The Progressive Mutagenic Effects of Acidic Bile Refluxate in Hypopharyngeal Squamous Cell Carcinogenesis: New Insights


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
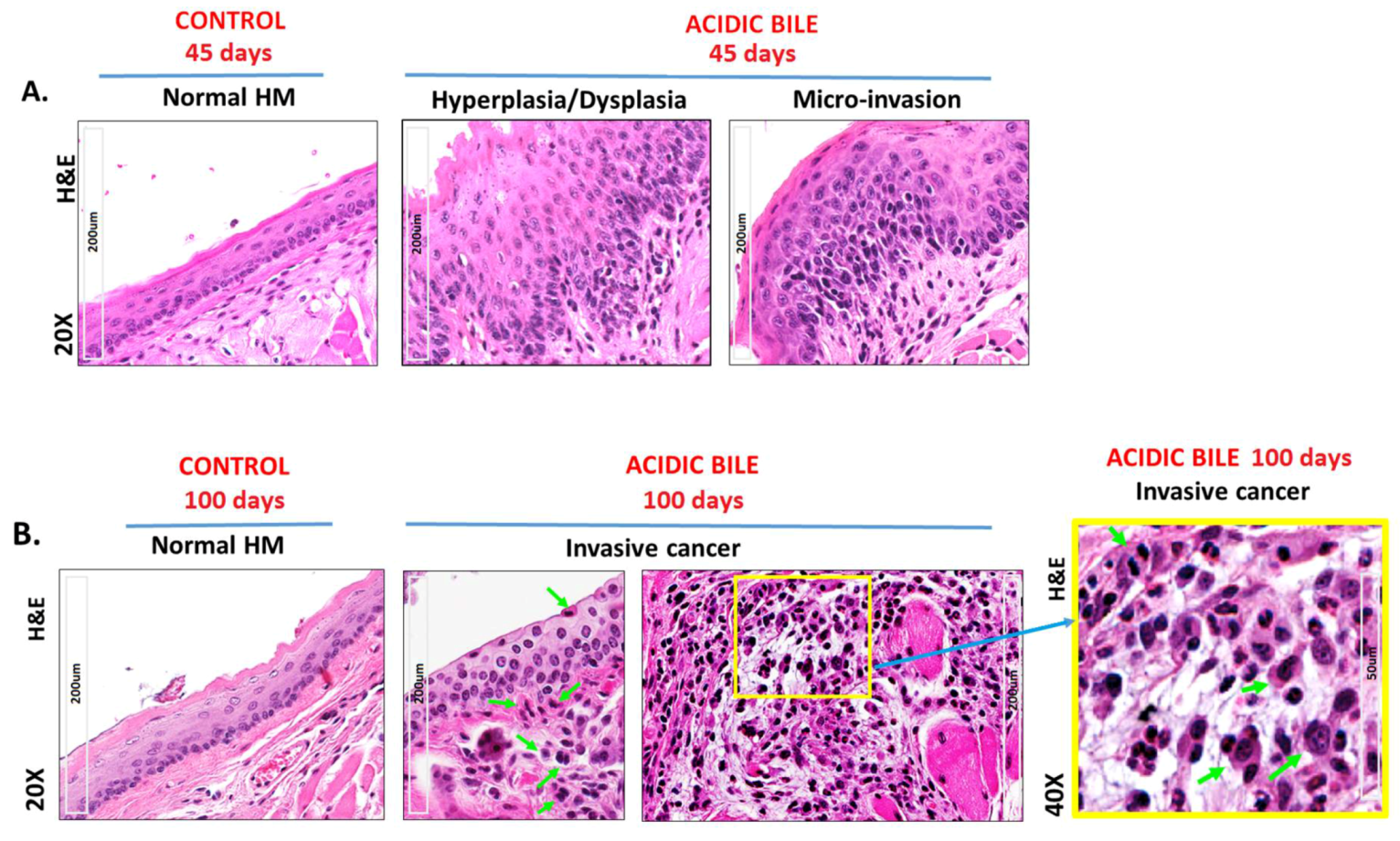
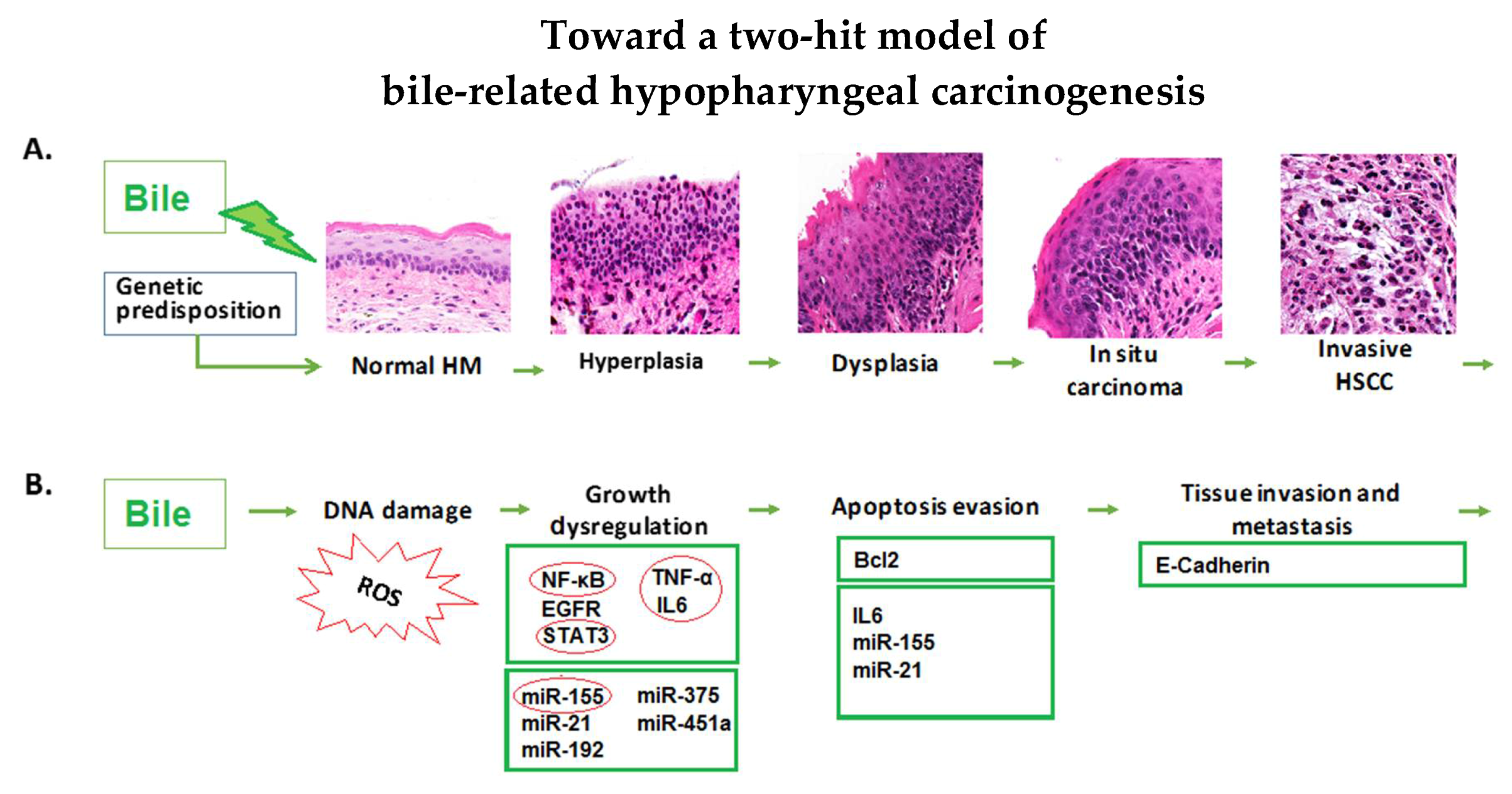
2.1. Long-Term Exposure of HM to Acidic Bile Progressively Induced a Malignant Histopathologic Phenotype
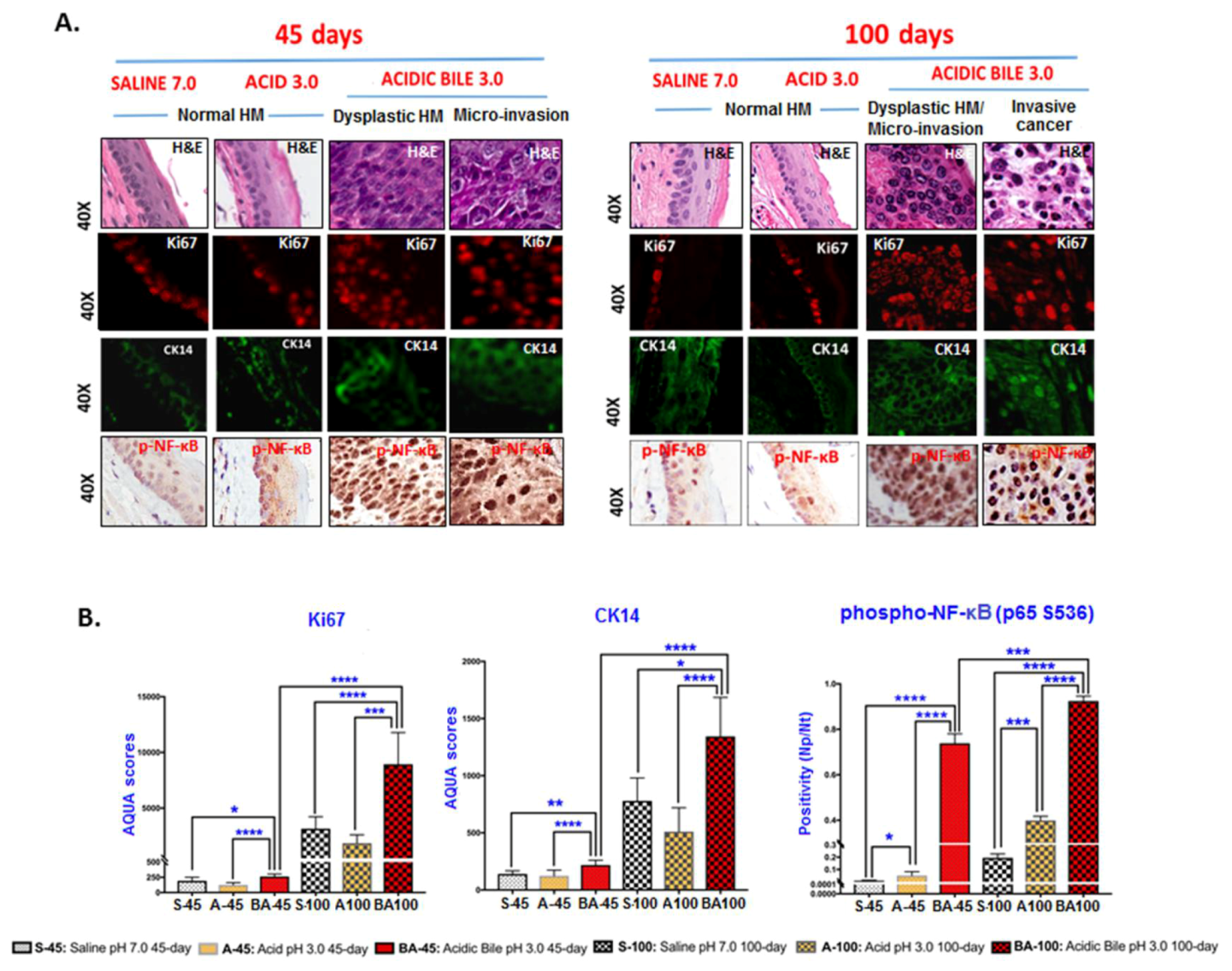
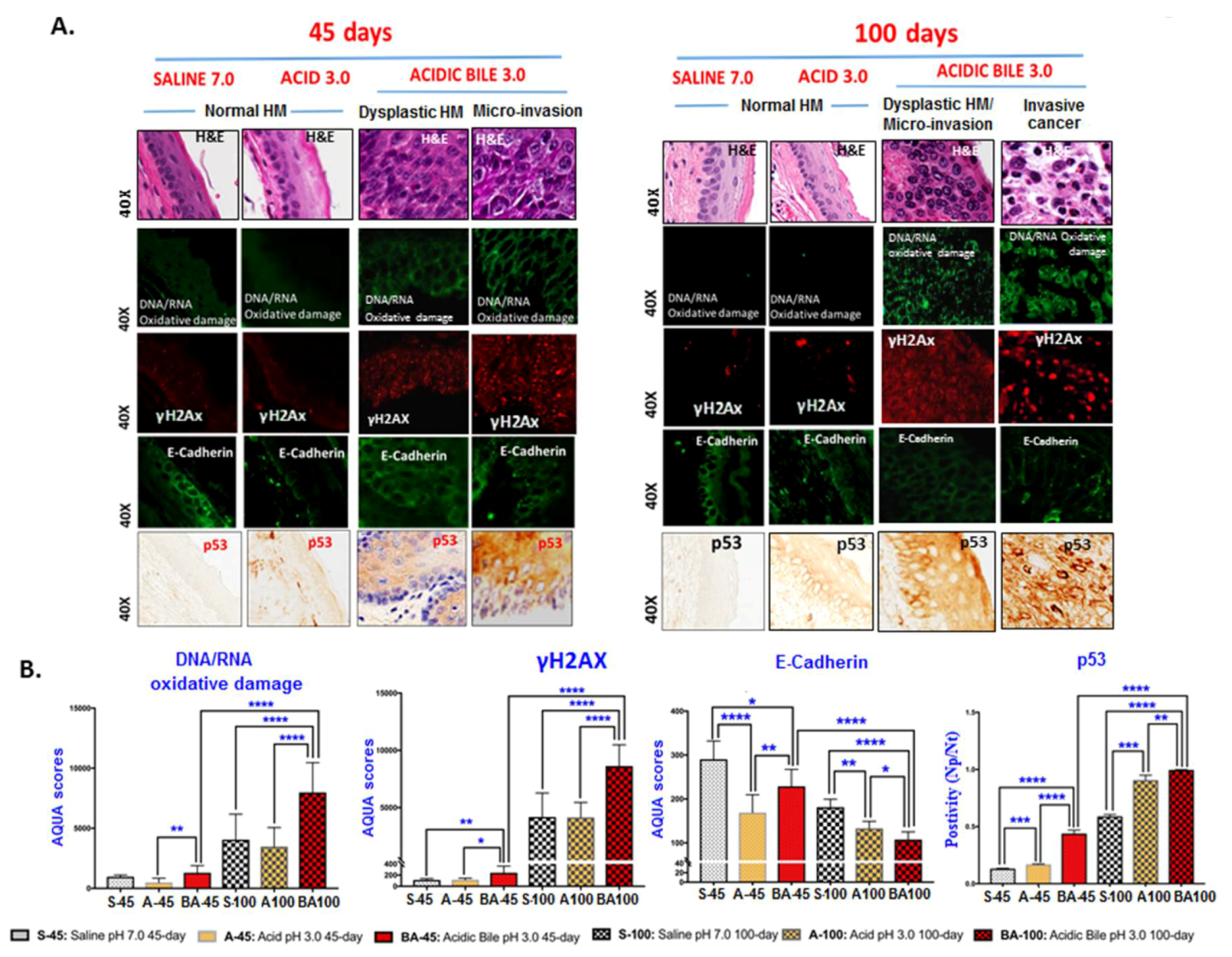
2.2. Long-Term Exposure to Acidic Bile Progressively Induced Increased NF-κB Activation and Elevated Expression Levels of p53 in HM
2.3. Correlation Analysis
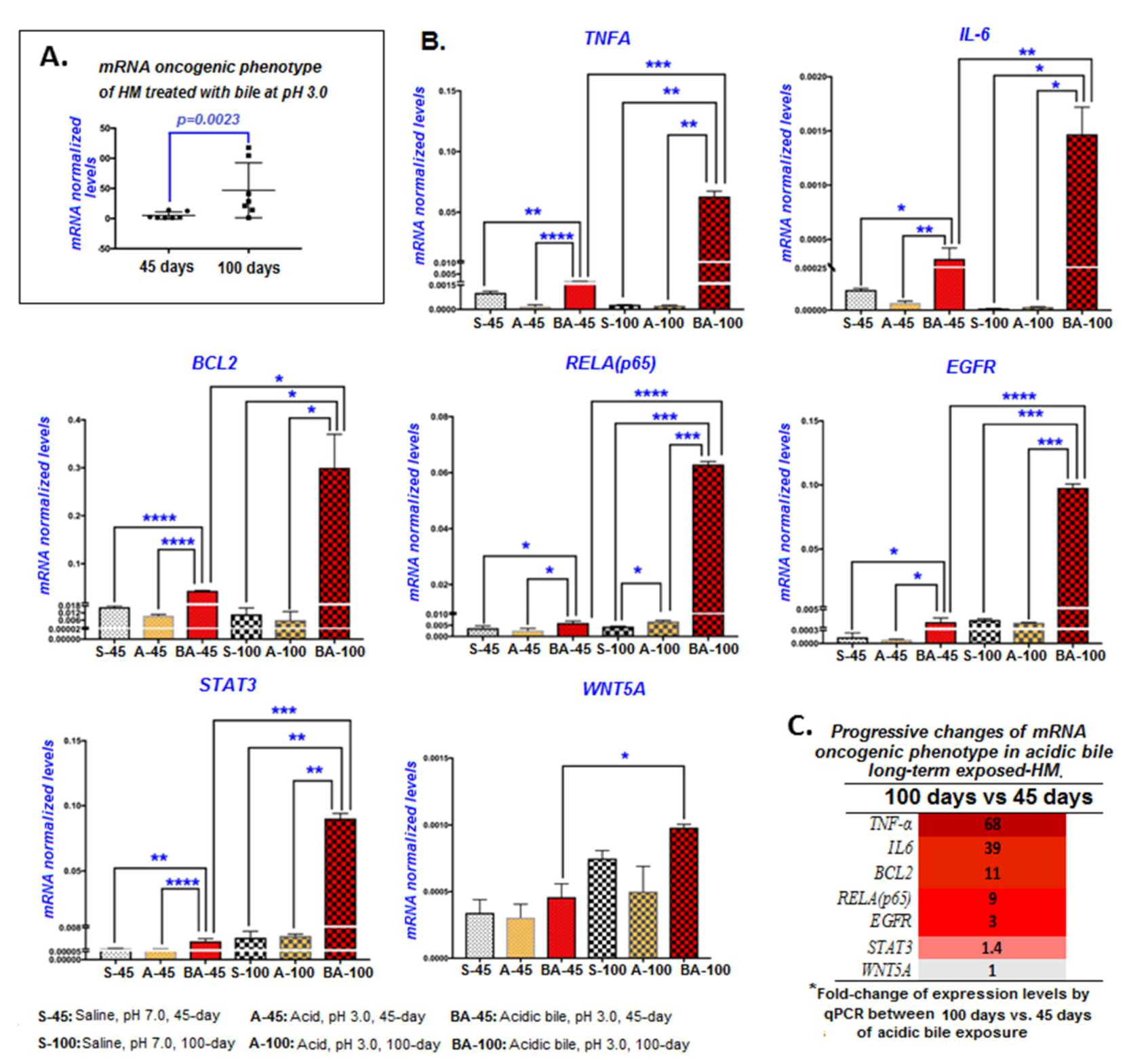
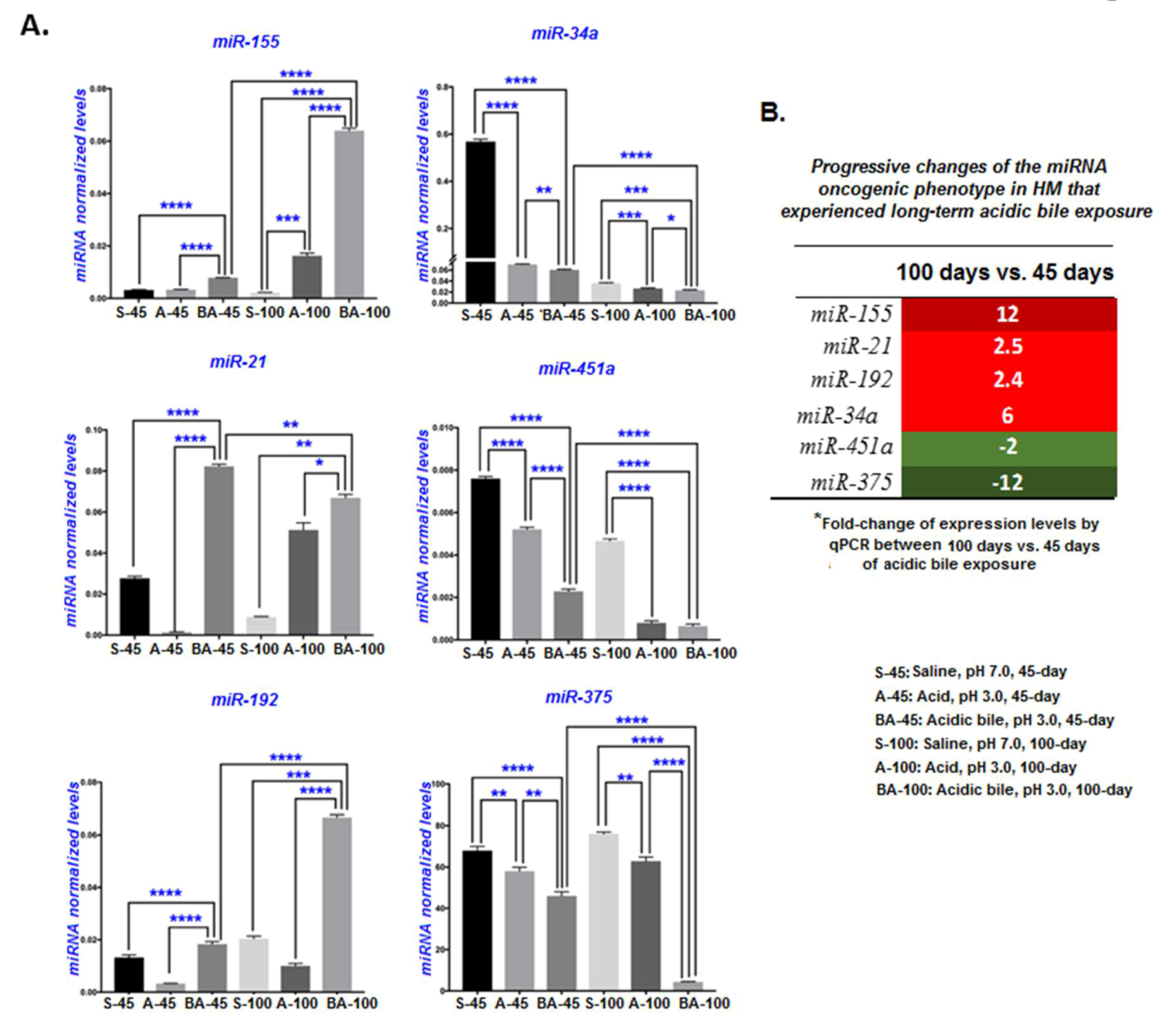
2.4. Long-Term Exposure of HM to Acidic Bile Progressively Induced Increased Transcriptional Levels of an Oncogenic Phenotype
2.5. Long-Term Exposure of HM to Acidic Bile Progressively Induced Increased Deregulations of Inflammatory-Related miR-155 and the Tumor Suppressors miR-375 and miR-451a
3. Discussion
4. Materials and methods
4.1. In Vivo Model
4.2. Tissue Examination
4.3. Immunohistochemical, Gene Expression, and miRNA Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- American Cancer Society. Laryngeal and Hypopharyngeal Cancer. 2019. Available online: https://www.cancer.org/cancer/laryngeal-and-hypopharyngeal-cancer/about/key-statistics.html (accessed on 8 January 2019).
- Chang, D.C.; Chen, A.W.; Lo, Y.S.; Chuang, Y.C.; Chen, M.K. Factors associated with suicidal ideation risk in head and neck cancer: A longitudinal study. Laryngoscope 2019, 129, 2491–2495. [Google Scholar] [CrossRef] [PubMed]
- Curado, M.P.; Hashibe, M. Recent changes in the epidemiology of head and neck cancer. Curr. Opin. Oncol. 