
Molecular Changes Induced in Melanoma by Cell Culturing in 3D Alginate Hydrogels

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture Conditions
2.2. Fluorescence Microscopy
2.3. Cell Cycle Quantification
2.4. Proliferation Analysis
2.5. Dynamic Mechanical Analysis (DMA)
2.6. RNA Isolation for RNA-Seq
2.7. RNA-Seq Library Preparation and Mapping
2.8. RNA-Seq Data Analysis
2.9. miRNA Transfection
2.10. mRNA Isolation and Reverse Transcription
2.11. miRNA Isolation and Reverse Transcription
2.12. Analysis of Gene and microRNA Expression by Quantitative PCR
2.13. In Silico Analysis
2.14. Statistical Analysis of Experimental Data
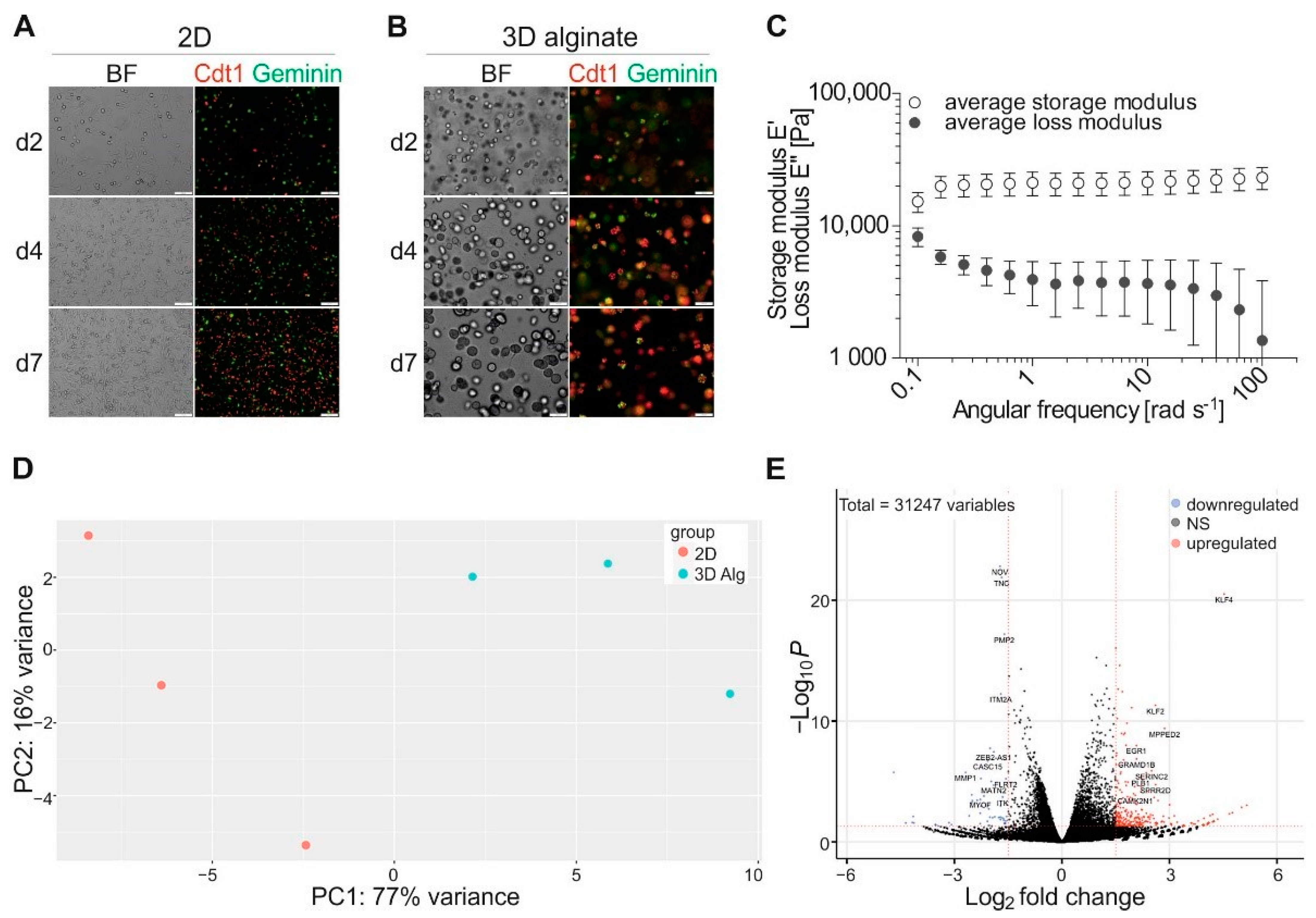
3. Results
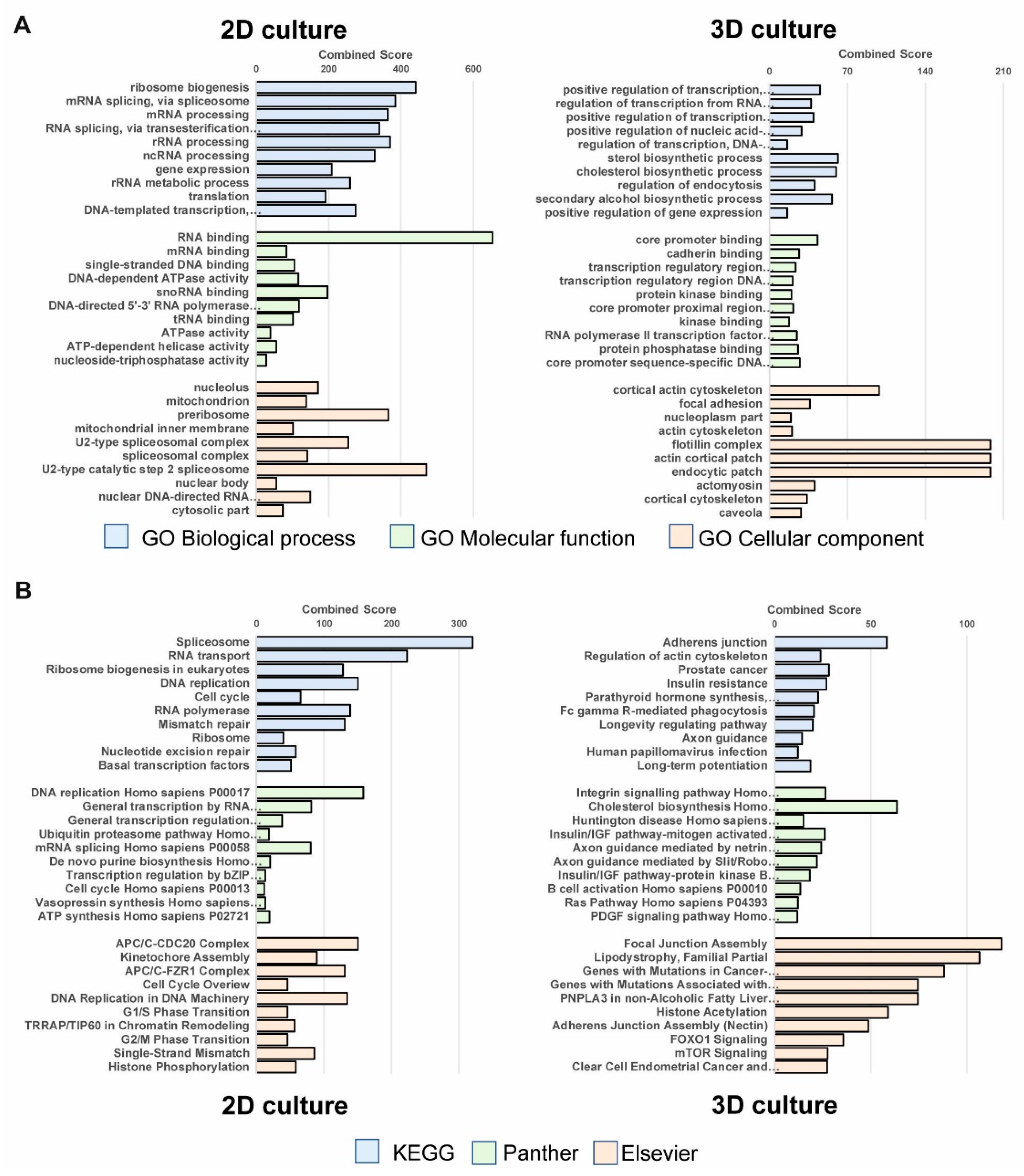
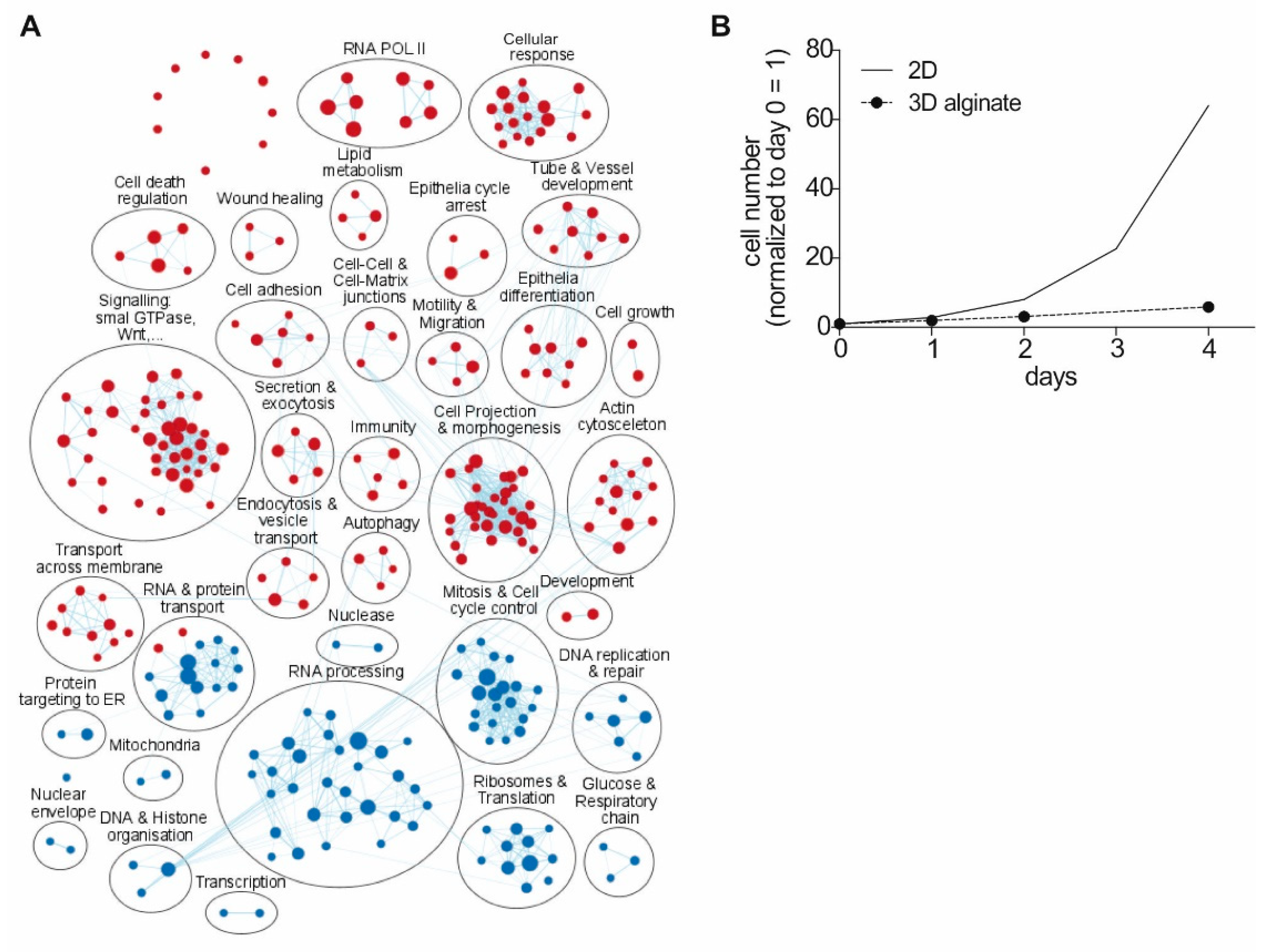
3.1. Functional Clustering of Differential Expressed Genes Reveals High Impact of Cell Culture Conditions on Gene Expression
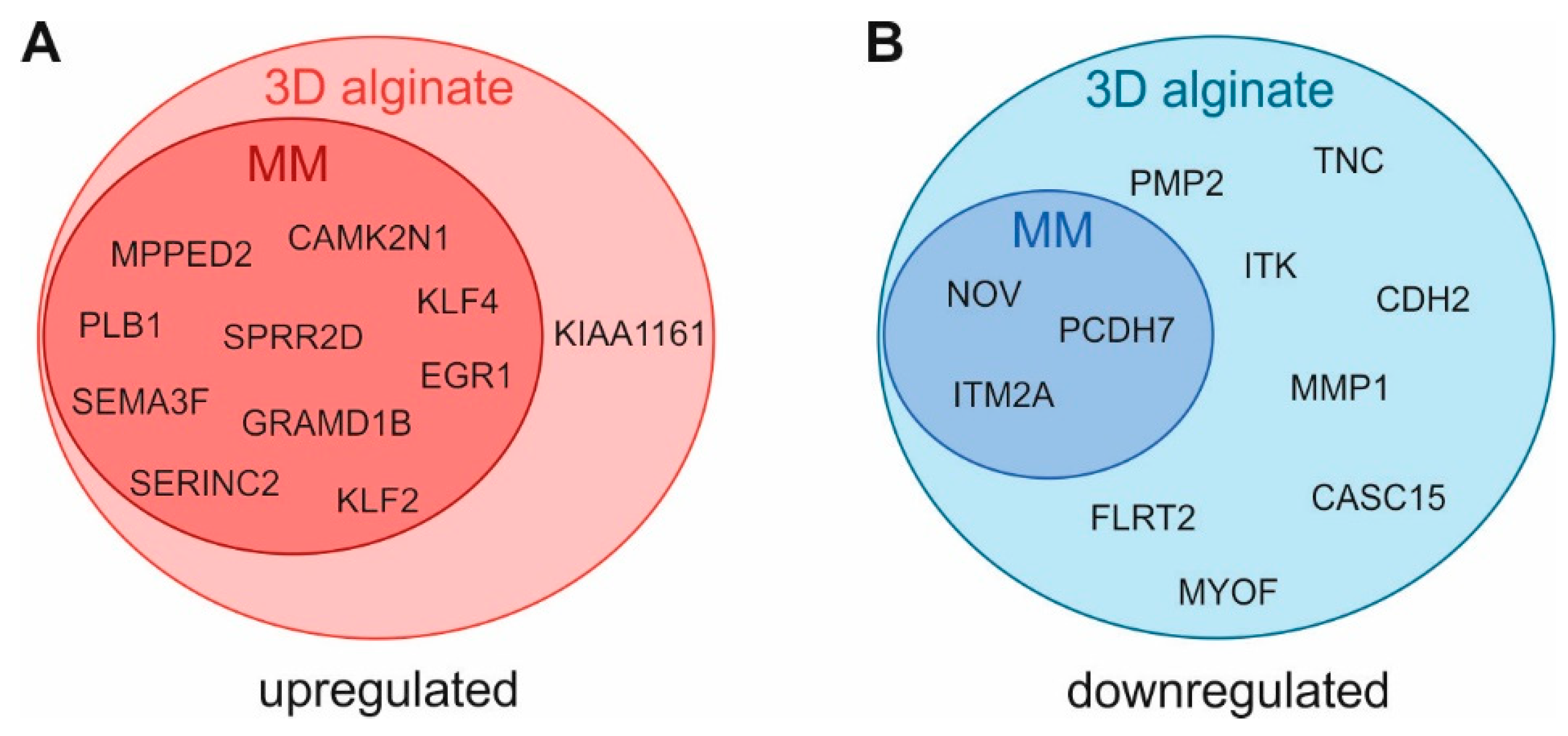
3.2. Regulation of Gene Expression Level by 3D Cultivation
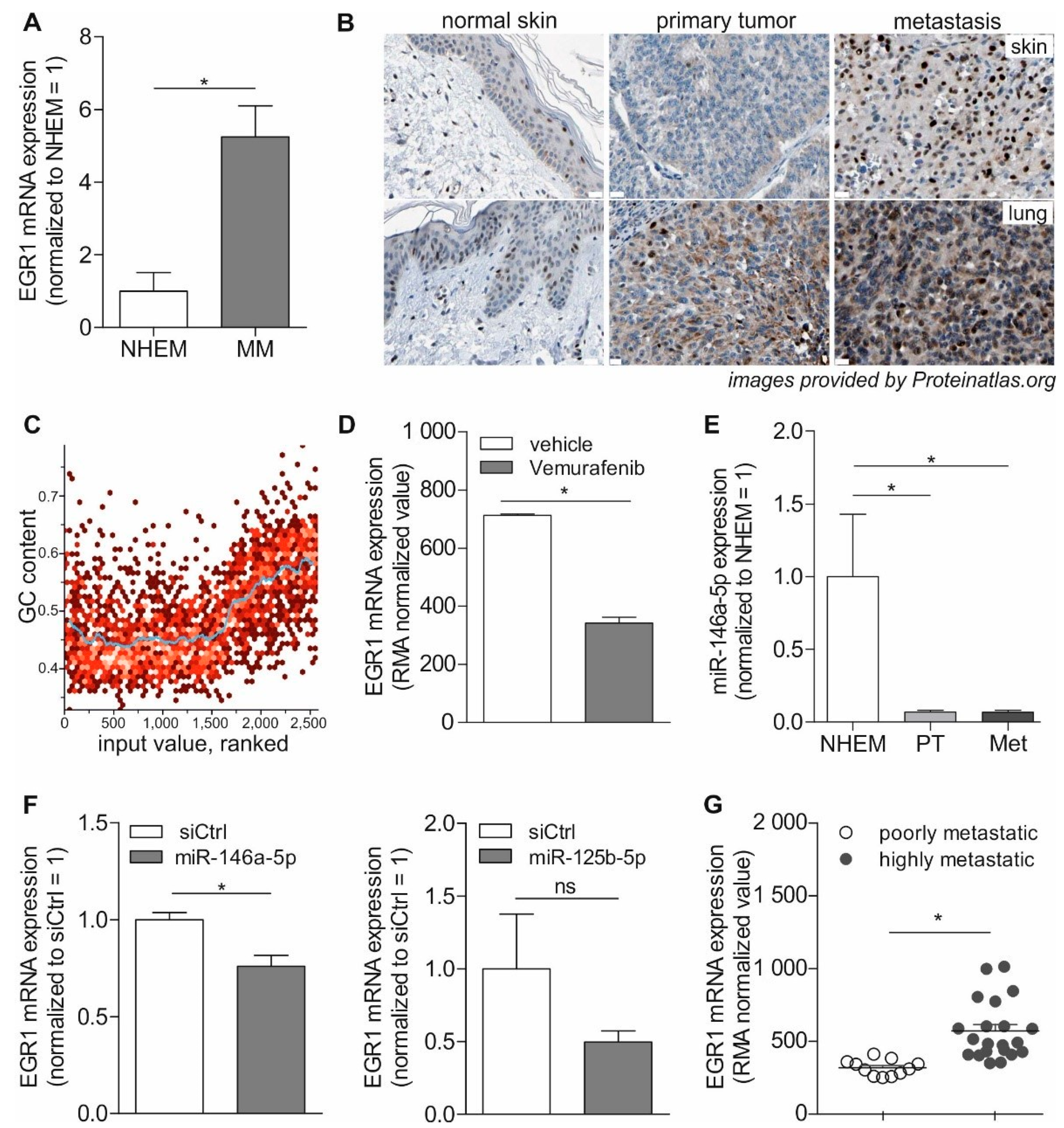
3.3. Role of EGR1 in Melanoma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fontoura, J.C.; Viezzer, C.; Dos Santos, F.G.; Ligabue, R.A.; Weinlich, R.; Puga, R.D.; Antonow, D.; Severino, P.; Bonorino, C. Comparison of 2D and 3D cell culture models for cell growth, gene expression and drug resistance. Mater. Sci. Eng. C 2020, 107, 110264. [Google Scholar] [CrossRef]
- Gruber, H.E.; Hanley, E.N. Human disc cells in monolayer vs 3D culture: Cell shape, division and matrix formation. BMC Musculoskelet. Disord. 2000, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Loessner, D.; Stok, K.S.; Lutolf, M.P.; Hutmacher, D.W.; Clements, J.A.; Rizzi, S.C. Bioengineered 3D platform to explore cell–ECM interactions and drug resistance of epithelial ovarian cancer cells. Biomaterials 2010, 31, 8494–8506. [Google Scholar] [CrossRef] [Green Version]
