
miR-92b-3p Regulates Cell Cycle and Apoptosis by Targeting CDKN1C, Thereby Affecting the Sensitivity of Colorectal Cancer Cells to Chemotherapeutic Drugs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Reagents
2.3. Animals
2.4. Small RNA Sequencing
2.5. Target Prediction
2.6. Cell Transfection
2.7. RNA Extraction and qRT-PCR
2.8. Cell Proliferation Assay
2.9. Drug Sensitivity Assay
2.10. Cell Cycle Assay
2.11. Apoptosis Assay
2.12. Dual Luciferase Reporter Assay
2.13. Western Blotting Analysis
2.14. Nude Mice Xenograft Tumor Assay
2.15. Statistical Analysis
3. Results
3.1. miR-92b-3p Is Highly Expressed in HCT8/T Cells and Induced by PTX
3.2. miR-92b-3p Maintains the Cell Viability in a Chemotherapy Setting
3.3. Overexpression of miR-92b-3p Desensitizes HCT8 Cells to Chemotherapeutic Drugs and Knockdown of miR-92b-3p Resensitizes HCT8/T Cells to Chemotherapeutic Drugs
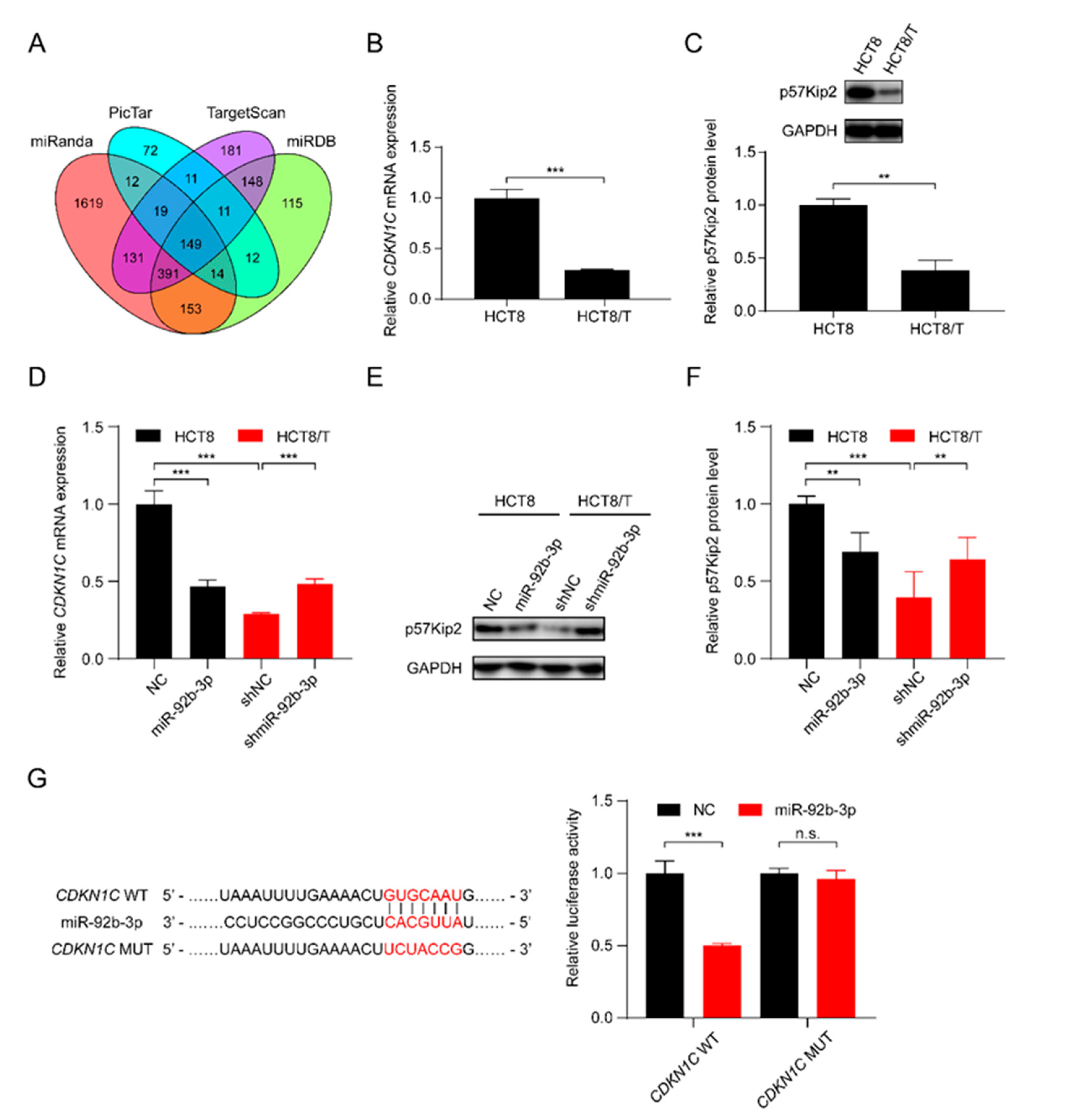
3.4. miR-92b-3p Directly Targets CDKN1C and Suppresses Its Expression
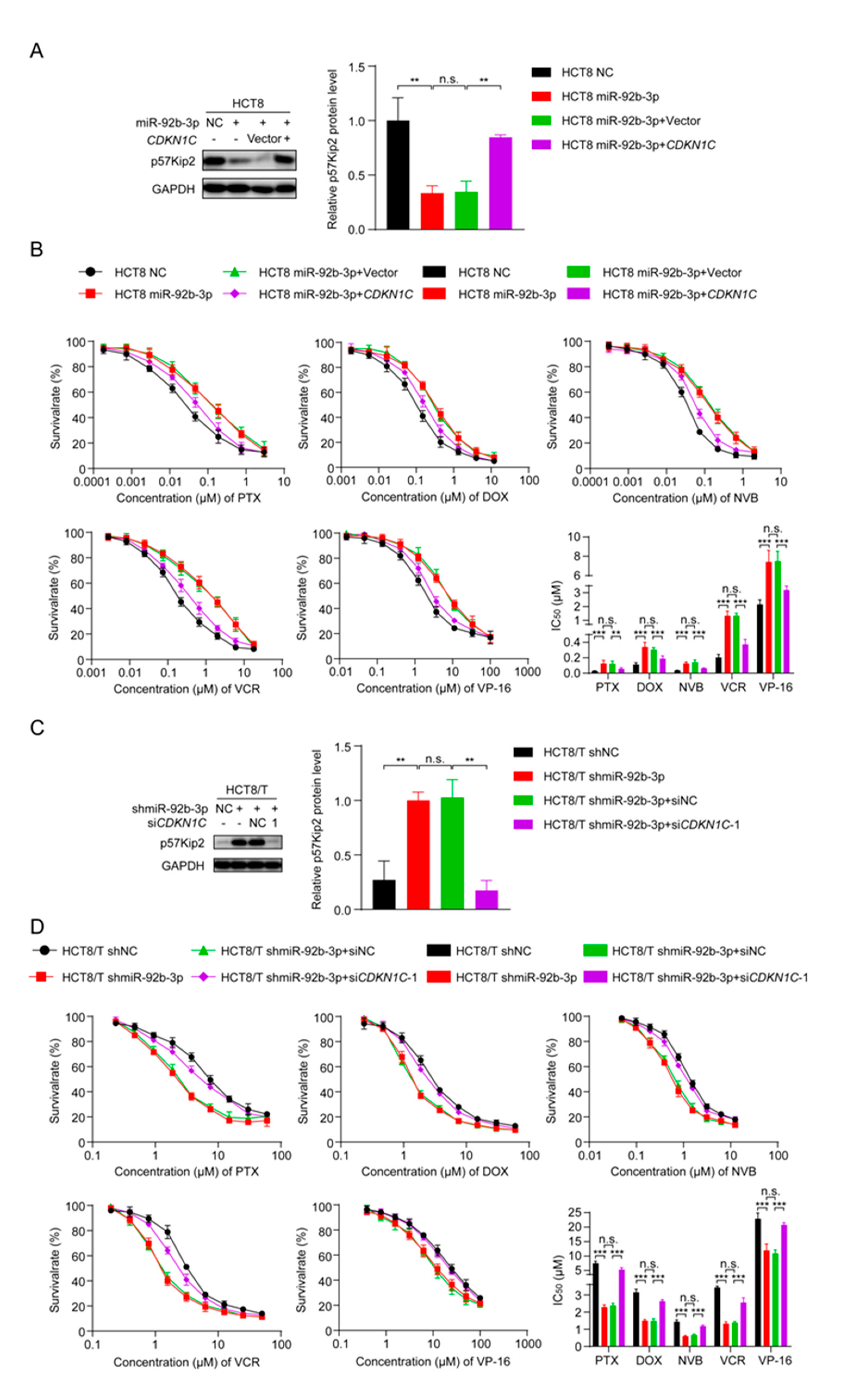
3.5. CDKN1C Silencing Reduces the Sensitivity of HCT8 Cells to Chemotherapeutic Drugs and Overexpression of CDKN1C Restores the Sensitivity of HCT8/T Cells to Chemotherapeutic Drugs
3.6. miR-92b-3p Regulates the Sensitivity of HCT8 and HCT8/T Cells to Chemotherapeutic Drugs by Targeting CDKN1C
3.7. The miR-92b-3p/CDKN1C Axis Mediates the Sensitivity of HCT8 and HCT8/T Cells to Chemotherapeutic Drugs through Regulating Cell Cycle and Apoptosis
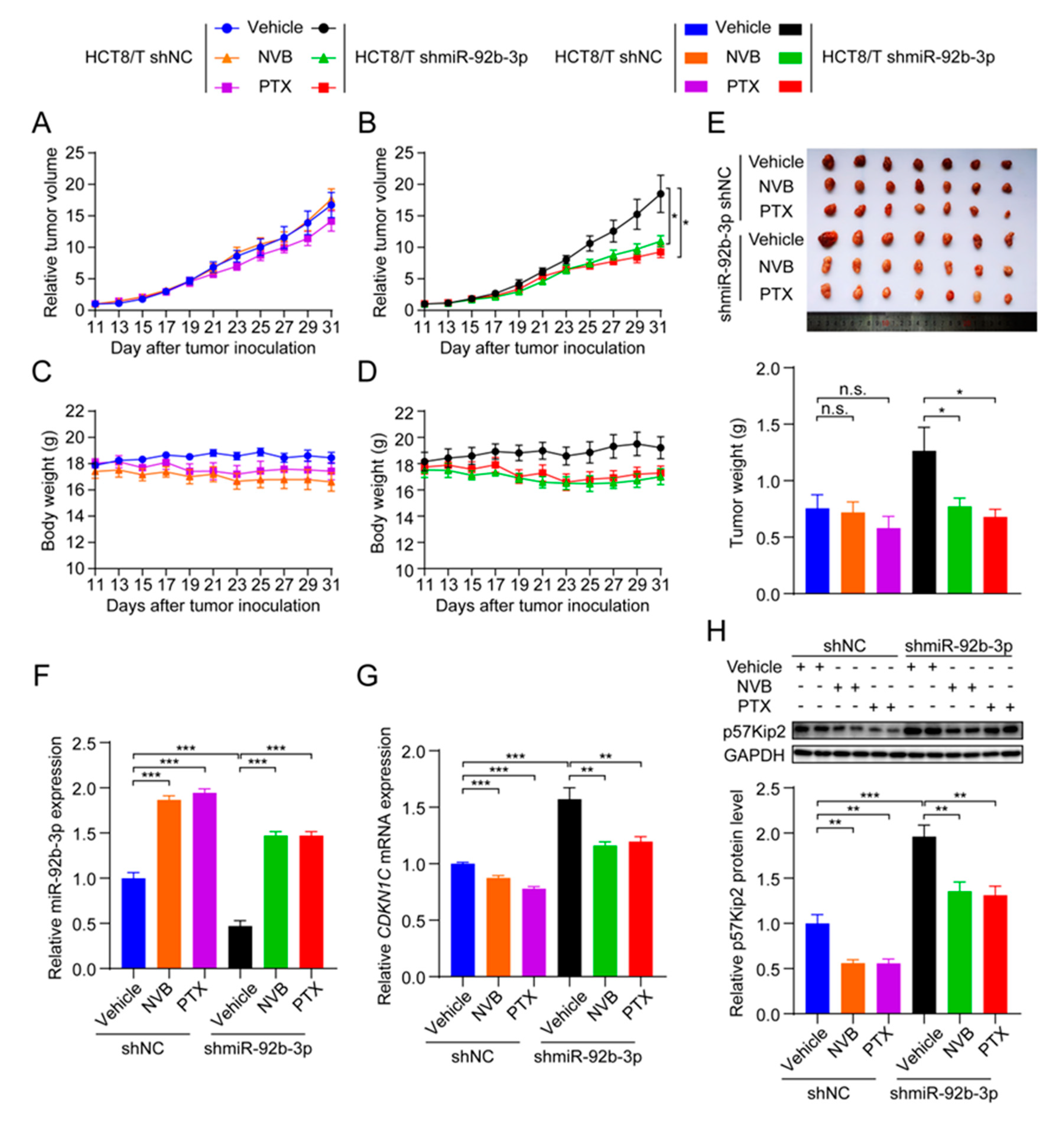
3.8. miR-92b-3p Suppresses the Sensitivity of HCT8/T Xenograft Tumors to Chemotherapeutic Drugs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Neerincx, M.; Poel, D.; Sie, D.L.S.; van Grieken, N.C.T.; Shankaraiah, R.C.; van der Wolf-de Lijster, F.S.W.; van Waesberghe, J.T.M.; Burggraaf, J.D.; Eijk, P.P.; Verhoef, C.; et al. Combination of a six microRNA expression profile with four clinicopathological factors for response prediction of systemic treatment in patients with advanced colorectal cancer. PLoS ONE 2018, 13, e0201809. