
Screening for the Key Proteins Associated with Rete Testis Invasion in Clinical Stage I Seminoma via Label-Free Quantitative Mass Spectrometry

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Ethics
2.2. Tissue Collection and Sample Preparation for LC–MS/MS
2.3. Protein Digestion and LC–MS/MS
2.4. GO and Pathway Analysis
2.5. Patient Samples and Cell Lines
2.6. Immunohistochemistry (IHC)
2.7. mRNA Expression Analysis in TGCT Cell Lines
2.8. Statistical Analysis
3. Results
3.1. RTI Promotes Protein Expression Change
3.2. GO Annotation and Functional Classification
3.3. Database Search
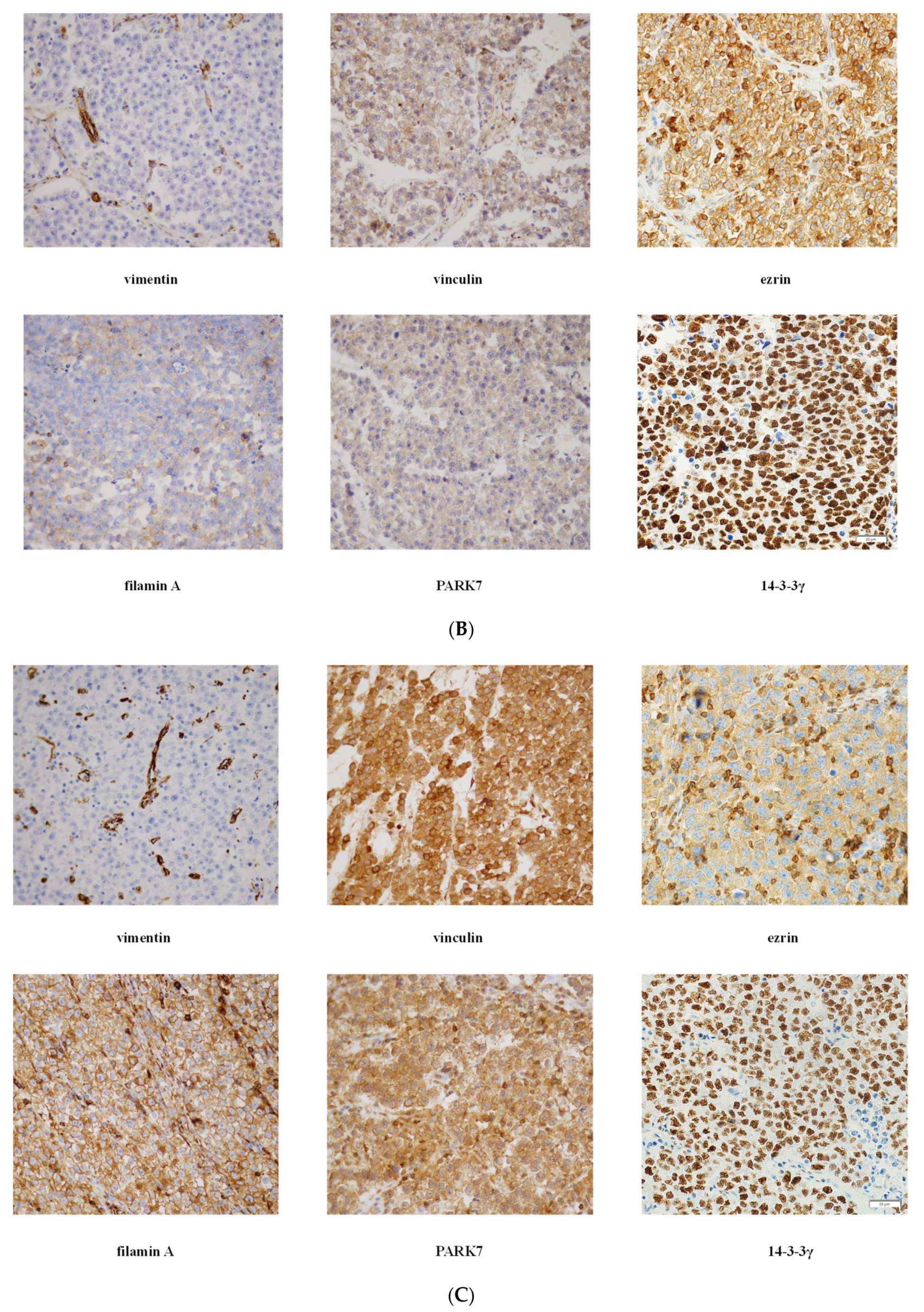
3.4. Validation of Mass Spectrometry Data by IHC
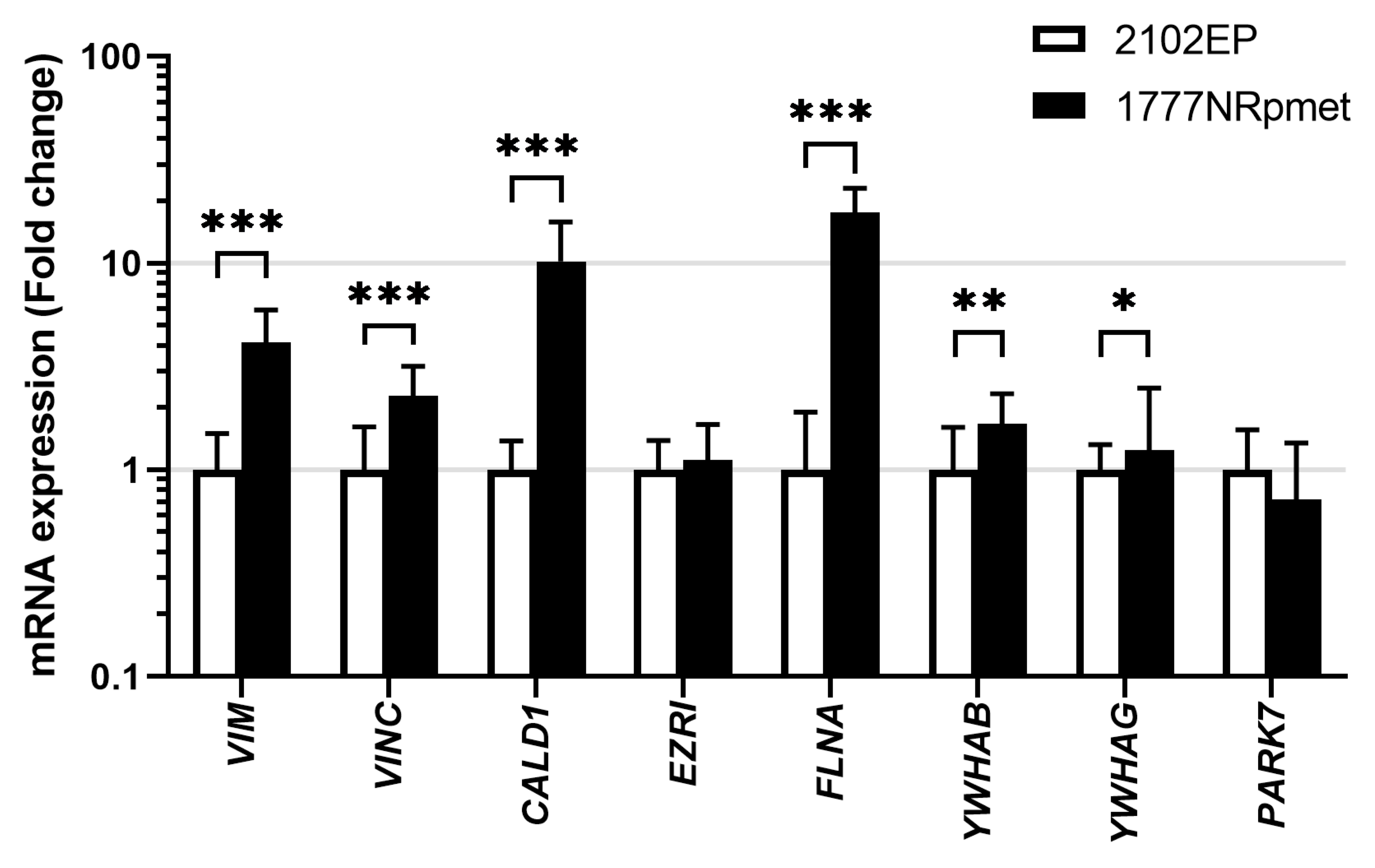
3.5. Expression in TGCT Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zengerling, F.; Kunath, F.; Jensen, K.; Ruf, C.; Schmidt, S.; Spek, A. Prognostic factors for tumor recurrence in patients with clinical stage I seminoma undergoing surveillance-A systematic review. Urol. Oncol. 2018, 36, 448–458. [Google Scholar] [CrossRef]
- Boormans, J.L.; de Mayor, C.J.; Marconi, L.; Yuan, Y.; Laguna Pes, M.P.; Bokemeyer, C.; Nicolai, N.; Algaba, F.; Oldenburg, J.; Albers, P. Testicular tumour size and rete testis invasion as prognostic factors for the risk of relapse of clinical stage I seminoma testis patients under surveillance: A systematic review by the testicular cancer guidelines panel. Eur. Urol. 2018, 73, 394–405. [Google Scholar] [CrossRef]
- Oliver, R.T.D.; Mason, M.D.; Mead, G.M.; von der Masse, H.; Rustin, G.J.S.; Joffe, J.K.; de Wit, R.; Aass, N.; Graham, J.D.; Coleman, R.; et al. Radiotherapy versus single-dose carboplatin in adjuvant treatment of stage I seminoma: A randomised trial. Lancet 2005, 366, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Fosså, S.D.; Horwich, A.; Russell, J.M.; Roberts, J.T.; Cullen, M.H.; Hodson, N.J.; Jones, W.G.; Yosef, H.; Duchesne, G.M.; Owen, J.R.; et al. Optimal planning target volume for stage I testicular seminoma: A Medical Research Council randomized trial. Medical Research Council Testicular Tumor Working Group. J. Clin. Oncol. 1999, 17, 1146. [Google Scholar] [CrossRef]
- Dieckmann, K.P.; Dralle-Filiz, I.; Matthies, C.; Heinzelbecker, J.; Bedke, J.; Ellinger, J.; Anheuser, P.; Souchon, R.; Pichlmeier, U. German Testicular Cancer Study Group. Testicular seminoma clinical stage 1: Treatment outcome on a routine care level. J. Cancer Res. Clin. Oncol. 2016, 142, 1599–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travis, L.B.; Beard, C.; Allan, J.M.; Dahl, A.A.; Feldman, D.R.; Oldenburg, J.; Daugaard, G.; Kelly, J.L.; Dolan, M.E.; Hannigan, R.; et al. Testicular cancer survivorship: Research strategies and recommendations. J. Natl. Cancer Inst. 2010, 102, 1114–1130. [Google Scholar] [CrossRef]
