
MicroRNAs Targeting HIF-2α, VEGFR1 and/or VEGFR2 as Potential Predictive Biomarkers for VEGFR Tyrosine Kinase and HIF-2α Inhibitors in Metastatic Clear-Cell Renal Cell Carcinoma

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Selection
2.2. Study Objectives and Endpoints
2.3. miRNA Extraction and Next-Generation Sequencing (NGS)
2.4. mRNA Extraction and qRT-PCR
2.5. Selection of miRNAs
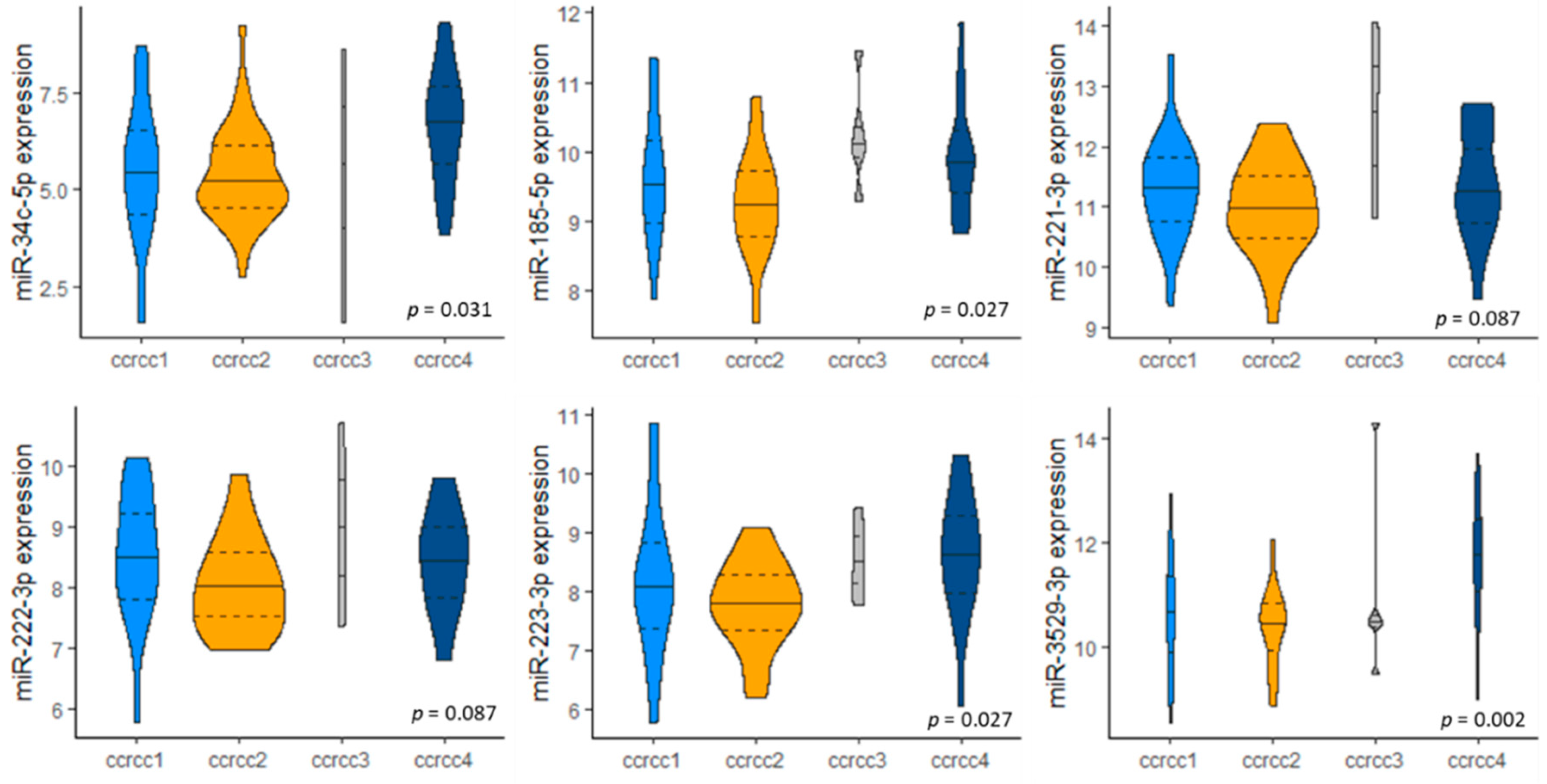
2.6. Molecular Subtypes
2.7. Clinical Data
3. Results
3.1. Included Patients
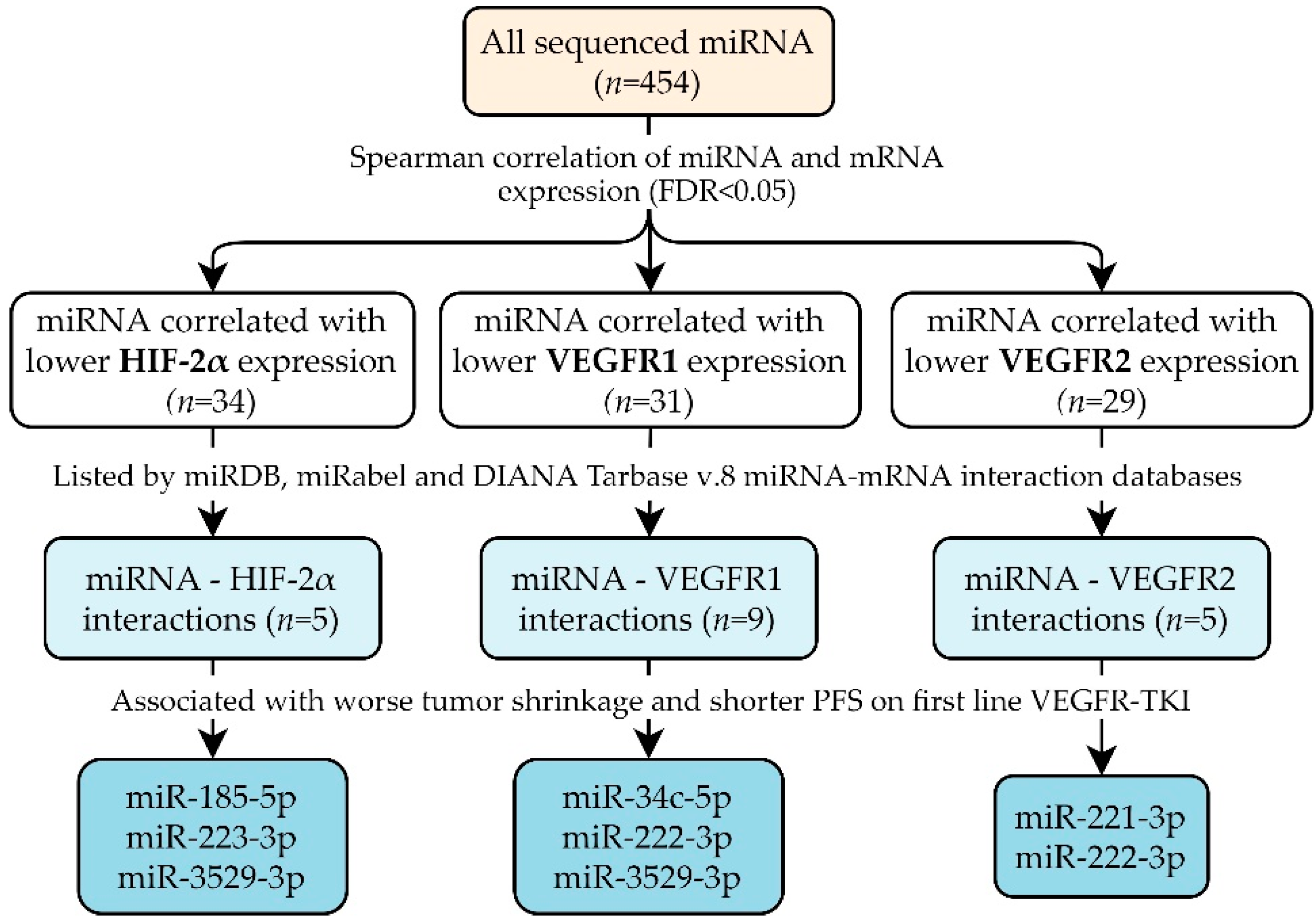
3.2. miRNAs Potentially Targeting HIF-2α, VEGFR1 and/or VEGFR2
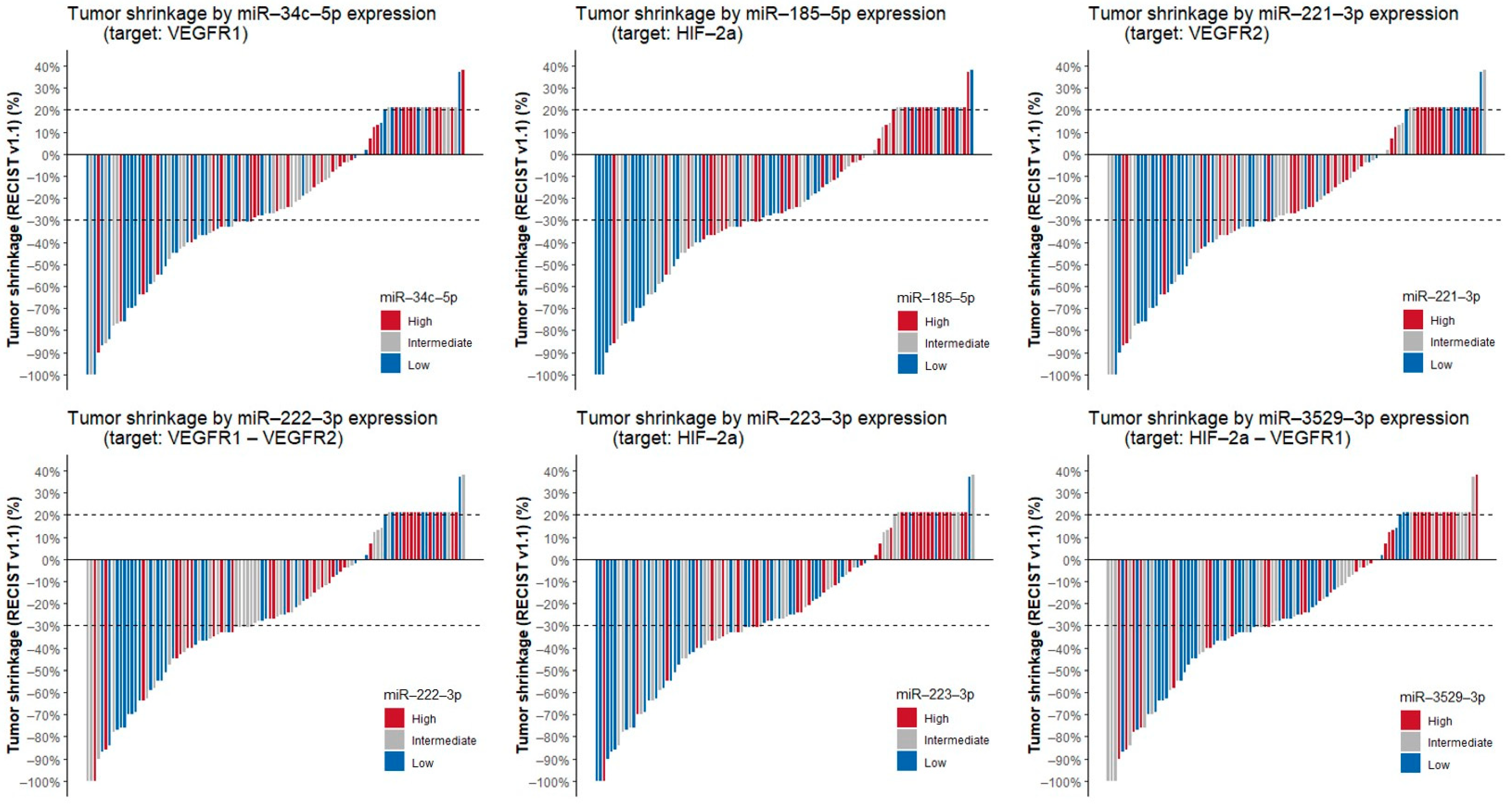
3.3. Correlation of miRNAs with Tumor Shrinkage, PFS and OS
3.4. miRNA Expression in Molecular Subclassification ccrcc1-4
4. Discussion
4.1. miR-34c-5p Targets VEGFR1
4.2. miR-185-5p Targets HIF-2α
4.3. miR-221-3p and miR-222-3p Both Target VEGFR2, miR-222-3p Targets VEGFR1
4.4. miR-223-3p Targets HIF-2α
4.5. miR-3529-3p Targets Both HIF-2α and VEGFR1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis. Prim. 2017, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Linehan, W.M.; Ricketts, C.J. The Cancer Genome Atlas of renal cell carcinoma: Findings and clinical implications. Nat. Rev. Urol. 2019, 16, 539–552. [Google Scholar] [CrossRef]
- Motzer, R.J.; Hutson, T.E.; Tomczak, P.; Michaelson, M.D.; Bukowski, R.M.; Rixe, O.; Oudard, S.; Negrier, S.; Szczylik, C.; Kim, S.T.; et al. Sunitinib versus Interferon Alfa in Metastatic Renal-Cell Carcinoma. N. Engl. J. Med. 2007, 356, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, T.H.; Li, Q.; Ma, H.-W.; Key, J.; Zhang, L.; Chen, R.; Garcia, J.A.; Naidoo, J.; Longgood, J.; Frantz, D.E.; et al. Allosteric inhibition of hypoxia inducible factor-2 with small molecules. Nat. Chem. Biol. 2013, 9, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Courtney, K.D.; Ma, Y.; de Leon, A.D.; Christie, A.; Xie, Z.; Woolford, L.; Singla, N.; Joyce, A.; Hill, H.; Madhuranthakam, A.J.; et al. HIF-2 Complex Dissociation, Target Inhibition, and Acquired Resistance with PT2385, a First-in-Class HIF-2 Inhibitor, in Patients with Clear Cell Renal Cell Carcinoma. Clin. Cancer Res. 2020, 26, 793–803. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Kaelin, W.G., Jr. Targeting the HIF2–VEGF axis in renal cell carcinoma. Nat. Med. 2020, 26, 1519–1530. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Bauer, T.M.; Papadopoulos, K.P.; Plimack, E.R.; Merchan, J.R.; McDermott, D.F.; Michaelson, M.D.; Appleman, L.J.; Thamake, S.; Perini, R.F.; et al. Inhibition of hypoxia-inducible factor-2α in renal cell carcinoma with belzutifan: A phase 1 trial and biomarker analysis. Nat. Med. 2021, 27, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Kotecha, R.R.; Motzer, R.J.; Voss, M.H. Towards individualized therapy for metastatic renal cell carcinoma. Nat. Rev. Clin. Oncol. 2019, 16, 621–633. [Google Scholar] [CrossRef]
