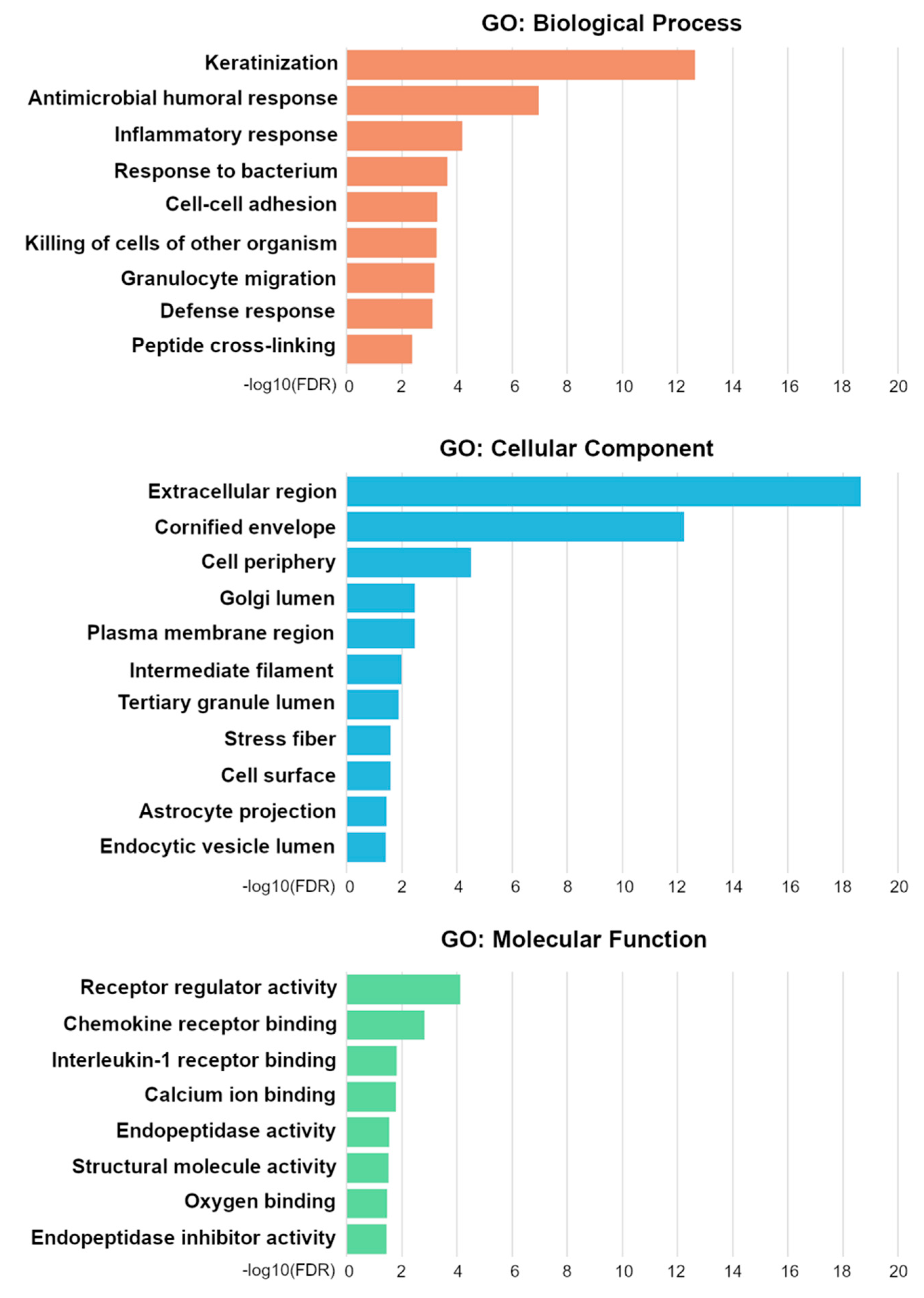
2.8. Differential mRNA Expression and Gene Ontology Enrichment Analysis
3. Results
3.1. Patient Characteristics
3.2. PRKCA Is Frequently Overexpressed in Young OTSCC Patients
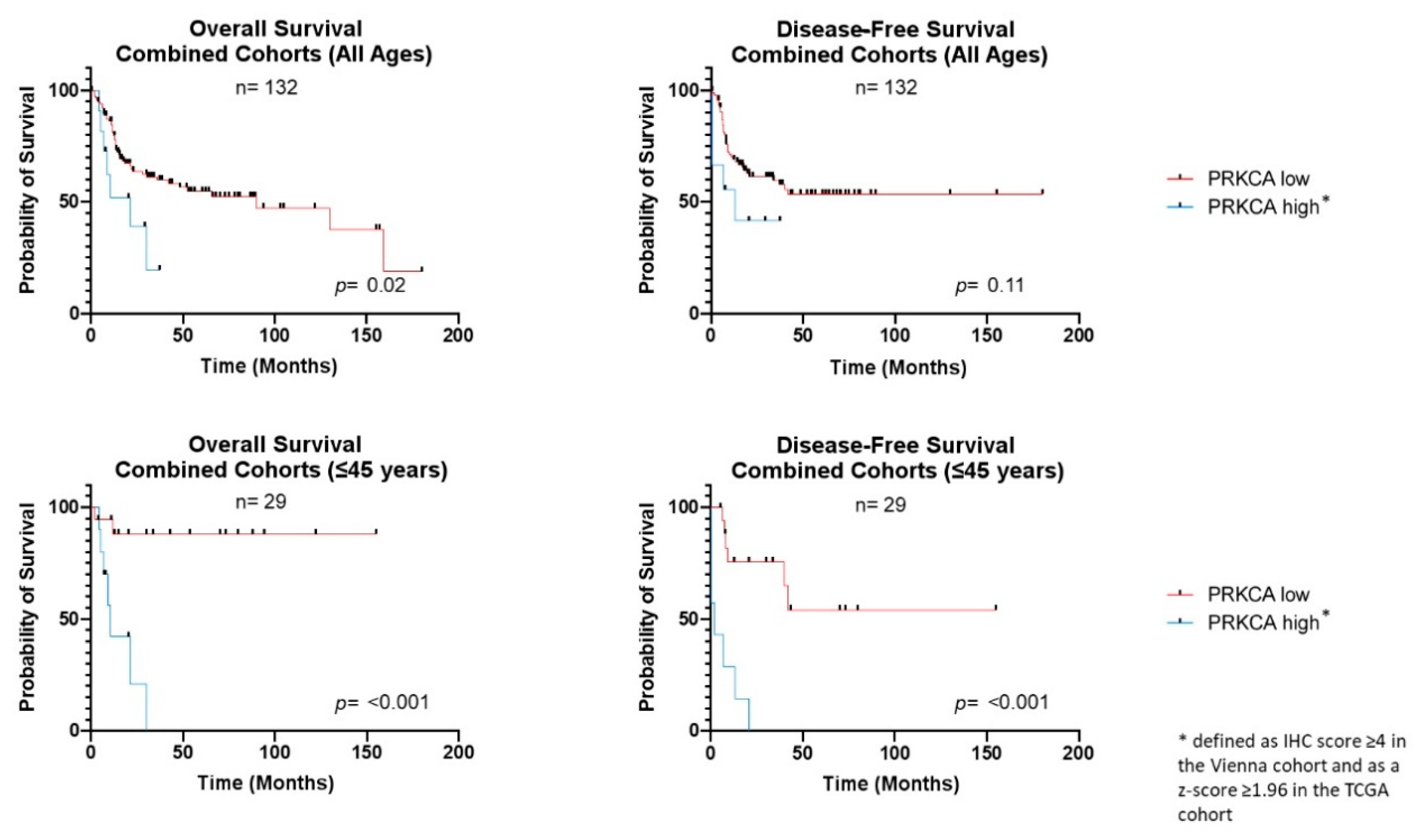
3.3. PRKCA Overexpression Is Associated with Adverse Clinical Outcome
3.4. PRKCA Overexpression Is Frequent in Alcohol and Tobacco Negative OTSCC
3.5. Messenger RNA Expression Profiles Differ Significantly between PRKCA Positive and PRKCA Negative OTSCC
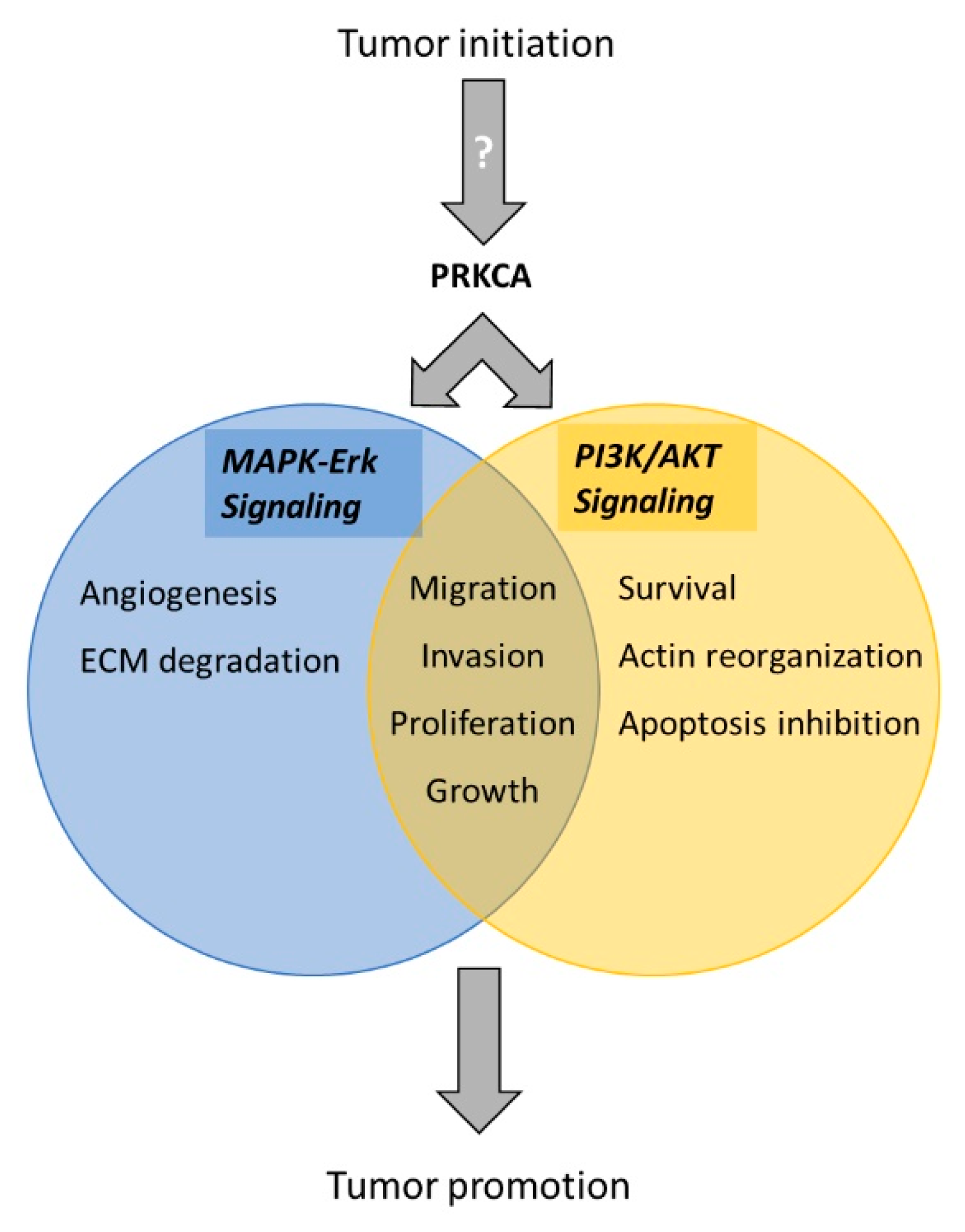
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN. CA. Cancer J. Clin. 2018. [Google Scholar] [CrossRef] [Green Version]
- Reshmi, S.C.; Saunders, W.S.; Kudla, D.M.; Ragin, C.R.; Gollin, S.M. Chromosomal instability and marker chromosome evolution in oral squamous cell carcinoma. Genes Chromosom. Cancer 2004. [Google Scholar] [CrossRef]
- Siebers, T.J.H.; Bergshoeff, V.E.; Otte-Höller, I.; Kremer, B.; Speel, E.J.M.; Van Der Laak, J.A.W.M.; Merkx, M.A.W.; Slootweg, P.J. Chromosome instability predicts the progression of premalignant oral lesions. Oral Oncol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Forastiere, A.; Koch, W.; Trotti, A.; Sidransky, D. Head and neck cancer. N. Engl. J. Med. 2001. [Google Scholar] [CrossRef] [PubMed]
- Pires, F.R.; Ramos, A.B.; de Oliveira, J.B.C.; Tavares, A.S.; de Luz, P.S.R.; dos Santos, T.C.R.B. Oral squamous cell carcinoma: Clinicopathological features from 346 cases from a single oral pathology service during an 8-year period. J. Appl. Oral Sci. 2013. [Google Scholar] [CrossRef]
- Troeltzsch, M.; Knösel, T.; Eichinger, C.; Probst, F.; Troeltzsch, M.; Woodlock, T.; Mast, G.; Ehrenfeld, M.; Otto, S. Clinicopathologic features of oral squamous cell carcinoma: Do they vary in different age groups? J. Oral Maxillofac. Surg. 2014. [Google Scholar] [CrossRef] [PubMed]
