1. Introduction
Large cohort-based genome-wide association studies provided us with the tools and knowledge to understand numerous phenotypic traits and diseases by single nucleotide polymorphisms (SNPs) and short (<50 bp) insertions and deletions in the genome. It is, however, more challenging to assess the role of the larger structural variations which has proven to be important for the regulation and function of many gene products. This is particularly the case for copy number variations (CNVs) [
1] which were first described in healthy humans [
2] but have since been associated with diseases, especially neurodevelopmental disorders and cancer [
3,
4]. CNVs are estimated to contribute to 4.8–9.5% of the genome and one or multiple exon copy number changes can affect gene expression levels or induce chromosomal rearrangements causing various disorders and diseases [
5,
6].
The current clinical standard method for CNV assessment remains array-based CNV identification, either from array-based comparative hybridization or SNP-array approaches [
7,
8]. While these arrays provide relatively accurate, cost-effective, and precise identification of CNVs, the use of short-read sequencing (or next generation sequencing, NGS), is not limited to the specific regions included on the arrays, has higher potential to identify novel CNVs, and has higher resolution at predicting both the breakpoints and shorter CNVs [
9]. Long-read sequencing is still cost-preventive for routine diagnostics and uniquely suited for structural variants (SVs). The alternative-NGS-bears the potential of a single assay for complete genomic analysis that allows for automated identification of both SNPs and SVs from the same data. CNV calling from NGS does, however, create new challenges such as dealing with variable coverage across the genome, alignment bias for deletions, read-length limit and insensitiveness towards repetitive and breakpoint regions [
10]. Furthermore, short-read sequencing increases mapping ambiguity consequently increasing the complexity of CNV detection [
10]. This is particularly true for whole exome sequencing (WES) or targeted gene panels, as the sequencing coverage and read depth in different areas are highly variable.
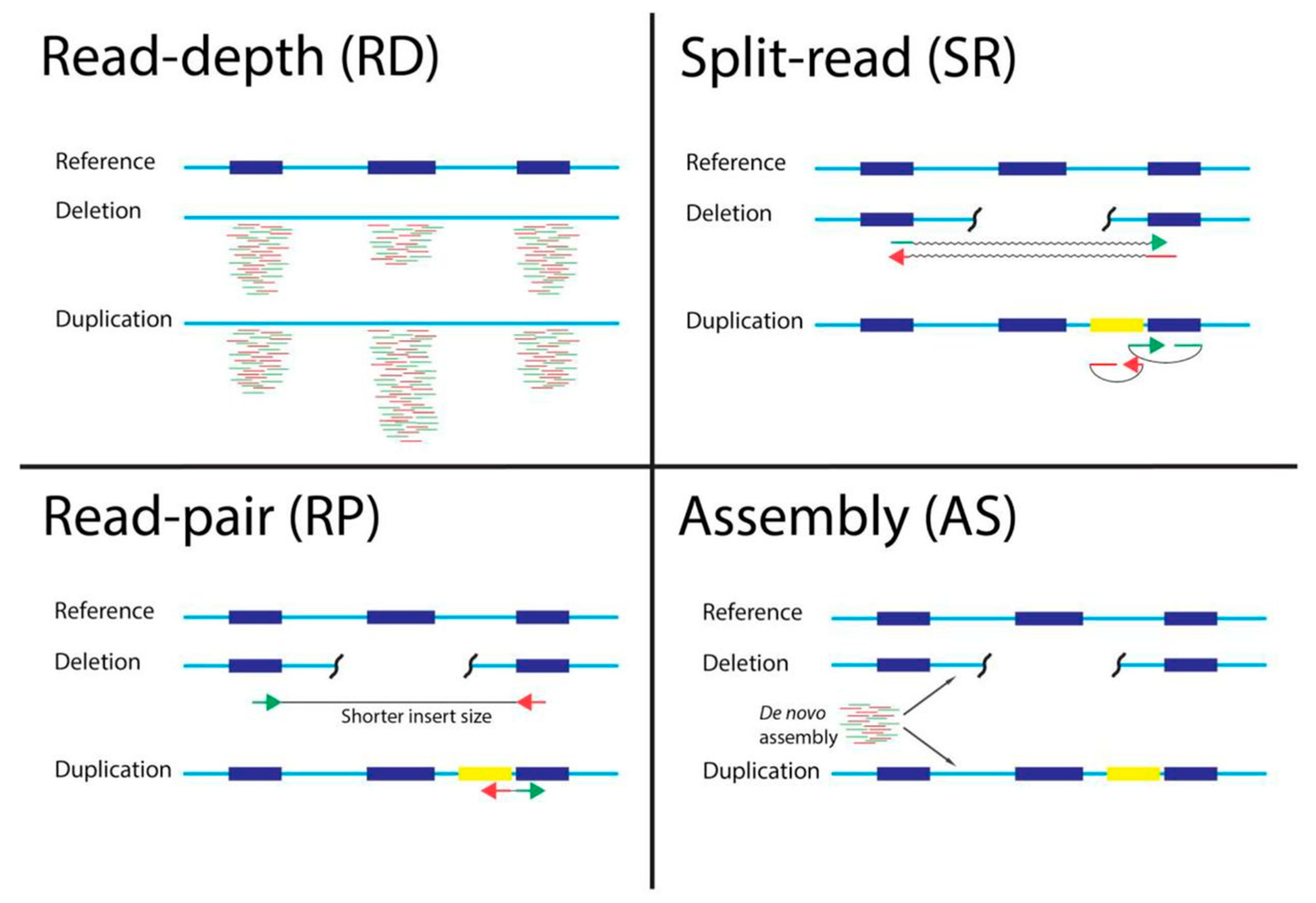
CNV calling algorithms can be based on one or more approaches: read-pair (RP), read-depth (RD), split read (SR), or assembly (AS) algorithms [
11] (
Figure 1). Most CNV calling tools are based on RD algorithms predicting CNVs from the changes based on read coverage in different areas of the genome.
With the increasing attention for the identification of SVs, including CNVs, and their clinical use, multiple SV databases have been established, such as CNV tracks in the UCSC database [
12,
13] or gnomAD SV [
14]. These databases help to evaluate predicted CNVs more accurately; however, the correct CNV identification itself remains challenging. In recent years, a number of tools for calling CNVs from NGS sequencing data, have been developed [
11]. Although a few recent studies [
15,
16] have benchmarked newly available SV callers, many benchmark studies have often been performed using known NGS datasets based on a single sample or a limited number of samples [
9,
11,
15,
16,
17], and a lack of comprehensive benchmark of various SV callers using multiple samples has been a major challenge for SV (including CNV) detection.
Here, we report an evaluation of 11 CNV detection tools for NGS and aim to identify the most reliable and clinically applicable software, whether based on WES or whole genome sequencing (WGS) originating from multiple individuals. We used the standard CytoScan HD SNP-array as a reference method for CNV detection and assessed the potential increase in sensitivity of WGS in comparison to WES. We selected the best performing tools and attempted to optimize the CNV calling to improve precision. Finally, we suggest a combination of four tools (GATK gCNV, Lumpy, DELLY, and cn.MOPS) for a balanced CNV recall and precision.