- Diminutto, A.; Basso, U.; Maruzzo, M.; Morelli, F.; De Giorgi, U.; Perin, A.; Fraccon, A.P.; Lo Re, G.; Rizzi, A.; Sava, T.; et al. Adjuvant carboplatin treatment in 115 patients with stage I seminoma: Retrospective multicenter survey. Clin. Genitourin. Cancer 2016, 14, e161–e169. [Google Scholar] [CrossRef]
- Fischer, S.; Tandstad, T.; Wheater, M.; Porfiri, E.; Flechon, A.; Aparicio, J.; Klingbiel, D.; Skrbinc, B.; Basso, U.; Shamash, J.; et al. Outcome of men with relapse after adjuvant carboplatin for clinical stage I seminoma. J. Clin. Oncol. 2017, 35, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, C.; Cathomas, R.; Wheater, M.; Klingbiel, D.; Fehr, M.; Bennett, J.; Markham, H.; Lee, C.; Crabb, S.J.; Geldart, T. Treatment outcome and patterns of relapse following adjuvant carboplatin for stage I testicular seminomatous germ-cell tumour: Results from a 17-year UK experience. Ann. Oncol. 2015, 26, 1865–1870. [Google Scholar] [CrossRef]
- Warde, P.; Specht, L.; Horwich, A.; Oliver, T.; Panzarella, T.; Gospodarowicz, M.; von der Maase, H. Prognostic factors for relapse in stage I seminoma managed by surveillance: A pooled analysis. J. Clin. Oncol. 2002, 20, 4448–4452. [Google Scholar] [CrossRef] [PubMed]
- Von der Maase, H.; Specht, L.; Jacobsen, G.K.; Jakobsen, A.; Madsen, E.L.; Pedersen, M.; Rørth, M.; Schultz, H. Surveillance following orchidectomy for stage I seminoma of the testis. Eur. J. Cancer 1993, 29, 1931–1934. [Google Scholar] [CrossRef]
- Valdevenito, J.P.; Gallegos, I.; Fernandez, C.; Acevedo, C.; Palma, R. Correlation between primary tumor pathologic features and presence of clinical metastasis at diagnosis of testicular seminoma. Urology 2007, 70, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, J.; Fossa, S.D.; Nuver, J.; Heidenreich, A.; Schmoll, H.J.; Bokemeyer, C.; Horwich, A.; Beyer, J.; Kataja, V.; on behalf of the EMSO Guidelines Working Group. Testicular seminoma and non-seminoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24, vi125–vi132. [Google Scholar] [CrossRef]
- Albers, P.; Albrecht, W.; Algaba, F.; Bokemeyer, C.; Cohn-Cedermark, G.; Fizazi, K.; Horwich, A.; Laguna, M.P.; Nicolai, N.; Oldenburg, J. Guidelines on testicular cancer: 2015 update. Eur. Urol. 2015, 68, 1054–1068. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.; Daugaard, G.; Tyldesley, S.; Atenafu, E.G.; Panzarella, T.; Kollmannsberger, C.; Warde, P. Evaluation of a prognostic model for risk of relapse in stage I seminoma surveillance. Cancer Med. 2015, 4, 155–160. [Google Scholar] [CrossRef]
