
Identification of CNGB1 as a Predictor of Response to Neoadjuvant Chemotherapy in Muscle-Invasive Bladder Cancer

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
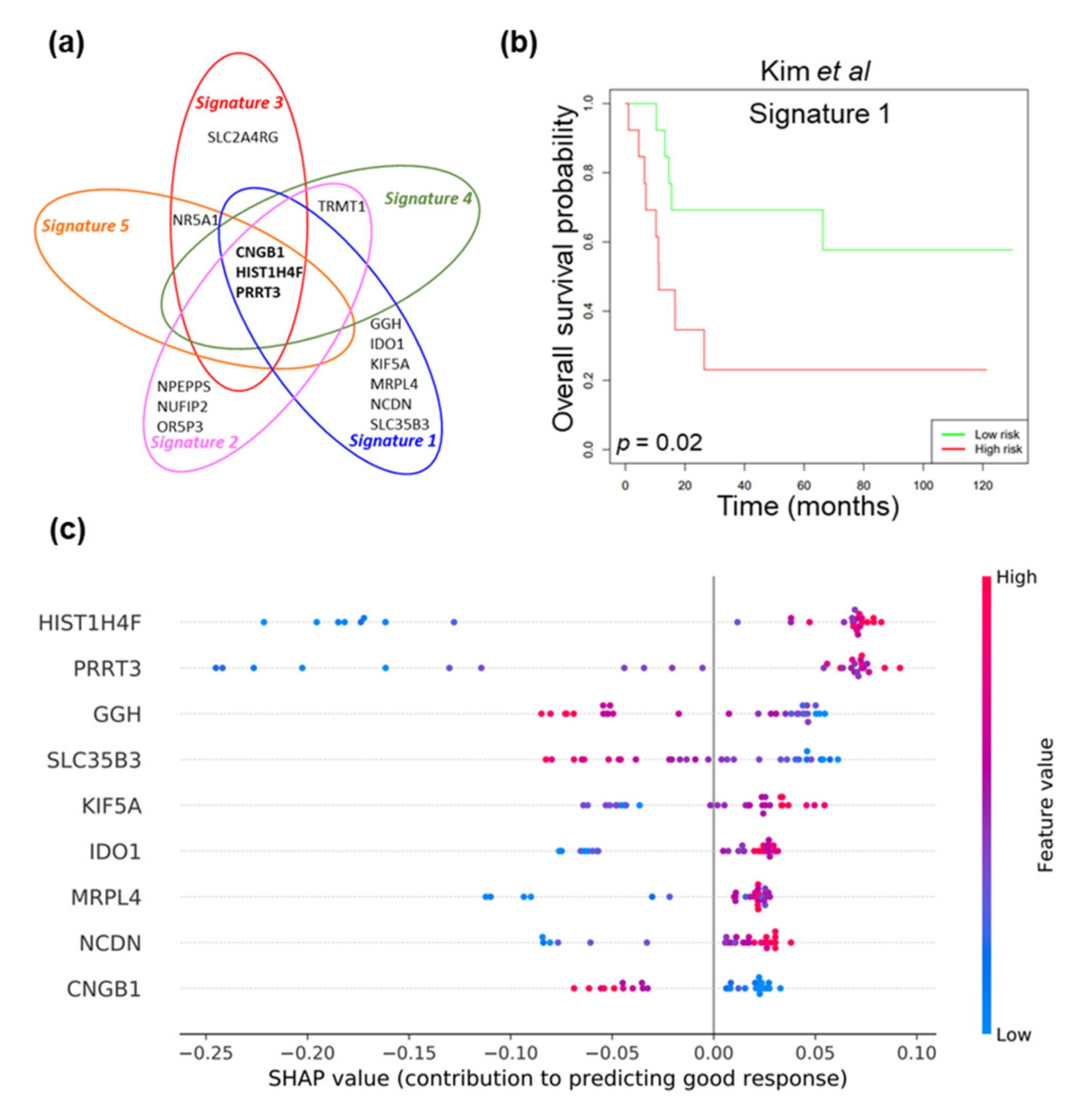
2.1. Machine Learning Approach to Identify Gene Signatures Predicting Response to NAC
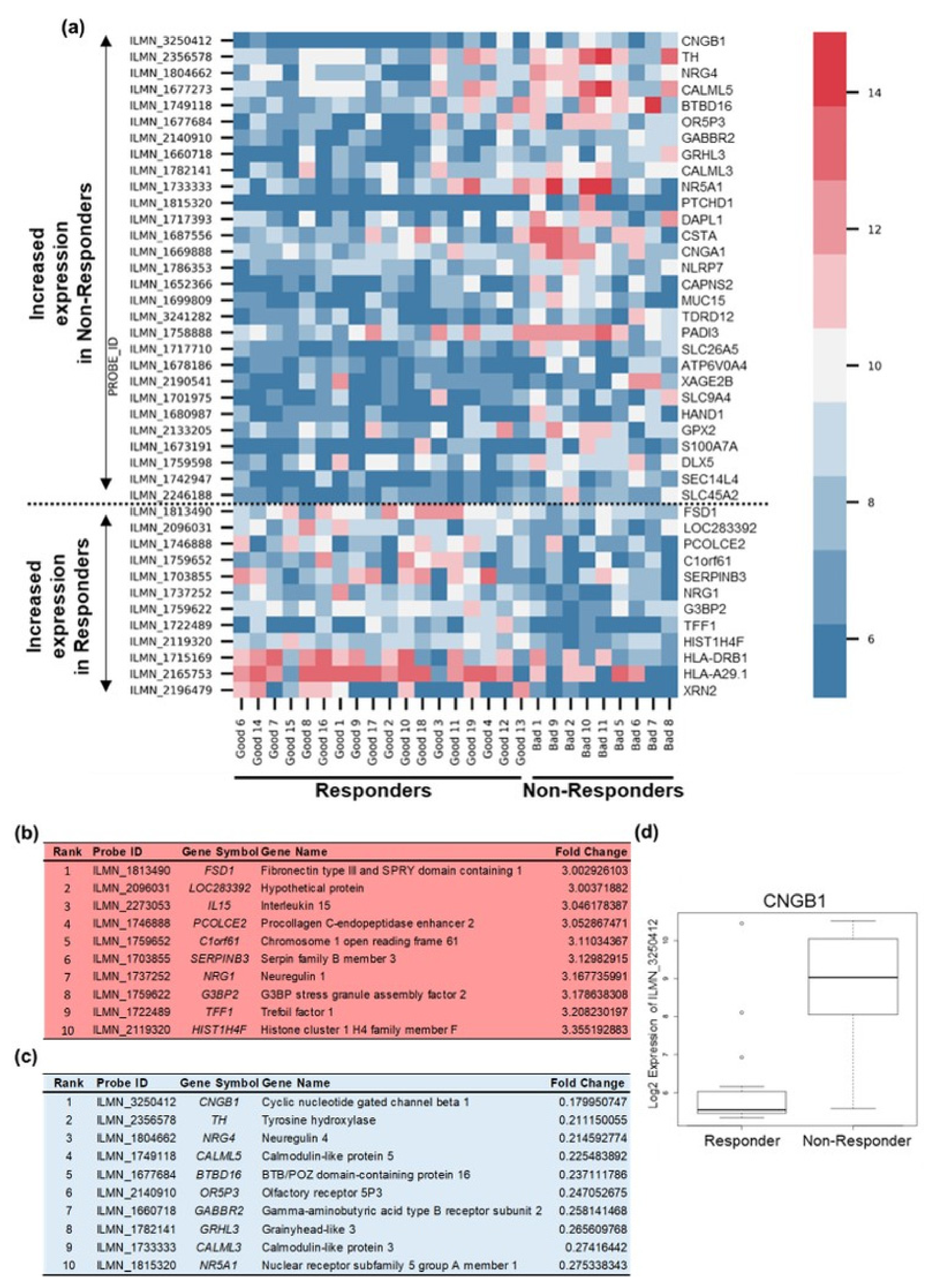
2.2. Differential Gene Expression Analysis to Corroborate Machine Learning Data
2.3. CNGB1 Is Upregulated in ‘Non-Responders’ to NAC and Associates with MIBC Patient Survival
2.4. In Vitro Functional Validation of CNGB1 in MIBC Cells
3. Discussion
4. Materials and Methods
4.1. Patient Cohorts
4.2. Tissue Sampling and Gene Expression Profiling
4.3. Machine Learning Identification of Reduced Biomarker Panels
4.4. Differential Expression Analysis
4.5. Immunostaining of Tissue Microarrays
4.6. Cell Culture
4.7. Western Blotting
4.8. siRNA Transfections
4.9. Cell Growth Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerniak, B.; Dinney, C.; McConkey, D. Origins of Bladder Cancer. Annu. Rev. Pathol. 2016, 11, 149–174. [Google Scholar] [CrossRef]
- Witjes, J.A.; Bruins, H.M.; Cathomas, R.; Compérat, E.M.; Cowan, N.C.; Gakis, G.; Hernández, V.; Espinós, E.L.; Lorch, A.; Neuzillet, Y.; et al. European Association of Urology Guidelines on Muscle-invasive and Metastatic Bladder Cancer: Summary of the 2020 Guidelines. Eur. Urol. 2020, 79, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.P.; Lieskovsky, G.; Cote, R.; Groshen, S.; Feng, A.C.; Boyd, S.; Skinner, E.; Bochner, B.; Thangathurai, D.; Mikhail, M.; et al. Radical cystectomy in the treatment of invasive bladder cancer: Long-term results in 1,054 patients. J. Clin. Oncol. 2001, 19, 666–675. [Google Scholar] [CrossRef]
- Advanced Bladder Cancer (ABC) Meta-analysis Collaboration. Neoadjuvant chemotherapy in invasive bladder cancer: A systematic review and meta-analysis. Lancet 2003, 361, 1927–1934. [Google Scholar] [CrossRef]
- Vale, C.L. Neoadjuvant chemotherapy in invasive bladder cancer: Update of a systematic review and meta-analysis of individual patient data advanced bladder cancer (ABC) meta-analysis collaboration. Eur. Urol. 2005, 48, 202–205, discussion 205–206. [Google Scholar] [CrossRef]
