The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response

 ,
,
Abstract
:1. Introduction
2. Gut Microbiota and CRC
2.1. Gut Microbiota Composition in CRC
2.2. Gut Microbiota Dysbiosis, Inflammation and CRC
2.3. Pathogenic Bacteria and CRC
3. Gut Microbiota-Derived Metabolites and CRC
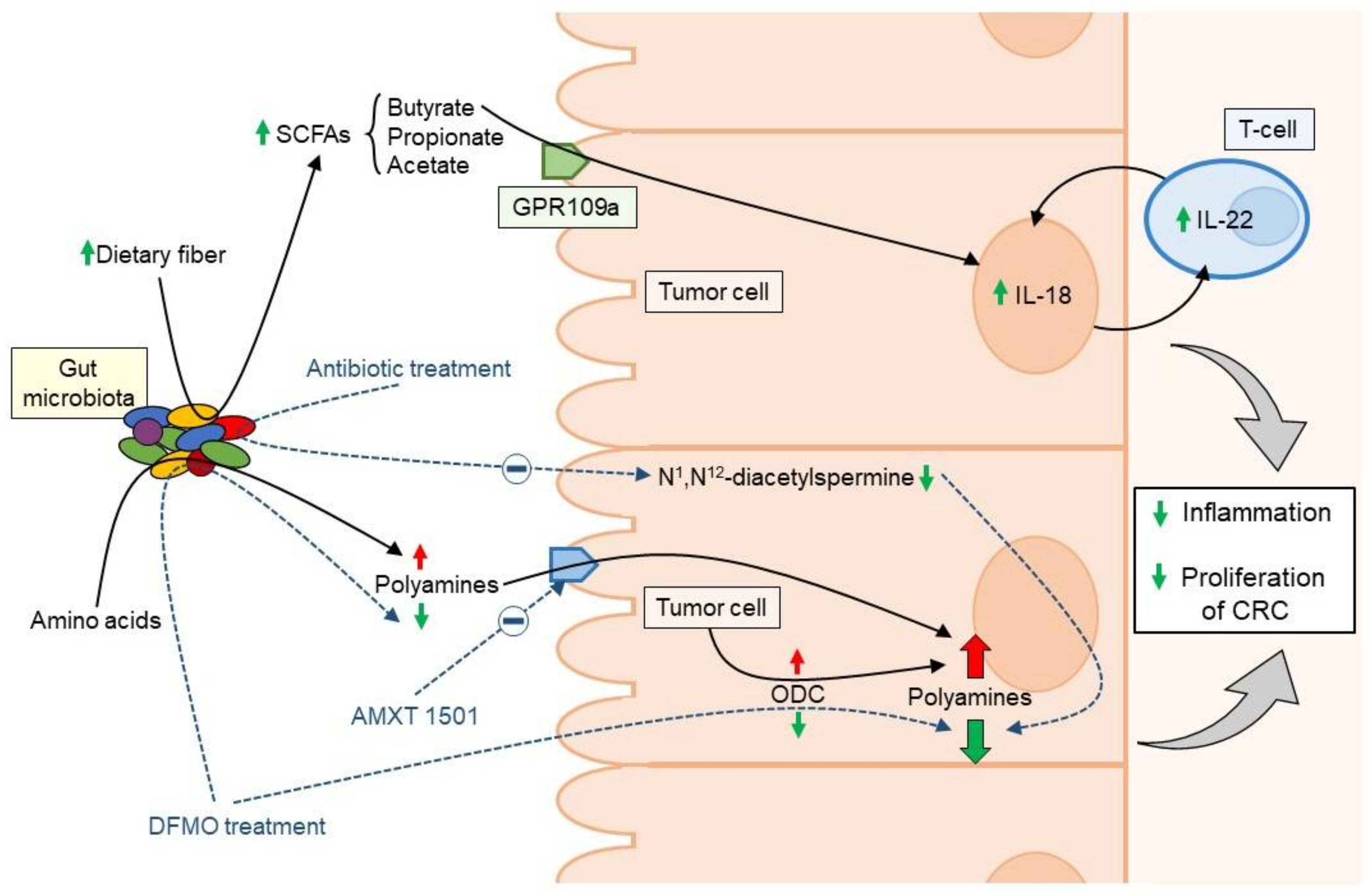
3.1. Gut Microbiota-Derived Polyamines and CRC
3.2. Short Chain Fatty Acid Metabolism and CRC
4. Antibiotic-Microbiome Link and CRC Risk
5. Dietary Mediators of CRC Carcinogenesis
5.1. Dietary Fiber
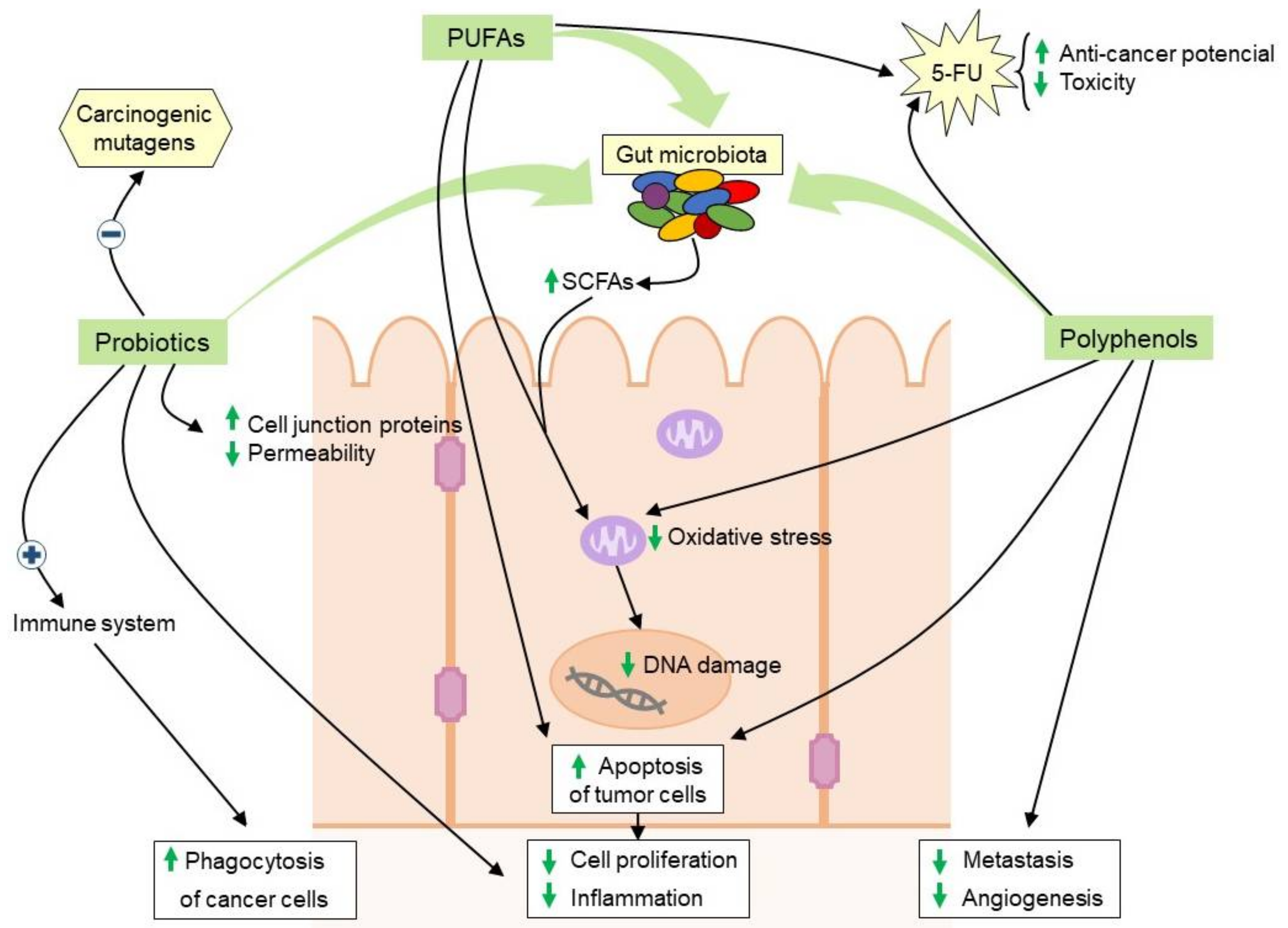
5.2. Diets Rich in Polyunsaturated Fatty Acids
5.3. Bioactive Polyphenols
5.4. Probiotics
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, A.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.K.; Kelly, S.A.; Legge, R.; Ma, F.; Low, S.J.; Kim, J.; Zhang, M.; Oh, P.L.; Nehrenberg, D.; Hua, K.; et al. Individuality in gut microbiota composition is a complex polygenic trait shaped by multiple environmental and host genetic factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18933–18938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial Community Variation in Human Body Habitats Across Space and Time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef]
- Bhat, M.I.; Kapila, R. Dietary metabolites derived from gut microbiota: Critical modulators of epigenetic changes in mammals. Nutr. Rev. 2017, 75, 374–389. [Google Scholar] [CrossRef]
- Jahani-Sherafat, S.; Alebouyeh, M.; Moghim, S.; Amoli, H.A.; Safaei, H.G. Role of gut microbiota in the pathogenesis of colorectal cancer; a review article. Gastroenterol. Hepatol. Bed Bench 2018, 11, 101–109. [Google Scholar]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, N.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in Globocan 2012. Int. J. Cancer 2014, 136, 359–386. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma–carcinoma sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsu, G.; Li, X.; Zhou, H.; Sheng, J.; Wong, S.H.; Wu, W.K.; Ng, S.C.; Tsoi, H.; Dong, Y.; Zhang, N.; et al. Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat. Commun. 2015, 6, 8727. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, J.Q.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2015, 66, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.J.M.; Collins, P.D. Colon Cancer: A Civilization Disorder. Dig. Dis. 2011, 29, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Cipe, G.; Idiz, U.O.; Firat, D.; Bektasoglu, H. Relationship between intestinal microbiota and colorectal cancer. World J. Gastrointest. Oncol. 2015, 7, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Zackular, J.P.; Baxter, N.; Iverson, K.D.; Sadler, W.D.; Petrosino, J.F.; Chen, G.Y.; Schloss, P.D. The Gut Microbiome Modulates Colon Tumorigenesis. mBio 2013, 4, 692–713. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef]
- Saffarian, A.; Mulet, C.; Regnault, B.; Amiot, A.; Tran-Van-Nhieu, J.; Ravel, J.; Sobhani, I.; Sansonetti, P.J.; Pédron, T.; Barnich, N.; et al. Crypt- and Mucosa-Associated Core Microbiotas in Humans and Their Alteration in Colon Cancer Patients. mBio 2019, 10, 1315–1319. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2011, 6, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.J.; Rawls, J.F.; Randall, T.A.; Burcal, L.; Mpande, C.N.; Jenkins, N.; Jovov, B.; Abdo, Z.; Sandler, R.S.; Keku, T.O. Molecular characterization of mucosal adherent bacteria and associations with colorectal adenomas. Gut Microbes 2010, 1, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Wu, W.; Qin, H. Probiotics modify human intestinal mucosa-associated microbiota in patients with colorectal cancer. Mol. Med. Rep. 2015, 12, 6119–6127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.; Sinha, R.; Pei, Z.; Dominianni, C.; Wu, J.; Shi, J.; Goedert, J.J.; Hayes, R.B.; Yang, L. Human gut microbiome and risk for colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 1907–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viljoen, K.S.; Dakshinamurthy, A.; Goldberg, P.; Blackburn, J.M. Quantitative Profiling of Colorectal Cancer-Associated Bacteria Reveals Associations between Fusobacterium spp., Enterotoxigenic Bacteroides fragilis (ETBF) and Clinicopathological Features of Colorectal Cancer. PLoS ONE 2015, 10, e0119462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial controlador–passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Genet. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.N.; Araujo-Perez, F.; Azcarate-Peril, M.A.; Yeh, J.J.; Sandler, R.S.; Keku, T.O. Fusobacterium Is Associated with Colorectal Adenomas. PLoS ONE 2013, 8, e53653. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Dominianni, C.; Shapiro, J.A.; Church, T.R.; Wu, J.; Miller, G.; Yuen, E.; Freiman, H.; Lustbader, I.; Salik, J.; et al. The gut microbiota in conventional and serrated precursors of colorectal cancer. Microbiome 2016, 4, 69. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e16. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kundu, P.; Seow, S.W.; de Matos, C.T.; Aronsson, L.; Chin, K.C.; Kärre, K.; Pettersson, S.; Greicius, G. Gut microbiota accelerate tumor growth via c-jun and STAT3 phosphorylation in APC Min/+ mice. Carcinogenesis 2012, 33, 1231–1238. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K.; Clausen, B.E.; Kaisho, T.; Tsujimura, T.; Terada, N.; Förster, I.; Akira, S. Enhanced Th1 activity and development of chronic enterocolitis in mice devoid of Stat3 in macrophages and neutrophils. Immunity 1999, 10, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Uronis, J.M.; Mühlbauer, M.; Herfarth, H.H.; Rubinas, T.C.; Jones, G.; Jobin, C. Modulation of the Intestinal Microbiota Alters Colitis-Associated Colorectal Cancer Susceptibility. PLoS ONE 2009, 4, e6026. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.-Y.; Österreicher, C.H.; Hung, K.E.; et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 2012, 491, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostovcikova, K.; Kverka, M.; Zakostelska, Z.; Hudcovic, T.; Hrncir, T.; Stepankova, R.; Rossmann, P.; Ridl, J.; Kostovcikova, K.; Mrázek, J.; et al. Altered gut microbiota promotes colitis-associated cancer in IL-1 receptor-associated kinase M-deficient mice. Inflamm. Bowel Dis. 2013, 19, 1266–1277. [Google Scholar] [CrossRef] [Green Version]
- Coelho, T.R.; Almeida, L.; Lazo, P.A. JC virus in the pathogenesis of colorectal cancer, an etiological agent or another component in a multistep process? Virol. J. 2010, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damin, D.C.; Ziegelmann, P.K.; Damin, A.P. Human papillomavirus infection and colorectal cancer risk: A meta-analysis. Color. Dis. 2013, 15, 420. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, G.; Zhou, H.; Wu, W.K.K.; Wong, S.H.; Coker, O.O.; Dai, Z.; Li, X.; Szeto, C.H.; Sugimura, N.; Lam, T.Y.-T.; et al. Alterations in Enteric Virome Are Associated With Colorectal Cancer and Survival Outcomes. Gastroenterology 2018, 155, 529–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannigan, G.; Duhaime, M.B.; Ruffin, M.; Koumpouras, C.C.; Schloss, P.D. Diagnostic Potential and Interactive Dynamics of the Colorectal Cancer Virome. mBio 2018, 9, 2248–2318. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.-W.; Dong, X.; Pan, P.; Chen, K.-W.; Fan, J.-X.; Cheng, S.-X.; Zhang, X.-Z. Phage-guided modulation of the gut microbiota of mouse models of colorectal cancer augments their responses to chemotherapy. Nat. Biomed. Eng. 2019, 3, 717–728. [Google Scholar] [CrossRef]
