
Pyridazinone Derivatives Limit Osteosarcoma-Cells Growth In Vitro and In Vivo

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Reagents
2.2. Compounds Preparation
2.3. Cell Lines
2.4. Mitochondrial Activity
2.5. cAMP Measurement
2.6. Protein Quantification
2.7. Cell Count and Morphological Parameters
2.8. DNA Quantification
2.9. Cell Proliferation
2.10. Apoptosis
2.10.1. AnnexinV/Pi Staining
2.10.2. Caspase-3 Activity
2.11. Senescence
2.12. MMPs Activity
2.13. Cell Migration
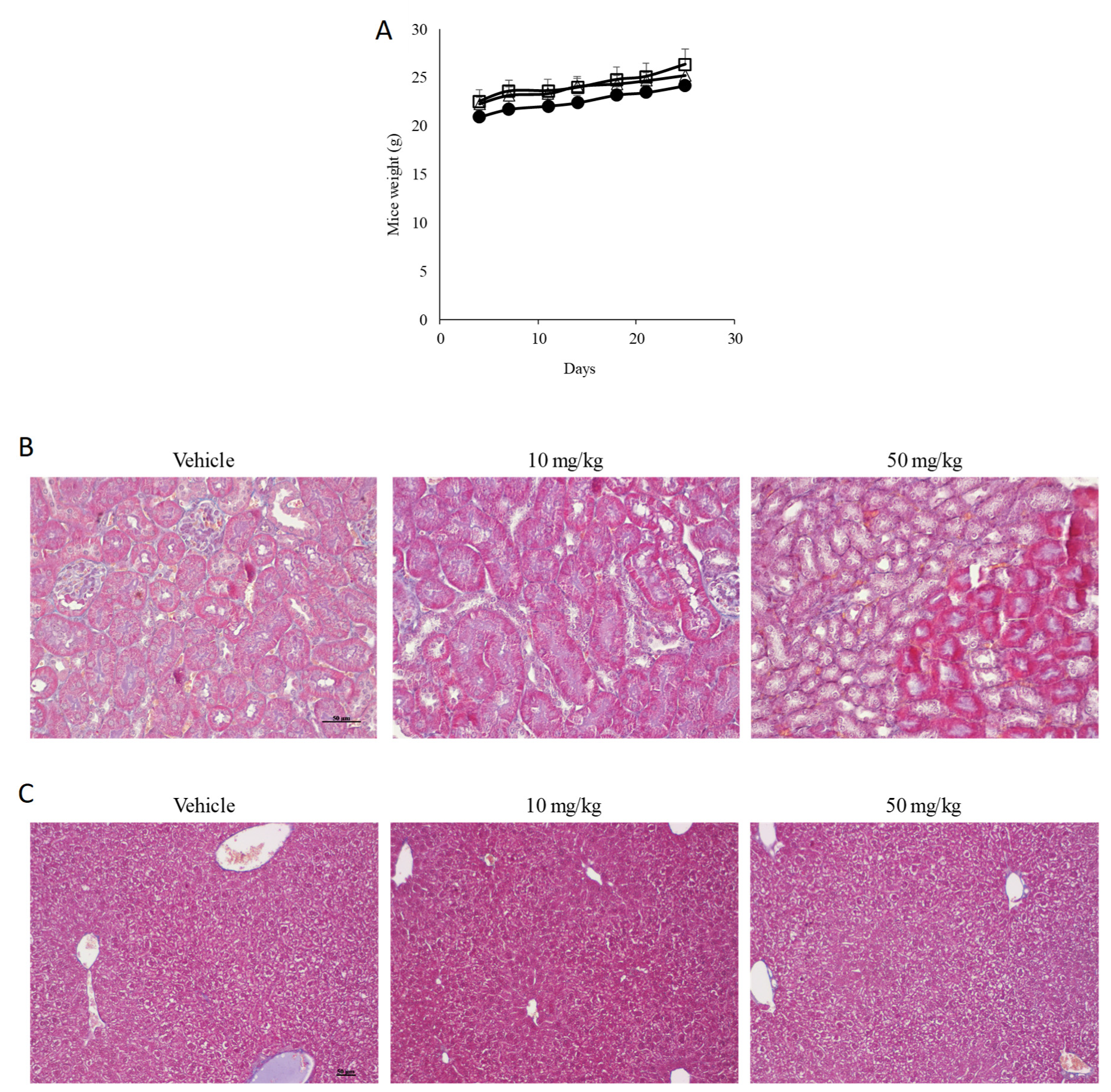
2.14. In Vivo Experiments
2.15. Histology
2.16. µCT Analysis
2.17. Graphical Representation of Data and Statistical Analysis
3. Results
3.1. Pyridazinone Scaffold-Based Compounds Decrease Human Osteosarcoma Cell Line Metabolic Activity
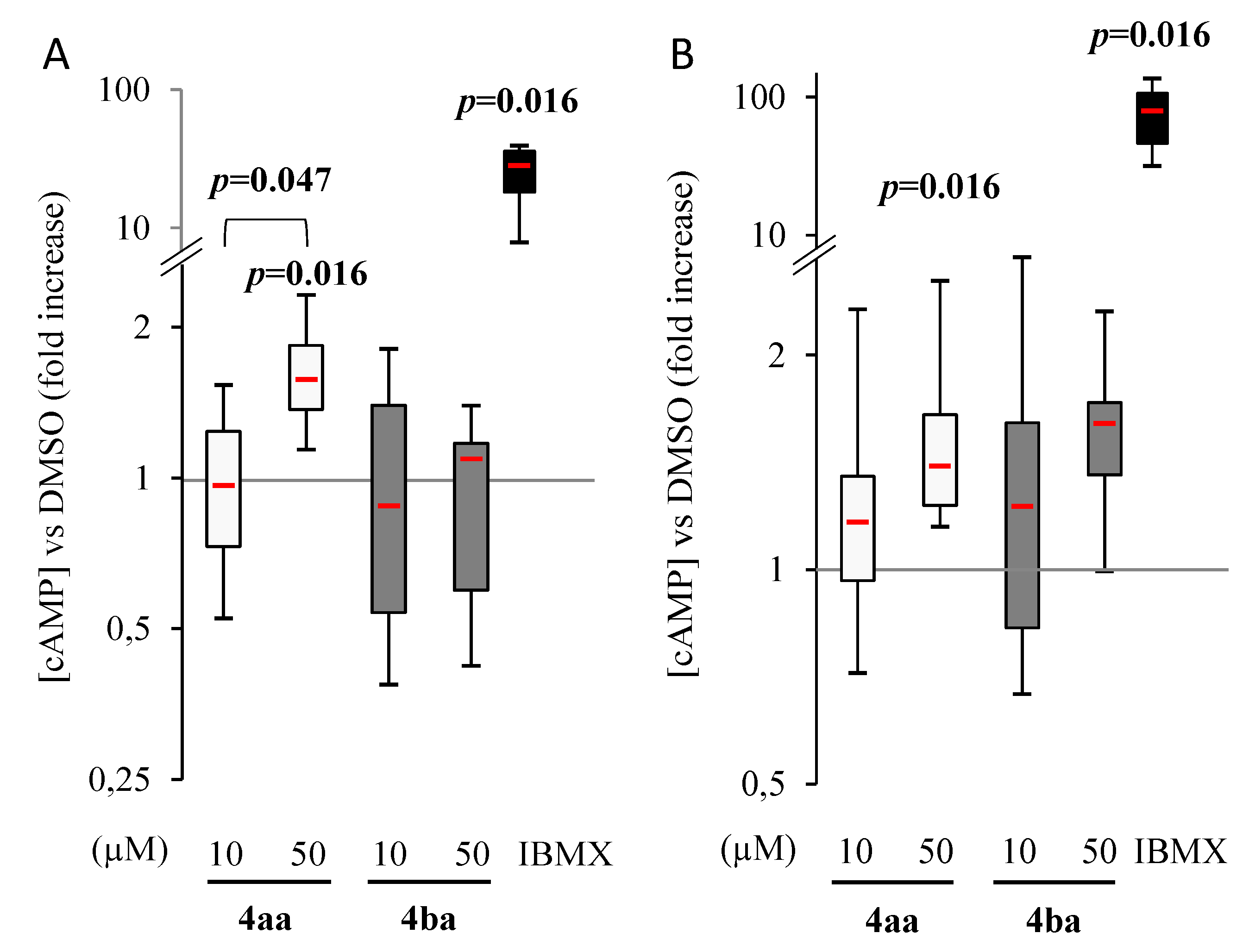
3.2. Pyridazinone Scaffold-Based Compounds Increase Intracellular cAMP
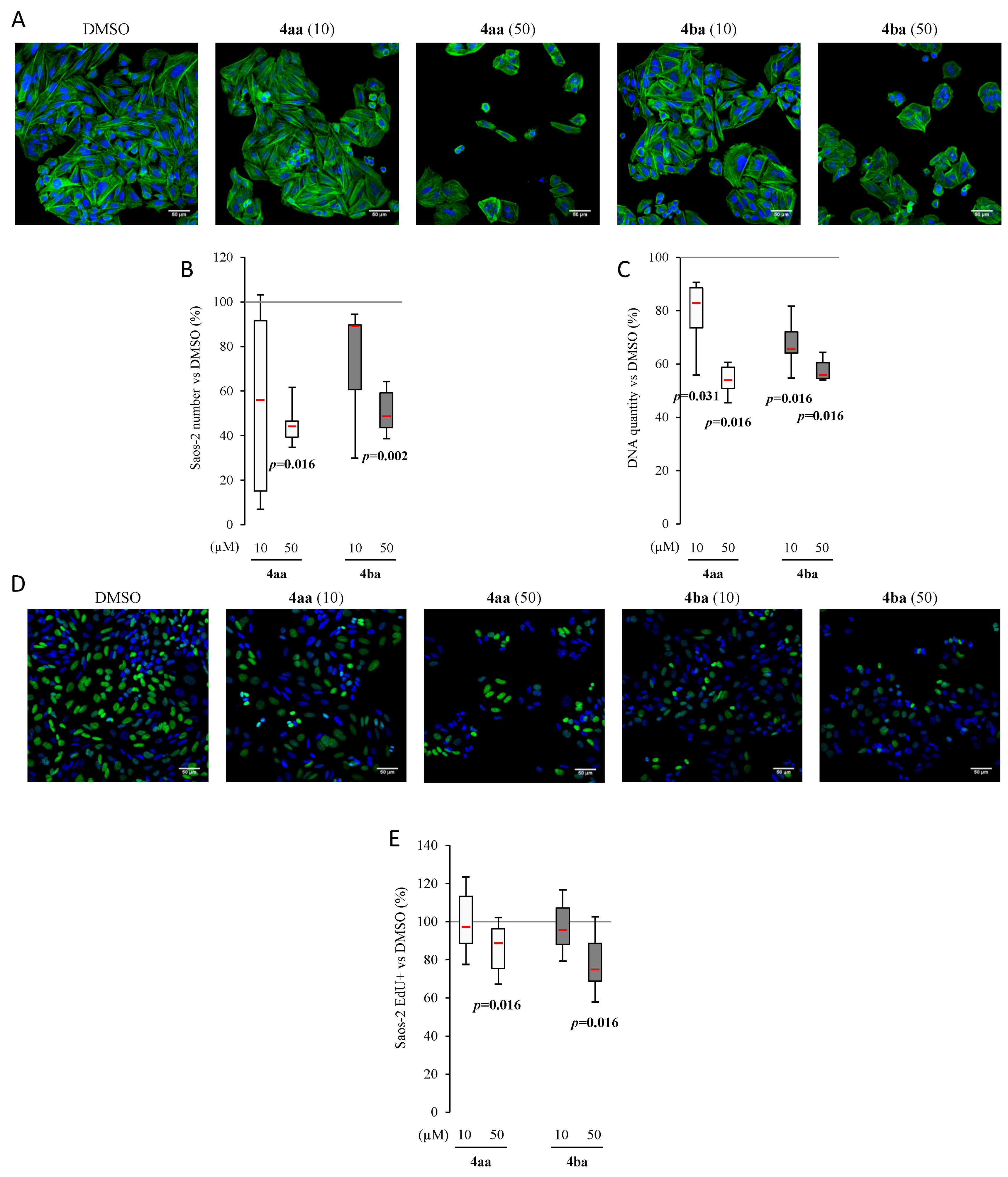
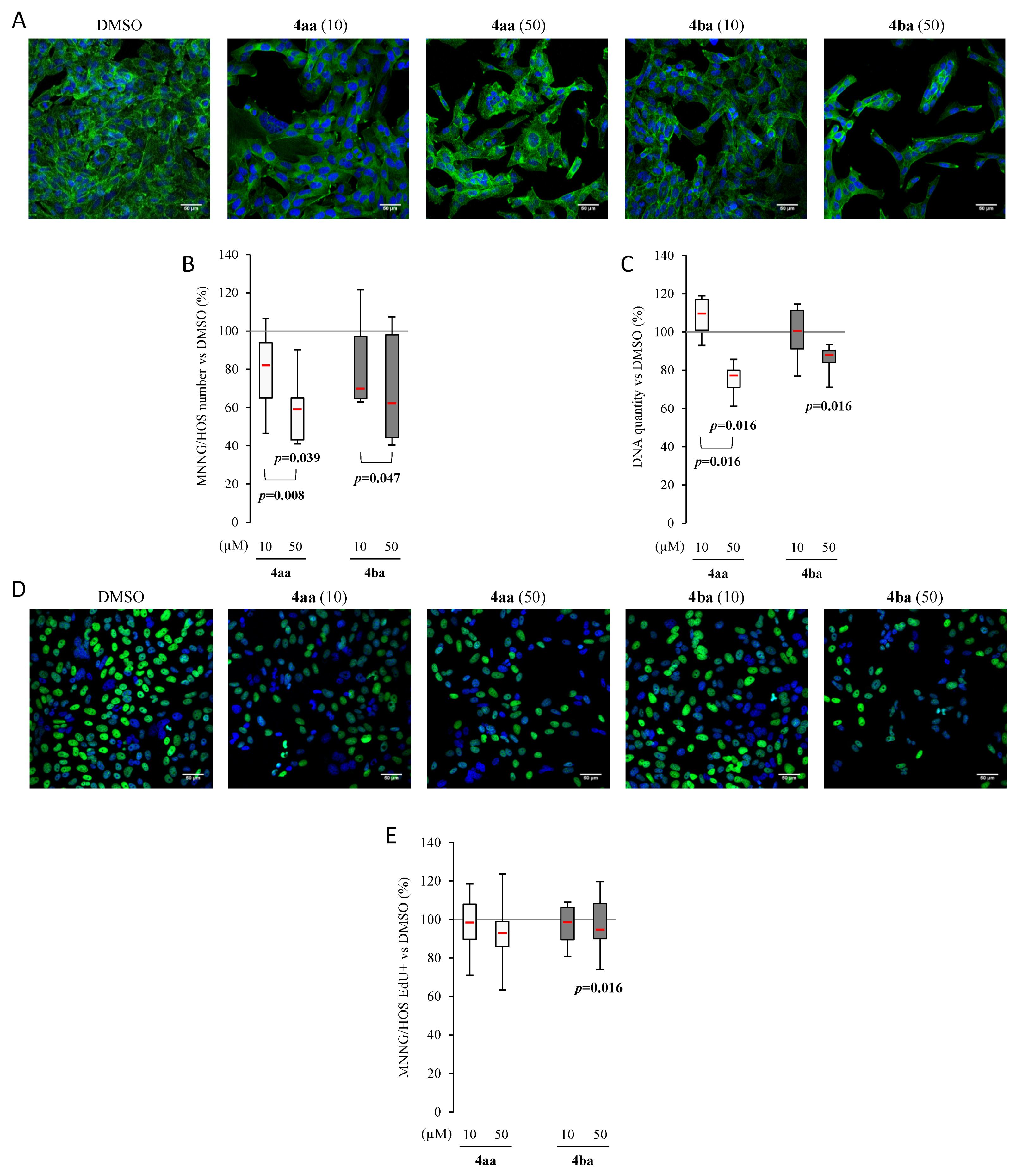
3.3. Pyridazinone Scaffold-Based Compounds Decrease Osteosarcoma Cell Line Proliferation
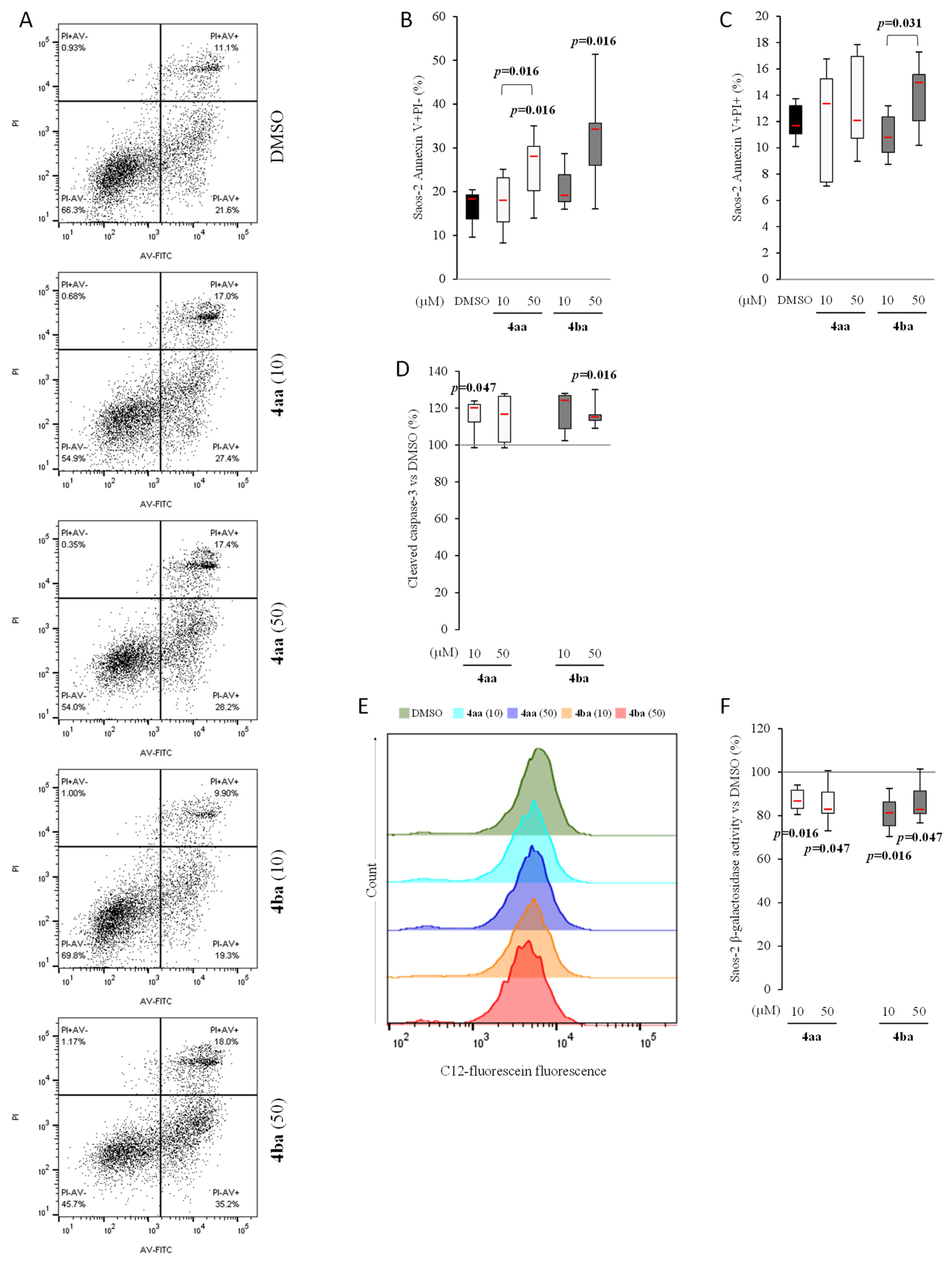
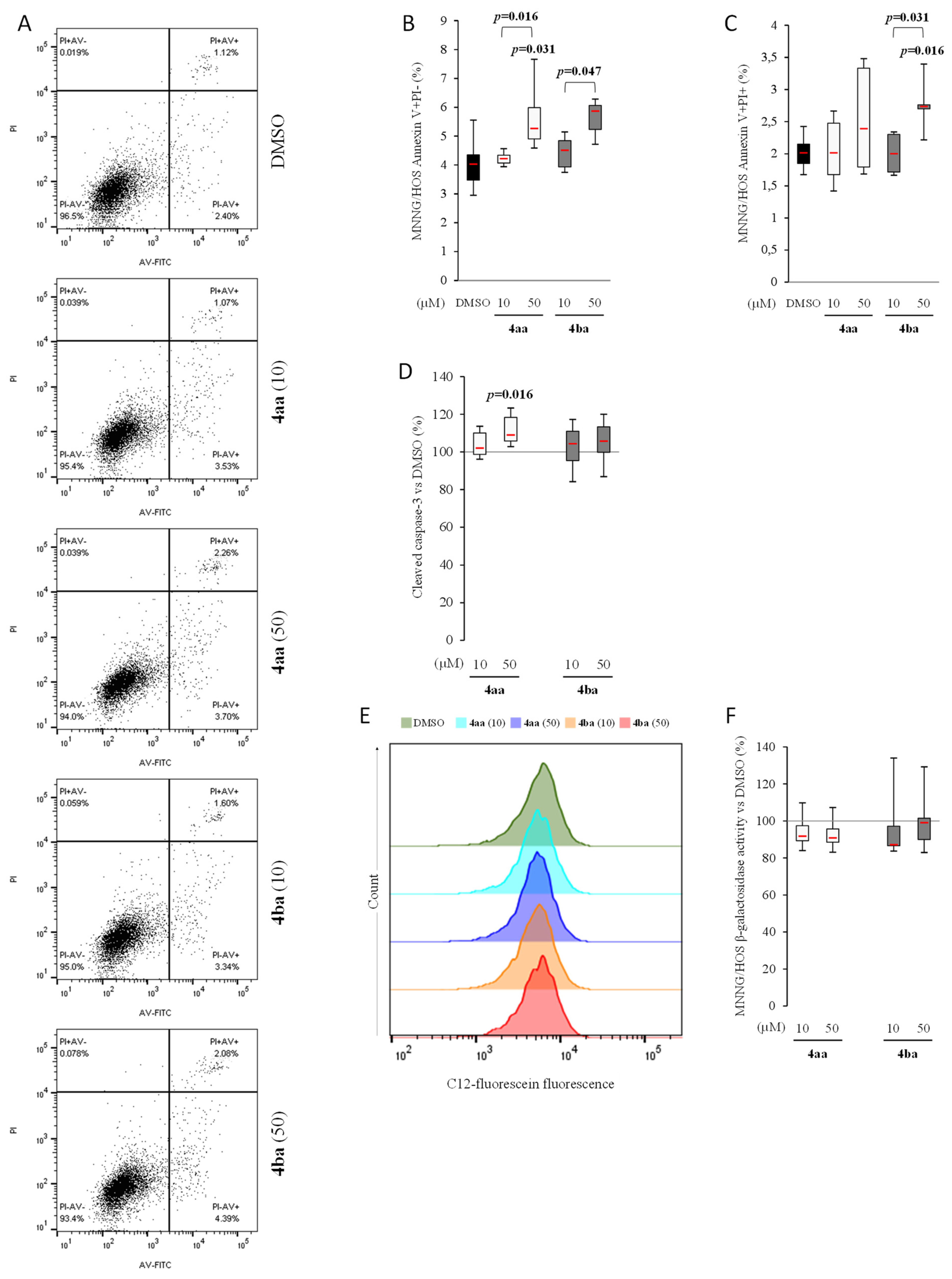
3.4. Pyridazinone Scaffold-Based Compounds Increase Osteosarcoma Cell Apoptosis
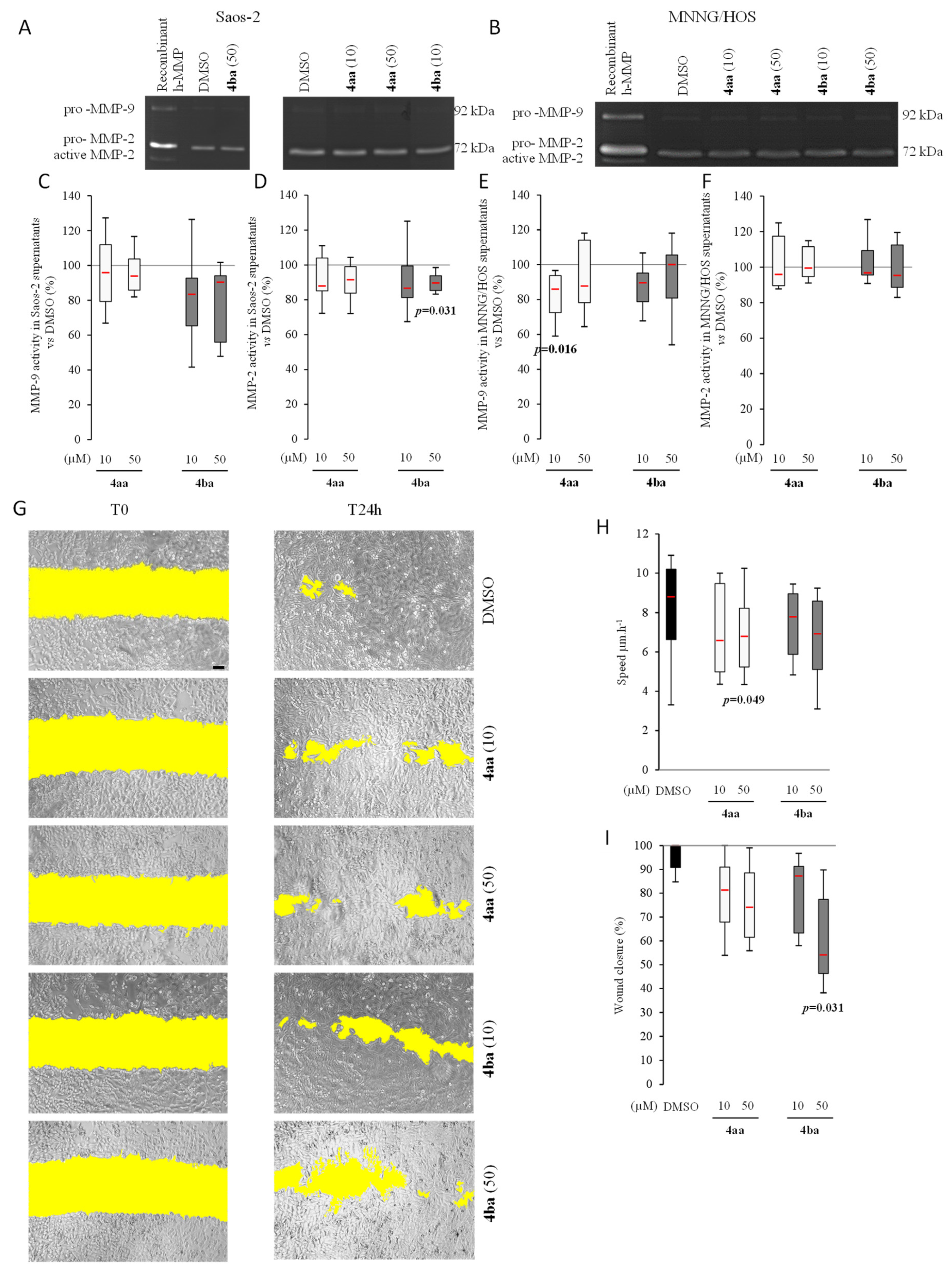
3.5. Pyridazinone Scaffold-Based Compounds Reduce MNNG Migratory Capabilities In Vitro
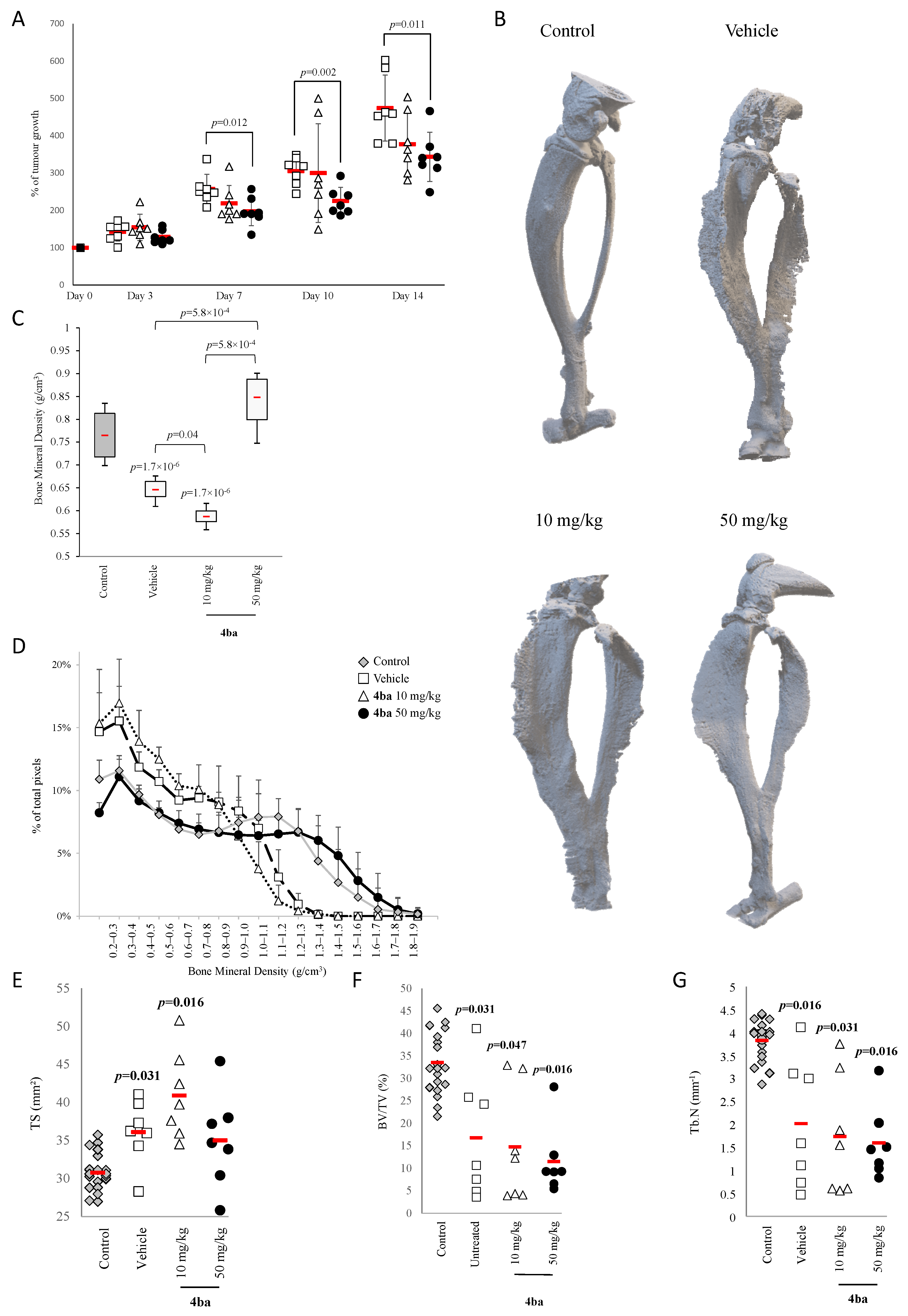
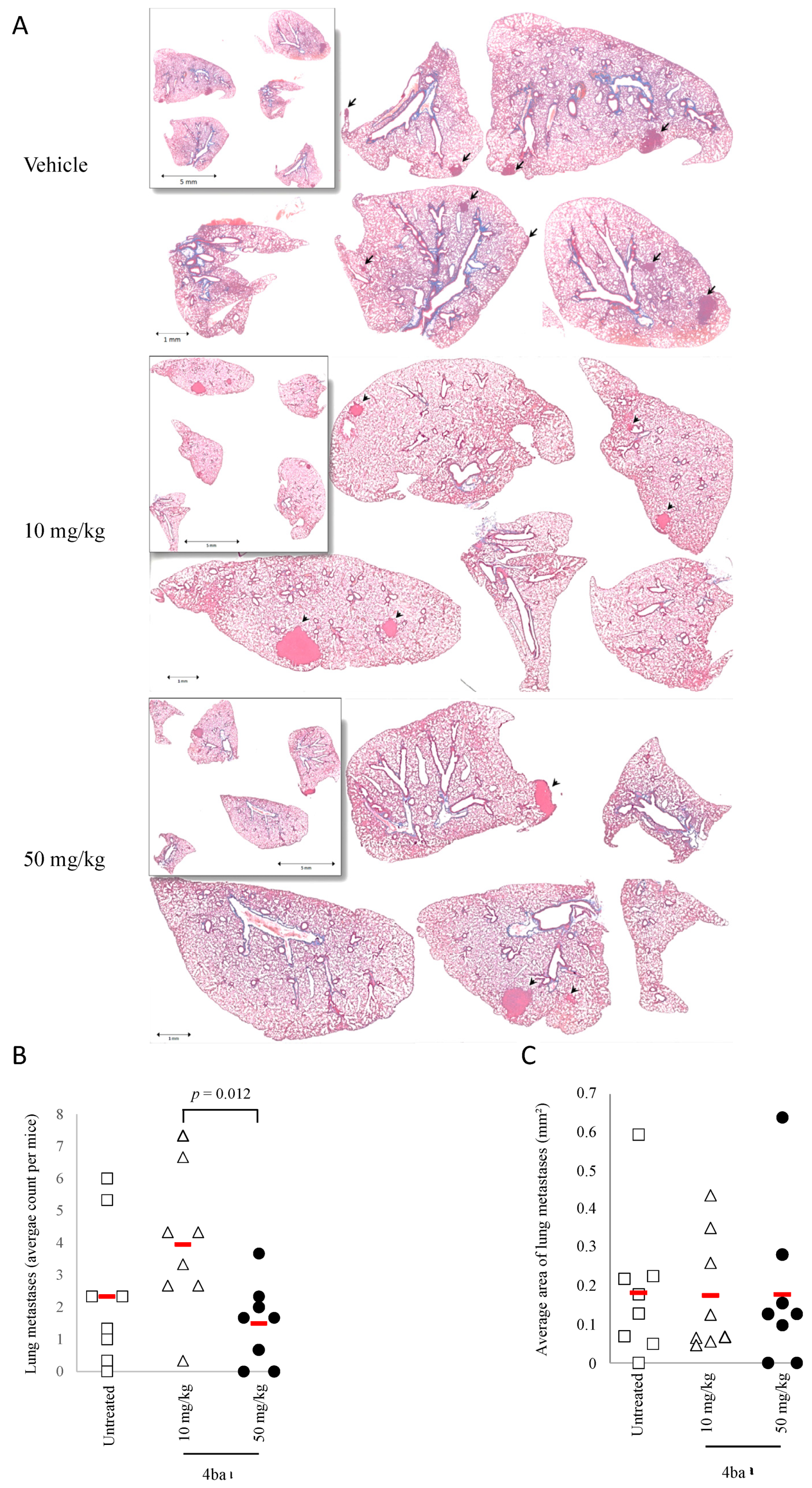
3.6. Pyridazinone Scaffold-Based Compounds Reduce Orthotopic Tumor Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mirabello, L.; Troisi, R.J.; Savage, S.A. International osteosarcoma incidence patterns in children and adolescents, middle ages and elderly persons. Int. J. Cancer 2009, 125, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawad, M.U.; Cheung, M.C.; Clarke, J.; Koniaris, L.G.; Scully, S.P. Osteosarcoma: Improvement in survival limited to high-grade patients only. J. Cancer Res. Clin. Oncol. 2011, 137, 597–607. [Google Scholar] [CrossRef]
- Anninga, J.K.; Gelderblom, H.; Fiocco, M.; Kroep, J.R.; Taminiau, A.H.; Hogendoorn, P.C.; Egeler, R.M. Chemotherapeutic adjuvant treatment for osteosarcoma: Where do we stand? Eur. J. Cancer 2011, 47, 2431–2445. [Google Scholar] [CrossRef]
