The Roles of ROS Generation in RANKL-Induced Osteoclastogenesis: Suppressive Effects of Febuxostat

 ,
,  , and
, and 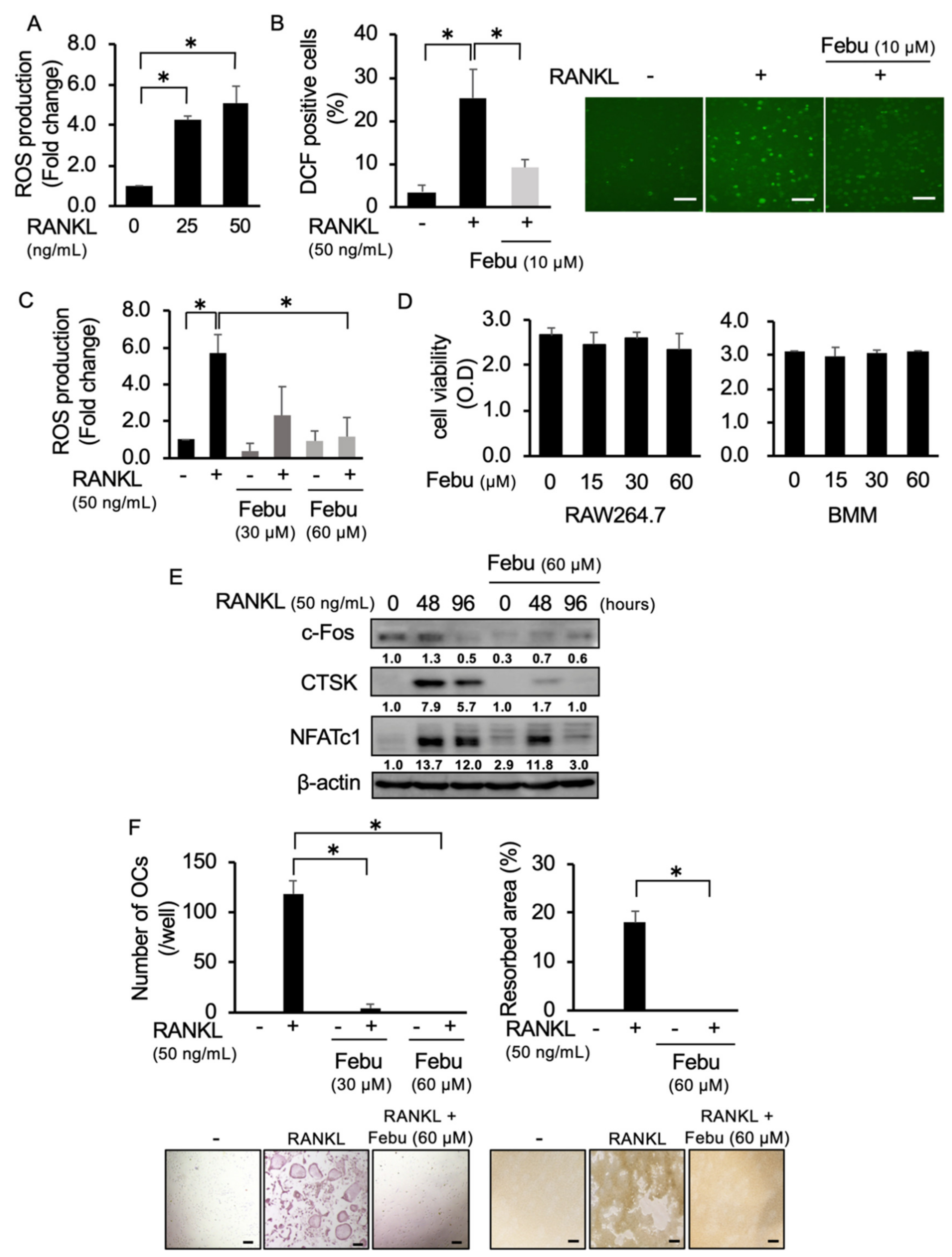{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Febuxostat