2. Materials and Methods
2.1. Sequencing and Read Alignments
Genomic DNA (gDNA) was extracted from whole blood samples using a liquid handling automated station (Tecan, Männedorf, Switzerland). WES was performed from 200 ng of gDNA. Fragmentation was done on Covaris S2 (Agilent, Santa Clara, CA, USA) to approximately 300 base pair fragments and adaptor ligation was performed using KAPA HTP Library Preparation Kit. Exomes were enriched with SureSelectXT Clinical Research Exome kit (Agilent Technologies, Santa Clara, CA, USA). Paired-end sequencing with average read depth of at least 50× and 150 bp read length was performed using HiSeq2500 or NextSeq500 platforms from Illumina. For WGS analysis, sequencing libraries were prepared from 500 ng gDNA using Nextera DNA Flex library prep kit (Illumina, San Diego, CA, USA), according to the manufacturer’s instructions. WGS libraries were sequenced on Illumina NovaSeq6000 with sequencing depth of at least 30× and 150 bp read length. Sequenced reads were trimmed and aligned to the human reference genome (hg19/GRCh37) using BWA MEM v.0.7.12 software [
18].
2.2. Selection of Normals
CNVkit, CODEX2, ExomeDepth, and GATK gCNV require a group of samples to represent healthy control genomes. Normal samples should be produced with the same technical protocol, thus presenting a similar pattern of technical noise. Due to the nature of our experimental set up, no healthy control samples were available to create the Panel of Normals. Normal-like samples, concluded to have no CNVs by CytoScan HD SNP-array, were selected instead. Seventy whole genomes were chosen as WGS normals. Ninety-four exome samples were chosen as WES normals.
2.3. MLPA
Multiplex ligation-dependent probe amplification (MLPA) analysis was performed according to the manufacturer’s instructions (MRC-Holland, Amsterdam, the Netherlands) using appropriate MLPA Kits for BRCA1 (NM_00007294), BRCA2 (NM_000059), FLCN (NM_144997), MSH2 (NM_000251), MSH6 (NM_000179), PALB2 (NM_024675), PMS2 (NM_000535), VHL (NM_000551).
2.4. CytoScan HD SNP-Array
gDNA was isolated using the liquid handling automated station (Tecan). Purified DNA was quantified using the Qubit instrument (Life Technologies, Carlsbad, CA, USA). CytoScan HD SNP-array (Thermofisher Scientific, Waltham, MA, USA), which contains 2.67 million genome-wide markers, was performed on extracted DNA from samples GB01-GB38, according to the manufacturer’s instructions. Result files were analyzed using Nexus Copy Number software 10.0 (BioDiscovery, El Segundo, CA, USA) using NCBI Build 37 as reference. The samples were pre-processed by systematic correction (Quadratic), probes were recentered by the median and applying the mean of Combine Replicates Between Arrays. Subsequently, data were processed by SNP-FASST2 Segmentation with a significance threshold of 1.0 × 10−8 and max contiguous probe spacing of 1000 Kbp with a minimum of 3 probes per segment. The following thresholds were applied for calling CNVs: High Gain = 0.7; Gain = 0.23; Loss = −0.37; Big Loss = −1.1 and heterozygous imbalance threshold of 0.4. All gains or losses not covered by an allelic imbalance event were considered as false-positive and removed. Moreover, independently of the automatically generated CNV calls, each sample was visually inspected for CNVs using Nexus Copy Number software 10.0 (BioDiscovery, El Segundo, CA, USA).
2.5. NA12878 Gold Standard
Gold standard for NA12878 CNVs was produced by the 1000 Genomes Project. It contains only high confidence CNVs and the list of all CNVs was obtained from Haraksingh et al., 2017 [
8]. In total, 2076 CNVs of 51–453,313 bp sizes were used for CNV calling software evaluation. We have performed NA12878 WGS sequencing in-house while WES data was obtained from:
ftp-trace.ncbi.nih.gov/giab/ftp/data/NA12878/Garvan_NA12878_HG001_HiSeq_Exome/ (accessed on 1 April 2019).
2.6. CNVnator
CNVnator [
19] was run using the default parameters and recommendations from the authors with the bin size of 100 for all WGS samples.
2.7. CLC Genomics Benchmark
CLC Genomics Benchmark uses fastq files as input to the CNV calling workflow which are subsequently mapped to the reference genome by their internal read mapping tool [
20]. The mapped reads of the samples under investigation and the control samples are then used for calling CNVs. The called CNVs can be exported as BED files.
2.8. GATK gCNV
GATK gCNV [
21] calling is composed of two workflows: model creation and individual sample calling.
During model creation, the Panel of Normals of WGS and WES composed by 70 and 94 samples respectively, were generated. For WGS, intervals of 1000 bp and 0 bp padding were produced and filtered following GATK recommendations. No intervals were generated for WES samples. Read counts were measured in the exome regions and whole genome intervals and ploidy models were generated. Finally, a model per chromosome was produced using GermlineVariantCall. Benchmark samples were subsequently run, following the same procedure as the Panel of Normals, but including the available models during ploidy determination and germline variant call. Chromosomal calls were finally merged using PostprocessGermlineCNVCalls.
2.9. DELLY
Duplications and deletions were called by using DELLY [
22]. Only one library size was available, and it was provided at a time. Each of the samples only contained one read-group and no further specifications were given. The variant call was performed searching for duplication events (−t DUP) and deletions (−t DEL). The resulting bcf files were merged using bcftools concat.
2.10. cn.MOPS
cn.MOPS [
23] was used to call CNVs on all WES and WGS samples following the vignette [
24]. Due to memory constraints, WGS sample bam files were grouped together in batches of 11 to 12 samples before analysis, and each batch was run as follows. Using the provided getReadCountsFromBAM function, read counts were extracted from the bam files for chromosomes 1–22 using a window length parameter WL = 500 in order to achieve roughly 50–100 reads per window. The cn.mops function was used to call CNVs and integer copy numbers were extracted using calcIntegerCopyNumbers.
For the WES samples, all bam files were run together as a single batch. A bed file with the targeted regions was converted into GRanges format and used to extract read counts in the regions with the getSegmentReadCountsFromBAM function. CNVs were then called using the exome cn.mops function and integer copy numbers calculated as above.
2.11. CNVkit
CNVkit [
25] was used to call CNVs on all WES and WGS samples, grouped together by library type and according to the authors guidelines [
26]. Briefly, we used the default parameters, restricting the analyses to the WGS mappable regions and, in the case of WES, to the captured regions also.
2.12. Control-FREEC
Control-FREEC 11.5 [
27,
28] was used to process the 39 WGS samples of the benchmark starting from BAM files. Default parameter values (forceGCcontentNormalization = 0, minCNAlength = 1, coefficientOfVariation = 0.05) were used, however, ploidy was set to 2, mateOrientation to “FR” for Illumina paired-end reads, and the sex of each sample was supplied (two males and seven females). In this case, the hg19 reference used for alignment was used and mappability of the genome was disregarded (read length = 151). Sambamba 0.6.7 [
29], BEDTools 2.27.1 [
30], Samtools 1.9 [
31,
32], and R 3.5.0 [
33] were used as dependencies. The script ‘assess_significance.R’ provided by the Control-FREEC developers was used to add
p-values to the detected CNVs.
2.13. Manta
Manta [
34] was used to infer deletions and duplications from both WES and WGS sequence data using default parameters.
2.14. LUMPY
For each of our WGS sample bam files, we used the LUMPY Express wrapper [
35] to call CNVs as described in the official documentation on GitHub (
https://github.com/arq5x/lumpy-sv, accessed on 1 November 2021): discordants were extracted with samtools view, filtering out reads by flag 1294; split reads were extracted using the provided extractSplitReads_BwaMem script. Both were sorted and then provided as input to the lumpyexpress utility together with the original bam to output a vcf giving structural variants, from which we obtained CNVs as the called deletions and duplications.
