
Periostin Exon-21 Antibody Neutralization of Triple-Negative Breast Cancer Cell-Derived Periostin Regulates Tumor-Associated Macrophage Polarization and Angiogenesis

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Anti-Human Periostin Antibody
2.2. Cell Culture
2.3. Human Periostin Exon-21 Splicing Reporter
2.4. RNA Sequence
2.5. Cell Line-Derived Xenograft Model Studies in Mice
2.6. Histological Analysis
2.7. Quantitative Realtime PCR
2.8. Western Blot Analysis
2.9. PCR Primer List
2.10. Statistics
3. Results
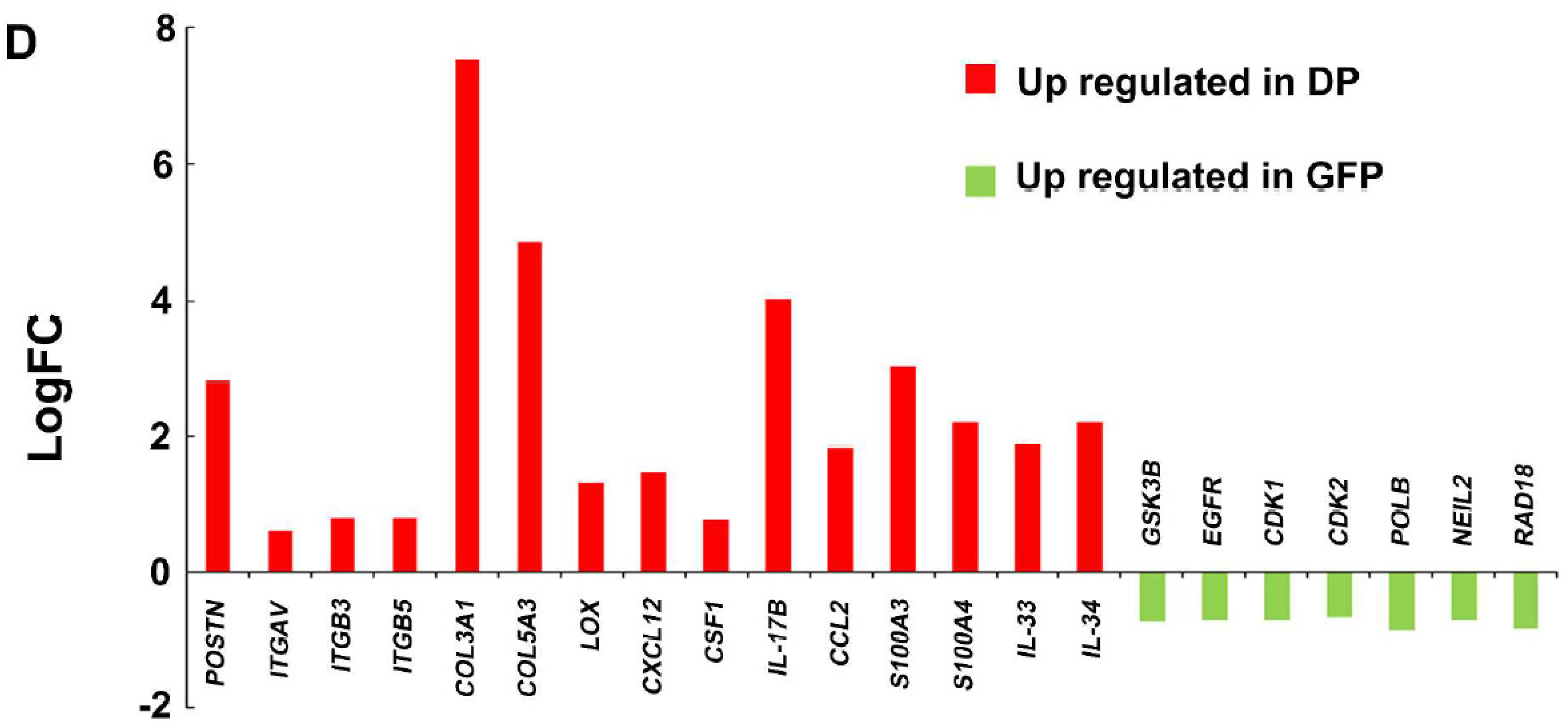
3.1. Development of SUM159PT TNBC Cells with A Fluorescence-Based Periostin Exon-21 Splicing Reporter
3.2. Characterization of GFP Single- and Double-Positive SUM159PT TNBC Cells
3.3. Antitumor Activity of Pn-21 Ab in Xenografts of SUM159PT Double-Positive Cells
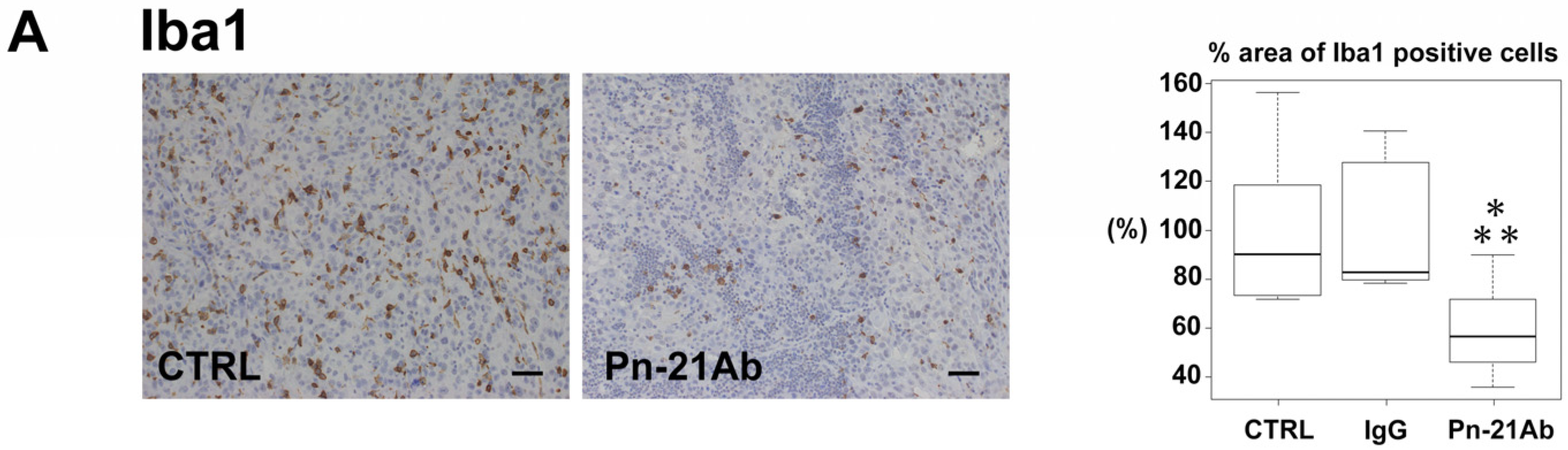
3.4. Pn Enhances TAM M2 Polarization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, H.; Ren, G. Epithelial-mesenchymal transition and drug resistance in breast cancer (Review). Int. J. Oncol. 2015, 47, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Higgins, M.J.; Baselga, J. Targeted therapies for breast cancer. J. Clin. Investig. 2011, 121, 3797–3803. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, Y.; Taniyama, Y.; Sanada, F.; Morishita, R.; Nakamori, S.; Morimoto, K.; Yeung, K.T.; Yang, J. Periostin blockade overcomes chemoresistance via restricting the expansion of mesenchymal tumor subpopulations in breast cancer. Sci. Rep. 2018, 8, 4013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniyama, Y.; Katsuragi, N.; Sanada, F.; Azuma, J.; Iekushi, K.; Koibuchi, N.; Okayama, K.; Ikeda-Iwabu, Y.; Muratsu, J.; Otsu, R.; et al. Selective Blockade of Periostin Exon 17 Preserves Cardiac Performance in Acute Myocardial Infarction. Hypertension 2016, 67, 356–361. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nishiyama, T.; Shimizu, K.; Kadota, K. TCC: An R package for comparing tag count data with robust normalization strategies. BMC Bioinform. 2013, 14, 219. [Google Scholar] [CrossRef] [Green Version]