- Bacci, G.; Longhi, A.; Fagioli, F.; Briccoli, A.; Versari, M.; Picci, P. Adjuvant and neoadjuvant chemotherapy for osteosarcoma of the extremities: 27 year experience at Rizzoli Institute, Italy. Eur. J. Cancer 2005, 41, 2836–2845. [Google Scholar] [CrossRef] [Green Version]
- Smeland, S.; Bielack, S.S.; Whelan, J.; Bernstein, M.; Hogendoorn, P.; Krailo, M.D.; Gorlick, R.; Janeway, K.A.; Ingleby, F.C.; Anninga, J.; et al. Survival and prognosis with osteosarcoma: Outcomes in more than 2000 patients in the EURAMOS-1 (European and American Osteosarcoma Study) cohort. Eur. J. Cancer 2019, 109, 36–50. [Google Scholar] [CrossRef] [Green Version]
- Perkins, S.M.; Shinohara, E.T.; DeWees, T.; Frangoul, H. Outcome for children with metastatic solid tumors over the last four decades. PLoS ONE 2014, 9, e100396. [Google Scholar] [CrossRef] [PubMed]
- Beavo, J.A.; Brunton, L.L. Cyclic nucleotide research—Still expanding after half a century. Nat. Rev. Mol. Cell Biol. 2002, 3, 710–718. [Google Scholar] [CrossRef]
- Graves, L.M.; Lawrence, J.C., Jr. Insulin, growth factors, and cAMP: Antagonism in the signal transduction pathways. Trends Endocrinol. Metab. 1996, 7, 43–50. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.J.; Huang, X.Y. cAMP inhibits cell migration by interfering with Rac-induced lamellipodium formation. J. Biol. Chem. 2008, 20, 13799–13805. [Google Scholar] [CrossRef] [Green Version]
- Insel, P.A.; Zhang, L.; Murray, F.; Yokouchi, H.; Zambon, A.C. Cyclic AMP is both a pro-apoptotic and anti-apoptotic second messenger. Acta Physiol. 2012, 204, 277–287. [Google Scholar] [CrossRef]
- Martinez, A.; Gil, C. cAMP-specific phosphodiesterase inhibitors: Promising drugs for inflammatory and neurological diseases. Expert Opin. Ther. Pat. 2014, 24, 1311–1321. [Google Scholar] [CrossRef]