- Osanto, S.; Qin, Y.; Buermans, H.P.; Berkers, J.; Lerut, E.; Goeman, J.J.; Van Poppel, H. Genome-Wide MicroRNA Expression Analysis of Clear Cell Renal Cell Carcinoma by Next Generation Deep Sequencing. PLoS ONE 2012, 7, e38298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, N.M.A.; Bao, T.T.; Grigull, J.; Youssef, Y.M.; Girgis, A.; Diamandis, M.; Fatoohi, E.; Metias, M.; Honey, R.J.; Stewart, R.; et al. miRNA Profiling for Clear Cell Renal Cell Carcinoma: Biomarker Discovery and Identification of Potential Controls and Consequences of miRNA Dysregulation. J. Urol. 2011, 186, 1077–1083. [Google Scholar] [CrossRef]
- Qin, S.; Shi, X.; Wang, C.; Jin, P.; Ma, F. Transcription Factor and miRNA Interplays Can Manifest the Survival of ccRCC Patients. Cancers 2019, 11, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, L.; Liang, J.; Deng, X.; Wu, J. miRNAs in Prediction of Prognosis in Clear Cell Renal Cell Carcinoma. BioMed. Res. Int. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Chen, L.; Wang, G.; Xiao, Y.; Ju, L.; Wang, X. Identification of a three-miRNA signature as a novel potential prognostic biomarker in patients with clear cell renal cell carcinoma. J. Cell. Biochem. 2019, 120, 13751–13764. [Google Scholar] [CrossRef] [PubMed]
- Go, H.; Kang, M.J.; Kim, P.-J.; Lee, J.-L.; Park, J.Y.; Park, J.-M.; Ro, J.Y.; Cho, Y.M. Development of Response Classifier for Vascular Endothelial Growth Factor Receptor (VEGFR)-Tyrosine Kinase Inhibitor (TKI) in Metastatic Renal Cell Carcinoma. Pathol. Oncol. Res. 2019, 25, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Khella, H.W.Z.; Butz, H.; Ding, Q.; Rotondo, F.; Evans, K.R.; Kupchak, P.; Dharsee, M.; Latif, A.; Pasic, M.D.; Lianidou, E.; et al. miR-221/222 Are Involved in Response to Sunitinib Treatment in Metastatic Renal Cell Carcinoma. Mol. Ther. 2015, 23, 1748–1758. [Google Scholar] [CrossRef]
- Gámez-Pozo, A.; Antón-Aparicio, L.M.; Bayona, C.; Borrega, P.; Gallegos Sancho, M.I.; García-Domínguez, R.; de Portugal, T.; Ramos-Vázquez, M.; Pérez-Carrión, R.; Bolós, M.V.; et al. MicroRNA Expression Profiling of Peripheral Blood Samples Predicts Resistance to First-line Sunitinib in Advanced Renal Cell Carcinoma Patients. Neoplasia 2012, 14, 1144–IN14. [Google Scholar] [CrossRef] [Green Version]
- Verbiest, A.; Couchy, G.; Job, S.; Zucman-Rossi, J.; Caruana, L.; Lerut, E.; Oyen, R.; de Reyniès, A.; Laguerre, B.; Rioux-Leclercq, N.; et al. Molecular Subtypes of Clear Cell Renal Cell Carcinoma Are Associated With Outcome During Pazopanib Therapy in the Metastatic Setting. Clin. Genitourin. Cancer 2018, 16, e605–e612. [Google Scholar] [CrossRef]
- Beuselinck, B.; Job, S.; Becht, E.; Karadimou, A.; Verkarre, V.; Couchy, G.; Giraldo, N.; Rioux-Leclercq, N.; Molinié, V.; Sibony, M.; et al. Molecular Subtypes of Clear Cell Renal Cell Carcinoma Are Associated with Sunitinib Response in the Metastatic Setting. Clin. Cancer Res. 2015, 21, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- García-Donas, J.; Beuselinck, B.; Inglada-Pérez, L.; Graña, O.; Schöffski, P.; Wozniak, A.; Bechter, O.; Apellániz-Ruiz, M.; Leandro-García, L.J.; Esteban, E.; et al. Deep sequencing reveals microRNAs predictive of antiangiogenic drug response. JCI Insight 2016, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beuselinck, B.; Verbiest, A.; Couchy, G.; Job, S.; De Reynies, A.; Meiller, C.; Albersen, M.; Verkarre, V.; Lerut, E.; Méjean, A.; et al. Pro-angiogenic gene expression is associated with better outcome on sunitinib in metastatic clear-cell renal cell carcinoma. Acta Oncol. 