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Pichler, M.; Stiegelbauer, V.; Vychytilova-Faltejskova, P.; Ivan, C.; Ling, H.; Winter, E.; Zhang, X.; Goblirsch, M.; Wulf-Goldenberg, A.; Ohtsuka, M.; et al. Genome-Wide miRNA Analysis Identifies miR-188-3p as a novel prognostic marker and molecular factor involved in colorectal carcinogenesis. Clin. Cancer Res. 2017, 23, 1323–1333. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Xu, Y.; Zhang, Q.; Yang, Y.; Mou, Y.; Gao, Y.; Chen, R.; Chen, C.; Dai, P. MiR-483-3p regulates oxaliplatin resistance by targeting FAM171B in human colorectal cancer cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.H.; Lyskjaer, I.; Jersie-Christensen, R.R.; Tarpgaard, L.S.; Primdal-Bengtson, B.; Nielsen, M.M.; Pedersen, J.S.; Hansen, T.P.; Hansen, F.; Olsen, J.V.; et al. miR-625-3p regulates oxaliplatin resistance by targeting MAP2K6-p38 signalling in human colorectal adenocarcinoma cells. Nat. Commun. 2016, 7, 12436. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.; Dowden, S.; Colwell, B.; Collins, L.L.; Berry, S. Navigating later lines of treatment for advanced colorectal cancer—optimizing targeted biological therapies to improve outcomes. Cancer Treat. Rev. 2014, 40, 1171–1181. [Google Scholar] [CrossRef]
- Rasmussen, M.H.; Jensen, N.F.; Tarpgaard, L.S.; Qvortrup, C.; Romer, M.U.; Stenvang, J.; Hansen, T.P.; Christensen, L.L.; Lindebjerg, J.; Hansen, F.; et al. High expression of microRNA-625-3p is associated with poor response to first-line oxaliplatin based treatment of metastatic colorectal cancer. Mol. Oncol. 2013, 7, 637–646. [Google Scholar] [CrossRef]