- Tandstad, T.; Smaaland, R.; Solberg, A.; Bremnes, R.M.; Langberg, C.W.; Laurell, A.; Stierner, U.K.; Ståhl, O.; Cavallin-Ståhl, E.K.; Klepp, O.H.; et al. Management of seminomatous testicular cancer: A binational prospective population-based study from the Swedish norwegian testicular cancer study group. J. Clin. Oncol. 2011, 29, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, A.; Cheng, T.; Zhang, J.; Trpkov, K. Testicular hilum and vascular invasion predict advanced clinical stage in nonseminomatous germ cell tumors. Mod. Pathol. 2013, 26, 579–586. [Google Scholar] [CrossRef]
- Dry, S.M.; Renshaw, A.A. Extratesticular extension of germ cell tumors preferentially occurs at the hilum. Am. J. Clin. Pathol. 1999, 111, 534–538. [Google Scholar] [CrossRef]
- Zhu, Q.; Sun, Y.; Zhou, Q.; He, Q.; Qian, H. Identification of key genes and pathways by bioinformatics analysis with TCGA RNA sequencing data in hepatocellular carcinoma. Mol. Clin. Oncol. 2018, 9, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Raman, P.; Maddipati, R.; Lim, K.H.; Tozeren, A. Pancreatic cancer survival analysis defines a signature that predicts outcome. PLoS ONE 2018, 13, e0201751. [Google Scholar] [CrossRef] [PubMed]
- Bosquet, J.G.; Marchion, D.C.; Chon, H.; Lancaster, J.M.; Chanock, S. Analysis of chemotherapeutic response in ovarian cancers using publicly available high-throughput data. Cancer Res. 2014, 74, 3902–3912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, X.; Sherman, B.T.; Huang, D.W.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A stateful web service to facilitate gene/protein list analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronson, D.L.; Vessella, R.L.; Fraley, E.E. Differentiation potential of human embryonal carcinoma cell lines. Cell Differ. 1984, 15, 129–132. [Google Scholar] [CrossRef]
- Andrews, P.W.; Bronson, D.L.; Benham, F.; Strickland, S.; Knowles, B.B. A comparative study of eight cell lines derived from human testicular teratocarcinoma. Int. J. Cancer 1980, 26, 269–280. [Google Scholar] [CrossRef]
- Roška, J.; Wachsmannová, L.; Hurbanová, L.; Šestáková, Z.; Mueller, T.; Jurkovičová, D.; Chovanec, M. Differential gene expression in cisplatin-resistant and -sensitve testicular germ cell tumor cell lines. Oncotarget 2020, 11, 4735–4753. [Google Scholar] [CrossRef]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to build a bridge from a population-based to a more "personalized" approach to cancer staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Verrill, C.; Yilmaz, A.; Srigley, J.R.; Amin, M.B.; Comperat, E.; Egevad, L.; Ulbright, T.M.; Tickoo, S.K.; Berney, D.M.; Epstein, J.I. Reporting and staging of testicular germ cell tumors: The international society of urological pathology (ISUP) testicular cancer consultation conference recommendations. Am. J. Surg. Pathol. 2017, 41, e22–e32. [Google Scholar] [CrossRef]
- Huang, R.Y.; Guilford, P.; Thiery, J.P. Early events in cell adhesion and polarity during epithelial-mesenchymal transition. J. Cell Sci. 2012, 125, 4417–4422. [Google Scholar] [CrossRef] [Green Version]