- Sturgis, E.M.; Cinciripini, P.M. Trends in head and neck cancer incidence in relation to smoking prevalence. Cancer 2007. [Google Scholar] [CrossRef]
- Moore, S.R.; Johnson, N.W.; Pierce, A.M.; Wilson, D.F. The epidemiology of tongue cancer: A review of global incidence. Oral Dis. 2000. [Google Scholar] [CrossRef]
- Hussein, A.A.; Helder, M.N.; de Visscher, J.G.; Leemans, C.R.; Braakhuis, B.J.; de Vet, H.C.W.; Forouzanfar, T. Global incidence of oral and oropharynx cancer in patients younger than 45 years versus older patients: A systematic review. Eur. J. Cancer 2017. [Google Scholar] [CrossRef]
- Tota, J.E.; Anderson, W.F.; Coffey, C.; Califano, J.; Cozen, W.; Ferris, R.L.; St. John, M.; Cohen, E.E.W.; Chaturvedi, A.K. Rising incidence of oral tongue cancer among white men and women in the United States, 1973–2012. Oral Oncol. 2017. [Google Scholar] [CrossRef]
- Harris, S.L.; Kimple, R.J.; Hayes, D.N.; Couch, M.E.; Rosenman, J.G. Never-smokers, never-drinkers: Unique clinical subgroup of young patients with head and neck squamous cell cancers. Head Neck 2010. [Google Scholar] [CrossRef] [PubMed]
- Poling, J.S.; Ma, X.J.; Bui, S.; Luo, Y.; Li, R.; Koch, W.M.; Westra, W.H. Human papillomavirus (HPV) status of non-tobacco related squamous cell carcinomas of the lateral tongue. Oral Oncol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Castellsagué, X.; Alemany, L.; Quer, M.; Halec, G.; Quirós, B.; Tous, S.; Clavero, O.; Alòs, L.; Biegner, T.; Szafarowski, T.; et al. HPV Involvement in Head and Neck Cancers: Comprehensive Assessment of Biomarkers in 3680 Patients. J. Natl. Cancer Inst. 2016. [Google Scholar] [CrossRef]
- Zafereo, M.E.; Xu, L.; Dahlstrom, K.R.; Viamonte, C.A.; El-Naggar, A.K.; Wei, Q.; Li, G.; Sturgis, E.M. Squamous cell carcinoma of the oral cavity often overexpresses p16 but is rarely driven by human papillomavirus. Oral Oncol. 2016, 56, 47–53. [Google Scholar] [CrossRef] [Green Version]