- Meisenberg, C.; Gilbert, D.C.; Chalmers, A.; Haley, V.; Gollins, S.; Ward, S.E.; El-Khamisy, S.F. Clinical and cellular roles for TDP1 and TOP1 in modulating colorectal cancer response to irinotecan. Mol. Cancer Ther. 2015, 14, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, H.; Assaraf, Y.G.; Zhao, K.; Xu, X.; Xie, J.; Yang, D.H.; Chen, Z.S. Overcoming ABC transporter-mediated multidrug resistance: Molecular mechanisms and novel therapeutic drug strategies. Drug Resist. Updates 2016, 27, 14–29. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Voortman, J.; Dingemans, A.M.; Voeller, D.M.; Pham, T.; Wang, Y.; Giaccone, G. MicroRNA expression and clinical outcome of small cell lung cancer. PLoS ONE 2011, 6, e21300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Jaarsveld, M.T.; Helleman, J.; Boersma, A.W.; van Kuijk, P.F.; van Ijcken, W.F.; Despierre, E.; Vergote, I.; Mathijssen, R.H.; Berns, E.M.; Verweij, J.; et al. miR-141 regulates KEAP1 and modulates cisplatin sensitivity in ovarian cancer cells. Oncogene 2013, 32, 4284–4293. [Google Scholar] [CrossRef]
- Nishida, K.; Kuwano, Y.; Rokutan, K. The MicroRNA-23b/27b/24 cluster facilitates colon cancer cell migration by targeting FOXP2. Cancers 2020, 12, 174. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Glaser, S.S.; Francis, H.; DeMorrow, S.; Han, Y.; Passarini, J.D.; Stokes, A.; Cleary, J.P.; Liu, X.; Venter, J.; et al. Functional analysis of microRNAs in human hepatocellular cancer stem cells. J. Cell Mol. Med. 2012, 16, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Leidinger, P.; Blin, N.; Meese, E. miRNA: Small molecules as potential novel biomarkers in cancer. Curr. Med. Chem. 2010, 17, 4427–4432. [Google Scholar] [CrossRef] [PubMed]
- Kiss, I.; Mlcochova, J.; Souckova, K.; Fabian, P.; Poprach, A.; Halamkova, J.; Svoboda, M.; Vyzula, R.; Slaby, O. MicroRNAs as outcome predictors in patients with metastatic colorectal cancer treated with bevacizumab in combination with FOLFOX. Oncol. Lett. 2017, 14, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, A.; Romero-Cordoba, S.; Plantamura, I.; Cosentino, G.; Hidalgo-Miranda, A.; Tagliabue, E.; Iorio, M.V. MiR-302b as a combinatorial therapeutic approach to improve cisplatin chemotherapy efficacy in human triple-negative breast cancer. Cancers 2020, 12, 2261. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Zhang, Z.; Shang, Y.; Jiang, X.; Dong, J.; Yu, P.; Nie, Y.; Zhao, Q. miR-23b-3p regulates the chemoresistance of gastric cancer cells by targeting ATG12 and HMGB2. Cell Death Dis. 2015, 6, e1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristobal, I.; Rubio, J.; Santos, A.; Torrejon, B.; Carames, C.; Imedio, L.; Mariblanca, S.; Luque, M.; Sanz-Alvarez, M.; Zazo, S.; et al. MicroRNA-199b downregulation confers resistance to 5-Fluorouracil treatment and predicts poor outcome and response to neoadjuvant chemoradiotherapy in locally advanced rectal cancer patients. Cancers 2020, 12, 1655. [Google Scholar] [CrossRef]
- Dan, T.; Shastri, A.A.; Palagani, A.; Buraschi, S.; Neill, T.; Savage, J.E.; Kapoor, A.; DeAngelis, T.; Addya, S.; Camphausen, K.; et al. miR-21 plays a dual role in tumor formation and cytotoxic response in breast tumors. Cancers 2021, 13, 888. [Google Scholar] [CrossRef]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids--the mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.J. MicroRNAs as promising biomarkers in cancer diagnostics. Biomark. Res. 2014, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Neerincx, M.; Buffart, T.E.; Mulder, C.J.; Meijer, G.A.; Verheul, H.M. The future of colorectal cancer: Implications of screening. Gut 2013, 62, 1387–1389. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.B.; Cai, L.; Lin, S.J.; Lu, J.L.; Yao, Y.; Zhou, L.F. The miR-92b functions as a potential oncogene by targeting on Smad3 in glioblastomas. Brain Res. 2013, 1529, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Wang, H.; Jiang, M.; Yan, Y.; Li, W.; Xu, H.; Huang, Q.; Lu, Y.; Chen, J. The E3 ubiquitin ligase CHIP/miR-92b/PTEN regulatory network contributes to tumorigenesis of glioblastoma. Am. J. Cancer Res. 2017, 7, 289–300. [Google Scholar]