- Luan, C.; Xie, L.; Yang, X.; Miao, H.; Lv, N.; Zhang, R.; Xiao, X.; Hu, Y.; Liu, Y.; Wu, N.; et al. Dysbiosis of Fungal Microbiota in the Intestinal Mucosa of Patients with Colorectal Adenomas. Sci. Rep. 2015, 5, 7980. [Google Scholar] [CrossRef]
- Coker, O.O.; Nakatsu, G.; Dai, R.Z.; Wu, W.K.K.; Wong, S.H.; Ng, S.C.; Chan, F.K.L.; Sung, J.J.; Yu, J. Enteric fungal microbiota dysbiosis and ecological alterations in colorectal cancer. Gut 2018, 68, 654–662. [Google Scholar] [CrossRef]
- Gao, R.; Kong, C.; Li, H.; Huang, L.; Qu, X.; Qin, N.; Qin, H. Dysbiosis signature of mycobiota in colon polyp and colorectal cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2457–2468. [Google Scholar] [CrossRef]
- Grivennikov, S. Inflammation and colorectal cancer: Colitis-associated neoplasia. Semin. Immunopathol. 2012, 35, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Francescone, R.; Hou, V.; Grivennikov, S. Microbiome, inflammation, and cancer. Cancer J. 2014, 20, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzutsev, A.; Goldszmid, R.S.; Viaud, S.; Zitvogel, L.; Trinchieri, G. The role of the microbiota in inflammation, carcinogenesis, and cancer therapy. Eur. J. Immunol. 2014, 45, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Bongers, G.; Pacer, M.E.; Geraldino, T.H.; Chen, L.; He, Z.; Hashimoto, D.; Furtado, G.C.; Ochando, J.; Kelley, K.A.; Clemente, J.C.; et al. Interplay of host microbiota, genetic perturbations, and inflammation promotes local development of intestinal neoplasms in mice. J. Exp. Med. 2014, 211, 457–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Elinav, E.; Huber, S.; Strowig, T.; Hao, L.; Hafemann, A.; Jin, C.; Wunderlich, C.; Wunderlich, T.; Eisenbarth, S.C.; et al. Microbiota-induced activation of epithelial IL-6 signaling links inflammasome-driven inflammation with transmissible cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 9862–9867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, W.E.; Moore, L.H. Intestinal floras of populations that have a high risk of CRC cancer. Appl. Environ. Microbiol. 1995, 61, 3202–3207. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, A.C.; Shields, C.E.D.; Wu, S.; Huso, D.L.; Wu, X.; Stewart, T.M.; Hacker-Prietz, A.; Rabizadeh, S.; Woster, P.M.; Sears, C.L.; et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15354–15359. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Shin, J.; Zhang, G.; Cohen, M.B.; Franco, A.; Sears, C. The Bacteroides fragilis Toxin Binds to a Specific Intestinal Epithelial Cell Receptor. Infect. Immun. 2006, 74, 5382–5390. [Google Scholar] [CrossRef] [Green Version]
- Vizcaino, M.I.; Crawford, J. The colibactin warhead crosslinks DNA. Nat. Chem. 2015, 7, 411–417. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/β-catenin modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef]
- Brennan, C.A.; Garrett, W.S. Fusobacterium nucleatum—symbiont, opportunist and oncobacterium. Nat. Rev. Genet. 2018, 17, 156–166. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Genet. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.M.; Manghi, P.; Asnicar, F.; Pasolli, E.; Armanini, F.; Zolfo, M.; Beghini, F.; Manara, S.; Karcher, N.; Pozzi, C.; et al. Metagenomic analysis of colorectal cancer datasets identifies cross-cohort microbial diagnostic signatures and a enlace with choline degradation. Nat. Med. 2019, 25, 667–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soda, K. The mechanisms by which polyamines accelerate tumor spread. J. Exp. Clin. Cancer Res. 2011, 30, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casero, R.A.; Stewart, T.M.; Pegg, A.E. Polyamine metabolism and cancer: Treatments, challenges and opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Love, R.R.; Surawicz, T.S.; Morrissey, J.F.; Verma, A.K. Levels of colorectal ornithine decarboxylase activity in patients with colon cancer, a family history of nonpolyposis hereditary colorectal cancer, and adenomas. Cancer Epidemiol. Biomark. Prev. 1992, 1, 195–198. [Google Scholar]
- Linsalata, M.; Russo, F.; Cavallini, A.; Berloco, P.; di Leo, A. Polyamines, diamine oxidase, and ornithine decarboxylase activity in colorectal cancer and in normal surrounding mucosa. Dis. Colon Rectum 1993, 36, 662–667. [Google Scholar] [CrossRef]
- Meyskens, F.L.; McLaren, C.E.; Pelot, D.; Fujikawa-Brooks, S.; Carpenter, P.M.; Hawk, E.; Kelloff, G.; Lawson, M.J.; Kidao, J.; McCracken, J.; et al. Difluoromethylornithine Plus Sulindac for the Prevention of Sporadic Colorectal Adenomas: A Randomized Placebo-Controlled, Double-Blind Trial. Cancer Prev. Res. 2008, 1, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Raj, K.P.; Zell, J.A.; Rock, C.L.; McLaren, C.; Zoumas-Morse, C.; Gerner, E.W.; Meyskens, F.L. Role of dietary polyamines in a phase III clinical trial of difluoromethylornithine (DFMO) and sulindac for prevention of sporadic colorectal adenomas. Br. J. Cancer 2013, 108, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Molina, B.; Queipo-Ortuño, M.I.; Lambertos, A.; Tinahones, F.J.; Peñafiel, R. Dietary and Gut Microbiota Polyamines in Obesity- and Age-Related Diseases. Front. Nutr. 2019, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Cocchi, S.; Suzzi, G. Polyamines and Gut Microbiota. Front. Nutr. 2019, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessels, J.; Kingma, A.W.; Ferwerda, H.; Keu, J.; Berg, G.A.V.D.; Muskiet, F.A.J. Microbial flora in the gastrointestinal tract abolishes cytostatic effects of α-difluoromethylornithinein vivo. Int. J. Cancer 1989, 43, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Quemener, V.; Blanchard, Y.; Chamaillard, L.; Havouis, R.; Cipolla, B.; Moulinoux, J.P. Polyamine deprivation: A new tool in cancer treatment. Anticancer Res. 1994, 14, 443–448. [Google Scholar] [PubMed]