- de Morais, E.F.; Mafra, R.P.; Gonzaga, A.K.G.; de Souza, D.L.B.; Pinto, L.P.; da Silveira, É.J.D. Prognostic Factors of Oral Squamous Cell Carcinoma in Young Patients: A Systematic Review. J. Oral Maxillofac. Surg. 2017. [Google Scholar] [CrossRef]
- Hilly, O.; Shkedy, Y.; Hod, R.; Soudry, E.; Mizrachi, A.; Hamzany, Y.; Bachar, G.; Shpitzer, T. Carcinoma of the oral tongue in patients younger than 30 years: Comparison with patients older than 60 years. Oral Oncol. 2013. [Google Scholar] [CrossRef]
- Jeon, J.-H.; Kim, M.G.; Park, J.Y.; Lee, J.H.; Kim, M.J.; Myoung, H.; Choi, S.W. Analysis of the outcome of young age tongue squamous cell carcinoma. Maxillofac. Plast. Reconstr. Surg. 2017. [Google Scholar] [CrossRef]
- Pickering, C.R.; Zhang, J.; Neskey, D.M.; Zhao, M.; Jasser, S.A.; Wang, J.; Ward, A.; Tsai, C.J.; Alves, M.V.O.; Zhou, J.H.; et al. Squamous cell carcinoma of the oral tongue in young non-smokers is genomically similar to tumors in older smokers. Clin. Cancer Res. 2014. [Google Scholar] [CrossRef] [Green Version]
- dos Santos Costa, S.F.; Brennan, P.A.; Gomez, R.S.; Fregnani, E.R.; Santos-Silva, A.R.; Martins, M.D.; de Castro-Junior, G.; Rahimi, S.; Fonseca, F.P. Molecular basis of oral squamous cell carcinoma in young patients: Is it any different from older patients? J. Oral Pathol. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Coates, P.J.; Boldrup, L.; Wang, L.; Krejci, A.; Hupp, T.; Fahraeus, R.; Norberg-Spaak, L.; Sgaramella, N.; Wilms, T.; et al. Copy number variation: A prognostic marker for young patients with squamous cell carcinoma of the oral tongue. J. Oral Pathol. Med. 2019. [Google Scholar] [CrossRef]