- Wang, K.; Wang, X.; Zou, J.; Zhang, A.; Wan, Y.; Pu, P.; Song, Z.; Qian, C.; Chen, Y.; Yang, S.; et al. miR-92b controls glioma proliferation and invasion through regulating Wnt/beta-catenin signaling via Nemo-like kinase. Neuro Oncol. 2013, 15, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhuo, H.Z.; Wu, J.Y.; Lin, L.Y.; Huang, Z.L.; Lu, J.X.; Cheng, K.L. MiR-92b inhibits proliferation and invasion of lung cancer by targeting EZH2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3166–3173. [Google Scholar] [CrossRef]
- Long, M.; Zhan, M.; Xu, S.; Yang, R.; Chen, W.; Zhang, S.; Shi, Y.; He, Q.; Mohan, M.; Liu, Q.; et al. miR-92b-3p acts as a tumor suppressor by targeting Gabra3 in pancreatic cancer. Mol. Cancer 2017, 16, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Jing, C.; Li, L.; Huang, F.; Ding, F.; Wang, B.; Lin, D.; Luo, A.; Liu, Z. MicroRNA-92b represses invasion-metastasis cascade of esophageal squamous cell carcinoma. Oncotarget 2016, 7, 20209–20222. [Google Scholar] [CrossRef] [Green Version]
- Creff, J.; Besson, A. Functional versatility of the CDK inhibitor p57(Kip2). Front. Cell Dev. Biol. 2020, 8, 584590. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Tian, T.; Nan, K.; Wang, W. p57: A multifunctional protein in cancer (Review). Int. J. Oncol. 2010, 36, 1321–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ren, D.; Wu, X.; Lin, X.; Ye, L.; Lin, C.; Wu, S.; Zhu, J.; Peng, X.; Song, L. miR-1266 contributes to pancreatic cancer progression and chemoresistance by the STAT3 and NF-kappaB signaling pathways. Mol. Ther. Nucleic. Acids 2018, 11, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Krek, A.; Grun, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Borriello, A.; Caldarelli, I.; Bencivenga, D.; Criscuolo, M.; Cucciolla, V.; Tramontano, A.; Oliva, A.; Perrotta, S.; Della Ragione, F. p57(Kip2) and cancer: Time for a critical appraisal. Mol. Cancer Res. 2011, 9, 1269–1284. [Google Scholar] [CrossRef] [Green Version]