- Sanada, F.; Muratsu, J.; Otsu, R.; Shimizu, H.; Koibuchi, N.; Uchida, K.; Taniyama, Y.; Yoshimura, S.; Rakugi, H.; Morishita, R. Local Production of Activated Factor X in Atherosclerotic Plaque Induced Vascular Smooth Muscle Cell Senescence. Sci. Rep. 2017, 7, 17172. [Google Scholar] [CrossRef] [Green Version]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Lambert, A.W.; Wong, C.K.; Ozturk, S.; Papageorgis, P.; Raghunathan, R.; Alekseyev, Y.; Gower, A.C.; Reinhard, B.M.; Abdolmaleky, H.M.; Thiagalingam, S. Tumor Cell-Derived Periostin Regulates Cytokines That Maintain Breast Cancer Stem Cells. Mol. Cancer Res. 2016, 14, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Ringnér, M.; Fredlund, E.; Häkkinen, J.; Borg, Å.; Staaf, J. GOBO: Gene Expression-Based Outcome for Breast Cancer Online. PLoS ONE 2011, 6, e17911. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liu, J.; Wang, Z.; Huang, Y.; Liu, W.; Zhu, X.; Cai, Y.; Fang, X.; Lin, S.; Yuan, L.; et al. Periostin Contributes to the Acquisition of Multipotent Stem Cell-Like Properties in Human Mammary Epithelial Cells and Breast Cancer Cells. PLoS ONE 2013, 8, e72962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Tzanakakis, G.; Kavasi, R.-M.; Voudouri, K.; Berdiaki, A.; Spyridaki, I.; Tsatsakis, A.; Nikitovic, D. Role of the extracellular matrix in cancer-associated epithelial to mesenchymal transition phenomenon. Dev. Dyn. 2018, 247, 368–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laoui, D.; Movahedi, K.; Van Overmeire, E.; Bossche, J.V.D.; Schouppe, E.; Mommer, C.; Nikolaou, A.; Morias, Y.; De Baetselier, P.; Van Ginderachter, J. Tumor-associated macrophages in breast cancer: Distinct subsets, distinct functions. Int. J. Dev. Biol. 2011, 55, 861–867. [Google Scholar] [CrossRef]
- Lindsten, T.; Hedbrant, A.; Ramberg, A.; Wijkander, J.; Solterbeck, A.; Eriksson, M.; Delbro, D.; Erlandsson, A. Effect of macrophages on breast cancer cell proliferation, and on expression of hormone receptors, uPAR and HER-2. Int. J. Oncol. 2017, 51, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.