2018, 57, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quillet, A.; Saad, C.; Ferry, G.; Anouar, Y.; Vergne, N.; Lecroq, T.; Dubessy, C. Improving Bioinformatics Prediction of microRNA Targets by Ranks Aggregation. Front. Genet. 2020, 10, 1330. [Google Scholar] [CrossRef] [Green Version]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA–gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, D.Y.C.; Xie, W.; Regan, M.M.; Warren, M.A.; Golshayan, A.R.; Sahi, C.; Eigl, B.J.; Ruether, J.D.; Cheng, T.; North, S.; et al. Prognostic Factors for Overall Survival in Patients With Metastatic Renal Cell Carcinoma Treated With Vascular Endothelial Growth Factor–Targeted Agents: Results From a Large, Multicenter Study. J. Clin. Oncol. 2009, 27, 5794–5799. [Google Scholar] [CrossRef]
- Islam, F.; Gopalan, V.; Vider, J.; Lu, C.-T.; Lam, A.K.-Y. MiR-142-5p act as an oncogenic microRNA in colorectal cancer: Clinicopathological and functional insights. Exp. Mol. Pathol. 2018, 104, 98–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.; Zhai, B.; Zheng, Y.; Ren, R.; Han, M.; Wang, X. Transcatheter arterial infusion chemotherapy increases expression level of miR-142-5p in stage III colorectal cancer. Indian J. Cancer 2015, 52, 47. [Google Scholar] [CrossRef]
- Ho, J.J.D.; Metcalf, J.L.; Yan, M.S.; Turgeon, P.J.; Wang, J.J.; Chalsev, M.; Petruzziello-Pellegrini, T.N.; Tsui, A.K.Y.; He, J.Z.; Dhamko, H.; et al. Functional Importance of Dicer Protein in the Adaptive Cellular Response to Hypoxia. J. Biol. Chem. 2012, 287, 29003–29020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.-J.; Lu, L.-G.; Tao, K.-Z.; Chen, D.-F.; Xia, Q.; Weng, J.-J.; Zhu, F.; Wang, X.-P.; Zheng, P. MicroRNA-185 suppresses growth and invasion of colon cancer cells through inhibition of the hypoxia-inducible factor-2α pathway in vitro and in vivo. Mol. Med. Rep. 2014, 10, 2401–2408. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Qi, W.; Dai, S.; Zou, G.; Liu, W.; Yu, B.; Tang, J. microRNA-223 promotes autophagy to aggravate lung ischemia-reperfusion injury by inhibiting the expression of transcription factor HIF2α. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 319, L1–L10. [Google Scholar] [CrossRef]
- Choi, Y.-C.; Yoon, S.; Jeong, Y.; Yoon, J.; Baek, K. Regulation of vascular endothelial growth factor signaling by miR-200b. Mol. Cells 2011, 32, 77–82. [Google Scholar] [CrossRef]
- Lee, J.S.; Song, D.W.; Park, J.H.; Kim, J.O.; Cho, C.; Kim, D.H. miR-374 promotes myocardial hypertrophy by negatively regulating vascular endothelial growth factor receptor-1 signaling. BMB Rep. 2017, 50, 208–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, H.; Zhang, J.; Pan, T.; Ai, S.; Tang, L.; Wang, F. miR-378a enhances the sensitivity of liver cancer to sorafenib by targeting VEGFR, PDGFRβ and c-Raf. Mol. Med. Rep. 2018, 17, 4581–4588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Tu, Y.; Wan, L.; Bu, L.; Huang, T.; Sun, X.; Wang, K.; Shen, B. In Vivo Monitoring of Angiogenesis Inhibition via Down-Regulation of Mir-21 in a VEGFR2-Luc Murine Breast Cancer Model Using Bioluminescent Imaging. PLoS ONE 2013, 8, e71472. [Google Scholar] [CrossRef]