2.15. ExomeDepth
ExomeDepth [
36] was used to detect CNVs from WES sequence data following the authors recommended best practices [
37]. Since ExomeDepth takes advantage of the tight correlation structures between large numbers of samples when building a reference sample, we used our reference samples as a Panel of Normals. Even though these reference samples are not ideal (since they are not from the same batch), they are a good representation of the samples in our laboratory, as well as the experimental design most users will encounter (related samples from the same lab, but not the same batch).
2.16. CODEX2
To run CODEX2 [
38], we grouped all the WES samples together with 50 extra Panel of Normals WES samples. This batch of samples was then analyzed for each of chromosomes 1 through 22 by following the provided documentation from GitHub (
https://github.com/yuchaojiang/CODEX2, accessed on 1 November 2021), using default or suggested parameters and functions except where otherwise noted. We used CODEX2 without specifying negative control samples, using the normalize_null function for normalization. For chromosomes 1, 4, 6, and 14, the glm.fit procedure in CODEX2 did not converge, despite increasing the parameter K = 1:10 as per the author’s suggestion.
2.17. Data Processing and Plotting
All tools were run using Snakemake [
39], post-processing of the called CNVs was carried out using Python 3.6 and R 3.5.0 [
33]. Scripts for running the tools are available on GitHub (
https://github.com/cphgeno/CNVbench, accessed on 1 November 2021). The rtracklayer package [
40] was used for main processing of the files, and ggplot2 [
41], ComplexHeatmap [
42], and RColorBrewer [
43] were the key resources for plotting.
2.18. Fraction Curve Generation
The analyzed data was split into four data sets: NA12878 WES, GB01-GB08 WES, NA12878 WGS, and GB01-GB38 WES. For each data set, and for each tool that called CNVs for the dataset in question, we filtered the CNV calls according to a confidence metric provided by the tool itself (see below). Specifically, we subset the tool data set on every confidence metric percentile from 0 to 99 by taking all CNV calls with confidence scores less/greater (as appropriate for the metric) than or equal to a given percentile. Recall and precision were then calculated and plotted for the resulting, filtered CNV calls.
Note that some tools use only a few discrete confidence values or use a single value for a large proportion of the calls. Therefore, a cutoff at the 99th percentile does not necessarily contain 1% of the called CNVs if, for example, 50% of the CNVs in the call set are assigned the best confidence score.
The confidence metrics used for each tool were as follows. CLC Genomics Workbench: Absolute fold change; CNVkit: Mean squared standard error of log2 of the copy number; CNVnator: t-statistic p-value; cn.MOPS: Median informative/non-informative ratio value; CODEX2: Likelihood ratio; ControlFREEC: Wilcoxon rank sum test p-value; DELLY: Genotype quality values; ExomeDepth: Observed/expected read ratio; GATK gCNV: CNQ scores (difference between the two best genotype Phred-scaled log posteriors); Lumpy: Number of pieces of evidence supporting the variant across all samples; Manta: CNV quality score.
2.19. Performance Profiling
Snakemake’s built-in capabilities for benchmarking runtime and memory usage were used to measure wall-clock time and peak resident set size for calling CNVs on sample NA12878. Tools were tested on a HP Apollo 6000 System ProLiant XL230a Gen9 Server blade (Hewlett-Packard, Palo Alto, CA, USA), on a node with 28 64-bit Intel Xeon E5-2683 v3 @2.00 GHz CPUs available, and 128 GB, DDR4 @2133 MHz RAM.
3. Results
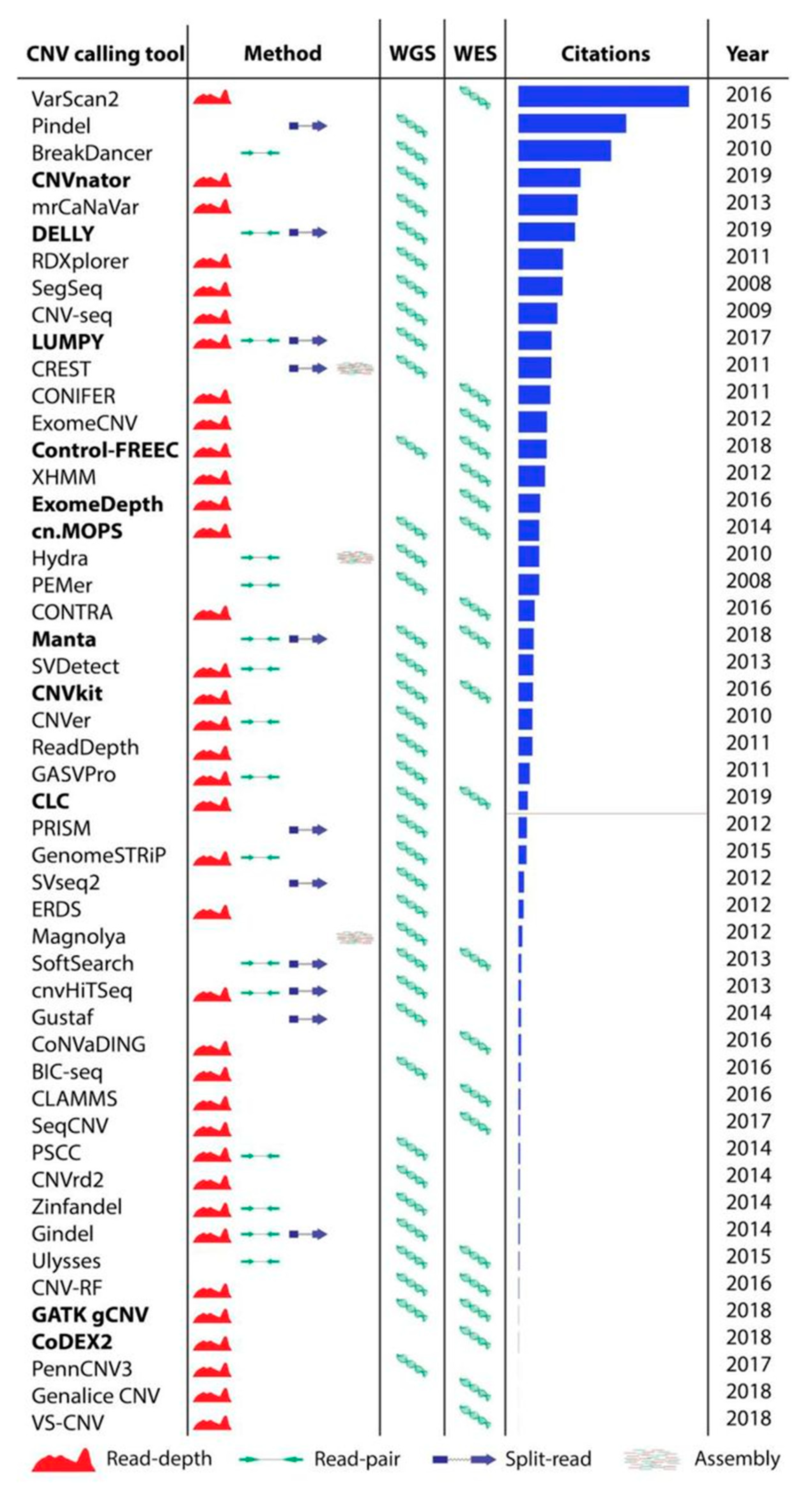
3.1. Review of 50 Most Popular CNV Calling Tools
We reviewed 50 most popular tools for CNV calling (
Figure 2,
Table S1). The tools were included in the benchmark if they were: (1) developed for calling CNVs from WES or WGS data, (2) developed for germline CNV calling, (3) recently developed or highly cited (>100 citations as of March, 2019, using the number of citations as the only available proxy for popularity of use), (4) still maintained, and (5) the latest versions of the tool was more recent than 5 years. However, several tools passing all these criteria were not suitable for inclusion in this benchmark (detailed information in
Table S1). After applying the selection criteria, 11 tools (in bold in
Figure 2) were selected for further performance evaluation.