- Petersen, O.W.; Rønnov-Jessen, L.; Howlett, A.R.; Bissell, M.J. Interaction with basement membrane serves to rapidly distinguish growth and differentiation pattern of normal and malignant human breast epithelial cells. Proc. Natl. Acad. Sci. USA 1992, 89, 9064–9068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickman, J.A.; Graeser, R.; de Hoogt, R.; Vidic, S.; Brito, C.; Gutekunst, M.; van der Kuip, H.; IMI PREDECT consortium. Three-dimensional models of cancer for pharmacology and cancer cell biology: Capturing tumor complexity in vitro/ex vivo. Biotechnol. J. 2014, 9, 1115–1128. [Google Scholar] [CrossRef]
- Klöck, G.; Pfeffermann, A.; Ryser, C.; Gröhn, P.; Kuttler, B.; Hahn, H.-J.; Zimmermann, U. Biocompatibility of mannuronic acid-rich alginates. Biomaterials 1997, 18, 707–713. [Google Scholar] [CrossRef]
- Rowley, J.A.; Madlambayan, G.; Mooney, D.J. Alginate hydrogels as synthetic extracellular matrix materials. Biomaterials 1999, 20, 45–53. [Google Scholar] [CrossRef]
- Neves, M.I.; Moroni, L.; Barrias, C.C. Modulating alginate hydrogels for improved biological performance as cellular 3D microenvironments. Front. Bioeng. Biotechnol. 2020, 8, 665. [Google Scholar] [CrossRef]
- Ivanovska, J.; Zehnder, T.; Lennert, P.; Sarker, B.; Boccaccini, A.R.; Hartmann, A.; Schneider-Stock, R.; Detsch, R. Biofabrication of 3D alginate-based hydrogel for cancer research: Comparison of cell spreading, viability, and adhesion characteristics of colorectal HCT116 tumor cells. Tissue Eng. Part C Methods 2016, 22, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Read, G.H.; Miura, N.; Carter, J.L.; Kines, K.T.; Yamamoto, K.; Devasahayam, N.; Cheng, J.Y.; Camphausen, K.A.; Krishna, M.C.; Kesarwala, A.H. Three-dimensional alginate hydrogels for radiobiological and metabolic studies of cancer cells. Colloids Surf. B Biointerfaces 2018, 171, 197–204. [Google Scholar] [CrossRef]