- Polyak, K.; Weinberg, R.A. Transitions between epithelial and mesenchymal states: Acquisition of malignant and stem cell traits. Nat. Rev. Cancer 2009, 9, 265–273. [Google Scholar] [CrossRef]
- Gardino, A.K.; Yaffe, M.B. 14-3-3 proteins as signaling integration points for cell cycle control and apoptosis. Semin. Cell Dev. Biol. 2011, 22, 688–695. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Sehgal, L.; Bose, A.; Gulvady, A.; Senapati, P.; Thorat, R.; Basu, S.; Bhatt, K.; Hosing, A.S.; Balyan, R.; et al. 14-3-3gamma prevents centrosome amplification and neoplastic progression. Sci. Rep. 2016, 6, 26580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Lee, J.K.; Bae, Y.; Lee, B.S.; Kim, E.; Cho, C.H.; Ryoo, K.; Yoo, J.; Kim, C.H.; Yi, G.S.; et al. Suppression of 14-3-3gamma-mediated surface expression of ANO1 inhibits cancer progression of glioblastoma cells. Sci. Rep. 2016, 6, 26413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, W.; Liu, X.; Chen, W.; Li, Q.; Martinez, J.D. Overexpression of 14-3-3gamma causes polyploidization in H322 lung cancer cells. Mol. Carcinog. 2007, 46, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.O.; Kim, S.R.; Lim, K.H.; Kim, J.H.; Ajjappala, B.; Lee, H.J.; Choi, J.I. Deubiquitinating enzyme USP37 regulating oncogenic function of 14-3-3gamma. Oncotarget 2015, 6, 36551–36576. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, E.; Mimae, T.; Ito, M.; Kadoya, T.; Miyata, Y.; Ito, A.; Okada, M. Breast cancer cell motility is promoted by 14-3-3gamma. Breast Cancer 2019, 26, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.N.; Chen, C.H.; Sheu, J.C.; Lee, H.S.; Huang, G.T.; Yu, C.Y.; Lu, F.J.; Chow, L.P. Identification of human hepatocellular carcinoma-related biomarkers by two-dimensional difference gel electrophoresis and mass spectrometry. J. Proteome Res. 2005, 4, 2062–2069. [Google Scholar] [CrossRef]
- Ko, B.S.; Lai, I.R.; Chang, T.C.; Liu, T.A.; Chen, S.C.; Wang, J.; Jan, Y.J.; Liou, J.Y. Involvement of 14-3-3gamma overexpression in extrahepatic metastasis of hepatocellular carcinoma. Hum. Pathol. 2011, 42, 129–135. [Google Scholar] [CrossRef]
- Xu, G.; Wang, J.; Wu, F.; Wang, N.; Zhou, W.; Wang, Q.; Pan, W.; Ao, G.; Yang, J. YAP and 14-3-3gamma are involved in HS-OA-induced growth inhibition of hepatocellular carcinoma cells: A novel mechanism for hydrogen sulfide releasing oleanolic acid. Oncotarget 2016, 7, 52150–52165. [Google Scholar] [CrossRef]