- Grossman, H.B.; Natale, R.B.; Tangen, C.M.; Speights, V.O.; Vogelzang, N.J.; Trump, D.L.; deVere White, R.W.; Sarosdy, M.F.; Wood, D.P., Jr.; Raghavan, D.; et al. Neoadjuvant chemotherapy plus cystectomy compared with cystectomy alone for locally advanced bladder cancer. N. Engl. J. Med. 2003, 349, 859–866. [Google Scholar] [CrossRef] [PubMed]
- International Collaboration of Trialists. International phase III trial assessing neoadjuvant cisplatin, methotrexate, and vinblastine chemotherapy for muscle-invasive bladder cancer: Long-term results of the BA06 30894 trial. J. Clin. Oncol. 2011, 29, 2171–2177. [Google Scholar] [CrossRef] [Green Version]
- Choueiri, T.K.; Jacobus, S.; Bellmunt, J.; Qu, A.; Appleman, L.J.; Tretter, C.; Bubley, G.J.; Stack, E.C.; Signoretti, S.; Walsh, M.; et al. Neoadjuvant dose-dense methotrexate, vinblastine, doxorubicin, and cisplatin with pegfilgrastim support in muscle-invasive urothelial cancer: Pathologic, radiologic, and biomarker correlates. J. Clin. Oncol. 2014, 32, 1889–1894. [Google Scholar] [CrossRef]
- Font, A.; Taron, M.; Gago, J.L.; Costa, C.; Sánchez, J.J.; Carrato, C.; Mora, M.; Celiz, P.; Perez, L.; Rodríguez, D.; et al. BRCA1 mRNA expression and outcome to neoadjuvant cisplatin-based chemotherapy in bladder cancer. Ann. Oncol. 2011, 22, 139–144. [Google Scholar] [CrossRef]
- Liu, D.; Plimack, E.R.; Hoffman-Censits, J.; Garraway, L.A.; Bellmunt, J.; Van Allen, E.; Rosenberg, J.E. Clinical Validation of Chemotherapy Response Biomarker ERCC2 in Muscle-Invasive Urothelial Bladder Carcinoma. JAMA Oncol. 2016, 2, 1094–1096. [Google Scholar] [CrossRef] [Green Version]
- Van Allen, E.M.; Mouw, K.W.; Kim, P.; Iyer, G.; Wagle, N.; Al-Ahmadie, H.; Zhu, C.; Ostrovnaya, I.; Kryukov, G.V.; O’Connor, K.W.; et al. Somatic ERCC2 mutations correlate with cisplatin sensitivity in muscle-invasive urothelial carcinoma. Cancer Discov. 2014, 4, 1140–1153. [Google Scholar] [CrossRef] [Green Version]
- Als, A.B.; Dyrskjøt, L.; von der Maase, H.; Koed, K.; Mansilla, F.; Toldbod, H.E.; Jensen, J.L.; Ulhøi, B.P.; Sengeløv, L.; Jensen, K.M.E.; et al. Emmprin and survivin predict response and survival following cisplatin-containing chemotherapy in patients with advanced bladder cancer. Clin. Cancer Res. 2007, 13 (15 Pt 1), 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Groenendijk, F.H.; de Jong, J.; Fransen van de Putte, E.E.; Michaut, M.; Schlicker, A.; Peters, D.; Velds, A.; Nieuwland, M.; van den Heuvel, M.M.; Kerkhoven, R.M.; et al. ERBB2 Mutations Characterize a Subgroup of Muscle-invasive Bladder Cancers with Excellent Response to Neoadjuvant Chemotherapy. Eur. Urol. 2016, 69, 384–388. [Google Scholar] [CrossRef]