- Asif, M. A mini review on biological activities of pyridazinone derivatives as antiulcer, antisecretory, antihistamine and particularly against histamine H3R. Mini Rev. Med. Chem. 2015, 14, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, W.; Shaquiquzzaman, M.; Akhter, M.; Verma, G.; Khan, M.F.; Alam, M.M. The therapeutic journey of pyridazinone. Eur. J. Med. Chem. 2016, 123, 256–281. [Google Scholar] [CrossRef]
- Wermuth, C.G. Are pyridazines privileged structures? Med. Chem. Comm. 2011, 2, 935–941. [Google Scholar] [CrossRef]
- Chung, K.F. Phosphodiesterase inhibitors in airways disease. Eur. J. Pharmacol. 2006, 533, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Houslay, M.D.; Schafer, P.; Zhang, K.Y. Keynote review: Phosphodiesterase-4 as a therapeutic target. Drug Discov. Today 2005, 10, 1503–1519. [Google Scholar] [CrossRef]
- Gavaldà, A.; Roberts, R.S. Phosphodiesterase-4 inhibitors: A review of current developments (2010–2012). Expert Opin. Ther. Pat. 2013, 23, 997–1016. [Google Scholar] [CrossRef]
- Malinka, W.; Redzicka, A.; Lozach, O. New derivatives of pyrrolo(3,4-d)pyridazinone and their anticancer effects. Il Farmaco 2004, 59, 457–462. [Google Scholar] [CrossRef]
- Savai, R.; Pullamsetti, S.S.; Banat, G.A.; Weissmann, N.; Ghofrani, H.A.; Grimminger, F.; Schermuly, R.T. Targeting cancer with phosphodiesterase inhibitors. Expert Opin. Investig. Drugs 2010, 19, 117–131. [Google Scholar] [CrossRef]
- Rathish, I.G.; Javed, K.; Ahmad, S.; Bano, S.; Alam, M.S.; Akhter, M.; Pillai, K.K.; Ovais, S.; Samim, M. Synthesis and evaluation of anticancer activity of some novel 6-aryl-2-(p-sulfamylphenyl)-pyridazin-3(2H)-ones. Eur. J. Med. Chem. 2012, 49, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Dai, M.; Cui, R.; Wang, Y.; Li, C.; Peng, X.; Zhao, J.; Wang, B.; Dai, Y.; Feng, D.; et al. Design, synthesis and biological evaluation of pyrazolo [3,4-d]pyridazinone derivatives as covalent FGFR inhibitors. Acta Pharm. Sin. B 2021, 11, 781–794. [Google Scholar] [CrossRef]