- Smit, T.; Calitz, C.; Willers, C.; Svitina, H.; Hamman, J.; Fey, S.J.; Gouws, C.; Wrzesinski, K. Characterization of an alginate encapsulated LS180 spheroid model for anti-colorectal cancer compound screening. ACS Med. Chem. Lett. 2020, 11, 1014–1021. [Google Scholar] [CrossRef]
- Cavo, M.; Fato, M.; Peñuela, L.; Beltrame, F.; Raiteri, R.; Scaglione, S. Microenvironment complexity and matrix stiffness regulate breast cancer cell activity in a 3D in vitro model. Sci. Rep. 2016, 6, 35367. [Google Scholar] [CrossRef] [Green Version]
- Schmid, R.; Schmidt, S.K.; Hazur, J.; Detsch, R.; Maurer, E.; Boccaccini, A.R.; Hauptstein, J.; Teßmar, J.; Blunk, T.; Schrüfer, S. Comparison of Hydrogels for the Development of Well-Defined 3D Cancer Models of Breast Cancer and Melanoma. Cancers 2020, 12, 2320. [Google Scholar] [CrossRef]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szczesniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Kappelmann-Fenzl, M.; Gebhard, C.; Matthies, A.O.; Kuphal, S.; Rehli, M.; Bosserhoff, A.K. C-Jun drives melanoma progression in PTEN wild type melanoma cells. Cell Death Dis. 2019, 10, 584. [Google Scholar] [CrossRef] [Green Version]
- Scolyer, R.A.; Rawson, R.V.; Gershenwald, J.E.; Ferguson, P.M.; Prieto, V.G. Melanoma pathology reporting and staging. Mod. Pathol. 2020, 33, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Zbytek, B.; Carlson, J.A.; Granese, J.; Ross, J.; Mihm, M.; Slominski, A. Current concepts of metastasis in melanoma. Expert Rev. Dermatol. 