- Hou, W.-Z.; Chen, X.-L.; Wu, W.; Hang, C.-H. MicroRNA-370-3p inhibits human vascular smooth muscle cell proliferation via targeting KDR/AKT signaling pathway in cerebral aneurysm. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1080–1087. [Google Scholar] [PubMed]
- Wang, X.H.; Chen, L. MicroRNA-370 suppresses the retinal capillary endothelial cell growth by targeting KDR gene. Bratisl. Med. J. 2017, 118, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Krebs, M.; Solimando, A.G.; Kalogirou, C.; Marquardt, A.; Frank, T.; Sokolakis, I.; Hatzichristodoulou, G.; Kneitz, S.; Bargou, R.; Kübler, H.; et al. miR-221-3p Regulates VEGFR2 Expression in High-Risk Prostate Cancer and Represents an Escape Mechanism from Sunitinib In Vitro. J. Clin. Med. 2020, 9, 670. [Google Scholar] [CrossRef] [Green Version]
- Pourmir, I.; Noel, J.; Simonaggio, A.; Oudard, S.; Vano, Y.-A. Update on the most promising biomarkers of response to immune checkpoint inhibitors in clear cell renal cell carcinoma. World J. Urol. 2021, 1–9. [Google Scholar] [CrossRef]
- Xu, Z.; Huang, B.; Zhang, Q.; He, X.; Wei, H.; Zhang, D. NOTCH1 regulates the proliferation and migration of bladder cancer cells by cooperating with long non-coding RNA HCG18 and microRNA-34c-5p. J. Cell. Biochem. 2019, 120, 6596–6604. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Wang, G.; Liu, H.; Xiong, C. SATB2 targeted by methylated miR-34c-5p suppresses proliferation and metastasis attenuating the epithelial-mesenchymal transition in colorectal cancer. Cell Prolif. 2018, 51, e12455. [Google Scholar] [CrossRef]
- Daugaard, I.; Knudsen, A.; Kjeldsen, T.E.; Hager, H.; Hansen, L.L. The association between miR-34 dysregulation and distant metastases formation in lung adenocarcinoma. Exp. Mol. Pathol. 2017, 102, 484–491. [Google Scholar] [CrossRef]
- Re, M.; Magliulo, G.; Gioacchini, F.M.; Bajraktari, A.; Bertini, A.; Çeka, A.; Rubini, C.; Ferrante, L.; Procopio, A.D.; Olivieri, F. Expression Levels and Clinical Significance of miR-21-5p, miR-let-7a, and miR-34c-5p in Laryngeal Squamous Cell Carcinoma. BioMed. Res. Int. 2017, 2017, 3921258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Wang, X.; Niu, X.; Jiao, R.; Li, X.; Wang, S. miR-34c-5p targets Notch1 and suppresses the metastasis and invasion of cervical cancer. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Petrozza, V.; Carbone, A.; Bellissimo, T.; Porta, N.; Palleschi, G.; Pastore, A.L.; Di Carlo, A.; Della Rocca, C.; Fazi, F. Oncogenic MicroRNAs Characterization in Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2015, 16, 29219–29225. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhong, W.; Zhu, M.; Li, M.; Yang, Z. miR-185 inhibits prostate cancer angiogenesis induced by the nodal/ALK4 pathway. BMC Urol. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhao, Y. MiR-185-5p Protects Against Angiogenesis in Polycystic Ovary Syndrome by Targeting VEGFA. Front. Pharmacol. 2020, 11, 1030. [Google Scholar] [CrossRef]