2009, 21, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Galli, J.; Cammarota, G.; De Corso, E.; Agostino, S.; Cianci, R.; Almadori, G.; Paludetti, G. Biliary laryngopharyngeal reflux: A new pathological entity. Curr. Opin. Otolaryngol. Head Neck Surg. 2006, 14, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Langevin, S.M.; Michaud, D.S.; Marsit, C.J.; Nelson, H.H.; Birnbaum, A.E.; Eliot, M.; Christensen, B.C.; McClean, M.D.; Kelsey, K.T. Gastric reflux is an independent risk factor for laryngopharyngeal carcinoma. Cancer Epidemiol. Biomarkers Prev. 2013, 22, 1061–1068. [Google Scholar] [CrossRef] [Green Version]
- Vaezi, M.; Richter, J. Double reflux: Double trouble. Gut 1999, 44, 590–592. [Google Scholar] [CrossRef] [Green Version]
- McQuaid, K.R.; Laine, L.; Fennerty, M.B.; Souza, R.; Spechler, S.J. Systematic review: The role of bile acids in the pathogenesis of gastro-oesophageal reflux disease and related neoplasia. Aliment. Pharmacol. Ther. 2011, 34, 146–165. [Google Scholar] [CrossRef]
- Kauer, W.K.; Stein, H.J. Role of acid and bile in the genesis of Barrett’s esophagus (Review). Chest Surg. Clin. N. Am. 2002, 12, 39–45. [Google Scholar] [CrossRef]
- Covington, M.F.; Krupinski, E.; Avery, R.J.; Kuo, P.H. Classification schema of symptomatic enterogastric reflux utilizing sincalide augmentation on hepatobiliary scintigraphy. J. Nucl. Med. Technol. 2014, 42, 198–202. [Google Scholar] [CrossRef]
- Galli, J.; Calò, L.; Agostino, S.; Cadoni, G.; Sergi, B.; Cianci, R.; Cammarota, G. Bile reflux as possible risk factor in laryngopharyngeal inflammatory and neoplastic lesions. Acta Otorhinolaryngol. Ital. 2003, 23, 377–382. [Google Scholar]
- Bayerdorffer, E.; Mannes, G.A.; Ochsenkuhn, T.; Dirschedl, P.; Wiebecke, B.; Paumgartner, G. Unconjugated secondary bile acids in the serum of patients with colorectal adenomas. Gut 1995, 36, 268–273. [Google Scholar] [CrossRef] [Green Version]
- Fein, M.; Peters, J.H.; Chandrasoma, P.; Ireland, A.P.; Oberg, S.; Ritter, M.P.; Bremner, C.G.; Hagen, J.A.; DeMeester, T.R. Duodenoesophageal reflux induces esophageal adenocarcinoma without exogenous carcinogen. J. Gastrointest. Surg. 1998, 2, 260–268. [Google Scholar] [CrossRef]