3.2. Datasets Used for the Benchmark Study
The datasets used for the benchmark are listed in
Table 1. Gold Standard sample NA12878 from 1000 Genomes Project was used with the CNVs which were published by Haraksingh et al., 2017 [
8]. For the in-house GB01-GB08 and GB09-GB38 samples the true reference was considered to be Nexus software-produced filtered CNV calls from CytoScan HD SNP-array, which have previously been shown to be among the best-performing array platforms [
8,
44]. To account for imperfections in the SNP-array CNV calling, we compared all CNV calls made by different CNV calling tools. Furthermore, selected CNVs were confirmed for the GB40-GB51 samples using multiplex ligation-dependent probe amplification (MLPA).
3.3. CNV Calling Tools Included in the Benchmark
We selected 11 tools for the benchmark. Eight tools use a read depth approach: CNVnator [
19], CLC Genomics Workbench [
45], GATK gCNV [
21], cn.MOPS [
23], ExomeDepth [
36], CNVkit [
25], CoDEX2 [
38], and Control-FREEC [
27]. Other SV callers that include CNVs, such as DELLY [
22], Manta [
34], and LUMPY [
35] use a combined approach and apply more than one CNV calling algorithm for more accurate predictions (
Figure 2; detailed tool algorithms are provided in
Supplementary File S1). All tools were run using the default parameters and following author recommendations when available. Most CNV calling tools are developed for either WES or WGS. However, cn.MOPS, CLC Genomics Workbench, CNVkit, Manta, and GATK gCNV tools are capable of calling germline CNVs using both WES and WGS data. For the NA12878 Gold Standard sample we also report CNVs called by Haraksingh et al., 2017 [
8] using the consensus of several methods, including non-NGS based approaches.
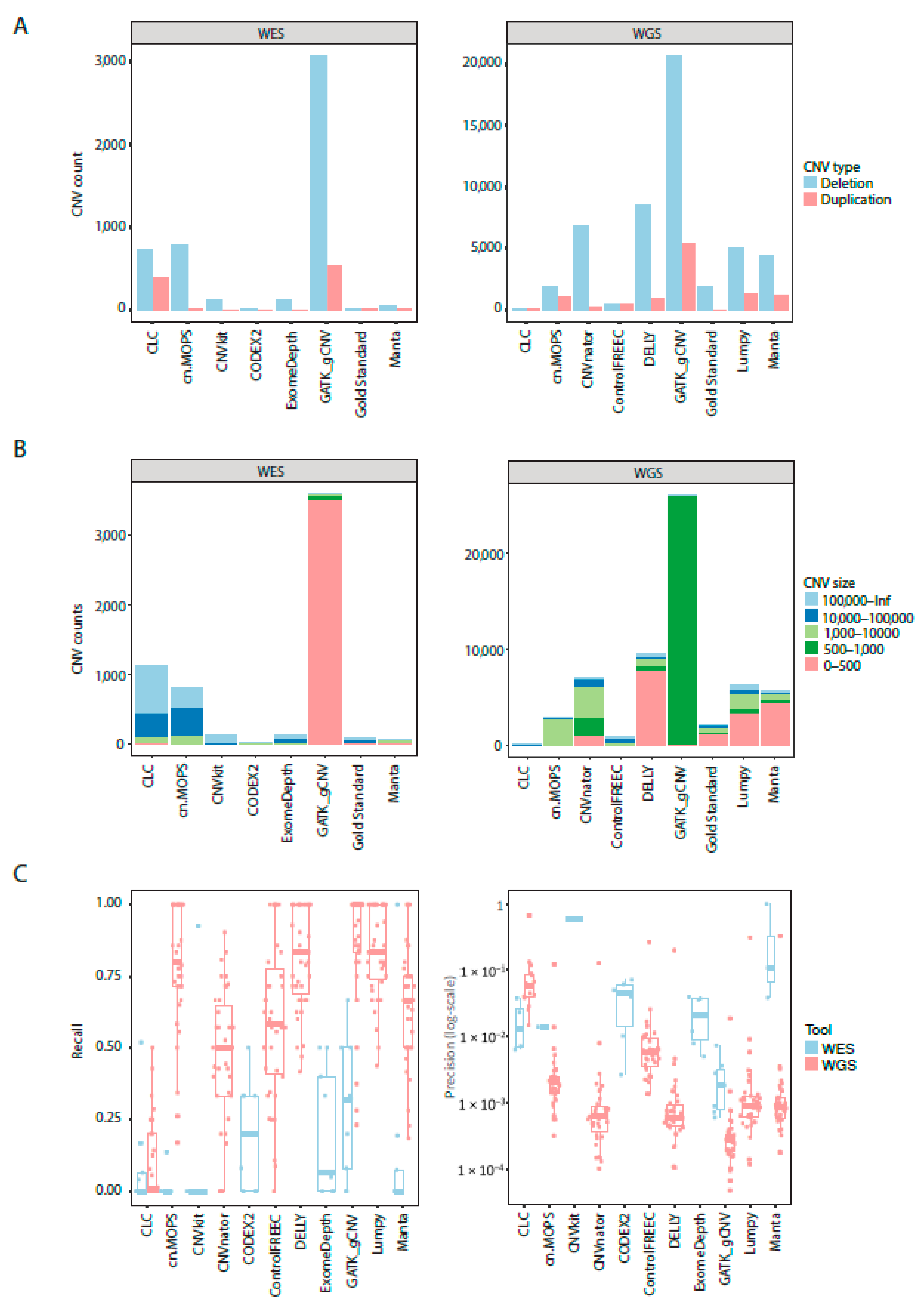
3.4. CNV Length and Type Distribution for CNV Calling Tools
All tools called more deletions than duplications for NA12878 Gold Standard sample (
Figure 3A). However, the total number of called CNVs varied greatly between the tools. GATK gCNV called more duplications and deletions in both WES and WGS samples compared to other tools. CODEX2 called the lowest number of deletions and duplications in WES samples and CLC Genomics Workbench called the lowest number of CNVs in WGS data. None of the CNV calls were filtered with a cutoff on confidence metrics, except where it was recommended by the authors of the tool (CODEX2) or the filtered files were created automatically (CLC Genomics Workbench). CODEX2 called a similar number of CNVs as provided in NA12878 Gold Standard true CNV set, but this did not equate that all the CNV calls were true positives. Similar patterns were observed for GB01-GB38 samples (
Figure S1).
CNV calls differed in lengths and frequencies among the tools in WES and WGS of NA12878 sample (
Figure 3B). CLC Genomics Workbench and cn.MOPS called a high number of CNVs longer than 10,000 bp while GATK gCNV called mainly CNVs shorter than 500 bp in WES and 500–1000 bp in WGS. GATK gCNV called shorter CNVs than any other tool. Furthermore, cn.MOPS, CNVnator, and Control-FREEC predicted more >1000 bp length CNVs than other tools for WGS NA12878 sample. Half of CNVs in NA12878 were shorter than 500 bp as per Gold Standard truth CNV set. Similar patterns were observed for GB01-GB08 WES and GB01-GB38 WGS samples (
Figure S2).