2008, 3, 569–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linck, L.; Liebig, J.; Voeller, D.; Eichner, N.; Lehmann, G.; Meister, G.; Bosserhoff, A. MicroRNA-sequencing data analyzing melanoma development and progression. Exp. Mol. Pathol. 2018, 105, 371–379. [Google Scholar] [CrossRef]
- Feuerer, L.; Lamm, S.; Henz, I.; Kappelmann-Fenzl, M.; Haferkamp, S.; Meierjohann, S.; Hellerbrand, C.; Kuphal, S.; Bosserhoff, A.K. Role of melanoma inhibitory activity in melanocyte senescence. Pigment Cell Melanoma Res. 2019, 32, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.-B.; Mascalchi, P.; Rodriguez, E.; Lin, Y.; Jodrell, D.I.; Richards, F.M.; Lyons, S.K. A quantitative FastFUCCI assay defines cell cycle dynamics at a single-cell level. J. Cell Sci. 2017, 130, 512–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Simms, D.; Cizdziel, P.E.; Chomczynski, P. TRIzol: A new reagent for optimal single-step isolation of RNA. Focus 1993, 15, 532–535. [Google Scholar]
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 22 February 2021).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GSEA|MSigDB. Available online: http://www.gsea-msigdb.org/gsea/msigdb/index.jsp (accessed on 22 February 2021).
- Montojo, J.; Zuberi, K.; Rodriguez, H.; Kazi, F.; Wright, G.; Donaldson, S.L.; Morris, Q.; Bader, G.D. GeneMANIA Cytoscape plugin: Fast gene function predictions on the desktop. Bioinformatics 2010, 26, 2927–2928. [Google Scholar] [CrossRef]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment map: A network-based method for gene-set enrichment visualization and interpretation. PLoS ONE 2010, 5, e13984. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 22 February 2021).