- Gamble, L.D.; Purgato, S.; Murray, J.; Xiao, L.; Yu, D.M.T.; Hanssen, K.M.; Giorgi, F.M.; Carter, D.R.; Gifford, A.J.; Valli, E.; et al. Inhibition of polyamine synthesis and uptake reduces tumor progression and prolongs survival in mouse models of neuroblastoma. Sci. Transl. Med. 2019, 11, e1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejea, C.M.; Wick, E.C.; Hechenbleikner, E.M.; White, J.R.; Welch, J.L.M.; Rossetti, B.J.; Peterson, S.N.; Snesrud, E.C.; Borisy, G.G.; Lazarev, M.; et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 18321–18326. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.H.; Dejea, C.M.; Edler, D.; Hoang, L.T.; Santidrian, A.F.; Felding, B.H.; Ivanisevic, J.; Cho, K.; Wick, E.C.; Hechenbleikner, E.M.; et al. Metabolism links bacterial biofilms and colon carcinogenesis. Cell Metab. 2015, 21, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Yang, X.; Zhang, R.; Li, J.; Xiao, X.; Hu, Y.; Chen, Y.; Yang, F.; Lu, N.; Wang, Z.; et al. Dysbiosis Signature of Fecal Microbiota in Colorectal Cancer Patients. Microb. Ecol. 2013, 66, 462–470. [Google Scholar] [CrossRef]
- Brown, D.; Rao, S.; Weir, T.L.; O’Malia, J.; Bazan, M.; Brown, R.J.; Ryan, E.P. Metabolomics and metabolic pathway networks from human colorectal cancers, adjacent mucosa, and stool. Cancer Metab. 2016, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, R.; Worschech, A.; Cardone, M.; Jones, Y.; Gyulai, Z.; Dai, R.-M.; Wang, E.; Ma, W.; Haines, D.; O’Huigin, C.; et al. MyD88-mediated signaling prevents development of adenocarcinomas of the colon: Role of interleukin 18. J. Exp. Med. 2010, 207, 1625–1636. [Google Scholar] [CrossRef]
- Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interfaz. Proc. Natl. Acad. Sci. USA 2008, 105, 20858–20863. [Google Scholar] [CrossRef] [Green Version]
- Dupaul-Chicoine, J.; Yeretssian, G.; Doiron, K.; Bergstrom, K.S.; McIntire, C.R.; Leblanc, P.M.; Meunier, C.; Turbide, C.; Gros, P.; Beauchemin, N.; et al. Control of Intestinal Homeostasis, Colitis, and Colitis-Associated Colorectal Cancer by the Inflammatory Caspases. Immunity 2010, 32, 367–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, I.C.; TeKippe, E.M.; Woodford, R.-M.T.; Uronis, J.M.; Holl, E.K.; Rogers, A.B.; Herfarth, H.H.; Jobin, C.; Ting, J.P.-Y. The NLRP3 inflammasome functions as a negative regulator of tumorigenesis during colitis-associated cancer. J. Exp. Med. 2010, 207, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Elinav, E.; Strowig, T.; Kau, A.; Henao-Mejia, J.; Thaiss, C.A.; Booth, C.J.; Peaper, D.R.; Bertin, J.; Eisenbarth, S.; Gordon, J.I.; et al. NLRP6 Inflammasome Regulates Colonic Microbial Ecology and Risk for Colitis. Cell 2011, 145, 745–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Martin, O.C.B.; Lin, C.; Naud, N.; Tache, S.; Raymond-Letron, I.; Corpet, D.E.; Pierre, F.H. Antibiotic Suppression of Intestinal Microbiota Reduces Heme-Induced Lipoperoxidation Associated with Colon Carcinogenesis in Rats. Nutr. Cancer 2014, 67, 119–125. [Google Scholar] [CrossRef]
- Dik, V.K.; van Oijen, M.G.H.; Smeets, H.M.; Siersema, P.D. Frequent Use of Antibiotics Is Associated with Colorectal Cancer Risk: Results of a Nested Case-Control Study. Dig. Dis. Sci. 2015, 61, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Boursi, B.; Haynes, K.; Mamtani, R.; Yang, Y.-X. Impact of antibiotic exposure on the risk of colorectal cancer. Pharmacoepidemiol. Drug Saf. 2015, 24, 534–542. [Google Scholar] [CrossRef]
- Zackular, J.P.; Baxter, N.T.; Chen, G.Y.; Schloss, P.D. Manipulation of the Gut Microbiota Reveals Role in Colon Tumorigenesis. mSphere 2015, 1, e00001-15. [Google Scholar] [CrossRef] [Green Version]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.S.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.G.; Eun, C.S.; Jo, S.V.; Lee, A.-R.; Park, C.H.; Han, D.S. The impact of gut microbiota manipulation with antibiotics on colon tumorigenesis in a murine model. PLoS ONE 2019, 14, e0226907. [Google Scholar] [CrossRef]
- Ma, W.; Chan, A.T. Antibiotic use and colorectal cancer: A causal association? Gut 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Chang, C.-H.; Lin, J.-W.; Wu, L.-C.; Chuang, L.-M.; Lai, M.-S. Infection, antibiotic therapy and risk of colorectal cancer: A nationwide nested case-control study in patients with Type 2 diabetes mellitus. Int. J. Cancer 2014, 135, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Wu, K.; Mehta, R.; Drew, D.; Song, M.; Lochhead, P.; Nguyen, L.H.; Izard, J.; Fuchs, C.S.; Garrett, W.S.; et al. Long-term use of antibiotics and risk of colorectal adenoma. Gut 2017, 67, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Montalban-Arques, A.; Scharl, M. Intestinal microbiota and colorectal carcinoma: Implications for pathogenesis, diagnosis, and therapy. eBioMedicine 2019, 48, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Haines, C.; Watson, A.J.M.; Hart, A.R.; Platt, M.J.; Pardoll, D.M.; E Cosgrove, S.; A Gebo, K.; Sears, C.L. Oral antibiotic use and risk of colorectal cancer in the United Kingdom, 1989–2012: A matched case–control study. Gut 2019, 68, 1971–1978. [Google Scholar] [CrossRef]
- Sanyaolu, L.N.; Oakley, N.J.; Nurmatov, U.; Dolwani, S.; Ahmed, H. Antibiotic exposure and the risk of colorectal adenoma and carcinoma: A systematic review and meta-analysis of observational studies. Color. Dis. 2019. [Google Scholar] [CrossRef]
- Armstrong, D.; Dregan, A.; Ashworth, M.; White, P.; McGee, C.; de Lusignan, S. The association between colorectal cancer and prior antibiotic prescriptions: Case control study. Br. J. Cancer 2020, 122, 912–917. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, S.; Li, H.; Yang, F.; Mushtaq, N.; Ullah, S.; Shi, Y.; An, C.; Xu, J. The influence of gut microbiota dysbiosis to the efficacy of 5-Fluorouracil treatment on colorectal cancer. Biomed. Pharmacother. 2018, 108, 184–193. [Google Scholar] [CrossRef]
- Abdel-Rahman, O.; Ghosh, S.; Walker, J. Outcomes of metastatic colorectal cancer patients in relationship to prior and concurrent antibiotics use; individual patient data analysis of three clinical trials. Clin. Transl. Oncol. 2020, 1–6. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leystra, A.; Clapper, M.L. Gut Microbiota Influences Experimental Outcomes in Mouse Models of Colorectal Cancer. Genes 2019, 10, 900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattimer, J.M.; Haub, M. Effects of Dietary Fiber and Its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Lazarova, D.L.; Bordonaro, M. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J. Gastrointest. Oncol. 2014, 6, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Bingham, S.; Day, N.; Luben, R.N.; Ferrari, P.; Slimani, N.; Norat, T.; Clavel-Chapelon, F.; Kesse, E.; Nieters, A.; Boeing, H.; et al. Dietary fibre in food and protection against colorectal cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC): An observational study. Lancet 2003, 361, 1496–1501. [Google Scholar] [CrossRef]
- Schatzkin, A.; Mouw, T.; Park, Y.; Subar, A.F.; Kipnis, V.; Hollenbeck, A.; Leitzmann, M.F.; E Thompson, F. Dietary fiber and whole-grain consumption in relation to colorectal cancer in the NIH-AARP Diet and Health Study. Am. J. Clin. Nutr. 2007, 85, 1353–1360. [Google Scholar] [CrossRef] [Green Version]
- Dahm, C.C.; Keogh, R.H.; A Spencer, E.; Greenwood, D.; Key, T.J.; Fentiman, I.S.; Shipley, M.J.; Brunner, E.J.; Cade, J.E.; Burley, V.; et al. Dietary Fiber and Colorectal Cancer Risk: A Nested Case-Control Study Using Food Diaries. J. Natl. Cancer Inst. 2010, 102, 614–626. [Google Scholar] [CrossRef] [Green Version]