- McAdam, E.; Haboubi, H.N.; Griffiths, A.P.; Baxter, J.N.; Spencer-Harty, S.; Davies, C.; Jenkins, G.J. Reflux composition influences the level of NF-κB activation and upstream kinase preference in oesophageal adenocarcinoma cells. Int. J. Cancer 2015, 136, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauer, W.K.; Peters, J.H.; DeMeester, T.R.; Ireland, A.P.; Bremner, C.G.; Hagen, J.A. Mixed reflux of gastric and duodenal juices is more harmful to the esophagus than gastric juice alone. The need for surgical therapy re-emphasized. Ann. Surg. 1995, 222, 525–531, discussion 531–533. [Google Scholar] [CrossRef] [PubMed]
- Nehra, D.; Howell, P.; Williams, C.P.; Pye, J.K.; Beynon, J. Toxic bile acids in gastro-oesophageal reflux disease: Influence of gastric acidity. Gut 1999, 44, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Vageli, D.P.; Prasad, M.L.; Sasaki, C.T. Gastro-duodenal fluid induced Nuclear Factor-κappaB activation and early pre-malignant alterations in murine hypopharyngeal mucosa. Oncotarget 2016, 7, 5892–5908. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, C.T.; Vageli, D.P. miR-21, miR-155, miR-192, and miR-375 Deregulations Related to NF-kappaB Activation in Gastroduodenal Fluid-Induced Early Preneoplastic Lesions of Laryngeal Mucosa In Vivo. Neoplasia 2016, 18, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, C.T.; Doukas, S.G.; Vageli, D.P. In Vivo Short-Term Topical Application of BAY 11-7082 Prevents the Acidic Bile-Induced mRNA and miRNA Oncogenic Phenotypes in Exposed Murine Hypopharyngeal Mucosa. Neoplasia 2018, 20, 374–386. [Google Scholar] [CrossRef]
- Vageli, D.P.; Doukas, S.G.; Spock, T.; Sasaki, C.T. Curcumin prevents the bile reflux-induced NF-κB-related mRNA oncogenic phenotype, in human hypopharyngeal cells. J. Cell Mol. Med. 2018, 22, 4209–4220. [Google Scholar] [CrossRef] [PubMed]
- Doukas, S.G.; Vageli, D.P.; Sasaki, C.T. NF-κB inhibition reverses acidic bile-induced miR-21, miR-155, miR-192, miR-34a, miR-375 and miR-451a deregulations in human hypopharyngeal cells. J. Cell Mol. Med. 2018, 22, 2922–2934. [Google Scholar] [CrossRef] [Green Version]