- Dietrich, P.; Kuphal, S.; Spruss, T.; Hellerbrand, C.; Bosserhoff, A.K. Wild-type KRAS is a novel therapeutic target for melanoma contributing to primary and acquired resistance to BRAF inhibition. Oncogene 2018, 37, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Schiffner, S.; Braunger, B.M.; de Jel, M.M.; Coupland, S.E.; Tamm, E.R.; Bosserhoff, A.K. Tg (Grm1) transgenic mice: A murine model that mimics spontaneous uveal melanoma in humans? Exp. Eye Res. 2014, 127, 59–68. [Google Scholar] [CrossRef]
- Ott, C.A.; Linck, L.; Kremmer, E.; Meister, G.; Bosserhoff, A.K. Induction of exportin-5 expression during melanoma development supports the cellular behavior of human malignant melanoma cells. Oncotarget 2016, 7, 62292. [Google Scholar] [CrossRef]
- GDS5085/8108370. Available online: https://www.ncbi.nlm.nih.gov/geo/tools/profileGraph.cgi?ID=GDS5085:8108370 (accessed on 15 February 2021).
- GDS3964/201694_s_at. Available online: https://www.ncbi.nlm.nih.gov/geo/tools/profileGraph.cgi?ID=GDS3964:201694_s_at (accessed on 15 February 2021).
- Naik, J.B.; Mishra, S. The compatibilizing effect of maleic anhydride on swelling properties of plant-fiber-reinforced polystyrene composites. Polym. Plast. Technol. Eng. 2006, 45, 923–927. [Google Scholar] [CrossRef]
- Menard, K.P. Dynamic Mechanical Analysis: A Practical Introduction; CRC Press: Boca Raton, FL, USA, 2008; ISBN 9781420053128. [Google Scholar]
- Kappelmann-Fenzl, M. Next Generation Sequencing and Data Analysis; Springer: Heidelberg, Germany, 2021; ISBN 9783030624897. [Google Scholar]
- Chen, E.Y. Enrichr. Available online: https://maayanlab.cloud/Enrichr/enrich?dataset=a0dc40ec16fe3a7872096e25c2d48713 (accessed on 12 February 2021).
- Chen, E.Y. Enrichr. Available online: https://maayanlab.cloud/Enrichr/enrich?dataset=880e2b2f8a6de05d98bd6fa689c974af (accessed on 12 February 2021).