- Razi, M.H.; Eftekhar, M.; Ghasemi, N.; Sheikhha, M.H.; Firoozabadi, A.D. Expression levels of circulatory mir-185-5p, vascular endothelial growth factor, and platelet-derived growth factor target genes in endometriosis. Int. J. Reprod. Biomed. 2020, 18, 347–358. [Google Scholar] [CrossRef]
- Ma, X.; Shen, D.; Li, H.; Zhang, Y.; Lv, X.; Huang, Q.; Gao, Y.; Li, X.; Gu, L.; Xiu, S.; et al. MicroRNA-185 inhibits cell proliferation and induces cell apoptosis by targeting VEGFA directly in von Hippel-Lindau–inactivated clear cell renal cell carcinoma. Urol. Oncol. Semin. Orig. Investig. 2015, 33, 169.e1–169.e11. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Tang, G. miR-185-5p targets ROCK2 and inhibits cell migration and invasion of hepatocellular carcinoma. Oncol. Lett. 2019, 17, 5087–5093. [Google Scholar] [CrossRef] [Green Version]
- Ostadrahimi, S.; Valugerdi, M.A.; Hassan, M.; Haddad, G.; Fayaz, S.; Parvizhamidi, M.; Mahdian, R.; Esfahani, P.F. miR-1266-5p and miR-185-5p Promote Cell Apoptosis in Human Prostate Cancer Cell Lines. Asian Pac. J. Cancer Prev. 2018, 19, 2305. [Google Scholar] [CrossRef]
- Yuan, H.-X.; Zhang, J.-P.; Kong, W.-T.; Liu, Y.-J.; Lin, Z.-M.; Wang, W.-P.; Guo, J.-M. Elevated microRNA-185 is associated with high vascular endothelial growth factor receptor 2 expression levels and high microvessel density in clear cell renal cell carcinoma. Tumor Biol. 2014, 35, 12757–12763. [Google Scholar] [CrossRef]
- Dias, F.; Teixeira, A.L.; Ferreira, M.; Adem, B.; Bastos, N.; Vieira, J.; Fernandes, M.; Sequeira, M.I.; Maurício, J.; Lobo, F.; et al. Plasmatic miR-210, miR-221 and miR-1233 profile: Potential liquid biopsies candidates for renal cell carcinoma. Oncotarget 2017, 8, 103315–103326. [Google Scholar] [CrossRef] [PubMed]
- Urbich, C.; Kuehbacher, A.; Dimmeler, S. Role of microRNAs in vascular diseases, inflammation, and angiogenesis. Cardiovasc. Res. 2008, 79, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Mujahid, S.; Nielsen, H.C.; Volpe, M.A.V. MiR-221 and miR-130a Regulate Lung Airway and Vascular Development. PLoS ONE 2013, 8, e55911. [Google Scholar] [CrossRef] [Green Version]
- Poliseno, L.; Tuccoli, A.; Mariani, L.; Evangelista, M.; Citti, L.; Woods, K.; Mercatanti, A.; Hammond, S.; Rainaldi, G. MicroRNAs modulate the angiogenic properties of HUVECs. Blood 2006, 108, 3068–3071. [Google Scholar] [CrossRef]
- Yoshizuka, M.; Nakasa, T.; Kawanishi, Y.; Hachisuka, S.; Furuta, T.; Miyaki, S.; Adachi, N.; Ochi, M. Inhibition of microRNA-222 expression accelerates bone healing with enhancement of osteogenesis, chondrogenesis, and angiogenesis in a rat refractory fracture model. J. Orthop. Sci. 2016, 21, 852–858. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, A.L.; Dias, F.; Ferreira, M.; Gomes, M.; Santos, J.I.; Lobo, F.; Maurício, J.; Machado, J.C.; Medeiros, R. Combined Influence of EGF+61G>A and TGFB+869T>C Functional Polymorphisms in Renal Cell Carcinoma Progression and Overall Survival: The Link to Plasma Circulating MiR-7 and MiR-221/222 Expression. PLoS ONE 2015, 10, e0103258. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.L.; Ferreira, M.; Silva, J.; Gomes, M.; Dias, F.; Santos, J.I.; Maurício, J.; Lobo, F.; Medeiros, R. Higher circulating expression levels of miR-221 associated with poor overall survival in renal cell carcinoma patients. Tumor Biol. 2014, 35, 4057–4066. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, C.; Lu, Z.; Guo, L.; Ge, Q. Analysis of serum genome-wide microRNAs for breast cancer detection. Clin. Chim. Acta 2012, 413, 1058–1065. [Google Scholar] [CrossRef]