- Dahm, C.C. Dietary fiber and colorectal cancer risk: A nested case-control study using food diaries. J. Natl. Cancer Inst. 2011, 103, 1484. [Google Scholar] [CrossRef] [Green Version]
- Hansen, L.; Skeie, G.; Landberg, R.; Lund, E.; Palmqvist, R.; Johansson, I.; Dragsted, L.O.; Egeberg, R.; Johnsen, N.F.; Christensen, J.; et al. Intake of dietary fiber, especially from cereal foods, is associated with lower incidence of colon cancer in the HELGA cohort. Int. J. Cancer 2011, 131, 469–478. [Google Scholar] [CrossRef]
- Song, M.; Wu, K.; Meyerhardt, J.A.; Ogino, S.; Wang, M.; Fuchs, C.S.; Giovannucci, E.L.; Chan, A.T. Fiber Intake and Survival After Colorectal Cancer Diagnosis. JAMA Oncol. 2018, 4, 71–79. [Google Scholar] [CrossRef]
- Moen, B.; Henjum, K.; Mage, I.; Knutsen, S.H.; Rud, I.; Hetland, R.B.; Paulsen, J.E. Effect of Dietary Fibers on Cecal Microbiota and Intestinal Tumorigenesis in Azoxymethane Treated A/J Min/+ Mice. PLoS ONE 2016, 11, e0155402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, R.S.; Nishihara, R.; Cao, Y.; Song, M.; Mima, K.; Qian, Z.R.; Nowak, J.A.; Kosumi, K.; Hamada, T.; Masugi, Y.; et al. Association of Dietary Patterns With Risk of Colorectal Cancer Subtypes Classified by Fusobacterium nucleatum in Tumor Tissue. JAMA Oncol. 2017, 3, 921–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deehan, E.C.; Yang, C.; Perez-Muñoz, M.E.; Nguyen, N.K.; Cheng, C.C.; Triador, L.; Zhang, Z.; Bakal, J.A.; Walter, J. Precision Microbiome Modulation with Discrete Dietary Fiber Structures Directs Short-Chain Fatty Acid Production. Cell Host Microbe 2020, 27, 389–404. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-M.; Yu, Y.-N.; Wang, J.-L.; Lin, Y.-W.; Kong, X.; Yang, C.-Q.; Yang, L.; Liu, Z.; Yuan, Y.-Z.; Liu, F.; et al. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keefe, S.J.D.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.M.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 6342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohoe, D.R.; Holley, D.; Collins, L.B.; Montgomery, S.A.; Whitmore, A.C.; Hillhouse, A.; Curry, K.P.; Renner, S.W.; Greenwalt, A.; Ryan, E.P.; et al. A gnotobiotic mouse model demonstrates that dietary fiber protects against colorectal tumorigenesis in a microbiota- and butyrate-dependent manner. Cancer Discov. 2014, 4, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Bishehsari, F.; Engen, P.; Preite, N.Z.; Tuncil, Y.E.; Naqib, A.; Shaikh, M.; Rossi, M.; Wilber, S.; Green, S.J.; Hamaker, B.R.; et al. Dietary Fiber Treatment Corrects the Composition of Gut Microbiota, Promotes SCFA Production, and Suppresses Colon Carcinogenesis. Genes 2018, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Burkitt, D.P. Epidemiology of cancer of the colon and rectum. Cancer 1971, 28, 3–13. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2007, 27, 104–119. [Google Scholar] [CrossRef]
- Fung, T.T.; Hu, F.B.; Schulze, M.B.; Pollak, M.; Wu, T.; Fuchs, C.S.; Giovannucci, E. A dietary pattern that is associated with C-peptide and risk of colorectal cancer in women. Cancer Causes Control. 2012, 23, 959–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2008, 136, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpato, M.; A Hull, M. Omega-3 polyunsaturated fatty acids as adjuvant therapy of colorectal cancer. Cancer Metastasis Rev. 2018, 37, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [Green Version]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; A Spencer, J.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2017, 67, 1974–1983. [Google Scholar] [CrossRef]
- Cho, Y.; Turner, N.D.; A Davidson, L.; Chapkin, R.S.; Carroll, R.J.; Lupton, J.R. Colon cancer cell apoptosis is induced by combined exposure to the n-3 fatty acid docosahexaenoic acid and butyrate through promoter methylation. Exp. Boil. Med. 2014, 239, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Chapkin, R.S.; Declercq, V.; Kim, E.; Fuentes, N.; Fan, Y.-Y. Mechanisms by Which Pleiotropic Amphiphilic n−3 PUFA Reduce Colon Cancer Risk. Curr. Color. Cancer Rep. 2014, 10, 442–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triff, K.; Kim, E.; Chapkin, R.S. Chemoprotective Epigenetic Mechanisms in a Colorectal Cancer Model: Modulation by n-3 PUFA in Combination With Fermentable Fiber. Curr. Pharmacol. Rep. 2015, 1, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.Y.; Turner, N.D.; Murphy, M.E.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R. In vivo regulation of colonic cell proliferation, differentiation, apoptosis, and P27Kip1 by dietary fish oil and butyrate in rats. Cancer Prev. Res. 2015, 8, 1076–1083. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, G.; D’Argenio, G.; Prossomariti, A.; Lembo, V.; Mazzone, G.; Candela, M.; Biagi, E.; Brigidi, P.; Vitaglione, P.; Fogliano, V.; et al. Eicosapentaenoic acid free fatty acid prevents and suppresses colonic neoplasia in colitis-associated colorectal cancer acting on Notch signaling and gut microbiota. Int. J. Cancer 2014, 135, 2004–2013. [Google Scholar] [CrossRef]
- Lee, J.Y.; Sim, T.-B.; Lee, J.-E.; Na, H.-K. Chemopreventive and Chemotherapeutic Effects of Fish Oil derived Omega-3 Polyunsaturated Fatty Acids on Colon Carcinogenesis. Clin. Nutr. Res. 2017, 6, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Nishihara, R.; Wu, K.; Qian, Z.R.; Kim, S.A.; Sukawa, Y.; Mima, K.; Inamura, K.; Masuda, A.; Yang, J.; et al. Marine ω-3 Polyunsaturated Fatty Acids and Risk for Colorectal Cancer According to Microsatellite Instability. J. Natl. Cancer Inst. 2015, 107, e007. [Google Scholar] [CrossRef] [Green Version]
- Lamprecht, S.A.; Lipkin, M. Chemoprevention of colon cancer by calcium, vitamin D and folate: Molecular mechanisms. Nat. Rev. Cancer 2003, 3, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Li, H.; Dong, J.; Dong, Y.; Wang, C.-Z. Expression profile of polyunsaturated fatty acids in colorectal cancer. World J. Gastroenterol. 2015, 21, 2405–2412. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-C.L.; Chapkin, R.S.; Lupton, J.R. Fish Oil Blocks Azoxymethane-Induced Rat Colon Tumorigenesis by Increasing Cell Differentiation and Apoptosis Rather Than Decreasing Cell Proliferation. J. Nutr. 1998, 128, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Zhang, X.; Meyerhardt, J.A.; Giovannucci, E.L.; Ogino, S.; Fuchs, C.S.; Chan, A.T. Marine ω-3 polyunsaturated fatty acid intake and survival after colorectal cancer diagnosis. Gut 2016, 66, 1790–1796. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Turner, N.D.; Davidson, L.; Chapkin, R.S.; Carroll, R.J.; Lupton, J.R. A chemoprotective fish oil/pectin diet enhances apoptosis via Bcl-2 promoter methylation in rat azoxymethane-induced carcinomas. Exp. Boil. Med. 2012, 237, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- Ng, Y.; Barhoumi, R.; Tjalkens, R.B.; Fan, Y.-Y.; Kolar, S.; Wang, N.; Lupton, J.R.; Chapkin, R.S. The role of docosahexaenoic acid in mediating mitochondrial membrane lipid oxidation and apoptosis in colonocytes. Carcinogenesis 2005, 26, 1914–1921. [Google Scholar] [CrossRef] [Green Version]