- Maroun, C.A.; Zhu, G.; Fakhry, C.; Gourin, C.G.; Seiwert, T.Y.; Vosler, P.S.; Tan, M.; Koch, W.; Eisele, D.W.; Pardoll, D.M.; et al. An Immunogenomic Investigation of Oral Cavity Squamous Cell Carcinoma in Patients Aged 45 Years and Younger. Laryngoscope 2020. Epub ahead. [Google Scholar] [CrossRef]
- Lingen, M.W.; Chang, K.W.; McMurray, S.J.; Solt, D.B.; Kies, M.S.; Mittal, B.B.; Haines, G.K.; Pelzer, H.J. Overexpression of p53 in squamous cell carcinoma of the tongue in young patients with no known risk factors is not associated with mutations in exons 5-9. Head Neck 2000. [Google Scholar] [CrossRef]
- Li, R.; Faden, D.L.; Fakhry, C.; Langelier, C.; Jiao, Y.; Wang, Y.; Wilkerson, M.D.; Pedamallu, C.S.; Old, M.; Lang, J.; et al. Clinical, genomic, and metagenomic characterization of oral tongue squamous cell carcinoma in patients who do not smoke. Head Neck 2015. [Google Scholar] [CrossRef]
- Sakamoto, K.; Imanishi, Y.; Tomita, T.; Shimoda, M.; Kameyama, K.; Shibata, K.; Sakai, N.; Ozawa, H.; Shigetomi, S.; Fujii, R.; et al. Overexpression of SIP1 and downregulation of e-cadherin predict delayed neck metastasis in stage I/II oral tongue squamous cell carcinoma after partial glossectomy. Ann. Surg. Oncol. 2012. [Google Scholar] [CrossRef]
- Wang, C.; Liu, X.; Huang, H.; Ma, H.; Cai, W.; Hou, J.; Huang, L.; Dai, Y.; Yu, T.; Zhou, X. Deregulation of Snai2 is associated with metastasis and poor prognosis in tongue squamous cell carcinoma. Int. J. Cancer 2012. [Google Scholar] [CrossRef] [Green Version]
- Albert, S.; Hourseau, M.; Halimi, C.; Serova, M.; Descatoire, V.; Barry, B.; Couvelard, A.; Riveiro, M.E.; Tijeras-Raballand, A.; De Gramont, A.; et al. Prognostic value of the chemokine receptor CXCR4 and epithelial-to- mesenchymal transition in patients with squamous cell carcinoma of the mobile tongue. Oral Oncol. 2012. [Google Scholar] [CrossRef]