- Kato, Y.; Zembutsu, H.; Takata, R.; Miya, F.; Tsunoda, T.; Obara, W.; Fujioka, T.; Nakamura, Y. Predicting response of bladder cancers to gemcitabine and carboplatin neoadjuvant chemotherapy through genome-wide gene expression profiling. Exp. Ther. Med. 2011, 2, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Takata, R.; Katagiri, T.; Kanehira, M.; Tsunoda, T.; Shuin, T.; Miki, T.; Namiki, M.; Kohri, K.; Matsushita, Y.; Fujioka, T.; et al. Predicting response to methotrexate, vinblastine, doxorubicin, and cisplatin neoadjuvant chemotherapy for bladder cancers through genome-wide gene expression profiling. Clin. Cancer Res. 2005, 11, 2625–2636. [Google Scholar] [CrossRef] [Green Version]
- Takata, R.; Katagiri, T.; Kanehira, M.; Shuin, T.; Miki, T.; Namiki, M.; Kohri, K.; Tsunoda, T.; Fujioka, T.; Nakamura, Y. Validation study of the prediction system for clinical response of M-VAC neoadjuvant chemotherapy. Cancer Sci. 2007, 98, 113–117. [Google Scholar] [CrossRef]
- Baras, A.S.; Gandhi, N.; Munari, E.; Faraj, S.; Shultz, L.; Marchionni, L.; Schoenberg, M.; Hahn, N.; Hoque, M.; Berman, D.; et al. Identification and Validation of Protein Biomarkers of Response to Neoadjuvant Platinum Chemotherapy in Muscle Invasive Urothelial Carcinoma. PLoS ONE 2015, 10, e0131245. [Google Scholar]
- Williams, P.D.; Cheon, S.; Havaleshko, D.M.; Jeong, H.; Cheng, F.; Theodorescu, D.; Lee, J.K. Concordant gene expression signatures predict clinical outcomes of cancer patients undergoing systemic therapy. Cancer Res. 2009, 69, 8302–8309. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.; Porten, S.; Kim, S.; Willis, D.; Plimack, E.R.; Hoffman-Censits, J.; Roth, B.; Cheng, T.; Tran, M.; Lee, I.-L.; et al. Identification of distinct basal and luminal subtypes of muscle-invasive bladder cancer with different sensitivities to frontline chemotherapy. Cancer Cell 2014, 25, 152–165. [Google Scholar] [CrossRef] [Green Version]
- McConkey, D.J.; Choi, W.; Shen, Y.; Lee, I.-L.; Porten, S.; Matin, S.F.; Kamat, A.M.; Corn, P.; Millikan, R.E.; Dinney, C.; et al. A Prognostic Gene Expression Signature in the Molecular Classification of Chemotherapy-naive Urothelial Cancer is Predictive of Clinical Outcomes from Neoadjuvant Chemotherapy: A Phase 2 Trial of Dose-dense Methotrexate, Vinblastine, Doxorubicin, and Cisplatin with Bevacizumab in Urothelial Cancer. Eur. Urol. 2016, 69, 855–862. [Google Scholar]