- Sofi, F.; Dinu, M.; Pagliai, G.; Pierre, F.; Gueraud, F.; Bowman, J.; Gerard, P.; Longo, V.; Giovannelli, L.; Caderni, G.; et al. Fecal microbiome as determinant of the effect of diet on colorectal cancer risk: Comparison of meat-based versus pesco-vegetarian diets (the MeaTIc study). Trials 2019, 20, 688–689. [Google Scholar] [CrossRef]
- Aglago, E.K.; Huybrechts, I.; Murphy, N.; Casagrande, C.; Nicolas, G.; Pischon, T.; Fedirko, V.; Severi, G.; Boutron-Ruault, M.-C.; Fournier, A.; et al. Consumption of Fish and Long-chain n-3 Polyunsaturated Fatty Acids Is Associated With Reduced Risk of Colorectal Cancer in a Large European Cohort. Clin. Gastroenterol. Hepatol. 2020, 18, 654–666. [Google Scholar] [CrossRef]
- Rani, I.; Sharma, B.; Kumar, S.; Kaur, S.; Agnihotri, N. Apoptosis mediated chemosensitization of tumor cells to 5-fluorouracil on supplementation of fish oil in experimental colon carcinoma. Tumor Boil. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, I.; Vaiphei, K.; Agnihotri, N. Supplementation of fish oil augments efficacy and attenuates toxicity of 5-fluorouracil in 1,2-dimethylhydrazine dihydrochloride/dextran sulfate sodium-induced colon carcinogenesis. Cancer Chemother. Pharmacol. 2014, 74, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Sebe, M.; Tsutsumi, R.; Yamaguchi, S.; Horikawa, Y.T.; Harada, N.; Oyama, T.; Kakuta, N.; Tanaka, K.; Tsutsumi, Y.M.; Nakaya, Y.; et al. The synergystic effects of omega-3 fatty acids against 5-fluorouracil-induced mucosal impairment in mice. BMC Nutr. 2016, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Ebadi, M.; Field, C.J.; Lehner, R.; Mazurak, V.C. Chemotherapy diminishes lipid storage capacity of adipose tissue in a preclinical model of colon cancer. Lipids Heal. Dis. 2017, 16, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, F.; Sorg, O.; Granci, V.; Lecumberri, E.; Miralbell, R.; Dupertuis, Y.M.; Pichard, C. Interaction of ω-3 polyunsaturated fatty acids with radiation therapy in two different colorectal cancer cell lines. Clin. Nutr. 2014, 33, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granci, V.; Cai, F.; Lecumberri, E.; Clerc, A.; Dupertuis, Y.M.; Pichard, C. Colon cancer cell chemosensitisation by fish oil emulsion involves apoptotic mitochondria pathway. Br. J. Nutr. 2012, 109, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Golkhalkhali, B.; Rajandram, R.; Paliany, A.S.; Ho, G.F.; Ishak, W.Z.W.; Johari, C.S.; Chin, K.F. Strain-specific probiotic (microbial cell preparation) and omega-3 fatty acid in modulating quality of life and inflammatory markers in colorectal cancer patients: A randomized controlled trial. Asia Pac. J. Clin. Oncol. 2017, 14, 179–191. [Google Scholar] [CrossRef]
- Siriwardhana, N.; Kalupahana, N.S.; Cekanova, M.; Lemieux, M.; Greer, B.; Moustaid-Moussa, N. Modulation of adipose tissue inflammation by bioactive food compounds. J. Nutr. Biochem. 2013, 24, 613–623. [Google Scholar] [CrossRef]
- Mileo, A.M.; Nisticò, P.; Miccadei, S. Polyphenols: Immunomodulatory and Therapeutic Implication in Colorectal Cancer. Front. Immunol. 2019, 10, 729. [Google Scholar] [CrossRef]
- Dykes, L. Phenolic Compounds in Cereal Grains and Their Health Benefits. Cereal Foods World 2007, 52, 105–111. [Google Scholar] [CrossRef]
- Vanamala, J.; Leonardi, T.; Patil, B.S.; Taddeo, S.S.; Murphy, M.E.; Pike, L.M.; Chapkin, R.S.; Lupton, J.R.; Turner, N.D. Suppression of colon carcinogenesis by bioactive compounds in grapefruit. Carcinogenesis 2005, 27, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 2016, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Miene, C.; Klenow, S.; Veeriah, S.; Richling, E.; Glei, M. Impact of apple polyphenols onGSTT2gene expression, subsequent protection of DNA and modulation of proliferation using LT97 human colon adenoma cells. Mol. Nutr. Food Res. 2009, 53, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A. Colonic metabolism of dietary polyphenols: Influence of structure on microbial fermentation products. Free. Radic. Boil. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications inhuman health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueñas, M.; Muñoz-Gonzalez, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A Survey of Modulation of Gut Microbiota by Dietary Polyphenols. BioMed Res. Int. 2015, 2015, 1–15. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, M.D.C.; Ribeiro-Vidal, H.; Esteban-Fernández, A.; Bartolomé, B.; Figuero, E.; Moreno-Arribas, M.V.; Sanz, M.; Herrera, D. Antimicrobial activity of red wine and oenological extracts against periodontal pathogens in a validated oral biofilm model. BMC Complement. Altern. Med. 2019, 19, 145. [Google Scholar] [CrossRef]
- Cueva, C.; Silva, M.; Pinillos, I.; Bartolomé, B.; Moreno-Arribas, M.V. Interplay between Dietary Polyphenols and Oral and Gut Microbiota in the Development of Colorectal Cancer. Nutrients 2020, 12, 625. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; de Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and Cancer Chemoprevention. Evid. Based Complement. Altern. Med. 2011, 2011, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Venâncio, V.P.; Cipriano, P.A.; Kim, H.; Antunes, L.M.G.; Talcott, S.T.; Mertens-Talcott, S.U. Cocoplum (Chrysobalanus icaco L.) anthocyanins exert anti-inflammatory activity inhuman colon cancer and non-malignant colon cells. Food Funct. 2017, 8, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Sung, B.; Kang, Y.J.; Kim, N.H.; Jang, J.-Y.; Hwang, S.Y.; Kim, M.; Lim, H.S.; Yoon, J.-H.; Chung, H.Y.; et al. Apigenin-induced apoptosis is enhanced by inhibition of autophagy formation in HCT116 human colon cancer cells. Int. J. Oncol. 2014, 44, 1599–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; de Castro, A.J.; Lee, H.J.; Smolarek, A.K.; So, J.Y.; Simi, B.; Wang, C.X.; Zhou, R.; Rimando, A.M.; Suh, N. Dietary intake of pterostilbene, a constituent of blueberries, inhibits the β-catenin/p65 downstream signaling pathway and colon carcinogenesis in rats. Carcinogenesis 2010, 31, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Jin, Y.; Hofseth, A.B.; Pena, E.; Habiger, J.; Chumanevich, A.; Poudyal, D.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P.; et al. Resveratrol suppresses colitis and colon cancer associated with colitis. Cancer Prev. Res. 2010, 3, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Ramiro, I.; Ramos, S.; López-Oliva, E.; Agis-Torres, A.; Bravo, L.; Goya, L.; Martín, M. Ángeles Cocoa polyphenols prevent inflammation in the colon of azoxymethane-treated rats and in TNF-α-stimulated Caco-2 cells. Br. J. Nutr. 2012, 110, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, A.P. Preclinical animal tumor models to study prevention of colon cancer recurrence by curcumin. Anim. Models Cancer Drug Discov. 2019, 293–307. [Google Scholar] [CrossRef]