- Woodcock, J.M.; Coolen, C.; Goodwin, K.L.; Baek, D.J.; Bittman, R.; Samuel, M.S.; Pitson, S.M.; Lopez, A.F. Destabilisation of dimeric 14-3-3 proteins as a novel approach to anti-cancer therapeutics. Oncotarget 2015, 6, 14522–14536. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, V.M.; Putnam, C.W.; Qi, W.; Martinez, J.D. P53 suppresses expression of the 14-3-3 gamma oncogene. BMC Cancer 2011, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Marine, J.C.; Francoz, S.; Maetens, M.; Wahl, G.; Toledo, F.; Lozano, G. Keeping p53 in check: Essential and synergistic functions of Mdm2 and Mdm4. Cell Death Differ. 2006, 13, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Marine, J.C.; Lozano, G. Mdm2-mediated ubiquitylation: P53 and beyond. Cell Death Differ. 2010, 17, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Regunath, K.; Jacq, X.; Prives, C. Cisplatin causes cell death via TAB1 regulation of p53/MDM2/MDMX circuitry. Genes Dev. 2013, 27, 1739–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Bismar, T.A.; Su, J.; Xu, B.; Kristiansen, G.; Varga, Z.; Teng, L.; Ingber, D.E.; Mammoto, A.; Kumar, R.; et al. Filamin A regulates focal adhesion disassembly and suppresses breast cancer cell migration and invasion. J. Exp. Med. 2010, 207, 2421–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Zhu, T.; Gao, D.; Zhang, Y.; Zhao, Q.; Liu, S.; Su, T.; Bernier, M.; Zhao, R. Filamin A expression correlates with proliferation and invasive properties of human metastatic melanoma tumors: Implications for survival in patients. J. Cancer Res. Clin. Oncol. 2014, 140, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Uramoto, H.; Akyurek, L.M.; Hanagiri, T. A positive relationship between filamin and VEGF in patients with lung cancer. Anticancer Res. 2010, 30, 3939–3944. [Google Scholar]
- Kircher, P.; Hermanns, C.; Nossek, M.; Drexler, M.K.; Grosse, R.; Fischer, M.; Sarikas, A.; Penkava, J.; Lewis, T.; Prywes, R.; et al. Filamin A interacts with the coactivator MKL1 to promote the activity of the transcription factor SRF and cell migration. Sci. Signal. 2015, 8, ra112. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.Q.; Zhang, T.P.; Zhao, W.J.; Liu, Z.W.; You, L.; Zhou, L.; Guo, J.C.; Zhao, Y.P. Filamin A: Insights into its exact role in cancers. Pathol. Oncol. Res. 2016, 22, 245–252. [Google Scholar] [CrossRef]
- Bedolla, R.G.; Wang, Y.; Asuncion, A.; Chamie, K.; Siddiqui, S.; Mudryj, M.M.; Prihoda, T.J.; Siddiqui, J.; Chinnaian, A.M.; Mehra, R.; et al. Nuclear versus cytoplasmic localization of filamin A in prostate cancer: Immunohistochemical correlation with metastases. Clin. Cancer Res. 2009, 15, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.M.; Liu, X.H.; Han, W.; Zhao, L.L.; Yuan, B.; Yuan, C.J. Differential expression of filamin A and its clinical significance in breast cancer. Oncol. Lett. 2013, 6, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S.P. Filamin A: Phenotypic diversity. Curr. Opin. Genet. Dev. 2005, 15, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.; Hebbard, G.; Trainer, A. Update on a previously reported male with a FLNA missense mutation. Eur. J. Hum. Genet. 2017, 25, 905–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, W.; Mruk, D.D.; Lie, P.P.; Lui, W.Y.; Cheng, C.Y. Filamin A is a regulator of blood-testis barrier assembly during postnatal development in the rat testis. Endocrinology 2012, 153, 5023–5035. [Google Scholar] [CrossRef] [Green Version]