- Liang, X.; Zheng, M.; Jiang, J.; Zhu, G.; Yang, J.; Tang, Y. Hypoxia-inducible factor-1 alpha, in association with TWIST2 and SNIP1, is a critical prognostic factor in patients with tongue squamous cell carcinoma. Oral Oncol. 2011. [Google Scholar] [CrossRef]
- Kang, F.W.; Gao, Y.; Que, L.; Sun, J.; Wang, Z.L. Hypoxia-inducible factor-1α overexpression indicates poor clinical outcomes in tongue squamous cell carcinoma. Exp. Ther. Med. 2013. [Google Scholar] [CrossRef] [Green Version]
- Almangush, A.; Heikkinen, I.; Mäkitie, A.A.; Coletta, R.D.; Läärä, E.; Leivo, I.; Salo, T. Prognostic biomarkers for oral tongue squamous cell carcinoma: A systematic review and meta-analysis. Br. J. Cancer 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Yang, Y.; Xiao, X.; Wang, C.; Zhang, X.; Wang, L.; Zhang, X.; Li, W.; Zheng, G.; Wang, S.; et al. Sox2 nuclear expression is closely associated with poor prognosis in patients with histologically node-negative oral tongue squamous cell carcinoma. Oral Oncol. 2011. [Google Scholar] [CrossRef]
- Huang, C.F.; Xu, X.R.; Wu, T.F.; Sun, Z.J.; Zhang, W.F. Correlation of ALDH1, CD44, OCT4 and SOX2 in tongue squamous cell carcinoma and their association with disease progression and prognosis. J. Oral Pathol. Med. 2014. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossman, R.L.; Heath, A.P.; Ferretti, V.; Varmus, H.E.; Lowy, D.R.; Kibbe, W.A.; Staudt, L.M. Toward a Shared Vision for Cancer Genomic Data. N. Engl. J. Med. 2016, 375, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Risso, D.; Schwartz, K.; Sherlock, G.; Dudoit, S. GC-content normalization for RNA-Seq data. BMC Bioinformatics 2011, 12, 480. [Google Scholar] [CrossRef] [Green Version]
- Berenblum, I. The Cocarcinogenic Action of Croton Resin. Cancer Res. 1941. Available online: https://cancerres.aacrjournals.org/content/1/1/44 (accessed on 24 April 2021).