-Q.; Waaijer, S.J.H.; Zwager, M.C.; De Vries, E.G.E.; Van Der Vegt, B.; Schröder, C.P. Tumor-associated macrophages in breast cancer: Innocent bystander or important player? Cancer Treat. Rev. 2018, 70, 178–189. [Google Scholar] [CrossRef] [Green Version]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-negative breast cancer: Challenges and opportunities of a heterogeneous disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef]
- Place, A.E.; Huh, S.J.; Polyak, K. The microenvironment in breast cancer progression: Biology and implications for treatment. Breast Cancer Res. 2011, 13, 227. [Google Scholar] [CrossRef] [Green Version]
- Pearce, O.M.T.; Delaine-Smith, R.M.; Maniati, E.; Nichols, S.; Wang, J.; Böhm, S.; Rajeeve, V.; Ullah, D.; Chakravarty, P.; Jones, R.R.; et al. Deconstruction of a Metastatic Tumor Micro-environment Reveals a Common Matrix Response in Human Cancers. Cancer Discov. 2018, 8, 304–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.; Hoersch, S.; Liu, H.; Carr, S.A.; Hynes, R.O. The Matrisome: In Silico Definition and In Vivo Characterization by Proteomics of Normal and Tumor Extracellular Matrices. Mol. Cell. Proteom. 2012, 11, m111.014647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Wu, K.; Wang, X.; Zhang, J.; Wang, L.; Jiang, Y.; Zhu, X.; Chen, W.; Yan, M. Periostin secreted by cancer-associated fibroblasts promotes cancer stemness in head and neck cancer by activating protein tyrosine kinase 7. Cell Death Dis. 2018, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Chen, X.; Liang, L.; Yu, L.; Wu, X.; Zhou, C.; Wang, Z.; Fan, L.; Hu, Z.; Liang, L.; et al. Periostin + cancer-associated fibroblasts promote lymph node metastasis by impairing the lymphatic endothelial barriers in cervical squamous cell carcinoma. Mol. Oncol. 2021, 15, 210–227. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, S.S.; Pinto, M.C.X.; Amorim, J.H.; Azevedo, V.A.C.; Resende, R.R.; Mintz, A.; Birbrair, A. Glioma Pericytes Promote Angio-genesis by Producing Periostin. Cell Mol. Neurobiol. 2020, 3, 1–8. [Google Scholar]