- Vageli, D.P.; Doukas, S.G.; Sasaki, C.T. Inhibition of NF-κB prevents the acidic bile-induced oncogenic mRNA phenotype, in human hypopharyngeal cells. Oncotarget 2017, 9, 5876–5891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doukas, P.G.; Vageli, D.P.; Doukas, S.G.; Sasaki, C.T. Temporal characteristics of NF-κB inhibition in blocking bile-induced oncogenic molecular events in hypopharyngeal cells. Oncotarget 2019, 10, 3339–3351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M. Nuclear factor-kappaB in cancer development and progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Loercher, A.; Lee, T.L.; Ricker, J.L.; Howard, A.; Geoghegen, J.; Chen, Z.; Sunwoo, J.B.; Sitcheran, R.; Chuang, E.Y.; Mitchell, J.B.; et al. Nuclear factor-kappaB is an important modulator of the altered gene expression profile and malignant phenotype in squamous cell carcinoma. Cancer Res. 2004, 64, 6511–6523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nottingham, L.K.; Yan, C.H.; Yang, X.; Si, H.; Coupar, J.; Bian, Y.; Cheng, T.F.; Allen, C.; Arun, P.; Gius, D.; et al. Aberrant IKKα and IKKβ cooperatively activate NF-κB and induce EGFR/AP1 signaling to promote survival and migration of head and neck cancer. Oncogene 2014, 33, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, J.D.; Grandis, J.R. The molecular pathogenesis of head and neck cancer. Cancer Biol. Ther. 2010, 9, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.T.; Ricker, J.L.; Chen, Z.; Van Waes, C. Role of activated nuclear factor-kappaB in the pathogenesis and therapy of squamous cell carcinoma of the head and neck. Head Neck 2007, 29, 959–971. [Google Scholar] [CrossRef]
- Lee, T.L.; Yang, X.P.; Yan, B.; Friedman, J.; Duggal, P.; Bagain, L.; Dong, G.; Yeh, N.T.; Wang, J.; Zhou, J.; et al. A novel nuclear factor-kappaB gene signature is differentially expressed in head and neck squamous cell carcinomas in association with TP53 status. Clin. Cancer Res. 2007, 13, 5680–5691. [Google Scholar] [CrossRef] [Green Version]