- Duquesnes, N.; Callot, C.; Jeannot, P.; Daburon, V.; Nakayama, K.I.; Manenti, S.; Davy, A.; Besson, A. p57(Kip2) knock-in mouse reveals CDK-independent contribution in the development of Beckwith-Wiedemann syndrome. J. Pathol. 2016, 239, 250–261. [Google Scholar] [CrossRef]
- Kazi, A.; Dou, Q.P. Cell cycle and drug sensitivity. Methods Mol. Med. 2005, 111, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Zhou, C.; Liu, H.; Gao, J.; Li, Q.; Mu, D.; Ma, Z. Rapamycin sensitizes T-ALL cells to dexamethasone-induced apoptosis. J. Exp. Clin. Cancer Res. 2010, 29, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakitina, T.V.; Vasilevskaya, I.A.; O’Dwyer, P.J. Inhibition of G1/S transition potentiates oxaliplatin-induced cell death in colon cancer cell lines. Biochem. Pharmacol. 2007, 73, 1715–1726. [Google Scholar] [CrossRef]
- Rossi, M.N.; Antonangeli, F. Cellular Response upon Stress: p57 Contribution to the Final Outcome. Mediat. Inflamm. 2015, 2015, 259325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuelsson, M.K.; Pazirandeh, A.; Okret, S. A pro-apoptotic effect of the CDK inhibitor p57(Kip2) on staurosporine-induced apoptosis in HeLa cells. Biochem. Biophys. Res. Commun. 2002, 296, 702–709. [Google Scholar] [CrossRef]
- Vlachos, P.; Nyman, U.; Hajji, N.; Joseph, B. The cell cycle inhibitor p57(Kip2) promotes cell death via the mitochondrial apoptotic pathway. Cell Death Differ. 2007, 14, 1497–1507. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drugs | IC50 (Mean ± SD, μM) | Fold Resistance | IC50 (Mean ± SD, μM) | Fold Reversal | ||
|---|---|---|---|---|---|---|
| HCT8 NC | HCT8 miR-92b-3p | HCT8/T shNC | HCT8/T shmiR-92b-3p | |||
| PTX | 0.0286 ± 0.0044 | 0.1279 ± 0.0154 *** | 4.47 | 6.92 ± 0.50 | 2.24 ± 0.28 *** | 3.09 |
| DOX | 0.1217 ± 0.0093 | 0.3418 ± 0.0318 *** | 2.81 | 2.73 ± 0.10 | 1.55 ± 0.06 *** | 1.76 |