- Taber, A.; Christensen, E.; Lamy, P.; Nordentoft, I.; Prip, F.; Lindskrog, S.V.; Birkenkamp-Demtröder, K.; Okholm, T.L.H.; Knudsen, M.; Pedersen, J.S.; et al. Molecular correlates of cisplatin-based chemotherapy response in muscle invasive bladder cancer by integrated multi-omics analysis. Nat. Commun. 2020, 11, 4858. [Google Scholar] [CrossRef]
- Font, A.; Domènech, M.; Benítez, R.; Rava, M.; Marqués, M.; Ramírez, J.L.; Pineda, S.; Domínguez-Rodríguez, S.; Gago, J.L.; Badal, J.; et al. Immunohistochemistry-Based Taxonomical Classification of Bladder Cancer Predicts Response to Neoadjuvant Chemotherapy. Cancers 2020, 12, 1784. [Google Scholar] [CrossRef] [PubMed]
- Kilari, D.; Iczkowski, K.A.; Pandya, C.; Robin, A.J.; Messing, E.M.; Guancial, E.; Kim, E.S. Copper Transporter-CTR1 Expression and Pathological Outcomes in Platinum-treated Muscle-invasive Bladder Cancer Patients. Anticancer Res. 2016, 36, 495–501. [Google Scholar]
- Hoffmann, A.C.; Wild, P.; Leicht, C.; Bertz, S.; Danenberg, K.D.; Danenberg, P.V.; Stöhr, R.; Stöckle, M.; Lehmann, J.; Schuler, M.; et al. MDR1 and ERCC1 expression predict outcome of patients with locally advanced bladder cancer receiving adjuvant chemotherapy. Neoplasia 2010, 12, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Qian, F.; Shen, L.; Chen, F.; Chen, J.; Shen, B. Computer-aided biomarker discovery for precision medicine: Data resources, models and applications. Brief. Bioinform. 2019, 20, 952–975. [Google Scholar] [CrossRef]
- Lazzarini, N.; Bacardit, J. RGIFE: A ranked guided iterative feature elimination heuristic for the identification of biomarkers. BMC Bioinform. 2017, 18, 322. [Google Scholar] [CrossRef] [Green Version]
- Lazzarini, N.; Runhaar, J.; Bay-Jensen, A.C.; Thudium, C.S.; Bierma-Zeinstra, S.M.A.; Henrotin, Y.; Bacardit, J. A machine learning approach for the identification of new biomarkers for knee osteoarthritis development in overweight and obese women. Osteoarthr. Cartil. 2017, 25, 2014–2021. [Google Scholar] [CrossRef] [Green Version]