3.5. Precision and Recall of CNV Calling Tools
Given CNVs from CytoScan HD SNP-array for GB01-GB38 samples and NA12878 Gold Standard truth CNV set, true positive (TP), true negative (TN), false positive (FP), and false negative (FN) CNV calls were identified for each of 39 samples for WGS, and 9 samples (GB01-GB08 and NA12878) for WES (
Table S2). The criteria of an overlap of 1 bp between the Cytoscan HD SNP-array called CNV and the NGS-based tool CNV call was used for the CNV call to be classified as a TP. Recall and precision were calculated using the following formulas:
GB01-GB38 samples had a total 2–103 (median 7) CNVs called by CytoScan HD SNP-array, whereas 2076 CNVs were called for NA12878 from the Gold Standard truth CNV set. For WES data, only CNVs covering exons were considered (1–63 (median 4) for GB01-GB08 and 233 for NA12878).
GATK gCNV recall was best for both WES and WGS data (
Figure 3C), followed by Lumpy, DELLY, cn.MOPS, and Manta. All tools performed poorly on the WES dataset by having lower levels of recall compared to WGS. While recall for WGS in all tools, except CLC Genomics Benchmark, was fair, precision was lacking for all the tools, with a maximum precision of 66.7%. Tools that called a higher total number of CNVs, also had higher recall, but lower precision. The only two tools which use CNV call filtering (CLC Genomics Workbench and CODEX2) had a low recall compared to the tools which did not filter CNVs as part of their default settings. Collectively, recall approached 1 for several tools, but came at the expense of precision, which was lower than 31% in WGS data for the four best recalling tools (GATK gCNV, Lumpy, DELLY, and cn.MOPS).
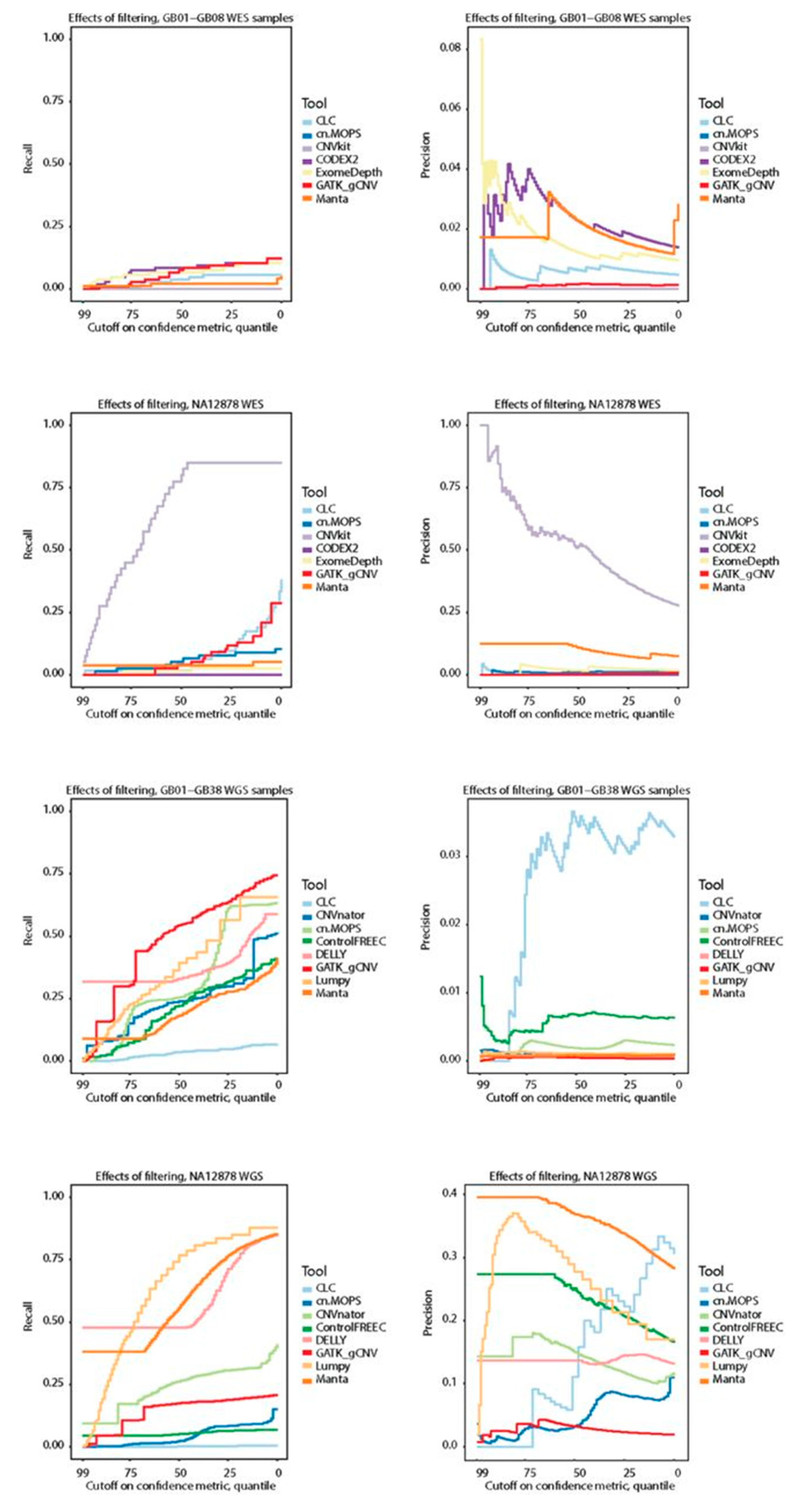
3.6. CNV Call Filtering Possibilities for CNV Calling Tools
To explore the possibility of filtering CNV calls to improve precision we analyzed recall and precision at sliding confidence cutoff values. Briefly, for each tool, we calculated recall and precision at different thresholds defined by the percentiles of the tool-defined confidence metric (see
Section 2 for details).
The recall on WES was low (ranging from 0 to 0.65,
Figure 3C), regardless of filtering, and that precision generally did not improve as a function of the threshold, and could not be easily interpreted as being asymptotically negative (
Figure 4). The exception to this was CNVkit, which displayed high recall and precision on the WES NA12878 sample and offered the possibility of meaningful filtering.
However, this performance of CNVkit was only seen for NA12878 sample and not for the remaining WES samples.
For WGS, recall for most tools decreased linearly as a function of filtering, the utility of which is therefore limited. Arguably, one exception for the GB01-GB38 WGS samples was GATK gCNV, which had proportionally good recall when filtering out the bottom 75th percentile. However, such a cutoff did not improve the precision for the tool. For the WGS NA12878 sample, DELLY and Manta had good performance when selecting only the over-represented top confidence values, while Lumpy also displayed diminishing returns at lower scores. In the case of Manta, precision decreased predictably with filtering, while Lumpy had a somewhat unpredictable precision fraction curve, and DELLY’s precision appeared unaffected by filtering. CLC Genomics Workbench had repeated patterns for recall in both NA12878 and GB01-GB38 suggesting a relative ranking metric for each run. Collectively, sorting CNVs on confidence metrics from the tools did not offer any meaningful threshold for controlling precision, due to asymptotically positive recall curves (more liberal inclusion resulted in more hits). We further found several unpredictable precision curves, and overuse of the maximum confidence value, which gave a percentile of top-scoring CNVs without any additional metrics to rank. Exceptions to these conclusions were found only for the NA12878 sample.