- Molinolo, A.A.; Amornphimoltham, P.; Squarize, C.H.; Castilho, R.M.; Patel, V.; Gutkind, J.S. Dysregulated molecular networks in head and neck carcinogenesis. Oral Oncol. 2009, 45, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Tili, E.; Michaille, J.J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Hu, A.; Huang, J.J.; Xu, W.H.; Jin, X.J.; Li, J.P.; Tang, Y.J.; Huang, X.F.; Cui, H.J.; Sun, G.B. miR-21 and miR-375 microRNAs as candidate diagnostic biomarkers in squamous cell carcinoma of the larynx: Association with patient survival. Am. J. Transl. Res. 2014, 6, 604–613. [Google Scholar] [PubMed]
- Zhao, X.D.; Zhang, W.; Liang, H.J.; Ji, W.Y. Overexpression of miR-155 promotes proliferation and invasion of human laryngeal squamous cell carcinoma via targeting SOCS1 and STAT3. PLoS ONE 2013, 8, e56395. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Zhan, G.; Ye, D.; Ren, Y.; Cheng, L.; Wu, Z.; Guo, J. MicroRNA-34a affects the occurrence of laryngeal squamous cell carcinoma by targeting the antiapoptotic gene survivin. Med. Oncol. 2010, 29, 2473–2480. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, I.; Kinoshita, T.; Hanazawa, T.; Kikkawa, N.; Chiyomaru, T.; Enokida, H.; Yamamoto, N.; Goto, Y.; Nishikawa, R.; Nakagawa, M.; et al. Identification of tumour suppressive microRNA-451a in hypopharyngeal squamous cell carcinoma based on microRNA expression signature. Br. J. Cancer 2014, 111, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Bus, P.; Siersema, P.D.; Verbeek, R.E.; van Baal, J.W. Upregulation of miRNA-143, -145, -192, and -194 in esophageal epithelial cells upon acidic bile salt stimulation. Dis. Esophagus 2014, 27, 591–600. [Google Scholar] [CrossRef]
- Li, J.; Wang, K.; Chen, X.; Meng, H.; Song, M.; Wang, Y.; Xu, X.; Bai, Y. Transcriptional activation of microRNA- 34a by NF-kappa B in human esophageal cancer cells. BMC Mol. Biol. 2012, 13, 4. [Google Scholar] [CrossRef] [Green Version]