- Swan, A.L.; Stekel, D.J.; Hodgman, C.; Allaway, D.; Alqahtani, M.H.; Mobasheri, A.; Bacardit, J. A machine learning heuristic to identify biologically relevant and minimal biomarker panels from omics data. BMC Genom. 2015, 16 (Suppl. 1), S2. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.J.; Kim, E.-J.; Kim, S.-K.; Kim, Y.-J.; Ha, Y.-S.; Jeong, P.; Kim, M.-J.; Yun, S.-J.; Lee, K.M.; Moon, S.-K.; et al. Predictive value of progression-related gene classifier in primary non-muscle invasive bladder cancer. Mol. Cancer 2010, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Kaupp, U.B.; Seifert, R. Cyclic nucleotide-gated ion channels. Physiol. Rev. 2002, 82, 769–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Gold, G.H. A cyclic nucleotide-gated conductance in olfactory receptor cilia. Nature 1987, 325, 442–444. [Google Scholar] [CrossRef]
- Chen, T.Y.; Peng, Y.-W.; Dhallan, R.S.; Ahamed, B.; Reed, R.R.; Yau, K.-W. A new subunit of the cyclic nucleotide-gated cation channel in retinal rods. Nature 1993, 362, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.J.; Gordon, S.E. Cooperativity and cooperation in cyclic nucleotide-gated ion channels. Biochemistry 2000, 39, 14003–14011. [Google Scholar] [CrossRef]
- Guo, C.C.; Majewski, T.; Zhang, L.; Yao, H.; Bondaruk, J.; Wang, Y.; Zhang, S.; Wang, Z.; Lee, J.G.; Lee, S.; et al. Dysregulation of EMT Drives the Progression to Clinically Aggressive Sarcomatoid Bladder Cancer. Cell Rep. 2019, 27, 1781–1793.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trudeau, M.C.; Zagotta, W.N. Dynamics of Ca2+-calmodulin-dependent inhibition of rod cyclic nucleotide-gated channels measured by patch-clamp fluorometry. J. Gen. Physiol. 2004, 124, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhao, Z.; Sun, Z.; Liu, C.; Cheng, X.; Ruge, F.; Yang, Y.; Jiang, W.G.; Ye, L. Increased expression of Psoriasin is correlated with poor prognosis of bladder transitional cell carcinoma by promoting invasion and proliferation. Oncol. Rep. 2020, 43, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.T.; Kim, J.; Yan, C.; Jeong, P.; Choi, S.Y.; Lee, O.J.; Chae, Y.B.; Yun, S.J.; Lee, S.C.; Kim, W.J. S100A9 and EGFR gene signatures predict disease progression in muscle invasive bladder cancer patients after chemotherapy. Ann. Oncol. 2014, 25, 974–979. [Google Scholar] [CrossRef]