3.7. Short CNVs Can Be Identified by NGS-Based CNV Calling Tools
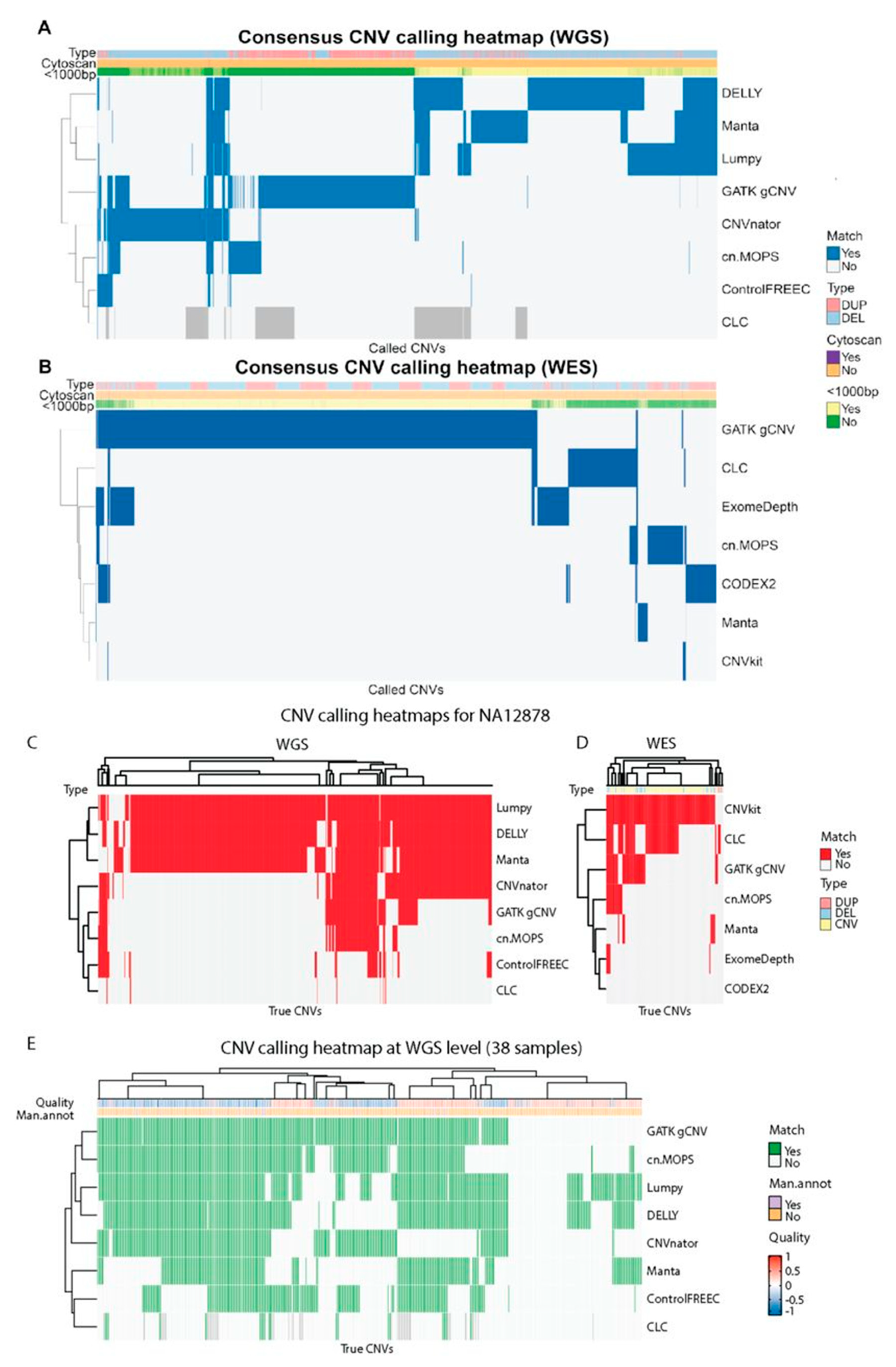
For the 38 WGS samples, DELLY and GATK gCNV called the most CNVs: 148,519 and 132,265, respectively. Manta, Lumpy, CNVnator, cn.MOPS, and Control-FREEC called 94,832, 93,166, 85,962, 36,491, and 13,160, respectively. CLC Genomics Workbench called 632 CNVs across the 29 samples it was run for. The tool was not run on all 38 samples due to computation time: it re-analyzes the base-level coverages of the control samples in every run, resulting in very long running times for WGS samples.
Tools with the same calling strategies had a higher overlap in called CNVs (
Figure 5A). For example, DELLY, Lumpy, and Manta displayed a large degree of CNV overlap in WGS samples and they all use RP and SR information for calling CNVs. CNVnator, GATK gCNV, and DELLY called a high number of unique CNVs in WGS data which were not called by any other tool. Furthermore, a total 51.9% of all called CNVs were shorter than 1000 bp. CNVs which were called by two or more tools were mostly short: less than 1000 bp. Such CNVs are known to be less often called by array-based CNV calling approaches [
8,
46]. Out of 407,671 CNVs called in the WGS samples, 74.4% were called only by a single tool. The percentages of CNVs called by 2–8 tools were 11.5, 9.5, 2.6, 0.8, 0.8, 0.4, and 0.1%, respectively (
Figure 5A).
For WES (
Figure 5B), CLC Genomics Workbench called 1268 CNVs, cn.MOPS-787, CNVkit-66, CODEX2-762, ExomeDepth-1123, GATK gCNV-7116, and Manta-174. The observed overlap between CNVs called between tools was lower than for WGS data: 90.3% of the total 9944 CNVs were called by a single tool, and the percentages for 2–7 tools were 6.8, 2.0, 0.6, 0.2, 0.04, and 0%, respectively. GATK gCNV called the highest number of unique CNVs while the CNV calls by two or more tools were mostly longer than 1000 bp. Despite the overlap between longer CNV calls, the majority (69.6%) of all CNV calls were shorter than 1000 bp.
Lumpy, DELLY, Manta, and partly CNVnator performed best on NA12878 WGS data while CNVkit recalled almost all CNVs present in NA12878 WES dataset (
Figure 5C,D). It is important to note that Manta and CNVnator were used for the generation of the NA12878 truth CNV set [
8] and the CNV calls might have been favored to overfitting. A more accurate picture of the tool’s performance can be obtained by evaluation of GB01-GB38 CNV calls.
Many CNVs confirmed by CytoScan HD SNP-array on WGS were called by multiple tools (
Figure 5E). While 25.6% of all called CNVs overlapped in two or more tools (
Figure 5A), more than 83.7% CNVs were overlapping with the CytoScan HD-confirmed CNV list (
Figure 5E). As for WES, in more than two thirds of the cases where CytoScan identified a CNV, none of the tools called it. All tools performed similarly poorly on WES data (
Figure 6) with cn.MOPS and CNVkit missing all the CNVs identified by CytoScan HD SNP-array.