- Welter, H.; Herrmann, C.; Fröhlich, T.; Flenkenthaler, F.; Eubler, K.; Schorle, H.; Nettersheim, D.; Mayerhofer, A.; Müller-Taubenberger, A. Filamin A orchestrates cytoskeletal structure, cell migration and stem cell characteristics in human seminoma TCam-2 cells. Cells 2020, 9, 2563. [Google Scholar] [CrossRef]
- Cheng, L.; Sung, M.T.; Cossu-Rocca, P.; Jones, T.D.; MacLennan, G.T.; De Jong, J.; Lopez-Beltran, A.; Montironi, R.; Looijenga, L.H.J. OCT4: Biological functions and clinical applications as a marker of germ cell neoplasia. J. Pathol. 2007, 211, 1–9. [Google Scholar] [CrossRef]
- Yue, J.; Wang, Q.; Lu, H.; Brenneman, M.; Fan, F.; Shen, Z. The cytoskeleton protein filamin-A is required for an efficient recombinational DNA double strand break repair. Cancer Res. 2009, 69, 7978–7985. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Shen, Z. Interaction with BRCA2 suggests a role for filamin-1 (hsFLNa) in DNA damage response. J. Biol. Chem. 2001, 276, 48318–48324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zengerling, F.; Muller, J.; Schrader, A.J.; Schrader, M. Clinical stage I seminoma. Is surveillance a new therapy standard? Der Urologe 2013, 52, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.; Carroll, B.; Sarallah, R.; Correia-Melo, C.; Ogrodnik, M.; Nelson, G. SQSTM1/p62 mediates crosstalk between autophagy and the UPS in DNA repair. Autophagy 2016, 12, 1917–1930. [Google Scholar] [CrossRef]
- Lyu, L.; Lin, T.C.; McCarty, N. TRIM44 mediated p62 deubiquitination enhances DNA damage repair by increasing nuclear FLNA and 53BP1 expression. Oncogene 2021, 40, 5116–5130. [Google Scholar] [CrossRef] [PubMed]
- Zeller, C.; Dai, W.; Steele, N.L.; Siddiq, A.; Walley, A.J.; Wilhelm-Benartzi, C.S.M.; Rizzo, S.; van der Zee, A.; Plumb, J.A.; Brown, R. Candidate DNA methylation drivers of acquired cisplatin resistance in ovarian cancer identified by methylome and expression profiling. Oncogene 2012, 31, 4567–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, W. Novel insights into PARK7 (DJ-1), a potential anti-cancer therapeutic target, and implications for cancer progression. J. Clin. Med. 2020, 9, 1256. [Google Scholar] [CrossRef]
- Tsuchiya, B.; Iwaya, K.; Kohno, N.; Kawate, T.; Akahoshi, T.; Matsubara, O.; Mukai, K. Clinical significance of DJ-1 as a secretory molecule: Retrospective study of DJ-1 expression at mRNA and protein levels in ductal carcinoma of the breast. Histopathology 2012, 61, 69–77. [Google Scholar] [CrossRef]