- Shiota, M.; Tsunoda, T.; Song, Y.; Yokomizo, A.; Tada, Y.; Oda, Y.; Naito, S. Enhanced S100 calcium-binding protein P expression sensitizes human bladder cancer cells to cisplatin. BJU Int. 2011, 107, 1148–1153. [Google Scholar] [CrossRef]
- Sandoval, J.; Mendez-Gonzalez, J.; Nadal, E.; Chen, G.; Carmona, F.J.; Sayols, S.; Moran, S.; Heyn, H.; Vizoso, M.; Gomez, A.; et al. A prognostic DNA methylation signature for stage I non-small-cell lung cancer. J. Clin. Oncol. 2013, 31, 4140–4147. [Google Scholar] [CrossRef]
- Izquierdo-Torres, E.; Hernández-Oliveras, A.; Fuentes-García, G.; Zarain-Herzberg, A. Calcium signaling and epigenetics: A key point to understand carcinogenesis. Cell Calcium 2020, 91, 102285. [Google Scholar] [CrossRef] [PubMed]
- Duenas-Gonzalez, A.; García-López, P.; Herrera, L.A.; Medina-Franco, J.L.; González-Fierro, A.; Candelaria, M. The prince and the pauper. A tale of anticancer targeted agents. Mol. Cancer 2008, 7, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, R.L.; Strassmaier, T.; Brady, J.D.; Karpen, J.W. The pharmacology of cyclic nucleotide-gated channels: Emerging from the darkness. Curr. Pharm. Des. 2006, 12, 3597–3613. [Google Scholar] [CrossRef]
- El-Mahdy, H.A.; El-Husseiny, A.A.; Kandil, Y.I.; Gamal El-Din, A.M. Diltiazem potentiates the cytotoxicity of gemcitabine and 5-fluorouracil in PANC-1 human pancreatic cancer cells through inhibition of P-glycoprotein. Life Sci. 2020, 262, 118518. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Martinez, T.R. Distribution-balanced stratified cross-validation for accuracy estimation. J. Exp. Theor. Artif. Intell. 2000, 12, 1–12. [Google Scholar] [CrossRef]
- Moreno-Torres, J.G.; Saez, J.A.; Herrera, F. Study on the impact of partition-induced dataset shift on k-fold cross-validation. IEEE Trans. Neural. Netw. Learn. Syst. 2012, 23, 1304–1312. [Google Scholar] [CrossRef]
- Lundberg, S.M.; Erion, G.; Chen, H.; DeGrave, A.; Prutkin, J.M.; Nair, B.; Katz, R.; Himmelfarb, J.; Bansal, N.; Lee, S.-I. From Local Explanations to Global Understanding with Explainable AI for Trees. Nat. Mach Intell. 2020, 2, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, A.C.; Veeratterapillay, R.; Williamson, S.C.; El-Sherif, A.; Sahay, N.; Thomas, H.D.; Mantilla, A.; Pickard, R.S.; Robson, C.N.; Heer, R. Side population in human non-muscle invasive bladder cancer enriches for cancer stem cells that are maintained by MAPK signalling. PLoS ONE 2012, 7, e50690. [Google Scholar] [CrossRef]