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The molecular signatures database hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Taddei, M.L.; Giannoni, E.; Morandi, A.; Ippolito, L.; Ramazzotti, M.; Callari, M.; Gandellini, P.; Chiarugi, P. Mesenchymal to amoeboid transition is associated with stem-like features of melanoma cells. Cell Commun. Signal. 2014, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Fane, M.E.; Chhabra, Y.; Smith, A.G.; Sturm, R.A. BRN 2, a POU erful driver of melanoma phenotype switching and metastasis. Pigment Cell Melanoma Res. 2019, 32, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Hoek, K.S.; Goding, C.R. Cancer stem cells versus phenotype-switching in melanoma. Pigment Cell Melanoma Res. 2010, 23, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, A.; Xu, H.; Krishnan, J.; Berger, S.I.; Mazloom, A.R.; Ma’ayan, A. ChEA: Transcription factor regulation inferred from integrating genome-wide ChIP-X experiments. Bioinformatics 2010, 26, 2438–2444. [Google Scholar] [CrossRef]
- Dietrich, P.; Kuphal, S.; Spruss, T.; Hellerbrand, C.; Bosserhoff, A.K. Micro RNA-622 is a novel mediator of tumorigenicity in melanoma by targeting Kirsten rat sarcoma. Pigment Cell Melanoma Res. 2018, 31, 614–629. [Google Scholar] [CrossRef]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 18 February 2021).
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- Expression of EGR1 in Melanoma—The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000120738-EGR1/pathology/melanoma#img (accessed on 18 February 2021).
- Tissue Expression of EGR1—Staining in Skin—The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000120738-EGR1/tissue/skin#img (accessed on 18 February 2021).
- Parmenter, T.J.; Kleinschmidt, M.; Kinross, K.M.; Bond, S.T.; Li, J.; Kaadige, M.R.; Rao, A.; Sheppard, K.E.; Hugo, W.; Pupo, G.M.; et al. Response of BRAF-mutant melanoma to BRAF inhibition is mediated by a network of transcriptional regulators of glycolysis. Cancer Discov. 2014, 4, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Shen, S.S.; Hoshida, Y.; Subramanian, A.; Ross, K.; Brunet, J.-P.; Wagner, S.N.; Ramaswamy, S.; Mesirov, J.P.; Hynes, R.O. Gene expression changes in an animal melanoma model correlate with aggressiveness of human melanoma metastases. Mol. Cancer Res. 2008, 6, 760–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GDS5670/8108370. Available online: https://www.ncbi.nlm.nih.gov/geo/tools/profileGraph.cgi?ID=GDS5670:8108370 (accessed on 15 February 2021).
- Contreras, J.R.; Palanichamy, J.K.; Tran, T.M.; Fernando, T.R.; Rodriguez-Malave, N.I.; Goswami, N.; Arboleda, V.A.; Casero, D.; Rao, D.S. MicroRNA-146a modulates B-cell oncogenesis by regulating Egr1. Oncotarget 2015, 6, 11023. [Google Scholar] [CrossRef]
- Mueller, D.W.; Rehli, M.; Bosserhoff, A.K. miRNA expression profiling in melanocytes and melanoma cell lines reveals miRNAs associated with formation and progression of malignant melanoma. J. Investig. Dermatol. 2009, 129, 1740–1751. [Google Scholar] [CrossRef] [Green Version]
- Kappelmann, M.; Kuphal, S.; Meister, G.; Vardimon, L.; Bosserhoff, A.K. MicroRNA miR-125b controls melanoma progression by direct regulation of c-Jun protein expression. Oncogene 2013, 32, 2984–2991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Liu, K.; Xiang, Z.; Yu, X.; Wang, P.; Ma, Q. MiR-125b-2-3p associates with prognosis of ccRCC through promoting tumor metastasis via targeting EGR1. Am. J. Transl. Res. 2020, 12, 5575–5585. [Google Scholar] [PubMed]
- Zhang, S.; Tao, X.; Cao, Q.; Feng, X.; Wu, J.; Yu, H.; Yu, Y.; Xu, C.; Zhao, H. lnc003875/miR-363/EGR1 regulatory network in the carcinoma-associated fibroblasts controls the angiogenesis of human placental site trophoblastic tumor (PSTT). Exp. Cell Res. 2020, 387, 111783. [Google Scholar] [CrossRef]
- Wu, X.; Cheng, J.; Li, P.; Yang, M.; Qiu, S.; Liu, P.; Du, J. Mechano-sensitive transcriptional factor Egr-1 regulates insulin-like growth factor-1 receptor expression and contributes to neointima formation in vein grafts. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.-S.; You, R.-I.; Cheng, C.-C.; Lee, M.-C.; Lin, T.-Y.; Hu, C.-T. Snail collaborates with EGR-1 and SP-1 to directly activate transcription of MMP 9 and ZEB1. Sci. Rep. 2017, 7, 12472. [Google Scholar] [CrossRef]
- Darnell, M.; Gu, L.; Mooney, D. RNA-seq reveals diverse effects of substrate stiffness on mesenchymal stem cells. Biomaterials 2018, 181, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Darnell, M.; O’Neil, A.; Mao, A.; Gu, L.; Rubin, L.L.; Mooney, D.J. Material microenvironmental properties couple to induce distinct transcriptional programs in mammalian stem cells. Proc. Natl. Acad. Sci. USA 2018, 115, E8368–E8377. [Google Scholar] [CrossRef] [Green Version]
- Winkelmann, R.; Sandrock, L.; Kirberg, J.; Jäck, H.-M.; Schuh, W. KLF2–a negative regulator of pre-B cell clonal expansion and B cell activation. PLoS ONE 2014, 9, e97953. [Google Scholar] [CrossRef]