- Cartledge, D.M.; Robbins, K.M.; Drake, K.M.; Sternberg, R.; Stabley, D.L.; Gripp, K.W.; Kolb, E.A.; Sol-Church, K.; Napper, A.D. Cytotoxicity of Zardaverine in Embryonal Rhabdomyosarcoma from a Costello Syndrome Patient. Front. Oncol. 2017, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, T.; Kobayashi, S.; Ebara, S.; Yoshimura, Y.; Horiuchi, H.; Tsutsumimoto, T.; Wakabayashi, S.; Takaoka, K. Phosphodiesterase inhibitors, pentoxifylline and rolipram, increase bone mass mainly by promoting bone formation in normal mice. Bone 2000, 27, 811–817. [Google Scholar] [CrossRef]
- Waki, Y.; Horita, T.; Miyamoto, K.; Ohya, K.; Kasugai, S. Effects of XT-44, a phosphodiesterase 4 inhibitor, in osteoblastgenesis and osteoclastgenesis in culture and its therapeutic effects in rat osteopenia models. Jpn. J. Pharmacol. 1999, 79, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, W.; Tian, X.Y.; Chen, J.; Setterberg, R.B.; Lundy, M.W.; Chmielzwski, P.; Froman, C.A.; Jee, W.S. Rolipram, a phosphodiesterase 4 inhibitor, prevented cancellous and cortical bone loss by inhibiting endosteal bone resorption and maintaining the elevated periosteal bone formation in adult ovariectomized rats. J. Musculoskelet. Neuronal Interact. 2007, 7, 119–130. [Google Scholar] [PubMed]
- Muff, R.; Nigg, N.; Gruber, P.; Walters, D.; Born, W.; Fuchs, B. Altered morphology, nuclear stability and adhesion of highly metastatic derivatives of osteoblast-like Saos-2 osteosarcoma cells. Anticancer Res. 2007, 27, 3973–3979. [Google Scholar]
- Li, C.; Li, G.; Liu, M.; Zhou, T.; Zhou, H. Paracrine effect of inflammatory cytokine-activated bone marrow mesenchymal stem cells and its role in osteoblast function. J. Biosci. Bioeng. 2016, 121, 213–219. [Google Scholar] [CrossRef]
- Barberot, C.; Moniot, A.; Allart-Simon, I.; Malleret, L.; Yegorova, T.; Laronze-Cochard, M.; Bentaher, A.; Médebielle, M.; Bouillon, J.-P.; Hénon, E.; et al. Synthesis and Biological Evaluation of Pyridazinone derivatives as Potential Anti-inflammatory Agents. Eur. J. Med. Chem. 2018, 146, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Allart-Simon, I.; Moniot, A.; Bisi, N.; Ponce-Vargas, M.; Audonnet, S.; Laronze-Cochard, M.; Sapi, J.; Hénon, E.; Velard, F.; Gérard, S. Pyridazinone derivatives as potential anti-inflammatory agents: Synthesis and biological evaluation as PDE4 inhibitors. RSC Med. Chem. 2021, 12, 584–592. [Google Scholar] [CrossRef]