- Dinney, C.P.; Fishbeck, R.; Singh, R.K.; Eve, B.; Pathak, S.; Brown, N.; Xie, B.; Fan, D.; Bucana, C.D.; Fidler, I.J.; et al. Isolation and characterization of metastatic variants from human transitional cell carcinoma passaged by orthotopic implantation in athymic nude mice. J. Urol. 1995, 154, 1532–1538. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Gender | Age | Preoperative Stage | Preoperative Grade | TNM Stage After RC | Response |
|---|---|---|---|---|---|---|
| good1 | Male | 71 | T2 | G2/3 | T0 N0 | Responder |
| good2 | Female | 68 | T2 | G2/3 | T0 N0 | Responder |
| good3 | Male | 74 | T2 | G3 | Tcis N0 | Responder |
| good4 | Male | 67 | T2 | G2/3 | T0 N0 | Responder |
| good5 | Female | 69 | T2 | G3 | T0 N0 | Responder |
| good6 | Male | 56 | T2 | G2 | Tis N0 | Responder |
| good7 | Male | 46 | T2 | G3 | T0 N0 | Responder |
| good8 | Male | 64 | T2 | G2/3 | T0 N1 * | Responder |
| good9 | Male | 63 | T2 | G3 | T0 N0 | Responder |
| good10 | Male | 70 | T4 | G3 | T0 N0 | Responder |
| good11 | Male | 69 | T2 | G3 | T0 N0 | Responder |
| good12 | Male | 63 | T2 | G2 | T1 N0 | Responder |
| good13 | Male | 58 | T1/2 | G3 | Tcis N0 | Responder |
| good14 | Male | 60 | T2 | G3 | T0 N0 | Responder |
| good15 | Male | 59 | T2 | G3 | T0 N0 | Responder |
| good16 | Female | 70 | T2 | G3 | Tis N0 | Responder |
| good17 | Male | 69 | T2 | G3 | T0 N0 | Responder |
| good18 | Female | 73 | T2 | G3 | T0 N0 | Responder |
| good19 | Male | 59 | T2 | G3 | T0 N0 | Responder |
| bad1 | Male | 58 | T2 | G3 | T4a N1 | Non-Responder |
| bad2 | Male | 71 | T2 | G3 | T3a N0 | Non-Responder |
| bad3 | Male | 74 | T2 | G3 | T3 N0 | Non-Responder |
| bad4 | Male | 61 | T2 | G3 | T3b N2 | Non-Responder |
| bad5 | Female | 63 | T2 | G2/3 | T3a N2 | Non-Responder |
| bad6 | Male | 65 | T2 | G3 | T2b N0 | Non-Responder |
| bad7 | Male | 60 | T2 | G3 | T3b N0 | Non-Responder |
| bad8 | Male | 64 | T1/2 | G3 | T2b N0 | Non-Responder |
| bad9 | Male | 74 | T2 | G2 | T3a N0 | Non-Responder |
| bad10 | Male | 58 | T2 | G3 | T3a N0 | Non-Responder |
| bad11 | Male | 50 | T2 | G2/3 | T4a N2 | Non-Responder |
| Signature | Gene Symbol | Gene Name | Prediction Accuracy |
|---|---|---|---|
| 1 | CNGB1 | cyclic nucleotide gated channel beta 1 | 1.000 (30/30*) |
| GGH | gamma-glutamyl hydrolase | ||
| HIST1H4F | histone cluster 1 H4, family member F | ||