- Li, T.-T.; Liu, M.-R.; Pei, D.-S. Friend or foe, the role of EGR-1 in cancer. Med. Oncol. 2020, 37, 7. [Google Scholar] [CrossRef]
- Bosserhoff, A.K. Melanoma Development; Springer: Heidelberg, Germany, 2017; ISBN 3319413198. [Google Scholar]
- Graf, S.A.; Heppt, M.V.; Wessely, A.; Krebs, S.; Kammerbauer, C.; Hornig, E.; Strieder, A.; Blum, H.; Bosserhoff, A.-K.; Berking, C. The myelin protein PMP2 is regulated by SOX10 and drives melanoma cell invasion. Pigment Cell Melanoma Res. 2019, 32, 424–434. [Google Scholar] [CrossRef]
- Hashimoto, H.; Olanrewaju, Y.O.; Zheng, Y.; Wilson, G.G.; Zhang, X.; Cheng, X. Wilms tumor protein recognizes 5-carboxylcytosine within a specific DNA sequence. Genes Dev. 2014, 28, 2304–2313. [Google Scholar] [CrossRef] [Green Version]
- Zandarashvili, L.; White, M.A.; Esadze, A.; Iwahara, J. Structural impact of complete CpG methylation within target DNA on specific complex formation of the inducible transcription factor Egr-1. FEBS Lett. 2015, 589, 1748–1753. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.-P.; Liu, C.; Fan, Y.; Mercola, D.; Adamson, E.D. Egr-1 negatively regulates human tumor cell growth via the DNA-binding domain. Cancer Res. 1995, 55, 5054–5062. [Google Scholar] [PubMed]
- Morawietz, H.; Ma, Y.-H.; Vives, F.; Wilson, E.; Sukhatme, V.P.; Holtz, J.; Ives, H.E. Rapid induction and translocation of Egr-1 in response to mechanical strain in vascular smooth muscle cells. Circ. Res. 1999, 84, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stula, M.; Orzechowski, H.-D.; Gschwend, S.; Vetter, R.; von Harsdorf, R.; Dietz, R.; Paul, M. Influence of sustained mechanical stress on Egr-1 mRNA expression in cultured human endothelial cells. Mol. Cell. Biochem. 2000, 210, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kunnen, S.J.; Malas, T.B.; Semeins, C.M.; Bakker, A.D.; Peters, D.J.M. Comprehensive transcriptome analysis of fluid shear stress altered gene expression in renal epithelial cells. J. Cell. Physiol. 2018, 233, 3615–3628. [Google Scholar] [CrossRef] [Green Version]
- Bougault, C.; Aubert-Foucher, E.; Paumier, A.; Perrier-Groult, E.; Huot, L.; Hot, D.; Duterque-Coquillaud, M.; Mallein-Gerin, F. Dynamic compression of chondrocyte-agarose constructs reveals new candidate mechanosensitive genes. PLoS ONE 2012, 7, e36964. [Google Scholar] [CrossRef] [Green Version]
- Tarcic, G.; Avraham, R.; Pines, G.; Amit, I.; Shay, T.; Lu, Y.; Zwang, Y.; Katz, M.; Ben-Chetrit, N.; Jacob-Hirsch, J. EGR1 and the ERK-ERF axis drive mammary cell migration in response to EGF. FASEB J. 2012, 26, 1582–1592. [Google Scholar] [CrossRef] [Green Version]
- Arakaki, K.; Uehara, A.; Higa-Nakamine, S.; Kakinohana, M.; Yamamoto, H. Increased expression of EGR1 and KLF4 by polysulfide via activation of the ERK1/2 and ERK5 pathways in cultured intestinal epithelial cells. Biomed. Res. 2020, 41, 119–129. [Google Scholar] [CrossRef]
- Alvarez, J.V.; Febbo, P.G.; Ramaswamy, S.; Loda, M.; Richardson, A.; Frank, D.A. Identification of a genetic signature of activated signal transducer and activator of transcription 3 in human tumors. Cancer Res. 2005, 65, 5054–5062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbolina, M.V.; Adley, B.P.; Ariztia, E.V.; Liu, Y.; Stack, M.S. Microenvironmental regulation of membrane type 1 matrix metalloproteinase activity in ovarian carcinoma cells via collagen-induced EGR1 expression. J. Biol. Chem. 2007, 282, 4924–4931. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.Y.; Kim, J.H.; Baker, A.; Lim, Y.; Lee, Y.H. Transcription factor Egr-1 is essential for maximal matrix metalloproteinase-9 transcription by tumor necrosis factor α. Mol. Cancer Res. 2010, 8, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Faggianelli-Conrozier, N.; Polyzou, A.; Chow, R.; Roth, S.; Trompouki, E.; Vermot, J. Complementary functions of the mechanosensitive factors egr1, klf2b and klf2a instruct the endocardial program. bioRxiv 2019, 718643, Preprint. [Google Scholar]
- Tang, T.; Zhu, Q.; Li, X.; Zhu, G.; Deng, S.; Wang, Y.; Ni, L.; Chen, X.; Zhang, Y.; Xia, T. Protease Nexin I is a feedback regulator of EGF/PKC/MAPK/EGR1 signaling in breast cancer cells metastasis and stemness. Cell Death Dis. 2019, 10, 649. [Google Scholar] [CrossRef] [Green Version]
- Elsevier. Cellular and Molecular Biology of Cancer Stem Cells in Melanoma: Possible Therapeutic Implications; Marzagalli, M., Raimondi, M., Fontana, F., Marelli, M.M., Moretti, R.M., Limonta, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Almairac, F.; Turchi, L.; Sakakini, N.; Debruyne, D.N.; Elkeurti, S.; Gjernes, E.; Polo, B.; Bianchini, L.; Fontaine, D.; Paquis, P. ERK-mediated loss of miR-199a-3p and induction of EGR1 act as a “toggle switch” of GBM cell dedifferentiation into NANOG-and OCT4-positive cells. Cancer Res. 2020, 80, 3236–3250. [Google Scholar] [CrossRef]