- Keklikoglou, I.; Kadioglu, E.; Bissinger, S.; Langlois, B.; Bellotti, A.; Orend, G.; Ries, C.H.; De Palma, M. Periostin Limits Tumor Response to VEGFA Inhibition. Cell Rep. 2018, 22, 2530–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-González, L.; Alonso, J. Periostin: A Matricellular Protein With Multiple Functions in Cancer Development and Pro-gression. Front. Oncol. 2018, 8, 225. [Google Scholar] [CrossRef]
- Jayasingam, S.D.; Citartan, M.; Thang, T.H.; Zin, A.A.M.; Ang, K.C.; Ch’Ng, E.S. Evaluating the Polarization of Tumor-Associated Macrophages Into M1 and M2 Phenotypes in Human Cancer Tissue: Technicalities and Challenges in Routine Clinical Practice. Front. Oncol. 2020, 9, 1512. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Xiong, S.; Mao, Y.; Chen, M.; Ma, X.; Zhou, X.; Ma, Z.; Liu, F.; Huang, Z.; Luo, Q.; et al. Periostin promotes immuno-suppressive premetastatic niche formation to facilitate breast tumour metastasis. J. Pathol. 2016, 239, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ke, S.Q.; Huang, Z.; Flavahan, W.; Fang, X.; Paul, J.; Wu, L.; Sloan, A.E.; McLendon, R.E.; Li, X.; et al. Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat. Cell Biol. 2015, 17, 170–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward (5′ to 3′) | Reverse (5′ to 3′) |
|---|---|---|
| periostin exon 21 | GGTCACCAAGGTCACCAAATTC | CCTGAAGTCAACTTGGCTCTCAC |
| periostin exon20/22 | GTTACAAGAAGACACACCCGTG | CCTGAAGTCAACTTGGCTCTCAC |
| GAPDH | GGATTTGGTCGTATTGGG | GGAAGATGGTGATGGGATT |
| ITGAV | AGGAGAAGGTGCCTACGAAGCT | GCACAGGAAAGTCTTGCTAAGGC |
| ITGB3 | CATGGATTCCAGCAATGTCCTCC | TTGAGGCAGGTGGCATTGAAGG |
| ITGB5 | GCCTTTCTGTGAGTGCGACAAC | CCGATGTAACCTGCTGGCACT |
| COL3A1 | TGGTCTGCAAGGAATGCCTGGA | TCTTTCCCTGGGACACCATCAG |
| COL5A3 | GAGAGGAGAACTGGGCTTCCAA | TAGAGGTCCCACTTCTCCTGTC |
| LOX | ACAATGTTGTGCGCTGTGAC | CCACTTCAGAACACCAGGCA |
| CXCL12 | CTCAACACTCCAAACTGTGCCC | CTCCAGGTACTCCTGAATCCAC |
| CSF1 | TGAGACACCTCTCCAGTTGCTG | GCAATCAGGCTTGGTCACCACA |
| IL-17A | GCTGTGGATGTCCAACAAGAGG | TCCTGCATGGTGAAGGGGTTCA |
| CCL2 | CCCCAGTCACCTGCTGTTAT | TGGAATCCTGAACCCACTTC |
| S100A3 | CAAATACAAGCTCTGCCAGGCG | TCGCAGTCCTTGTTGGTGTCCA |
| S100A4 | CAGAACTAAAGGAGCTGCTGACC | CTTGGAAGTCCACCTCGTTGTC |