- Gérard, S.; Raoul, M.; Sapi, J. A New Approach to the Synthesis of N-Alkylated 2-Substituted Azetidin-3-ones. Eur. J. Org. Chem. 2006, 10, 2440–2445. [Google Scholar] [CrossRef]
- Bartoli, G.; Bosco, M.; Carlone, A.; Pesciaioli, F.; Sambri, L.; Melchiorre, P. Organocatalytic asymmetric Friedel-Crafts alkylation of indoles with simple alpha,beta-unsaturated ketones. Org. Lett. 2007, 9, 1403–1405. [Google Scholar] [CrossRef]
- Hong, L.; Sun, W.; Liu, C.; Wang, L.; Wong, K.; Wang, R. Enantioselective Friedel–Crafts Alkylation of 4,7-Dihydroindoles with Enones Catalyzed by Primary–Secondary Diamines. Chem. Eur. J. 2009, 15, 11105–11108. [Google Scholar] [CrossRef]
- Adachi, S.; Tanaka, F.; Watanabe, K.; Harada, T. Oxazaborolidinone-Catalyzed Enantioselective Friedel−Crafts Alkylation of Furans and Indoles with α,β-Unsaturated Ketones. Org. Lett. 2009, 11, 5206–5209. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 12, 1798–1806. [Google Scholar] [CrossRef]
- Laheij, A.M.; de Soet, J.J.; Veerman, E.C.; Bolscher, J.G.; van Loveren, C. The influence of oral bacteria on epithelial cell migration in vitro. Mediat. Inflamm. 2013, 2013, 154532. [Google Scholar] [CrossRef] [Green Version]
- Crenn, V.; Biteau, K.; Amiaud, J.; Dumars, C.; Guiho, R.; Vidal, L.; Le Nail, L.-R.; Heymann, D.; Moreau, A.; Gouin, F.; et al. Bone microenvironment has an influence on the histological response of osteosarcoma to chemotherapy: Retrospective analysis and preclinical modeling. Am. J. Cancer Res. 2017, 11, 2333–2349. [Google Scholar]
- Campbell, G.M.; Sophocleous, A. Quantitative analysis of bone and soft tissue by micro-computed tomography: Applications to ex vivo and in vivo studies. Bonekey Rep. 2014, 20, 564. [Google Scholar] [CrossRef] [Green Version]