- Simons, C.C.J.M.; Hughes, L.A.; Arts, I.C.; Goldbohm, R.A.; Brandt, P.A.V.D.; Weijenberg, M.P. Dietary flavonol, flavone and catechin intake and risk of colorectal cancer in the Netherlands Cohort Study. Int. J. Cancer 2009, 125, 2945–2952. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Barupal, D.K.; Rothwell, J.A.; Jenab, M.; Fedirko, V.; Romieu, I.; Aleksandrova, K.; Overvad, K.; Kyrø, C.; Tjønneland, A.; et al. Dietary flavonoid intake and colorectal cancer risk in the European prospective investigation into cancer and nutrition (EPIC) cohort. Int. J. Cancer 2017, 140, 1836–1844. [Google Scholar] [CrossRef]
- Nimptsch, K.; Zhang, X.; Cassidy, A.; Song, M.; O’Reilly, ÉJ.; Lin, J.H.; Pischon, T.; Rimm, E.B.; Willett, W.C.; Fuchs, C.S.; et al. Habitual intake of flavonoid subclasses and risk of colorectal cancer in 2 large prospective cohorts. Am. J. Clin. Nutr. 2015, 103, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Xavier, C.P.R.; Lima, C.F.; Rohde, M.; Pereira-Wilson, C. Quercetin enhances 5-fluorouracil-induced apoptosis in MSI colorectal cancer cells through p53 modulation. Cancer Chemother. Pharmacol. 2011, 68, 1449–1457. [Google Scholar] [CrossRef]
- Hakim, L.; Alias, E.; Makpol, S.; Ngah, W.Z.W.; Morad, N.A.; Yusof, Y.A.M. Gelam Honey and Ginger Potentiate the Anti Cancer Effect of 5-FU against HCT 116 Colorectal Cancer Cells. Asian Pac. J. Cancer Prev. 2014, 15, 4651–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montrose, D.C.; Horelik, N.A.; Madigan, J.P.; Stoner, G.D.; Wang, L.-S.; Bruno, R.S.; Park, H.J.; Giardina, C.; Rosenberg, D.W. Anti-inflammatory effects of freeze-dried black raspberry powder in ulcerative colitis. Carcinogenesis 2010, 32, 343–350. [Google Scholar] [CrossRef] [PubMed]
- McFadden, R.-M.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The Role of Curcumin in Modulating Colonic Microbiota During Colitis and Colon Cancer Prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Buhrmann, C.; Kraehe, P.; Shayan, P.; Lueders, C.; Goel, A. Curcumin Chemosensitizes 5-Fluorouracil Resistant MMR-Deficient Human Colon Cancer Cells in High Density Cultures. PLoS ONE 2014, 9, e85397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrmann, C.; Shayan, P.; Kraehe, P.; Popper, B.; Goel, A.; Shakibaei, M. Resveratrol induces chemosensitization to 5-fluorouracil through up-regulation of intercellular junctions, Epithelial-to-mesenchymal transition and apoptosis in colorectal cancer. Biochem. Pharmacol. 2015, 98, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, L.; Ni, Z.; Sun, J.; Gao, H.; Cheng, Z.; Xu, J.; Qiu, Y. Resveratrol induces AMPK-dependent MDR1 inhibition in colorectal cancer HCT116/L-OHP cells by preventing activation of NF-κB signaling and suppressing cAMP-responsive element transcriptional activity. Tumor Boil. 2015, 36, 9499–9510. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Yoshino, K.; Kitajima, M. Possibility of preventing colorectal carcinogenesis with probiotics. Hepatogastroenterology 2011, 57, 1411–1415. [Google Scholar]
- Drago, L. Probiotics and Colon Cancer. Microorganisms 2019, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Hatakka, K.; Holma, R.; El-Nezami, H.; Suomalainen, T.; Kuisma, M.; Saxelin, M.; Poussa, T.; Mykkänen, H.; Korpela, R. The influence of Lactobacillus rhamnosus LC705 together with Propionibacterium freudenreichii ssp. shermanii JS on potentially carcinogenic bacterial activity inhuman colon. Int. J. Food Microbiol. 2008, 128, 406–410. [Google Scholar] [CrossRef]
- Lin, C.; Cai, X.; Zhang, J.; Wang, W.; Sheng, Q.; Hua, H.; Zhou, X. Role of Gut Microbiota in the Development and Treatment of Colorectal Cancer. Digesiton 2018, 100, 72–78. [Google Scholar] [CrossRef]
- Vinderola, G.; Perdigon, G.; Duarte, J.; Farnworth, E.; Matar, C. Effects of the oral administration of the exopolysaccharide produced by Lactobacillus kefiranofaciens on the gut mucosal immunity. Cytokine 2006, 36, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Perdigón, G. The Probiotic Bacterium Lactobacillus casei Induces Activation of the Gut Mucosal Immune System through Innate Immunity. Clin. Vaccine Immunol. 2006, 13, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozkurt, H.S.; Quigley, E.M.; Kara, B. Bifidobacterium animalissubspecieslactisengineered to produce mycosporin-like amino acids in colorectal cancer prevention. SAGE Open Med. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Wang, S.; Ma, L.; Lu, W.; Zhao, X.; Yang, Y.; Li, X.; Wang, H.; Sun, W. Acupuncture: How objective is the evidence from randomised clinical trials? BJOG Int. J. Obstet. Gynaecol. 2011, 118, 1141–1142. [Google Scholar] [CrossRef]
- Rafter, J.; Bennett, M.; Caderni, G.; Clune, Y.; Hughes, R.; Karlsson, P.C.; Klinder, A.; O’Riordan, M.; O’Sullivan, G.C.; Pool-Zobel, B.; et al. Dietary synbiotics reduce cancer risk factors in polypectomized and colon cancer patients. Am. J. Clin. Nutr. 2007, 85, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Hibberd, A.; Lyra, A.; Ouwehand, A.C.; Rolny, P.; Lindegren, H.; Cedgård, L.; Wettergren, Y. Intestinal microbiota is altered in patients with colon cancer and modified by probiotic intervention. BMJ Open Gastroenterol. 2017, 4, e000145. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Xu, L.; Zhangab, D.; Wu, Z. Effect of probiotics on small intestinal bacterial overgrowth in patients with gastric and colorectal cancer. Turk. J. Gastroenterol. 2016, 27, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Xin, Y.; Zhang, C.; Wu, D.; Ding, D.; Tang, L.; Owusu, L.; Bai, J.; Li, W. Fermentation supernatants of Lactobacillus delbrueckii inhibit growth of human colon cancer cells and induce apoptosis through a caspase 3-dependent pathway. Oncol. Lett. 2014, 7, 1738–1742. [Google Scholar] [CrossRef] [Green Version]
- Konishi, H.; Fujiya, M.; Tanaka, H.; Ueno, N.; Moriichi, K.; Sasajima, J.; Ikuta, K.; Akutsu, H.; Tanabe, H.; Kohgo, Y. Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis. Nat. Commun. 2016, 7, 12365. [Google Scholar] [CrossRef] [PubMed]
- Pouncey, A.L.; Scott, A.J.; Alexander, J.L.; Marchesi, J.R.; Kinross, J.M. Gut microbiota, chemotherapy and the host: The influence of the gut microbiota on cancer treatment. Ecancermedicalscience 2018, 12, 868. [Google Scholar] [CrossRef] [Green Version]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal Bacteria Control Cancer Response to Therapy by Modulating the Tumor Microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-W.; Liu, C.-Y.; Lee, H.-C.; Huang, Y.-H.; Li, L.-H.; Chiau, J.-S.C.; Wang, T.-E.; Chu, C.-H.; Shih, S.-C.; Tsai, T.-H.; et al. Lactobacillus casei Variety rhamnosus Probiotic Preventively Attenuates 5-Fluorouracil/Oxaliplatin-Induced Intestinal Injury in a Syngeneic Colorectal Cancer Model. Front. Microbiol. 2018, 9, 983. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Tang, W.; Fan, X.; Wu, G. Intestinal microbiota: A novel perspective in colorectal cancer biotherapeutics. Onco Targets Ther. 2018, 11, 4797–4810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, V.; Le, D.T. Efficacy of PD-1 blockade in tumors with MMR deficiency. Immunotherapy 2016, 8, 1–3. [Google Scholar] [CrossRef]