- Niedel, J.E.; Kuhn, L.J.; Vandenbark, G.R. Phorbol diester receptor copurifies with protein kinase C. Proc. Natl. Acad. Sci. USA 1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.-J.; Pan, W.-W.; Liu, S.-B.; Shen, Z.-F.; Xu, Y.; Hu, L.-L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, M.; De Santis, M.C.; Braccini, L.; Gulluni, F.; Hirsch, E. PI3K/AKT signaling pathway and cancer: An updated review. Ann. Med. 2014, 46, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, J.; Flechner, L.; Hyun, T.; Yam, A.; Franke, T.F.; Pierce, J.H. Protein kinase C-α overexpression stimulates Akt activity and suppresses apoptosis induced by interleukin 3 withdrawal. Oncogene 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.K.; Galoforo, S.S.; Berns, C.M.; Martinez, A.A.; Corry, P.M.; Guan, K.L.; Lee, Y.J. Elevated levels of ERK2 in human breast carcinoma MCF-7 cells transfected with protein kinase Cα. Cell Prolif. 1996. [Google Scholar] [CrossRef]
- Salama, M.F.; Liu, M.; Clarke, C.J.; Espaillat, M.P.; Haley, J.D.; Jin, T.; Wang, D.; Obeid, L.M.; Hannun, Y.A. PKCα is required for Akt-mTORC1 activation in non-small cell lung carcinoma (NSCLC) with EGFR mutation. Oncogene 2019. [Google Scholar] [CrossRef]
- Wu, B.; Zhou, H.; Hu, L.; Mu, Y.; Wu, Y. Involvement of PKCα activation in TF/VIIa/PAR2-induced proliferation, migration, and survival of colon cancer cell SW620. Tumor Biol. 2013. [Google Scholar] [CrossRef]
- Mandil, R.; Ashkenazi, E.; Blass, M.; Kronfeld, I.; Kazimirsky, G.; Rosenthal, G.; Umansky, F.; Lorenzo, P.S.; Blumberg, P.M.; Brodie, C. Protein kinase Cα and protein kinase Cδ play opposite roles in the proliferation and apoptosis of glioma cells. Cancer Res. 2001, 61, 4612–4619. [Google Scholar]
- Zhang, H.X.; Liu, O.S.; Deng, C.; He, Y.; Feng, Y.Q.; Ma, J.A.; Hu, C.H.; Tang, Z.G. Genome-wide gene expression profiling of tongue squamous cell carcinoma by RNA-seq. Clin. Oral Investig. 2018, 22, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Cardesa, A.; Nadal, A. Carcinoma of the head and neck in the HPV era. Acta Dermatovenerol. Alpina Pannonica Adriat. 2011, 20, 161–173. [Google Scholar]
- Dmello, C.; Srivastava, S.S.; Tiwari, R.; Chaudhari, P.R.; Sawant, S.; Vaidya, M.M. Multifaceted role of keratins in epithelial cell differentiation and transformation. J. Biosci. 2019, 44, 33. [Google Scholar] [CrossRef]
- Plzák, J.; Bouček, J.; Bandúrová, V.; Kolář, M.; Hradilová, M.; Szabo, P.; Lacina, L.; Chovanec, M.; Smetana, K. The Head and Neck Squamous Cell Carcinoma Microenvironment as a Potential Target for Cancer Therapy. Cancers 2019, 11, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tampa, M.; Mitran, M.I.; Mitran, C.I.; Sarbu, M.I.; Matei, C.; Nicolae, I.; Caruntu, A.; Tocut, S.M.; Popa, M.I.; Caruntu, C.; et al. Mediators of Inflammation—A Potential Source of Biomarkers in Oral Squamous Cell Carcinoma. J. Immunol. Res. 2018, 2018, 1061780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winck, F.V.; Prado Ribeiro, A.C.; Ramos Domingues, R.; Ling, L.Y.; Riaño-Pachón, D.M.; Rivera, C.; Brandão, T.B.; Gouvea, A.F.; Santos-Silva, A.R.; Coletta, R.D.; et al. Insights into immune responses in oral cancer through proteomic analysis of saliva and salivary extracellular vesicles. Sci. Rep. 2015, 5, 16305. [Google Scholar] [CrossRef] [PubMed]