- Schosserer, M.; Grillari, J.; Breitenbach, M. The Dual Role of Cellular Senescence in Developing Tumors and Their Response to Cancer Therapy. Front. Oncol. 2017, 7, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwan, D.G.; Brunton, V.G.; Baillie, G.S.; Leslie, N.R.; Houslay, M.D.; Frame, M.C. Chemoresistant KM12C colon cancer cells are addicted to low cyclic AMP levels in a phosphodiesterase 4-regulated compartment via effects on phosphoinositide 3-kinase. Cancer Res. 2007, 67, 5248–5257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, M.; Murata, T.; Shimizu, K.; Sugiyama, T.; Nakagawa, T.; Manganiello, V.C.; Tagawa, T. Phosphodiesterase 4 in osteoblastic osteosarcoma cells as a potential target for growth inhibition. Anticancer Drugs 2003, 14, 377–381. [Google Scholar] [CrossRef]
- Ahlström, M.; Pekkinen, M.; Huttunen, M.; Lamberg-Allardt, C. Cyclic nucleotide phosphodiesterases (PDEs) in human osteoblastic cells; the effect of PDE inhibition on cAMP accumulation. Cell. Mol. Biol. Lett. 2005, 10, 305–319. [Google Scholar]
- Ahlström, M.; Pekkinen, M.; Huttunen, M.; Lamberg-Allardt, C. Dexamethasone down-regulates cAMP-phosphodiesterase in human osteosarcoma cells. Biochem. Pharmacol. 2005, 69, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Quan, H.; Xie, C.; Wang, L.; Hu, Y.; Lou, L. Phosphodiesterase 3/4 inhibitor zardaverine exhibits potent and selective antitumor activity against hepatocellular carcinoma both in vitro and in vivo independently of phosphodiesterase inhibition. PLoS ONE 2014, 9, e90627. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Liu, X.; Xu, S.; Tang, Q.; Duan, Y.; Xiao, Z.; Zhi, J.; Jiang, L.; Zheng, P.; Zhu, W. Discovery of novel pyrrolo-pyridine/pyrimidine derivatives bearing pyridazinone moiety as c-Met kinase inhibitors. Eur. J. Med. Chem. 2017, 141, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zheng, Y.; Wang, Y.; Sheng, W.; Li, Y.; Liu, X.; Si, S.; Shao, R.; Zhen, Y. A new compound of thiophenylated pyridazinone IMB5043 showing potent antitumor efficacy through ATM-Chk2 pathway. PLoS ONE 2018, 13, e0191984. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, D.A.; DeJesus, R.E.; Contreras, L.; Rodriguez-Palomares, I.A.; Villanueva, P.J.; Balderrama, K.S.; Monterroza, L.; Larragoity, M.; Varela-Ramirez, A.; Aguilera, R.J. A new pyridazinone exhibits potent cytotoxicity on human cancer cells via apoptosis and poly-ubiquitinated protein accumulation. Cell Biol. Toxicol. 2019, 35, 503–519. [Google Scholar] [CrossRef]