- Li, H.P.; Zeng, X.C.; Zhang, B.; Long, J.T.; Zhou, B.; Tan, G.S.; Zeng, W.X.; Chen, W.; Yang, J.Y. miR-451 inhibits cell proliferation in human hepatocellular carcinoma through direct suppression of IKK-β. Carcinogenesis 2013, 34, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Li, H.; Yang, X.; Shao, J.; Jang, M.; Guan, D.; Zou, S.; Van Waes, C.; Chen, Z.; Zhan, M. Unraveling regulatory programs for NFkappaB, p53 and microRNAs in head and neck squamous cell carcinoma. PLoS ONE 2013, 8, e73656. [Google Scholar]
- Sasaki, C.T.; Doukas, S.G.; Costa, J.; Vageli, D.P. Biliary reflux as a causal factor in hypopharyngeal carcinoma: New clinical evidence and implications. Cancer 2019, 125, 3554–3565. [Google Scholar] [CrossRef]
- Alam, H.; Sehgal, L.; Kundu, S.T.; Dalal, S.N.; Vaidya, M.M. Novel function of keratins 5 and 14 in proliferation and differentiation of stratified epithelial cells. Mol. Biol. Cell 2011, 22, 4068–4078. [Google Scholar] [CrossRef]
- Choudhari, S.K.; Chaudhary, M.; Gadbail, A.R.; Sharma, A.; Tekade, S. Oxidative and antioxidative mechanisms in oral cancer and precancer: A review. Oral Oncol. 2014, 50, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.; Gu, M.; Takahata, T.; Frederick, B.; Agarwal, C.; Siriwardana, S.; Agarwal, R.; Sclafani, R.A. Resveratrol Selectively Induces DNA Damage, Independent of Smad4 Expression, in Its Efficacy against Human Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2011, 17, 5402–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in Context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, F.S.; Hsu, D.W.; Zeheb, R.; Efird, J.T.; Okunieff, P.G.; Malkin, D.M. Mutant, wild type, or overall p53 expression: Freedom from clinical progression in tumours of astrocytic lineage. Br. J. Cancer 2004, 91, 1678–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascal, J.M. The comings and goings of PARP-1 in response to DNA damage. DNA Repair 2018, 71, 177–182. [Google Scholar] [CrossRef]