| NVB | 0.0377 ± 0.0024 | 0.1331 ± 0.0158 *** | 3.53 | 1.04 ± 0.04 | 0.51 ± 0.05 *** | 2.04 |
| VCR | 0.1949 ± 0.0207 | 1.3090 ± 0.1442 *** | 6.72 | 2.65 ± 0.05 | 1.26 ± 0.05 *** | 2.10 |
| VP-16 | 2.0910 ± 0.0995 | 7.4720 ± 1.2260 *** | 3.57 | 20.22 ± 0.54 | 11.92 ± 0.93 *** | 1.70 |
| Drugs | IC50 (Mean ± SD, μM) | Fold Resistance | IC50 (Mean ± SD, μM) | Fold Reversal | ||
|---|---|---|---|---|---|---|
| HCT8 siNC | HCT8 siCDKN1C-1 | HCT8/T Vector | HCT8/T CDKN1C | |||
| PTX | 0.0188 ± 0.0027 | 0.1034 ± 0.0182 *** | 5.50 | 6.27 ± 0.52 | 2.89 ± 0.23 *** | 2.17 |
| DOX | 0.1142 ± 0.0164 | 0.2999 ± 0.0625 *** | 2.63 | 2.69 ± 0.21 | 1.64 ± 0.12 *** | 1.64 |
| NVB | 0.0386 ± 0.0035 | 0.1168 ± 0.0192 *** | 3.03 | 0.96 ± 0.05 | 0.54 ± 0.04 *** | 1.78 |
| VCR | 0.1300 ± 0.0061 | 0.5744 ± 0.0864 *** | 4.42 | 2.53 ± 0.19 | 1.45 ± 0.20 *** | 1.74 |
| VP-16 | 1.9590 ± 0.1971 | 6.7030 ± 0.7310 *** | 3.42 | 18.89 ± 1.40 | 11.96 ± 1.53 *** | 1.58 |
| Drugs | IC50 (Mean ± SD, μM) | |||
|---|---|---|---|---|
| HCT8 NC | HCT8 miR-92b-3p | HCT8 miR-92b-3p+Vector | HCT8 miR-92b-3p+CDKN1C | |
| PTX | 0.0282 ± 0.0052 | 0.1252 ± 0.0421 *** | 0.1213 ± 0.0353 | 0.0566 ± 0.0140 ** |
| DOX | 0.1130 ± 0.0227 | 0.3400 ± 0.0606 *** | 0.3058 ± 0.0255 | 0.1896 ± 0.0347 *** |
| NVB | 0.0362 ± 0.0029 | 0.1268 ± 0.0193 *** | 0.1417 ± 0.0297 | 0.0618 ± 0.0062 *** |
| VCR | 0.2046 ± 0.0374 | 1.3190 ± 0.3520 *** | 1.3310 ± 0.1793 | 0.3749 ± 0.0643 *** |
| VP-16 | 2.1490 ± 0.3348 | 7.4280 ± 1.1860 *** | 7.5020 ± 1.0160 | 3.2020 ± 0.2855 *** |
| Drugs | IC50 (Mean ± SD, μM) | |||
|---|---|---|---|---|
| HCT8/T shNC | HCT8/T shmiR-92b-3p | HCT8/T shmiR-92b-3p+siNC | HCT8/T shmiR-92b-3p+siCDKN1C-1 | |
| PTX | 7.52 ± 0.68 | 2.29 ± 0.13 *** | 2.38 ± 0.13 | 5.33 ± 0.62 *** |
| DOX | 3.15 ± 0.18 | 1.50 ± 0.07 *** | 1.48 ± 0.13 | 2.63 ± 0.10 *** |
| NVB | 1.44 ± 0.11 | 0.59 ± 0.05 *** | 0.65 ± 0.04 | 1.18 ± 0.06 *** |
| VCR | 3.43 ± 0.08 | 1.33 ± 0.10 *** | 1.38 ± 0.07 | 2.56 ± 0.27 *** |