- Demaria, M.; O’Leary, M.N.; Chang, J.; Shao, L.; Liu, S.; Alimirah, F.; Koenig, K.; Le, C.; Mitin, N.; Deal, A.M.; et al. Cellular Senescence Promotes Adverse Effects of Chemotherapy and Cancer Relapse. Cancer Discov. 2017, 7, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillon, J.; Petit, C.; Toutain, B.; Guette, C.; Lelièvre, E.; Coqueret, O. Chemotherapy-induced senescence, an adaptive mechanism driving resistance and tumor heterogeneity. Cell Cycle 2019, 18, 2385–2397. [Google Scholar] [CrossRef]
- Oku, T.; Shimada, K.; Kenmotsu, H.; Ando, Y.; Kurisaka, C.; Sano, R.; Tsuiji, M.; Hasegawa, S.; Fukui, T.; Tsuji, T. Stimulation of Peritoneal Mesothelial Cells to Secrete Matrix Metalloproteinase-9 (MMP-9) by TNF-α: A Role in the Invasion of Gastric Carcinoma Cells. Int. J. Mol. Sci. 2018, 19, 3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Murata, T.; Shimizu, K.; Morita, H.; Inui, M.; Tagawa, T. Phosphodiesterase 4 regulates the migration of B16-F10 melanoma cells. Exp. Ther. Med. 2012, 4, 205–210. [Google Scholar] [CrossRef]
- Fleming, Y.M.; Frame, M.C.; Houslay, M.D. PDE4-regulated cAMP degradation controls the assembly of integrin-dependent actin adhesion structures and REF52 cell migration. J. Cell Sci. 2004, 117, 2377–2388. [Google Scholar] [CrossRef] [Green Version]
- David, E.; Cagnol, S.; Goujon, J.-Y.; Egorov, M.; Taurelle, J.; Benestea, C.; Morandeau, L.; Moal, C.; Sicard, M.; Pairel, S.; et al. 12b80—Hydroxybisphosphonate Linked Doxorubicin: Bone Targeted Strategy for Treatment of Osteosarcoma. Bioconjug. Chem. 2019, 30, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Gobin, B.; Huin, M.B.; Lamoureux, F.; Ory, B.; Charrier, C.; Lanel, R.; Battaglia, S.; Redini, F.; Lezot, F.; Blanchard, F.; et al. BYL719, a new α-specific PI3K inhibitor: Single administration and in combination with conventional chemotherapy for the treatment of osteosarcoma. Int. J. Cancer 2015, 136, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, P.; Pei, Y.; Zheng, K.; Wang, W.; Qiu, E.; Zhang, X. Addition of Zoledronate to Chemotherapy in Patients with Osteosarcoma Treated with Limb-Sparing Surgery: A Phase III Clinical Trial. Med. Sci. Monit. 2019, 25, 1429–1438. [Google Scholar] [CrossRef]