- Martín-Oliva, D.; O’Valle, F.; Muñoz-Gámez, J.A.; Valenzuela, M.T.; Nuñez, M.I.; Aguilar, M.; Ruiz de Almodóvar, J.M.; Garcia del Moral, R.; Oliver, F.J. Crosstalk between PARP-1 and NF-kappaB modulates the promotion of skin neoplasia. Oncogene 2004, 23, 5275–5283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassa, P.O.; Hottiger, M.O. A role of poly (ADP-ribose) polymerase in NF-kappaB transcriptional activation. Biol Chem. 1999, 380, 953–959. [Google Scholar] [CrossRef]
- Hassa, P.O.; Covic, M.; Hasan, S.; Imhof, R.; Hottiger, M.O. The Enzymatic and DNA Binding Activity of PARP-1 Are Not Required for NF-κB Coactivator Function. J. Biol. Chem. 2001, 276, 45588–45597. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, A.; Knauer, S.K.; Stauber, R.H. Nuclear receptors in head and neck cancer: Current knowledge and perspectives. Int. J. Cancer 2010, 126, 801–809. [Google Scholar] [CrossRef]
- Kovács, P.; Csonka, T.; Kovács, T.; Sári, Z.; Ujlaki, G.; Sipos, A.; Karányi, Z.; Szeőcs, D.; Hegedűs, C.; Uray, K.; et al. Lithocholic Acid, a Metabolite of the Microbiome, Increases Oxidative Stress in Breast Cancer. Cancers 2019, 11, 1255. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.; Bernstein, C.; Payne, C.M.; Dvorak, K. Bile acids as endogenous etiologic agents in gastrointestinal cancer. World J. Gastroenterol. 2009, 15, 3329–3340. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.; Li, H.; Yang, Z.; Hoque, A.; Xu, X. Inhibition of farnesoid X receptor controls esophageal cancer cell growth in vitro and in nude mouse xenografts. Cancer 2013, 119, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Zheng, J.B.; Qi, J.; Yang, K.; Wu, Y.H.; Wang, K.; Wang, C.B.; Sun, X.J. Bile acids promote gastric intestinal metaplasia by upregulating CDX2 and MUC2 expression via the FXR/NF-κB signalling pathway. Int. J. Oncol. 2019, 54, 879–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauer, W.K.; Peters, J.H.; DeMeester, T.R.; Feussner, H.; Ireland, A.P.; Stein, H.J.; Siewert, R.J. Composition and concentration of bile acid reflux into the esophagus of patients with gastroesophageal reflux disease. Surgery 1997, 122, 874–881. [Google Scholar] [CrossRef]