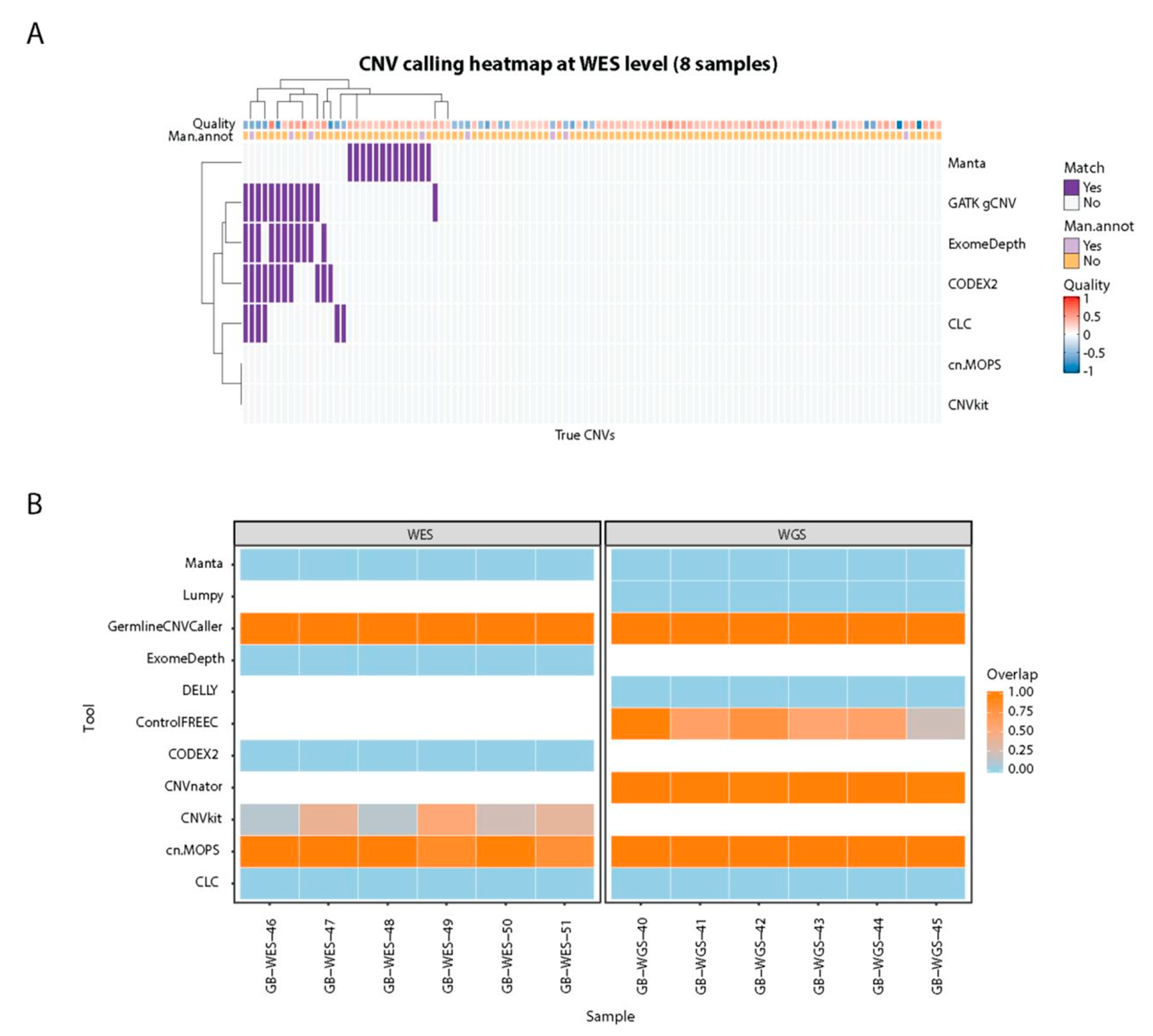
3.8. MLPA-Confirmed CNV Recall for CNV Calling Tools
To assess if tools accurately identify CNV breakends we used twelve MLPA-confirmed CNVs of varying sizes (1 exon to whole gene; deletions (
n = 11) or duplications (
n = 1),
Table S3) from six WES samples and six WGS samples (
Figure 6B). Six out of 11 tools (CLC Genomics Workbench, CODEX2, DELLY, ExomeDepth, Lumpy, and Manta) did not identify any of the MLPA-confirmed CNVs. Conversely, GATK gCNV, CNVkit and cn.MOPS identified all MLPA-confirmed CNVs in WES. GATK gCNV, cn.MOPS, CNVnator, and Control-FREEC identified all the CNVs confirmed by MLPA in WGS samples. CNVnator (WGS), cn.MOPS (WES), and CNVkit (WES) predicted shorter CNVs than the MLPA-defined truth, while GATK gCNV identified the full-length of both deleted and duplicated regions. Control-FREEC called all 6 CNVs in WGS samples but predicted shorter CNVs spanning 22.0–98.6% of true CNV length.
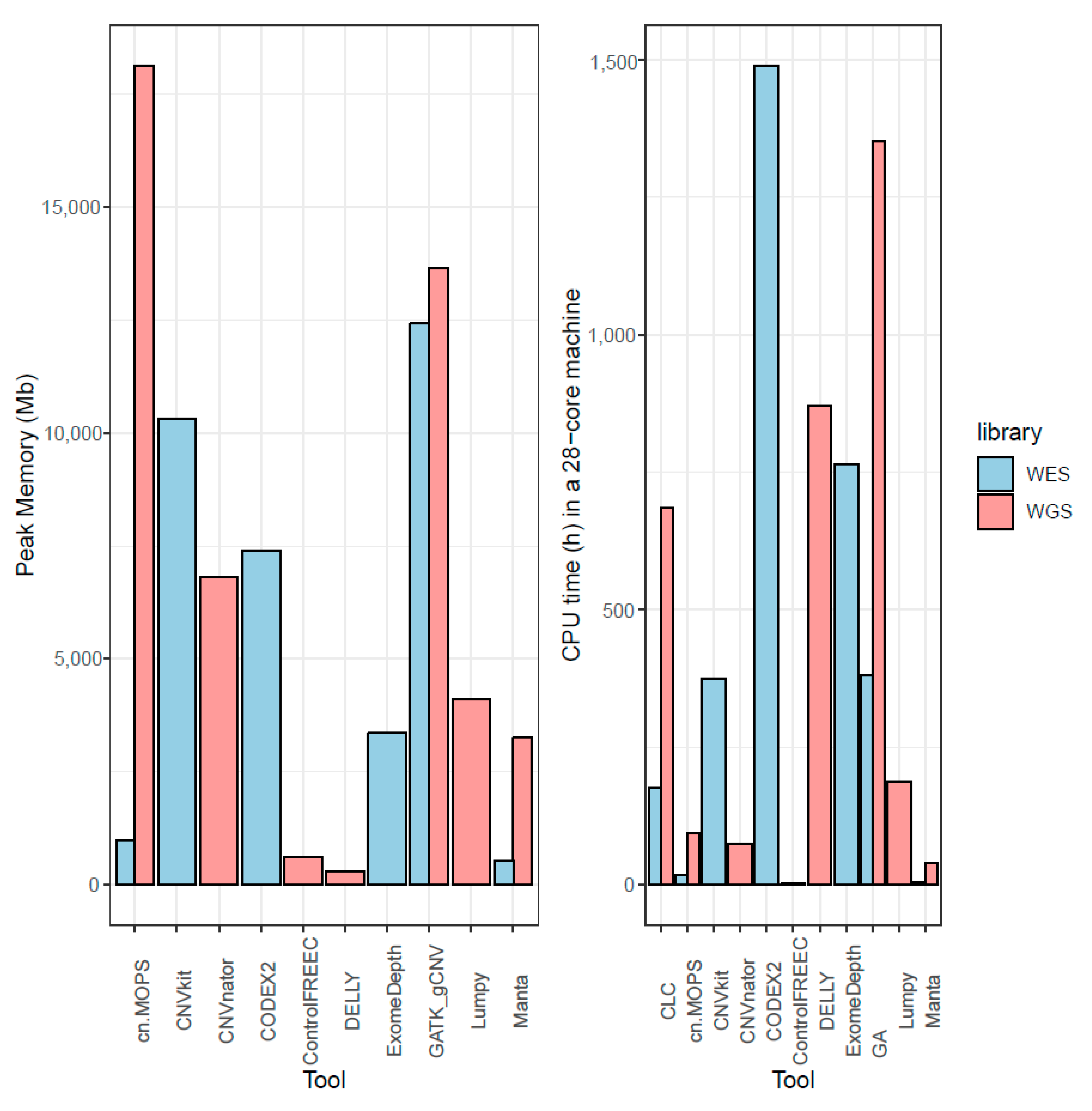
3.9. Memory and CPU Requirements for CNV Calling Tools
CPU and memory requirements were measured on a 28-core server grade cluster node, for the tools where it was possible to obtain an estimation on the NA12878 sample. Control-FREEC showed the best compromise between memory and CPU, both being low in WGS and even lower than requirements for other tools in WES (
Figure 7). Memory-wise, DELLY and Manta were the other two tools with the lowest needs; the latter also having short computational times, while the former had one of the highest, possibly due to the fact that insertions and deletions were called subsequently and not in parallel. Surprisingly, cn.MOPS also showed low memory requirements on exomes, but the highest in genomes. However, it also offered one of the lowest computational times both in WES and WGS.