| IL-33 | GCCTGTCAACAGCAGTCTACTG | TGTGCTTAGAGAAGCAAGATACTC |
| IL-34 | CCAAGGTGGAATCCGTGTTGTC | CACCTCACAGTCCTGCCAGTTT |
| E-cadherin | CTGAGAACGAGGCTAACG | TTCACATCCAGCACATCC |
| N-cadherin | GCTGATAGCCCGGTTTCACT | CCCAGGCTTTGATCCCTCTG |
| Vimentin | GGACCAGCTAACCAACGACA | AAGGTCAAGACGTGCCAGAG |
| Twist | TACGCCTTCTCGGTCTGGA | ACTGTCCATTTTCTCCTTCTCTGG |
| Snail 1 | GAGCTGACCTCCCTGTCAGA | GGCCTCCAAGGAAGAGACTG |
| Snail 2 | GGCTGGCCAAACATAAGCAG | TTGCCGCAGATCTTGCAAAC |
| Zeb 1 | GTGGCGGTAGATGGTAAT | CTGTTTGTAGCGACTGGA |
| Zeb 2 | ACCAGCCCTTTAGGAGTT | AGACCGACAGGCGGAATA |
| iNOS | GCTCTACACCTCCAATGTGACC | CTGCCGAGATTTGAGCCTCATG |
| IL-1b | CCACAGACCTTCCAGGAGAATG | GTGCAGTTCAGTGATCGTACAGG |
| IL-12a | TGCCTTCACCACTCCCAAAACC | CAATCTCTTCAGAAGTGCAAGGG |
| CD206 | CTCTGTTCAGCTATTGGACGC | CGGAATTTCTGGGATTCAGCTTC |
| CD163 | CCAGAAGGAACTTGTAGCCACAG | CAGGCACCAAGCGTTTTGAGCT |
| Fizz1 | GCAAGAAGCTCTCGTGTGCTA | AACATCCCACGAACCACAGCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujikawa, T.; Sanada, F.; Taniyama, Y.; Shibata, K.; Katsuragi, N.; Koibuchi, N.; Akazawa, K.; Kanemoto, Y.; Kuroyanagi, H.; Shimazu, K.; et al. Periostin Exon-21 Antibody Neutralization of Triple-Negative Breast Cancer Cell-Derived Periostin Regulates Tumor-Associated Macrophage Polarization and Angiogenesis. Cancers 2021, 13, 5072. https://doi.org/10.3390/cancers13205072
Fujikawa T, Sanada F, Taniyama Y, Shibata K, Katsuragi N, Koibuchi N, Akazawa K, Kanemoto Y, Kuroyanagi H, Shimazu K, et al. Periostin Exon-21 Antibody Neutralization of Triple-Negative Breast Cancer Cell-Derived Periostin Regulates Tumor-Associated Macrophage Polarization and Angiogenesis. Cancers. 2021; 13(20):5072. https://doi.org/10.3390/cancers13205072
Chicago/Turabian StyleFujikawa, Tatsuya, Fumihiro Sanada, Yoshiaki Taniyama, Kana Shibata, Naruto Katsuragi, Nobutaka Koibuchi, Kaori Akazawa, Yuko Kanemoto, Hidehito Kuroyanagi, Kenzo Shimazu, and et al. 2021. "Periostin Exon-21 Antibody Neutralization of Triple-Negative Breast Cancer Cell-Derived Periostin Regulates Tumor-Associated Macrophage Polarization and Angiogenesis" Cancers 13, no. 20: 5072. https://doi.org/10.3390/cancers13205072