- Fleskens, S.; Slootweg, P. Grading systems in head and neck dysplasia: Their prognostic value, weaknesses and utility (review). Head Neck Oncol. 2009, 1, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, N.; Blagus, R.; El-Mofty, S.K.; Helliwell, T.; Prasad, M.L.; Sandison, A.; Volavšek, M.; Wenig, B.M.; Zidar, N.; Cardesa, A. Evaluation of a new grading system for laryngeal squamous intraepithelial lesions-a proposed unified classification. Histopathology 2014, 65, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Conti, C.J.; Ward, J.M. Atlas of Laboratory Mouse Histology; Austion: Austin, TX, USA, 2004–2005.
- Sasaki, C.T.; Issaeva, N.; Vageli, D.P. In vitro model for gastroduodenal reflux-induced nuclear factor-kappaB activation and its role in hypopharyngeal carcinogenesis. Head Neck 2016, 38, E1381–E1391. [Google Scholar] [CrossRef] [PubMed]
- Doukas, S.G.; Cardoso, B.; Tower, J.I.; Vageli, D.P.; Sasaki, C.T. Biliary tumorigenic effect on hypopharyngeal cells is significantly enhanced by pH reduction. Cancer Med. 2019, 8, 4417–4427. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, C.T.; Doukas, S.G.; Costa, J.; Vageli, D.P. The Progressive Mutagenic Effects of Acidic Bile Refluxate in Hypopharyngeal Squamous Cell Carcinogenesis: New Insights. Cancers 2020, 12, 1064. https://doi.org/10.3390/cancers12051064
Sasaki CT, Doukas SG, Costa J, Vageli DP. The Progressive Mutagenic Effects of Acidic Bile Refluxate in Hypopharyngeal Squamous Cell Carcinogenesis: New Insights. Cancers. 2020; 12(5):1064. https://doi.org/10.3390/cancers12051064
Chicago/Turabian StyleSasaki, Clarence T., Sotirios G. Doukas, Jose Costa, and Dimitra P. Vageli. 2020. "The Progressive Mutagenic Effects of Acidic Bile Refluxate in Hypopharyngeal Squamous Cell Carcinogenesis: New Insights" Cancers 12, no. 5: 1064. https://doi.org/10.3390/cancers12051064