GATK gCNV and cn.MOPS used a lot of RAM at peak memory, and it is possible that more RAM per node than the available 128 GB per machine would have shortened the runtime by enabling better distribution of tasks. Computational requirements for creation of GATK model were not measured in this benchmark. Due to the batch caller nature of cn.MOPS, CNVkit, and CODEX2, many alignments have to be kept in memory at the same time, explaining the observed higher memory requirements.
4. Discussion
The results from this study contribute to the growing body of knowledge focused on the evaluation of structural variant characterization tools. Specifically, from the reviewed 50 CNV calling tools, we observed that many of the tools were either not maintained with the last updates applied more than 5 years ago, or not widely used. We included 11 widely used or newly developed CNV calling tools, which fulfilled our selection criteria, to benchmark their performance on CNV calling. Unlike previous studies focused on either a single sample or limited number of samples [
9,
11,
15,
16,
17], this study has benchmarked the performance of various SV callers on both WES and WGS data originating from multiple samples and evaluated their overall performance.
In order to establish a reference set of CNVs we used the best performing array-platform, in addition to NGS. CytoScan HD SNP-array technology was chosen as a sensitive and clinically adopted method to detect CNVs. It is important to note that because of the probe-dependent and genome-distributed nature of the array technology, not all short CNVs could be captured.
Therefore, the CNV calls classified as false positive (FP) in this benchmark should be interpreted carefully. Additionally, as CNV detection is a technically challenging task, none of the array-based standards in this study can ultimately be regarded as an absolute truth [
8]. Besides the in-house GB01-GB38 samples, which were analyzed by Cytoscan HD SNP-array, we included the well-studied NA12878 sample, for which extensive efforts have been made to confirm all CNVs, based on several platforms, and NGS-based CNV callers. The latter might introduce a bias for these samples in this benchmark, as two of the included tools were used in this evaluation (Manta and CNVnator). Furthermore, the NA12878 sample and its truth CNV set are also popular for testing and optimizing CNV tools, which could potentially explain the possible overfitting we observed e.g., for Manta and CNVkit, which had the highest discrepancy between NA12878 calls and calls on our cohort.
Tools with identical CNV calling strategy had a tendency to call the same CNVs, and, in general, read-depth based tools, or combinations including this strategy, performed best, when assessed on recall of CNVs from Cytoscan HP SNP-array, or NA12878 Gold Standard sample. The number of CNVs called varied more than a 100-fold; consequently, the recall rates for tools calling many CNVs were higher, and no systematic trade-off could be found to improve precision for these tools. In short, tools calling many CNVs hit the target more often, but high confidence CNVs were not generally showing a higher fraction of recall. For tools like DELLY and Lumpy, a combination of CNV metrics could be used to filter on the CNV calls as it is applied in SVTyper [
47].
CNVs selected for experimental validation with MLPA were selected based on targeted gene panel sequencing, and were, therefore, not biased by CNV calls from tools tested in this analysis. It was, however, striking that tools could be split into two groups: those that were able to recall all six independent CNVs and those that called none.
GATK gCNV caller performed best at CNV recall and is clearly the most sensitive tool for CNV identification for both WES and WGS data, but comes with poor precision, like all tools tested (highest precision mean <13% for WES and <6% for WGS). GATK gCNV is also the best performing tool when recalling MLPA-confirmed CNVs and estimating their breakends correctly, even if four other tools also recalled the CNVs. The good performance of GATK gCNV and cn.MOPS caller comes at a high computational cost and the former was almost twice as computationally expensive as the third-highest consuming WGS tool considering CPU/h and peak RAM usage.
The GnomAD database [
14] shows how CNV calls can be used clinically, but more research and larger cohort studies are needed for better annotation and inference of causation of CNVs. Our study shows that more work has to be done on collecting large and well-annotated datasets with CNV detection on several platforms, in order to drive the development of tools with improved precision on CNV calling from NGS data. The current state of tools for finding CNVs is suited for identifying complex traits in large cohorts, for which we suggest to use the overlap between several tools. Using rare CNVs called from NGS as a basis for genome-wide association studies is not currently advisable.
The future for NGS-based CNV calling tools is likely to rely on the utilization of a combination of long- and short-read sequencing [
48]. This is particularly true considering the need for CNV annotation that explains causative traits and which will require sequencing of large cohorts with two simultaneous protocols. Alternatively, future improvements on both price and error rate for long-read sequencing are needed. In terms of filtering CNV calling results, better annotation of CNVs is a clear optimization candidate. The ability to leverage additional genetic data, such as RNA-seq, or even static knowledge of genetic sites from associations to epigenetic mechanisms or regulation, may also guide the selection and prioritization process in the near future. In a clinical setting, production of background panels or databases to filter true common CNVs or common FPs called by each tool can greatly reduce the number of relevant CNVs presented for interpretation (data not shown), just like databases like gnomAD SV [
49] can be used to reduce the numbers of common CNVs. Although beyond the scope of this work, another interesting area that requires further investigation is how different CNV calling tools perform based on various SV sizes and read coverage, both of which are known to affect detection and accuracy of SV calling [
15,
16,
50]. In a similar manner, the distributions of SVs in biological regions (e.g., sex chromosomes) may require special attention, as specific SVs have been known to occur in a particular gender (e.g., [
51,
52]). Lastly, transcriptional regulation of the altered regions requires more investigations, so that the causative effect of CNVs can be elucidated, and potentially be predicted in each case. Our work has several limitations. First, we benchmarked only a limited set of tools; however, findings are in line with larger studies [
15], relying on single truth sets. Furthermore, the observed potential for overfitting to NA12878 Gold Standard sample by some tools complicated the accurate evaluation of recall and precision with a well-annotated dataset. Some tools (CNVkit, CODEX2, ExomeDepth and GATK gCNV) require training on a known dataset before application and we did not attempt to optimize this step or evaluate what influence it has on the tool’s performance since it is outside of the scope of this work. Moreover, we used whole genomes and whole exomes for WGS and WES sequencing respectively without any additional filtering on GC content or read mappability. Finally, the main limitation of our work is the lack of well-defined true CNV sets, therefore our analysis using CytoScan HD SNP-array calls vastly underestimates CNV call precision on the in-house data sets, but this caveat should not favor specific tools.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}