- Kawate, T.; Iwaya, K.; Kikuchi, R.; Kaise, H.; Oda, M.; Sato, E.; Hiroi, S.; Matsubara, O.; Kohno, M. DJ-1 protein expression as a predictor of pathological complete remission after neoadjuvant chemotherapy in breast cancer patients. Breast Cancer Res. Treat. 2013, 139, 51–59. [Google Scholar] [CrossRef]
- Kawate, T.; Iwaya, K.; Koshikawa, K.; Moriya, T.; Yamasaki, T.; Hasegawa, S.; Kaise, H.; Fujita, T.; Matsuo, H.; Nakamura, T.; et al. High levels of DJ-1 protein and isoelectric point 6.3 isoform in sera of breast cancer patients. Cancer Sci. 2015, 106, 938–943. [Google Scholar] [CrossRef]
- Ariga, H.; Takahashi-Niki, K.; Kato, I.; Maita, H.; Niki, T.; Iguchi-Ariga, S.M. Neuroprotective function of DJ-1 in Parkinson’s disease. Oxid. Med. Cell. Longev. 2013, 2013, 683920. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.Z.; Xiao, L.; Zhao, L.; Qiu, L.J.; Ma, Z.X.; Xu, X.W.; Liu, H.Y.; Zhou, T.T.; Wang, X.Y.; Tang, L.; et al. Molecular mechanism underlying hypoxic preconditioning-promoted mitochondrial translocation of DJ-1 in hypoxia/reoxygenation H9c2 cells. Molecules 2019, 25, 71. [Google Scholar] [CrossRef] [Green Version]
- Koch, S.; Mayer, F.; Honecker, F.; Schittenhelm, M.; Bokemeyer, C. Efficacy of cytotoxic agents used in the treatment of testicular germ cell tumours under normoxic and hypoxic conditions in vitro. Br. J. Cancer 2003, 89, 2133–2139. [Google Scholar] [CrossRef]
- Kalavska, K.; Cierna, Z.; Chovanec, M.; Takacova, M.; Svetlovska, D.; Miskovska, V.; Obertova, J.; Palacka, P.; Rajec, J.; Sycova-Mila, Z.; et al. Prognostic value of intratumoral carbonic anhydrase IX expression in testicular germ cell tumors. Oncol. Lett. 2017, 13, 2177–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonatelli, M.; Silva, E.C.A.; Carcano, F.M.; Zaia, M.G.; Lopes, L.F.; Scapulatempo-Neto, C.; Pinheiro, C. The Warburg effect is associated with tumor aggressiveness in testicular germ cell tumors. Front. Endocrinol. 2019, 10, 417. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | TCGA (Cancer Tissue) | Expression in Testicular Cancer | Prognostic Value |
|---|---|---|---|
| 14-3-3β | All | High/Medium | Liver cancer Endometrial cancer Lung cancer Head and neck cancer Breast cancer Renal cancer |
| 14-3-3γ | All | High/Medium | Renal cancer Cervical cancer Lung cancer Pancreatic cancer |
| Caldesmon | All | Weak/Negative | Renal cancer Melanoma |
| Ezrin | All | High/Medium | Pancreatic cancer Renal cancer Urothelial cancer Colorectal cancer Renal cancer |
| Filamin A | All | High/Medium | |
| PARK7 | All | High/Medium | NA |
| Vimentin | All | High/Medium | Endometrial cancer Renal cancer |
| Vinculin | All | Medium/Weak | Pancreatic cancer |
| Variable | RTI-Positive | % | RTI-Negative | % |