- Pfeifhofer, C.; Gruber, T.; Letschka, T.; Thuille, N.; Lutz-Nicoladoni, C.; Hermann-Kleiter, N.; Braun, U.; Leitges, M.; Baier, G. Defective IgG2a/2b Class Switching in PKCα −/− Mice. J. Immunol. 2006, 176, 6004–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, P.S.; Sutton, C.R.; Rao, S. Protein kinase C in the immune system: From signalling to chromatin regulation. Immunology 2015, 146, 508–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haabeth, O.A.W.; Lorvik, K.B.; Hammarström, C.; Donaldson, I.M.; Haraldsen, G.; Bogen, B.; Corthay, A. Inflammation driven by tumour-specific Th1 cells protects against B-cell cancer. Nat. Commun. 2011, 2, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Yang, Z.; Lu, N. A new role for the PI3K/Akt signaling pathway in the epithelial-mesenchymal transition. Cell Adh. Migr. 2015, 9, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.R.; Check, D.P.; Caporaso, N.E.; Travis, W.D.; Devesa, S.S. US lung cancer trends by histologic type. Cancer 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Li, M.; Wu, N.; Chen, Y.; Yang, F. Time trends in epidemiologic characteristics and imaging features of lung adenocarcinoma: A population study of 21,113 cases in China. PLoS ONE 2015. [Google Scholar] [CrossRef]
- Jiang, H.; Fu, Q.; Song, X.; Ge, C.; Li, R.; Li, Z.; Zeng, B.; Li, C.; Wang, Y.; Xue, Y.; et al. HDGF and prkca upregulation is associated with a poor prognosis in patients with lung adenocarcinoma. Oncol. Lett. 2019. [Google Scholar] [CrossRef] [Green Version]
- Isakov, N. Protein kinase C (PKC) isoforms in cancer, tumor promotion and tumor suppression. Semin. Cancer Biol. 2018. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | TCGA (n = 98) | Vienna (n = 34) | Combined (n = 132) | |||
|---|---|---|---|---|---|---|
| Age (years) | ≤45 (n = 15) | >45(n = 82) a | ≤45 (n = 14) | >45 (n = 20) | ≤45 (n = 29) | >45 (n = 103) |
| Sex | ||||||
| Male Female | 8 (53.3%) 7 (46.7%) | 55 (66.3%) 28 (33.7%) | 9 (64.3%) 5 (35.7%) | 11 (55%) 9 (45%) | 17 (58.6%) 12 (41.4%) | 66 (64.1%) 37 (35.9%) |
| T class.b | ||||||
| Tx T1 T2 T3 T4 | 0 3 (20%) 6 (40%) 3 (20%) 3 (20%) | 3 (2.6%) 11 (13.3%) 31 (37.3%) 26 (31.3%) 12 (14.5%) | 1 (7.1%) 6 (42.9%) 4 (28.6%) 1 (7.1%) 2 (14.3%) | 0 7 (35%) 8 (40%) 2 (10%) 3 (15%) | 1 (3.4%) 9 (31.1%) 10 (34.5%) 4 (13.8%) 5 (17.2%) | 3 (2.9%) 18 (17.5%) 39 (37.9%) 28 (27.2%) 15 (14.5%) |
| N class. | ||||||
| Nx N0 N1 N2 N3 | 0 8 (53.4%) 2 (13.3%) 5 (33.3%) 0 | 4 (4.8%) 40 (48.2%) 16 19.3%) 23 (27.7%) 0 | 1 (7.1%) 9 (64.3%) 2 (14.3%) 2 (14.3%) 0 | 0 14 (70%) 4 (20%) 2 (10%) 0 | 1 (3.4%) 17 (58.7%) 4 (13.8%) 7 (24.1%) 0 | 4 (3.9%) 54 (52.4%) 20 (19.4%) 25 (24.3%) 0 |
| M class. | ||||||
| Mx M0 M1 | 10 (66.7%) 5 (33.3%) 0 | 58 (69.9%) 25 (30.1%) 0 | 5 (35.7%) 9 (64.3%) 0 | 3 (15%) 17 (85%) 0 | 15 (51.7%) 14 (48.3%) 0 | 61(59.2%) 42 (40.8%) 0 |
| Tobacco | ||||||
| Yes No Unkn. | 10 (66.7%) 5 (33.3%) 0 | 26 (31.3%) 56 (67.5%) 1 (1.2%) | 10(71.5%) 3 (21.4%) 1 (7.1%) | 6 (30%) 14 (70%) 0 | 20 (69%) 8 (27.6%) 1 (3.4%) | 32 (31.1%) 70 (68%) 1 (0.9%) |
| Alcohol | ||||||
| Yes No Unkn. | 1 (6.7%) 6 (40%) 8 (53.3%) | 30 (36.1%) 5 (6%) 48 (57.9%) | 2 (14.3%) 11(78.6%) 1 ((7.1%) | 10 (50%) 10 (50%) 0 | 3 (10.3%) 17 (58.7%) 9 (31%) | 40 (38.8%) 15 (14.6%) 48 (46.6%) |
| Primary treatment | ||||||
| Surgery CR RT EPT | 15 (100%) 0 0 0 | 83 (100%) 0 0 0 | 11(78.6%) 2 (14.3%) 1 (7.1%) 0 | 9 (45%) 3 (15%) 2 (10%) 6 (30%) | 26 (89.7%) 2 (6.9%) 1 (3.4%) 0 | 92 (89.3%) 3 (2.9%) 2 (1.9%) 6 (5.8%) |
| Neck diss. | ||||||
| Yes No | 13 (86.7%) 2 (13.3%) | 76 (91.6%) 7 (8.4%) | 10(71.4%) 4 (28.6%) | 10 (50%) 10 (50%) | 23 (79.3%) 6 (20.7%) | 86 (83.5%) 17 (16.5%) |
| Adjuvant treatment | ||||||
| PORT CR SS None | n/a n/a n/a n/a | n/a n/a n/a n/a | 5 (35.8%) 1 (7.1%) 0 8 (57.1%) | 1 (5%) 0 3 (15%) 16 (80%) | n/a n/a n/a n/a | n/a n/a n/a n/a |