- Jiang, K.; Li, G.; Chen, W.; Song, L.; Wei, T.; Li, Z.; Gong, R.; Lei, J.; Shi, H.; Zhu, J. Plasma Exosomal miR-146b-5p and miR-222-3p are Potential Biomarkers for Lymph Node Metastasis in Papillary Thyroid Carcinomas. OncoTargets Ther. 2020, 13, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Lu, Y.; Wang, H.; Han, X.; Mao, J.; Li, J.; Yu, L.; Wang, B.; Fan, S.; Yu, X.; et al. miR-221/222 enhance the tumorigenicity of human breast cancer stem cells via modulation of PTEN/Akt pathway. Biomed. Pharmacother. 2016, 79, 93–101. [Google Scholar] [CrossRef]
- Rong, M.; Chen, G.; Dang, Y. Increased MiR-221 expression in hepatocellular carcinoma tissues and its role in enhancing cell growth and inhibiting apoptosis in vitro. BMC Cancer 2013, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Che, Q.; Qiu, H.; Bao, W.; Chen, X.; Lu, W.; Li, B.; Wan, X. Elevated MiR-222-3p Promotes Proliferation and Invasion of Endometrial Carcinoma via Targeting ERα. PLoS ONE 2014, 9, e87563. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, M.; Di Leva, G.; Romano, G.; Nuovo, G.; Suh, S.-S.; Ngankeu, A.; Taccioli, C.; Pichiorri, F.; Alder, H.; Secchiero, P.; et al. miR-221&222 Regulate TRAIL Resistance and Enhance Tumorigenicity through PTEN and TIMP3 Downregulation. Cancer Cell 2009, 16, 498–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravegnini, G.; Cargnin, S.; Sammarini, G.; Zanotti, F.; Bermejo, J.L.; Hrelia, P.; Terrazzino, S.; Angelini, S. Prognostic Role of miR-221 and miR-222 Expression in Cancer Patients: A Systematic Review and Meta-Analysis. Cancers 2019, 11, 970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacova, J.; Juracek, J.; Poprach, A.; Buchler, T.; Kopecky, J.; Fiala, O.; Svoboda, M.; Slaby, O. Candidate MicroRNA Biomarkers of Therapeutic Response to Sunitinib in Metastatic Renal Cell Carcinoma: A Validation Study in Patients with Extremely Good and Poor Response. Anticancer Res. 2018, 38, 2961–2965. [Google Scholar] [CrossRef]
- Vergho, D.C.; Kneitz, S.; Kalogirou, C.; Burger, M.; Krebs, M.; Rosenwald, A.; Spahn, M.; Löser, A.; Kocot, A.; Riedmiller, H.; et al. Impact of miR-21, miR-126 and miR-221 as Prognostic Factors of Clear Cell Renal Cell Carcinoma with Tumor Thrombus of the Inferior Vena Cava. PLoS ONE 2014, 9, e109877. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.-H.; Liu, Y.; Xiao, L.-M.; Chen, L.K.; Zheng, S.-Y.; Zeng, E.-M.; Li, D.-H.; Li, Y.-P. Silencing microRNA-221/222 cluster suppresses glioblastoma angiogenesis by suppressor of cytokine signaling-3-dependent JAK/STAT pathway. J. Cell. Physiol. 2019, 234, 22272–22284. [Google Scholar] [CrossRef]