| IDO1 | indoleamine 2,3-dioxygenase 1 | ||
| KIF5A | kinesin family member 5A | ||
| MRPL4 | mitochondrial ribosomal protein L4 | ||
| NCDN | neurochondrin | ||
| PRRT3 | proline-rich transmembrane protein 3 | ||
| SLC35B3 | solute carrier family 35, member B3 | ||
| 2 | CNGB1 | cyclic nucleotide gated channel beta 1 | 1.000 (30/30*) |
| HIST1H4F | histone cluster 1 H4, family member F | ||
| NPEPPS | aminopeptidase puromycin sensitive | ||
| NUFIP2 | nuclear fragile X mental retardation protein interacting protein 2 | ||
| OR5P3 | olfactory receptor family 5, subfamily P, member 3 | ||
| PRRT3 | proline-rich transmemembrane protein 3 | ||
| TRMT1 | tRNA methyltransferase 1 | ||
| 3 | CNGB1 | cyclic nucleotide gated channel beta 1 | 0.967 (29/30*) |
| HIST1H4F | histone cluster 1 H4, family member F | ||
| NR5A1 | nuclear receptor subfamily 5, group A, member 1 | ||
| PRRT3 | proline-rich transmembrane protein 3 | ||
| SLC2A4RG | solute carrier family 2 member 4 regulator | ||
| 4 | CNGB1 | cyclic nucleotide gated channel beta 1 | 0.967 (29/30*) |
| HIST1H4F | histone cluster 1 H4, family member F | ||
| PRRT3 | proline-rich transmembrane protein 3 | ||
| TRMT1 | tRNA methyltransferase 1 | ||
| 5 | CNGB1 | cyclic nucleotide gated channel beta 1 | 0.933 (28/30*) |
| HIST1H4F | histone cluster 1 H4, family member F | ||
| NR5A1 | nuclear receptor subfamily 5, group A, member 1 | ||
| PRRT3 | proline-rich transmembrane protein 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hepburn, A.C.; Lazzarini, N.; Veeratterapillay, R.; Wilson, L.; Bacardit, J.; Heer, R. Identification of CNGB1 as a Predictor of Response to Neoadjuvant Chemotherapy in Muscle-Invasive Bladder Cancer. Cancers 2021, 13, 3903. https://doi.org/10.3390/cancers13153903
Hepburn AC, Lazzarini N, Veeratterapillay R, Wilson L, Bacardit J, Heer R. Identification of CNGB1 as a Predictor of Response to Neoadjuvant Chemotherapy in Muscle-Invasive Bladder Cancer. Cancers. 2021; 13(15):3903. https://doi.org/10.3390/cancers13153903
Chicago/Turabian StyleHepburn, Anastasia C., Nicola Lazzarini, Rajan Veeratterapillay, Laura Wilson, Jaume Bacardit, and Rakesh Heer. 2021. "Identification of CNGB1 as a Predictor of Response to Neoadjuvant Chemotherapy in Muscle-Invasive Bladder Cancer" Cancers 13, no. 15: 3903. https://doi.org/10.3390/cancers13153903