- Routy, B.; le Chatelier, E.; de Rosa, L.; Duong, C.P.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1–based immunotherapy against epithelial tumors. Science 2017, 359, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Österlund, P.; Ruotsalainen, T.; Korpela, R.; Saxelin, M.; Ollus, A.; Valta, P.; Kouri, M.; Elomaa, I.; Joensuu, H. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: A randomised study. Br. J. Cancer 2007, 97, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Packey, C.D.; A Ciorba, M. Microbial influences on the small intestinal response to radiation injury. Curr. Opin. Gastroenterol. 2010, 26, 88–94. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Study (Reference) | Dietary Mediator | Type of Study | Species | Most Relevant Results |
|---|---|---|---|---|
| Dietary Fiber | ||||
| Lattimer et al. 2010 [93]) | Dietary Fiber (Arabinoxylan, Inulin, β-glucan, Pectin, Bran, Cellulose, Resistant Starch) | In vivo | Human | ↑ Excretion of bile acids, ↑ Production of fecal SCFAs ↑ Antioxidants ↓ Cancer prevalence |
| Zeng et al. 2014 [94] | Dietary Fiber | In vivo | Human | ↓ Fecal pH in the colon ↑ SCFA-producing gut bacteria ↑ Apoptosis of colon cancer cells ↓ Chronic inflammatory process and migration/ invasion of colon cancer cells |
| Deehan et al. 2020 [103] | Dietary Fiber | In vivo | Human | Modulation of the colon microbiota ↑ Saccharolytic fermentation ↑ Production of fecal SCFAs |
| Chen et al. 2013 [104] | Dietary Fiber | In vivo | Human | ↑ Production of SCFAs by healthy gut microbiota, ↓ Risk of advanced colorectal adenoma. |
| Burkitt et al. 1993 [108]; Bergman et al. 1990 [109]; Hamer et al. 2008 [110] | Dietary Fiber | In vivo | Human/ Mouse | ↑ Production of fecal SCFAs (especially butyrate) ↓ Fecal pH in the colon, ↓ Pathogenic organism proliferation ↓ DNA damage induction ↑ Apoptosis of colon cancer cells ↓ Proliferation of colon cancer cells. |
| Fung et al. 2012 [111]; Neish et al. 2009 [112] | Long-term dietary fiber intake | In vivo | Human | ↑ Abundance of Firmicutes abundance ↑ Immune modulatory and anti-inflammatory effects in the host |
| Bingham et al. 2003 [95] | Dietary Fiber | In vivo | Human | ↑ Total dietary fiber intake ↓ Risk of CRC |
| Schatzkin et al. 2007 [96] | Dietary Fiber (whole grains) | In vivo | Human | ↑ Whole grain food consumption ↓ Risk of CRC (modest) |
| Dahm et al. 2010 [97] | Dietary Fiber | In vivo | Human | ↑ Fiber intake ↓Risk of CRC |
| Hansen et al. 2012 [99] | Dietary Fiber (cereals) | In vivo | Human | ↑ Total dietary fiber ↓ Risk of CRC |
| Song M et al. 2018 [100] | Dietary Fiber (whole grains) | In vivo | Human | ↑ Survival rates of non-metastatic CRC |
| Moen et al. 2016 [101] | Dietary Fiber (inulin, cellulose, brewers spent grain) | In vivo | Mouse | Inulin intake change cecal microbiota ↓ Colonic tumorigenesis |
| Mehta et al. 2017 [102] | Dietary Fiber (whole grains) | In vivo | Human | ↓ Risk of developing Fusobacterium nucleatum-positive CRC |
| O’Keefe et al. 2015 [105] | Dietary fiber and fat | In vivo | Human | ↑ Saccharolytic fermentation ↑ Butyrogenesis ↓ Secondary bile acid synthesis ↓ Biomarkers of colon cancer risk |
| Donohoe et al. 2014 [106] | Dietary Fiber | In vivo | Mouse | ↑ Microbial fiber fermentation ↑ Butyrate production ↑ Protection against colorectal tumorigenesis. |
| Bishehsari et al. 2018 [107] | Dietary Fiber | In vivo | Mouse | ↑ SCFA-producing bacteria, ↓ Gut microbiota dysbiosis ↓ Polyposis incidence |
| Diets rich in polyunsaturated fatty acids | ||||
| Costantini et al. 2017 [114] | n-3 PUFAs | In vivo | Human/ Mouse | ↓ Relative abundance of Faecalibacterium ↑ Bacteroidetes and butyrate-producing bacteria (Lachnospiraceae family) |
| Cho et al. 2014 [116] | n-3 Fatty Acid Docosahexaenoic Acid and Butyrate | In vitro | Human | Epigenetic alterations (methylation of proapoptotic genes) ↑ Apoptosis of colon cancer cells |
| Chapkin et al. 2014 [117] | n-3 PUFAs | In vivo | Human/ Mouse | Alterations in the plasma membrane of colon cancer cells Epigenetic alterations ↑ Risk of developing CRC. |
| Triff et al. 2015 [118] | n-3 PUFA & Fiber | In vivo | Human/ Mouse | Regulation of nuclear receptor transcriptional activity ↓ Inflammatory cytokines ↑ Chemoprotection. |
| Hong et al. 2015 [119] | Fish oil & Butyrate | In vivo | Rat | ↑ Apoptosis of colon cancer cells ↓ Proliferation of colon cancer cells ↑ Protection against CRC |
| Lee et al. 2017 [121] | Fish oil & Butyrate | In vivo | Human | Modulation of CRC-related gene expression ↓ Inflammation ↑ Apoptosis of colon cancer cells |
| Chang et al. 1998 [125] | Fish oil & Fiber (pectin, cellulose) | In vivo | Rat | ↑ Apoptosis of colon cancer cells ↓ Proliferation of colon cancer cells ↓ Rate of CRC adenocarcinoma incidence |
| Cho et al. 2012 [127] | Fish oil & Pectin | In vivo | Rat | ↑ Apoptosis of colonocytes ↑ Chemoprotective capacity |
| Ng et al. 2005 [128] | Docosahexaenoic acid (DHA, 22:6 n-3) & butyrate | In vitro | Human | ↑ Mitochondrial lipid oxidation ↓ Mitochondrial membrane potential ↑ Apoptosis of colonocytes ↑ Chemoprotective effects |
| Sofi et al. 2019 [129] | Comparison of Meat-Based vs Pesco-Vegetarian Diets | In vivo | Human | Positive effect of pesco-vegetarian diet on gut microbiota ↓ Risk of CRC |
| Rani et al. 2017 [131]; Ran et al. 2014 [132]; Sebe et al. 2016 [133] | n-3 PUFAs& 5-FU | In vivo | Mouse | ↓ Tumor burden and DNA damage ↓ Mucosal deterioration, ↑ Apoptosis ↓ 5-FU-related toxicity (intestinal mucositis) ↑ 5-FU anti-cancer activity |
| Ebadi et al. 2017 [134] | PUFAs & irinotecan | In vivo | Rat | Modulation of adipose tissue mitochondrial function ↓ 5-FU-associated side effects |
| Cai et al. 2014 [135] | n-3 PUFAs | In vitro | Human | ↑ Lipid peroxidation, Modulation of the inflammatory response ↑ Apoptosis ↓ Cytotoxicity by radiation therapy |
| Granci et al. 2013 [136] | Fish oil& & 5-FU, oxaliplatin and irinotecan | In vitro | Human | ↑ Apoptosis ↓ Cytotoxic effects of 5-FU, oxaliplatin and irinotecan. |
| Volpato et al. 2018 [113] | n-3 PUFAs | In vitro /In vivo | Human/ Mouse | ↑ Butyrate-producing gut bacteria ↓ Inflammation; ↑ Apoptosis ↓ Proliferation of colon cancer cells |
| Watson et al. 2018 [115] | n-3 PUFAs | In vivo | Human | ↓ Gut microbiota dysbiosis ↓ Pathogenic gut bacteria ↑ SCFA-producing gut bacteria (Bifidobacterium, Roseburia and Lactobacillus). |
| Piazzi et al. 2014 [120] | Eicosapentaenoic Acid | In vivo | Mouse | ↑ Lactobacillus species in the gut microbiota ↓ Size of CRC tumors ↓ Proliferation colon cancer cells ↑ Apoptosis colon cancer cells |