- Bozec, A.; Zangari, J.; Butori-Pepino, M.; Ilie, M.; Lalvee, S.; Juhel, T.; Butori, C.; Brest, P.; Hofman, P.; Vouret-Craviari, V. MiR-223-3p inhibits angiogenesis and promotes resistance to cetuximab in head and neck squamous cell carcinoma. Oncotarget 2017, 8, 57174–57186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, G.-H.; Ma, P.-Z.; Song, X.-B.; Liu, N.; Zhang, T.; Wu, B. MicroRNA-223-3p Inhibits the Angiogenesis of Ischemic Cardiac Microvascular Endothelial Cells via Affecting RPS6KB1/hif-1a Signal Pathway. PLoS ONE 2014, 9, e108468. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Wang, X.; Wang, T.; Xing, J. MiR-223-3p promotes cell proliferation and metastasis by downregulating SLC4A4 in clear cell renal cell carcinoma. Aging 2019, 11, 615–633. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Yan, X.; Pan, Y.; Wang, Y.; Wang, N.; Li, L.; Liu, Y.; Chen, X.; Zhang, C.-Y.; Gu, H.; et al. MicroRNA-223 delivered by platelet-derived microvesicles promotes lung cancer cell invasion via targeting tumor suppressor EPB41L3. Mol. Cancer 2015, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Ma, W.; Zeng, Q.; Tan, J.; Cao, K.; Luo, L. Identification of miRNA-Based Signature as a Novel Potential Prognostic Biomarker in Patients with Breast Cancer. Dis. Markers 2019, 2019, 3815952. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Lu, Z.; Li, H.; Lu, J.; Guo, L.; Ge, Q. Next-Generation Sequencing of MicroRNAs for Breast Cancer Detection. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Wang, L.; Zhu, Z.; Gao, W.; Li, D.; Zhou, Z.; Chen, L.; Fu, C.-G. Downregulation of miR-423-5p Contributes to the Radioresistance in Colorectal Cancer Cells. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Patients | n = 109 | |
| Gender: male | 70 | 64.2% |
| Gender: female | 39 | 35.8% |
| Median age at diagnosis (years) | 62 | IQR: 55–68 |
| Median OS after diagnosis (months) | 46 | IQR: 21–110 |
| Median OS after stage IV (months) | 32 | IQR: 15–64 |
| IMDC risk group at start of first-line therapy | ||
| Favorable (n) | 13 | 11.9% |
| Intermediate (n) | 69 | 63.3% |
| Poor (n) | 27 | 24.8% |
| First-line targeted therapy | ||
| Sunitinib (n) | 68 | 62.4% |
| Pazopanib (n) | 30 | 10.1% |
| Sorafenib (n) | 11 | 27.5% |
| Molecular subtypes | ||
| ccrcc1 | 19/76 | 25% |
| ccrcc2 | 33/76 | 43.4% |
| ccrcc3 | 4/76 | 5.2% |
| ccrcc4 | 20/76 | 26.3% |
| miRNA | Interaction with HIF-2α | Interaction with VEGFR1 | Interaction with VEGFR2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rho | p-Value | Interaction Database | Experimental Validation | rho | p-Value | Interaction Database | Experimental Validation | rho | p-Value | Interaction Database | Experimental Validation | |
| miR-21-3p | 0.375 | 0.006 | yes 3 | – | 0.355 | 0.009 | – | – | 0.317 | 0.022 | yes 3 | [34] |
| miR-142-5p | 0.305 | 0.028 | yes 1,2,3 | [26,27] | 0.199 | 0.182 | – | – | 0.247 | 0.085 | – | – |
| miR-185-5p | 0.324 | 0.019 | yes 2,3 | [28,29] | 0.405 | 0.003 | – | – | 0.342 | 0.012 | – | – |
| miR-223-3p | 0.300 | 0.031 | yes 2 | [30] | 0.357 | 0.008 | – | – | 0.335 | 0.014 | – | – |
| miR-3529-3p | 0.330 | 0.016 | yes 1 | – | 0.358 | 0.008 | yes 1 | – | 0.297 | 0.032 | – | – |
| miR-34c-5p | 0.293 | 0.035 | – | – | 0.366 | 0.006 | yes 3 | – | 0.301 | 0.031 | – | – |
| miR-142-3p | 0.350 | 0.010 | – | – | 0.285 | 0.041 | yes 2,3 | [31] | 0.292 | 0.035 | – | – |
| miR-149-5p | 0.273 | 0.052 | – | – | 0.296 | 0.032 | yes 1,2 | – | 0.316 | 0.022 | yes 1,2 | – |