| Overall Survival (Combined Cohorts, All Ages) | ||||||||
| Analysis | Univariate | Multivariate | ||||||
| Parameter | Hazard Ratio | Lower 95% | Upper 95% | p-value | Hazard Ratio | Lower 95% | Upper 95% | p-value |
| Age | 1.01 | 0.99 | 1.03 | 0.19 | 1.02 | 0.99 | 1.04 | 0.15 |
| T class | 1.86 | 1.41 | 2.44 | <0.01 | 1.80 | 1.31 | 2.45 | <0.01 |
| N class | 1.38 | 1.15 | 1.64 | <0.01 | 1.24 | 0.99 | 1.2 | 0.05 |
| PRKCA | 3.62 | 1.18 | 11.45 | 0.02 | 3.57 | 1.53 | 8.56 | <0.01 |
| Smoking | 1.54 | 0.75 | 4.45 | 0.29 | 1.04 | 0.95 | 1.14 | 0.46 |
| Sex | 1.08 | 0.89 | 1.24 | 0.45 | 1.07 | 0.91 | 1.15 | 0.48 |
| Disease-Free Survival (Combined Cohorts, All Ages) | ||||||||
| Analysis | Univariate | Multivariate | ||||||
| Parameter | Hazard Ratio | Lower 95% | Upper 95% | p-value | Hazard Ratio | Lower 95% | Upper 95% | p-value |
| Age | 0.98 | 0.95 | 1.01 | 0.58 | 0.99 | 0.95 | 1.02 | 0.62 |
| T class | 0.72 | 0.34 | 1.49 | 0.26 | 0.86 | 0.40 | 1.83 | 0.70 |
| N class | 0.58 | 0.23 | 1.39 | 0.22 | 0.61 | 0.23 | 1.58 | 0.31 |
| PRKCA | 2.82 | 0.78 | 10.10 | 0.11 | 1.68 | 0.25 | 13.23 | 0.55 |
| Smoking | 0.98 | 0.68 | 5.03 | 0.55 | 0.97 | 0.88 | 4.66 | 0.58 |
| Sex | 1.24 | 0.66 | 3.32 | 0.69 | 1.13 | 0.76 | 3.02 | 0.64 |
| Cohort | Median Survival a (Months) | UV HR (95% CI) | UV p-Value c | MV HR (95% CI) | MV p-Value d | |
|---|---|---|---|---|---|---|
| PRKCA positiveb | PRKCA negative | |||||
| Vienna All OS | 15 | 130 | 34.34 (4.15–284.1) | 0.001 | - | - |
| Vienna Young OS | 15 | Undef. a | 141.8 (12.24–1642) | <0.001 | - | - |
| Vienna All DFS | 0.5 | Undef. a | 107.7 (10.03–1158) | <0.001 | - | - |
| Vienna Young DFS | 0.5 | Undef. a | 31.15 (3.56–272.7) | 0.002 | - | - |
| Vienna All TU at LFU | 15 | Undef. a | 53.07 (5.82–483.9) | <0.001 | - | - |
| Vienna Young TU at LFU | 15 | 94 | 44.25 (4.87–401.9) | <0.001 | - | - |
| TCGA DFS All | Undef. a | Undef. a | 0.58 (0.14–2.47) | 0.46 | - | - |
| TCGA OS All | Undef. a | 52.27 | 1.51 (0.38–5.95) | 0.56 | - | - |
| TCGA OS Young | 10.74 | Undef. a | 3.2 (0.47–21.6) | 0.23 | - | - |
| TCGA DFS Young | 6.83 | Undef. a | 16.5 (1.69–161.2) | 0.02 | - | - |
| Combined OS All | 21 | 90.05 | 3.62 (1.18–11.45) | 0.02 | 3.57 (1.53–8.56) | < 0.01 |
| Combined DFS All | 13 | Undef. a | 2.82 (0.78–10.10) | 0.11 | 1.68 (0.25–13.23) | 0.55 |
| Combined OS Young | 10.75 | Undef. a | 18.95 (4.03–88.87) | <0.001 | - | - |
| Combined DFS Young | 2.17 | Undef. a | 29.65 (5.96–147.40) | <0.001 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parzefall, T.; Schnoell, J.; Monschein, L.; Foki, E.; Liu, D.T.; Frohne, A.; Grasl, S.; Pammer, J.; Lucas, T.; Kadletz, L.; et al. PRKCA Overexpression Is Frequent in Young Oral Tongue Squamous Cell Carcinoma Patients and Is Associated with Poor Prognosis. Cancers 2021, 13, 2082. https://doi.org/10.3390/cancers13092082
Parzefall T, Schnoell J, Monschein L, Foki E, Liu DT, Frohne A, Grasl S, Pammer J, Lucas T, Kadletz L, et al. PRKCA Overexpression Is Frequent in Young Oral Tongue Squamous Cell Carcinoma Patients and Is Associated with Poor Prognosis. Cancers. 2021; 13(9):2082. https://doi.org/10.3390/cancers13092082
Chicago/Turabian StyleParzefall, Thomas, Julia Schnoell, Laura Monschein, Elisabeth Foki, David Tianxiang Liu, Alexandra Frohne, Stefan Grasl, Johannes Pammer, Trevor Lucas, Lorenz Kadletz, and et al. 2021. "PRKCA Overexpression Is Frequent in Young Oral Tongue Squamous Cell Carcinoma Patients and Is Associated with Poor Prognosis" Cancers 13, no. 9: 2082. https://doi.org/10.3390/cancers13092082