| Song et al. 2015 [122] | n-3 PUFAs | In vivo | Human | ↓ Risk of microsatellite instability ↑ DNA repair systems mismatch pathways |
| Yang et al. 2015 [124] | n-3 PUFAs | In vivo | Human | Different PUFA composition between normal and cancerous tissues ↓ Inflammation in CRC tumorigenesis. |
| Song et al. 2017 [126] | Marine ω-3 PUFAs | In vivo | Human | ↑ Intake of marine ω-3 after CRC diagnosis ↓ Risk of CRC-specific mortality. |
| Aglago et al. 2020 [130] | n-3 PUFAs | In vivo | Human | Regular intake of fish at recommended levels ↓ Risk of CRC |
| Golkhalkhali et al. 2018 [137] | n-3 PUFAs & probiotic supplement | In vivo | Human | ↑ Tolerability of capecitabine/oxaliplatin chemotherapy ↑ Quality of life markers ↓ Chemotherapy-induced symptoms (diarrhea and fatigue) |
| Bioactive polyphenols | ||||
| Mileo et al. 2019 [139] | Polyphenols | In vivo/In vitro | Human/ Mouse | ↑ Gut microbiota balance ↓ Proliferation of colon cancer cells ↑ Apoptosis of colon cancer cells |
| Miene et al. 2009 [143] | Polyphenols (Apple) | In vitro | Human | Polyphenols are metabolized by colonic microbiota ↓ DNA damage induced by oxidative stress in colonic adenoma cells |
| Gibellini et al. 2011 [150] | Quercetin | In vivo | Human | ↓ Proliferation of colon cancer cells ↑ Apoptosis of colon cancer cells |
| Venancio et al. 2017 [151] | Polyphenols (Cocoplum) | In vitro | Human | Anti-inflammatory activity and pro-oxidant effects |
| Lee Y et al. 2014 [152] | Apigenin | In vitro | Human | ↓ Cell cycle progression ↓ Autophagy ↑ Apoptosis |
| Xavier et al. 2011 [160] | Polyphenols | In vitro | Human | ↑ Apoptosis (in combination with 5-FU) |
| Hakim et al. 2014 [161] | Gelam Honey and Ginger | In vitro | Human | ↑ Anticancer activity of 5-FU |
| Montrose et al. 2015 [162] | Black Raspberry (Anthocyanins, simple phenols, ellagic acid and quercetin) | In vivo | Mouse | ↓ Expression of proinflammatory cytokines (TNF-α and IL-1β) ↓ Plasma levels of COX-2 and prostaglandin E2 ↑ Chemopreventive effect |
| McFadden et al. 2015 [163] | Curcumin | In vivo | Mouse | ↑ Microbial diversity ↓ Colonic tumor burden ↑ Chemopreventive effect |
| Shakibaei et al. 2014 [164] | Curcumin | In vitro | Human | ↑ Chemosensitization to 5-FU treatment |
| Buhrmann et al. 215 [165] | Resveratrol | In vitro | Human | ↑ Chemosensitization to 5-FU treatment |
| Wang et al. 2015 [166] | Resveratrol | In vitro | Human | ↓ Drug resistance (down-regulation of multi-drug resistant protein 1), ↓ Activation of NF-κB signaling ↓ Transcriptional activity of the cAMP-sensitive element |
| Paul et al. 2010 [153] | Pterostilbene (Blueberries) | In vivo | Rat | ↓ Colon tumorigenesis by regulating the Wnt/b-catenin-signaling pathway ↓ Inflammatory responses. |
| Cui et al. 2010 [154] | Resveratrol | In vivo | Mouse | ↓ Colitis-driven colon cancer incidence |
| Rodríguez-Ramiro et al. 2013 [155] | Polyphenols (Cocoa) | In vivo | Rat | Anti-inflammatory effect on the colonic tissue Chemoprevention in the early stages |
| Simons et al. 2009 [157] | Flavonol, Flavone and Catechin | In vivo | Human | ↓ Risk of CRC |
| Zamora-Ros et al. 2017 [158] | Flavonoid | In vivo | Human | No association between regular dietary intake of flavonoids and CRC risk |
| Sánchez et al. 2019 [148]; Cueva et al. 2020 [149] | Red wine Polyphenols | In vivo | Human | Modulation of the gut microbiota composition ↓ Growth of pathogenic bacterial species (F. nucleatum and P. Gingivalis) ↓ Adhesion to oral cells ↓ Risk of CRC |
| Probiotics | ||||
| Hatakka et al. 2008 [169] | Lactobacillus rhamnosus LC705 and Propionibacterium freudenreichii ssp. | In vivo | Human | Fecal counts of Lactobacilli and Propionibacteria ↓ β-glucosidase activity ↑ CRC prevention |
| Vinderola et al. 2006 [171] | Lactobacillus kefiranofaciens | In vivo | Human | Regulation of the immune system, ↑ Phagocytosis of tumor cells in early stages. |
| Galdeano et al. 2007 [172] | Lactobacillus casei | In vivo | Mouse | Induction of innate immunity influencing the clonal expansion of IgA B-cell population, ↓ Risk of CRC. |
| Bozkurt et al. 2019 [173] | Bifidobacterium animalis subsp. lactis | In vivo | Human/ Mouse | ↑ Production mycosporin-like amino acids Modulation of the immune system to regulate the proliferation and differentiation of intestinal epithelial cells, macrophages, lymphocytes and cytokine production |
| Liu et al. 2011 [174] | Lactobacillus plantarum, Lactobacillus acidophilus & Bifidobacterium Longum | In vivo | Human | ↑ Integrity of the intestinal barrier, ↑ Gut microbiota balance ↓ Post-operative infection rate |
| Rafter et al. 2007 [175] | Lactobacillus rhamnosus & Bifidobacterium lactis | In vivo | Human | Modulation of the gut microbiota composition ↓ Intestinal permeability, ↓ Cancer biomarkers (cell proliferation). |
| Hibberd et al. 2017 [176] | Bifidobacterium animalis subsp. lactis Bl-04 & Lactobacillus acidophilus NCFM | In vivo | Human | ↑ Abundance of butyrate-producing bacteria in tumor, mucosa and fecal samples |
| Liang et al. 2016 [177] | Bifidobacterium | In vivo | Human | ↓ Gut microbiota dysbiosis in CRC patients |
| Wan et al. 2014 [178] | Lactobacillus delbrueckii | In vitro | Cell line SW620 | ↓ Proliferation of colon cancer cells ↑ Apoptosis of colon cancer cells (via caspase 3 pathway) |
| Konishi et al. 2016 [179] | Lactobacillus casei strain ATCC 334 | In vivo | Human | ↑ Production of ferrichrome ↑ Apoptosis of colon cancer cells (via JNK pathway) ↓ Progression of CRC |
| Chang et al. 2018 [182] | Lactobacillus casei Variety rhamnosus & 5-FU/oxaliplatin | In vivo | Mouse | ↓ Intestinal mucositis derived from anticancer treatment. |
| Ding et al. 2018 [183]; Lee et al. 2016 [184]; Routy et al. 2018 [185] | Bifidobacterium & PD-1-based immunotherapy | In vivo | Human/ Mouse | ↓ Tumor growth ↓ Side effects induced by PD-1-based immunotherapy |
| Osterlund et al. 2007 [186] | Lactobacillus rhamnosus | In vivo | Human | ↓ Side effects (severe diarrhea, abdominal distress) induced by chemotherapy. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Alcoholado, L.; Ramos-Molina, B.; Otero, A.; Laborda-Illanes, A.; Ordóñez, R.; Medina, J.A.; Gómez-Millán, J.; Queipo-Ortuño, M.I. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers 2020, 12, 1406. https://doi.org/10.3390/cancers12061406
Sánchez-Alcoholado L, Ramos-Molina B, Otero A, Laborda-Illanes A, Ordóñez R, Medina JA, Gómez-Millán J, Queipo-Ortuño MI. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers. 2020; 12(6):1406. https://doi.org/10.3390/cancers12061406
Chicago/Turabian StyleSánchez-Alcoholado, Lidia, Bruno Ramos-Molina, Ana Otero, Aurora Laborda-Illanes, Rafael Ordóñez, José Antonio Medina, Jaime Gómez-Millán, and María Isabel Queipo-Ortuño. 2020. "The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response" Cancers 12, no. 6: 1406. https://doi.org/10.3390/cancers12061406