|---|---|---|---|---|
| All patients | 37 | 100 | 37 | 100 |
| TNM staging system | ||||
| pT1pNx | 12 | 32.4 | 19 | 51.4 |
| pT2pNx | 24 | 64.9 | 18 | 48.6 |
| pT3pNx | 0 | 0 | 0 | 0 |
| pT4pNx | 1 | 2.7 | 0 | 0 |
| Tumour diameter | ||||
| <4 cm | 18 | 58.1 | 16 | 48.5 |
| ≥4 cm | 13 | 41.9 | 17 | 51.5 |
| Therapy modality | ||||
| Carboplatin | 18 | 48.6 | 14 | 37.8 |
| Radiation | 14 | 37.8 | 13 | 35.1 |
| Surveillance | 4 | 10.8 | 8 | 21.6 |
| None/No evidence | 1 | 2.7 | 2 | 5.4 |
| Therapy response | ||||
| Favourable (no disease progression) | 35 | 94.6 | 35 | 94.6 |
| Unfavourable | 1 | 2.7 | 0 | 0 |
| Unknown | 1 | 2.7 | 2 | 5.4 |
| Variable (Positivity Localization—Grade) | RTI-Positive | % | RTI-Negative | % | p Value |
|---|---|---|---|---|---|
| 14-3-3γ (TC positivity) | 0.047 | ||||
| 0 | 1 | 2.7 | 6 | 16.2 | |
| 2 | 36 | 97.3 | 31 | 83.8 | |
| Ezrin (IC positivity) | 0.021 | ||||
| 0 | 0 | 0 | 5 | 13.5 | |
| 2 | 37 | 100 | 32 | 86.5 | |
| Filamin A (TC-M/C-G) | 0.020 | ||||
| 0 | 23 | 62.2 | 13 | 35.1 | |
| 1 | 14 | 37.8 | 24 | 64.9 | |
| PARK7 (TC-C-G) | 0.034 | ||||
| 0 | 5 | 13.5 | 0 | 0 | |
| 1 | 12 | 32.4 | 9 | 24.3 | |
| 2 | 20 | 54.1 | 28 | 75.7 | |
| Vimentin (TC positivity) | 0.314 | ||||
| 0 | 37 | 100 | 36 | 97.3 | |
| 1 | 0 | 0 | 1 | 2.7 | |
| Vinculin (IC-G) | 0.152 | ||||
| 0 | 37 | 100 | 35 | 94.6 | |
| 1 | 0 | 0 | 2 | 5.4 |
| Variable | OR | 95% CI | p Value |
|---|---|---|---|
| Tumour diameter | 0.222 | 0.054–0.916 | 0.037 |
| TNM stage | 5.027 | 1.307–19.343 | 0.019 |
| 14-3-3γ | 3.488 | 1.392–8.737 | 0.008 |
| Filamin A | 0.215 | 0.059–0.778 | 0.019 |
| PARK7 | 0.313 | 0.118–0.831 | 0.020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borszéková Pulzová, L.; Roška, J.; Kalman, M.; Kliment, J.; Slávik, P.; Smolková, B.; Goffa, E.; Jurkovičová, D.; Kulcsár, Ľ.; Lešková, K.; et al. Screening for the Key Proteins Associated with Rete Testis Invasion in Clinical Stage I Seminoma via Label-Free Quantitative Mass Spectrometry. Cancers 2021, 13, 5573. https://doi.org/10.3390/cancers13215573
Borszéková Pulzová L, Roška J, Kalman M, Kliment J, Slávik P, Smolková B, Goffa E, Jurkovičová D, Kulcsár Ľ, Lešková K, et al. Screening for the Key Proteins Associated with Rete Testis Invasion in Clinical Stage I Seminoma via Label-Free Quantitative Mass Spectrometry. Cancers. 2021; 13(21):5573. https://doi.org/10.3390/cancers13215573
Chicago/Turabian StyleBorszéková Pulzová, Lucia, Jan Roška, Michal Kalman, Ján Kliment, Pavol Slávik, Božena Smolková, Eduard Goffa, Dana Jurkovičová, Ľudovít Kulcsár, Katarína Lešková, and et al. 2021. "Screening for the Key Proteins Associated with Rete Testis Invasion in Clinical Stage I Seminoma via Label-Free Quantitative Mass Spectrometry" Cancers 13, no. 21: 5573. https://doi.org/10.3390/cancers13215573