| miR-222-3p | 0.355 | 0.008 | – | – | 0.417 | 0.002 | yes 3 | – | 0.410 | 0.002 | yes 1,2 | [15] |
| miR-370-3p | 0.254 | 0.075 | – | – | 0.347 | 0.010 | yes 1 | – | 0.280 | 0.047 | yes 1,2 | [35,36] |
| miR-378a-3p | 0.270 | 0.054 | – | – | 0.284 | 0.043 | yes 1 | [32,33] | 0.235 | 0.104 | – | – |
| miR-664a-5p | 0.270 | 0.055 | – | – | 0.282 | 0.044 | yes 1 | – | 0.241 | 0.094 | – | – |
| miR-1301-3p | 0.230 | 0.114 | – | – | 0.340 | 0.013 | yes 2 | – | 0.271 | 0.054 | – | – |
| miR-221-3p | 0.240 | 0.095 | – | – | 0.337 | 0.014 | – | – | 0.332 | 0.016 | yes 1,2 | [37] |
| miRNA | Tumor Shrinkage | PFS | OS | |||
|---|---|---|---|---|---|---|
| rho | p-Value | HR (95% CI) | p-Value | HR (95% CI) | p-Value | |
| miR-21-3p | 0.127 | 0.260 | 1.25 (1.05–1.49) | 0.025 | 1.37 (1.14–1.63) | 0.002 |
| miR-34c-5p | 0.356 | 0.003 | 1.20 (1.06–1.36) | 0.013 | 1.26 (1.11–1.44) | 0.002 |
| miR-142-3p | 0.153 | 0.198 | 1.08 (0.91–1.27) | 0.431 | 1.12 (0.95–1.32) | 0.197 |
| miR-142-5p | 0.147 | 0.198 | 1.03 (0.85–1.24) | 0.798 | 1.03 (0.85–1.25) | 0.770 |
| miR-149-5p | 0.151 | 0.198 | 1.44 (1.13–1.83) | 0.012 | 1.57 (1.23–2.00) | 0.002 |
| miR-185-5p | 0.277 | 0.022 | 1.70 (1.33–2.17) | <0.001 | 1.77 (1.37–2.27) | <0.001 |
| miR-221-3p | 0.254 | 0.028 | 1.58 (1.21–2.05) | 0.006 | 1.36 (1.07–1.74) | 0.025 |
| miR-222-3p | 0.233 | 0.043 | 1.31 (1.04–1.66) | 0.035 | 1.28 (1.03–1.60) | 0.053 |
| miR-223-3p | 0.334 | 0.004 | 1.28 (1.04–1.58) | 0.035 | 1.33 (1.07–1.65) | 0.025 |
| miR-370-3p | −0.005 | 0.963 | 1.14 (1.01–1.30) | 0.055 | 1.17 (1.03–1.33) | 0.038 |
| miR-378a-3p | 0.034 | 0.789 | 1.15 (0.92–1.44) | 0.293 | 1.22 (0.98–1.53) | 0.106 |
| miR-664a-5p | 0.107 | 0.330 | 1.14 (0.89–1.47) | 0.357 | 1.19 (0.93–1.53) | 0.183 |
| miR-1301-3p | 0.181 | 0.138 | 1.42 (1.08–1.86) | 0.025 | 1.45 (1.10–1.91) | 0.025 |
| miR-3529-3p | 0.255 | 0.028 | 1.38 (1.13–1.69) | 0.007 | 1.44 (1.18–1.76) | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinget, L.; Roussel, E.; Verbiest, A.; Albersen, M.; Rodríguez-Antona, C.; Graña-Castro, O.; Inglada-Pérez, L.; Zucman-Rossi, J.; Couchy, G.; Job, S.; et al. MicroRNAs Targeting HIF-2α, VEGFR1 and/or VEGFR2 as Potential Predictive Biomarkers for VEGFR Tyrosine Kinase and HIF-2α Inhibitors in Metastatic Clear-Cell Renal Cell Carcinoma. Cancers 2021, 13, 3099. https://doi.org/10.3390/cancers13123099
Kinget L, Roussel E, Verbiest A, Albersen M, Rodríguez-Antona C, Graña-Castro O, Inglada-Pérez L, Zucman-Rossi J, Couchy G, Job S, et al. MicroRNAs Targeting HIF-2α, VEGFR1 and/or VEGFR2 as Potential Predictive Biomarkers for VEGFR Tyrosine Kinase and HIF-2α Inhibitors in Metastatic Clear-Cell Renal Cell Carcinoma. Cancers. 2021; 13(12):3099. https://doi.org/10.3390/cancers13123099
Chicago/Turabian StyleKinget, Lisa, Eduard Roussel, Annelies Verbiest, Maarten Albersen, Cristina Rodríguez-Antona, Osvaldo Graña-Castro, Lucía Inglada-Pérez, Jessica Zucman-Rossi, Gabrielle Couchy, Sylvie Job, and et al. 2021. "MicroRNAs Targeting HIF-2α, VEGFR1 and/or VEGFR2 as Potential Predictive Biomarkers for VEGFR Tyrosine Kinase and HIF-2α Inhibitors in Metastatic Clear-Cell Renal Cell Carcinoma" Cancers 13, no. 12: 3099. https://doi.org/10.3390/cancers13123099