- Miyamoto, K.; Waki, Y.; Horita, T.; Kasugai, S.; Ohya, K. Reduction of bone loss by denbufylline, an inhibitor of phosphodiesterase 4. Biochem. Pharmacol. 1997, 54, 613–617. [Google Scholar] [CrossRef]
- Porwal, K.; Pal, S.; Bhagwati, S.; Siddiqi, M.I.; Chattopadhyay, N. Therapeutic potential of phosphodiesterase inhibitors in the treatment of osteoporosis: Scopes for therapeutic repurposing and discovery of new oral osteoanabolic drugs. Eur. J. Pharmacol. 2021, 899, 174015. [Google Scholar] [CrossRef]
- Ahn, J.H.; Cho, W.H.; Lee, J.A.; Kim, D.H.; Seo, J.H.; Lim, J.S. Bone mineral density change during adjuvant chemotherapy in pediatric osteosarcoma. Ann. Pediatr. Endocrinol. Metab. 2015, 20, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Weilbaecher, K.N.; Guise, T.A.; McCauley, L.K. Cancer to bone: A fatal attraction. Nat. Rev. Cancer 2011, 11, 411–425. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniot, A.; Braux, J.; Bour, C.; Guillaume, C.; Lamret, F.; Allart-Simon, I.; Audonnet, S.; Renault, S.; Rédini, F.; Laronze-Cochard, M.; et al. Pyridazinone Derivatives Limit Osteosarcoma-Cells Growth In Vitro and In Vivo. Cancers 2021, 13, 5992. https://doi.org/10.3390/cancers13235992
Moniot A, Braux J, Bour C, Guillaume C, Lamret F, Allart-Simon I, Audonnet S, Renault S, Rédini F, Laronze-Cochard M, et al. Pyridazinone Derivatives Limit Osteosarcoma-Cells Growth In Vitro and In Vivo. Cancers. 2021; 13(23):5992. https://doi.org/10.3390/cancers13235992
Chicago/Turabian StyleMoniot, Aurélie, Julien Braux, Camille Bour, Christine Guillaume, Fabien Lamret, Ingrid Allart-Simon, Sandra Audonnet, Sarah Renault, Francoise Rédini, Marie Laronze-Cochard, and et al. 2021. "Pyridazinone Derivatives Limit Osteosarcoma-Cells Growth In Vitro and In Vivo" Cancers 13, no. 23: 5992. https://doi.org/10.3390/cancers13235992