- Riddick, G.; Kotliarova, S.; Rodriguez, V.; Kim, H.S.; Linkous, A.; Storaska, A.J.; Ahn, S.; Walling, J.; Belova, G.; Fine, H.A. A core regulatory circuit in glioblastoma stem cells links MAPK activation to a transcriptional program of neural stem cell identity. Sci. Rep. 2017, 7, 43605. [Google Scholar] [CrossRef] [Green Version]
- Sakakini, N.; Turchi, L.; Bergon, A.; Holota, H.; Rekima, S.; Lopez, F.; Paquis, P.; Almairac, F.; Fontaine, D.; Baeza-Kallee, N. A positive feed-forward loop associating EGR1 and PDGFA promotes proliferation and self-renewal in glioblastoma stem cells. J. Biol. Chem. 2016, 291, 10684–10699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudmundsson, K.O.; Nguyen, N.; Oakley, K.; Han, Y.; Gudmundsdottir, B.; Liu, P.; Tessarollo, L.; Jenkins, N.A.; Copeland, N.G.; Du, Y. Prdm16 is a critical regulator of adult long-term hematopoietic stem cell quiescence. Proc. Natl. Acad. Sci. USA 2020, 117, 31945–31953. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Carroll, J.S.; Yee, E.; Thomas, D.D.; Wert-Lamas, L.; Neier, S.C.; Sheynkman, G.; Ritz, J.; Novina, C.D. The lncRNA SLNCR recruits the androgen receptor to EGR1-bound genes in melanoma and inhibits expression of tumor suppressor p21. Cell Rep. 2019, 27, 2493. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Cheng, J.; Wang, H.; Ding, N.; Zhou, F.; Ji, R.; Zhu, L.; Zhu, C.; Pan, Y. A novel regulatory loop miR-101/ANXA2/EGR1 mediates malignant characteristics of liver cancer stem cells. Carcinogenesis 2021, 42, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Katakam, S.K.; Pelucchi, P.; Cocola, C.; Reinbold, R.; Vlodavsky, I.; Greve, B.; Götte, M. Syndecan-1-dependent regulation of heparanase affects invasiveness, stem cell properties, and therapeutic resistance of Caco2 colon cancer cells. Front. Oncol. 2020, 10, 774. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term Description | ES | Term Description | ES | ||
|---|---|---|---|---|---|
| GO Biological Process | negative regulation of heterotypic cell-cell adhesion | 4.7 | Reactome | Rho GTPase cycle | 1.2 |
| cellular response to laminar fluid shear stress | 4.7 | signaling by GPCR | 0.9 | ||
| positive regulation of nitric oxide biosynthetic process | 3.9 | GPCR downstream signaling | 0.9 | ||
| GO Molecular Function | Ras guanyl-nucleotide exchange factor activity | 1.2 | KEGG Pathways | cAMP signaling pathway | 1.4 |
| RNA polymerase II regulatory region sequence-specific DNA binding | 1.0 | axon guidance | 1.3 | ||
| RNA polymerase II regulatory region DNA binding | 1.0 | regulation of actin cytoskeleton | 1.0 | ||
| GO Cellular Component | leading-edge membrane | 1.1 | Keywords | guanine-nucleotide releasing factor | 1.2 |
| site of polarized growth | 1.1 | SH3 domain | 1.1 | ||
| ruffle | 1.0 | actin-binding | 0.9 |
| Gene Symbol | Official Full Name | Log2FC | Gene Symbol | Official Full Name | Log2FC | ||
|---|---|---|---|---|---|---|---|
| upregulated | KLF4 | Kruppel-like factor 4 | 4.5 | downregulated | MMP1 | matrix metallopeptidase 1 | −2.7 |
| MPPED2 | metallophosphoesterase domain containing 2 | 2.9 | MYOF | myoferlin | −2.3 | ||
| SPRR2D | small proline-rich protein 2D | 2.6 | CASC15 | cancer susceptibility 15 | −2.1 | ||
| KLF2 | Kruppel-like factor 2 | 2.6 | PCDH7 | protocadherin 7 | −2.0 | ||
| SERINC2 | serine incorporator 2 | 2.5 | CDH2 | cadherin 2 | −2.0 | ||
| KIAA1161 | myogenesis regulation glycosidase | 2.2 | MATN2 | matrilin 2 | −1.9 | ||
| SEMA3F | semaphorin 3F | 2.2 | ZEB2-AS1 | ZEB2 antisense RNA 1 | −1.9 | ||
| PLB1 | phospholipase B1 | 2.2 | NOV/CCN3 | cellular communication network factor 3 | −1.7 | ||
| GRAMD1B | GRAM domain containing 1B | 2.1 | ITM2A | integral membrane protein 2A | −1.7 | ||
| EGR1 | early growth response 1 | 2.1 | TNC | tenascin C | −1.7 | ||
| CAMK2N1 | calcium/calmodulin-dependent protein kinase II inhibitor 1 | 2.0 | ITK | IL2 inducible T cell kinase | −1.7 | ||
| NANOGP4 | Nanog homeobox pseudogene 4 | 2.0 | PMP2 | peripheral myelin protein 2 | −1.6 | ||
| FLRT2 | fibronectin leucine-rich transmembrane protein 2 | −1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kappelmann-Fenzl, M.; Schmidt, S.K.; Fischer, S.; Schmid, R.; Lämmerhirt, L.; Fischer, L.; Schrüfer, S.; Thievessen, I.; Schubert, D.W.; Matthies, A.; et al. Molecular Changes Induced in Melanoma by Cell Culturing in 3D Alginate Hydrogels. Cancers 2021, 13, 4111. https://doi.org/10.3390/cancers13164111
Kappelmann-Fenzl M, Schmidt SK, Fischer S, Schmid R, Lämmerhirt L, Fischer L, Schrüfer S, Thievessen I, Schubert DW, Matthies A, et al. Molecular Changes Induced in Melanoma by Cell Culturing in 3D Alginate Hydrogels. Cancers. 2021; 13(16):4111. https://doi.org/10.3390/cancers13164111
Chicago/Turabian StyleKappelmann-Fenzl, Melanie, Sonja K. Schmidt, Stefan Fischer, Rafael Schmid, Lisa Lämmerhirt, Lena Fischer, Stefan Schrüfer, Ingo Thievessen, Dirk W. Schubert, Alexander Matthies, and et al. 2021. "Molecular Changes Induced in Melanoma by Cell Culturing in 3D Alginate Hydrogels" Cancers 13, no. 16: 4111. https://doi.org/10.3390/cancers13164111