| VP-16 | 22.88 ± 1.97 | 11.94 ± 2.23 *** | 10.91 ± 1.17 | 20.77 ± 0.78 *** |
| Cell Lines | Group | Body Weight (g) | Tumor Volume (mm3) | Tumor Weight (g) | RTV | T/C | TGI (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| D11 | D31 | D11 | D31 | ||||||
| HCT8/T shNC | Vehicle | 17.87 ± 0.20 | 18.44 ± 0.43 | 75.42 ± 8.31 | 1235.14 ± 182.40 | 0.76 ± 0.12 | 16.72 ± 2.00 | - | - |
| NVB | 17.40 ± 0.53 | 16.60 ± 0.71 | 74.80 ± 8.34 | 1259.77 ± 100.69 | 0.72 ± 0.09 | 17.65 ± 1.65 | 1.05 | −2.18 | |
| PTX | 18.03 ± 0.36 | 17.43 ± 0.66 | 74.54 ± 8.36 | 1050.67 ± 163.01 | 0.58 ± 0.10 | 14.17 ± 1.61 | 0.85 | 15.83 | |
| HCT8/T shmiR-92b-3p | Vehicle | 18.16 ± 0.71 | 19.21 ± 0.84 | 114.00 ± 17.20 | 1999.24 ± 336.23 | 1.26 ± 0.21 | 18.48 ± 2.96 | - | - |
| NVB | 17.53 ± 0.59 | 17.00 ± 0.60 | 113.20 ± 15.06 | 1162.95 ± 70.84 * | 0.77 ± 0.07 * | 10.96 ± 0.89 * | 0.65 | 44.32 | |
| PTX | 17.74 ± 0.53 | 17.29 ± 0.53 | 111.30 ± 16.92 | 992.94 ± 152.68 * | 0.68 ± 0.07 * | 9.29 ± 0.93 * | 0.55 | 53.23 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, F.; Yang, Z.; Gu, X.; Feng, L.; Xu, M.; Zhang, X. miR-92b-3p Regulates Cell Cycle and Apoptosis by Targeting CDKN1C, Thereby Affecting the Sensitivity of Colorectal Cancer Cells to Chemotherapeutic Drugs. Cancers 2021, 13, 3323. https://doi.org/10.3390/cancers13133323
Zhao F, Yang Z, Gu X, Feng L, Xu M, Zhang X. miR-92b-3p Regulates Cell Cycle and Apoptosis by Targeting CDKN1C, Thereby Affecting the Sensitivity of Colorectal Cancer Cells to Chemotherapeutic Drugs. Cancers. 2021; 13(13):3323. https://doi.org/10.3390/cancers13133323
Chicago/Turabian StyleZhao, Fangqing, Zhongmin Yang, Xiaofan Gu, Lixing Feng, Mingshi Xu, and Xiongwen Zhang. 2021. "miR-92b-3p Regulates Cell Cycle and Apoptosis by Targeting CDKN1C, Thereby Affecting the Sensitivity of Colorectal Cancer Cells to Chemotherapeutic Drugs" Cancers 13, no. 13: 3323. https://doi.org/10.3390/cancers13133323