Inhibits RANKL-Induced ROS Production and OC Formation
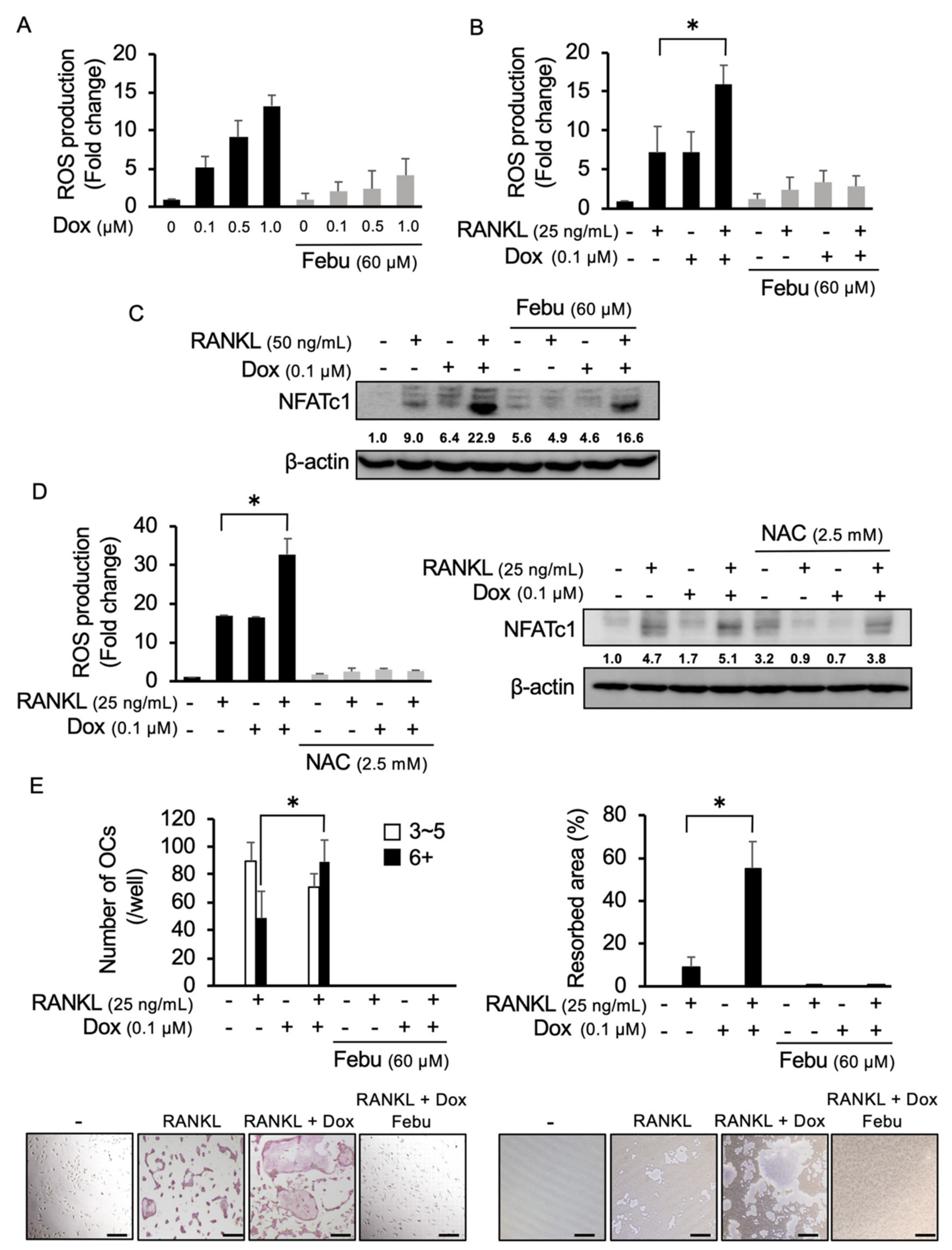
2.2. Dox Facilitates RANKL-Mediated Osteoclastogenesis Through ROS Production
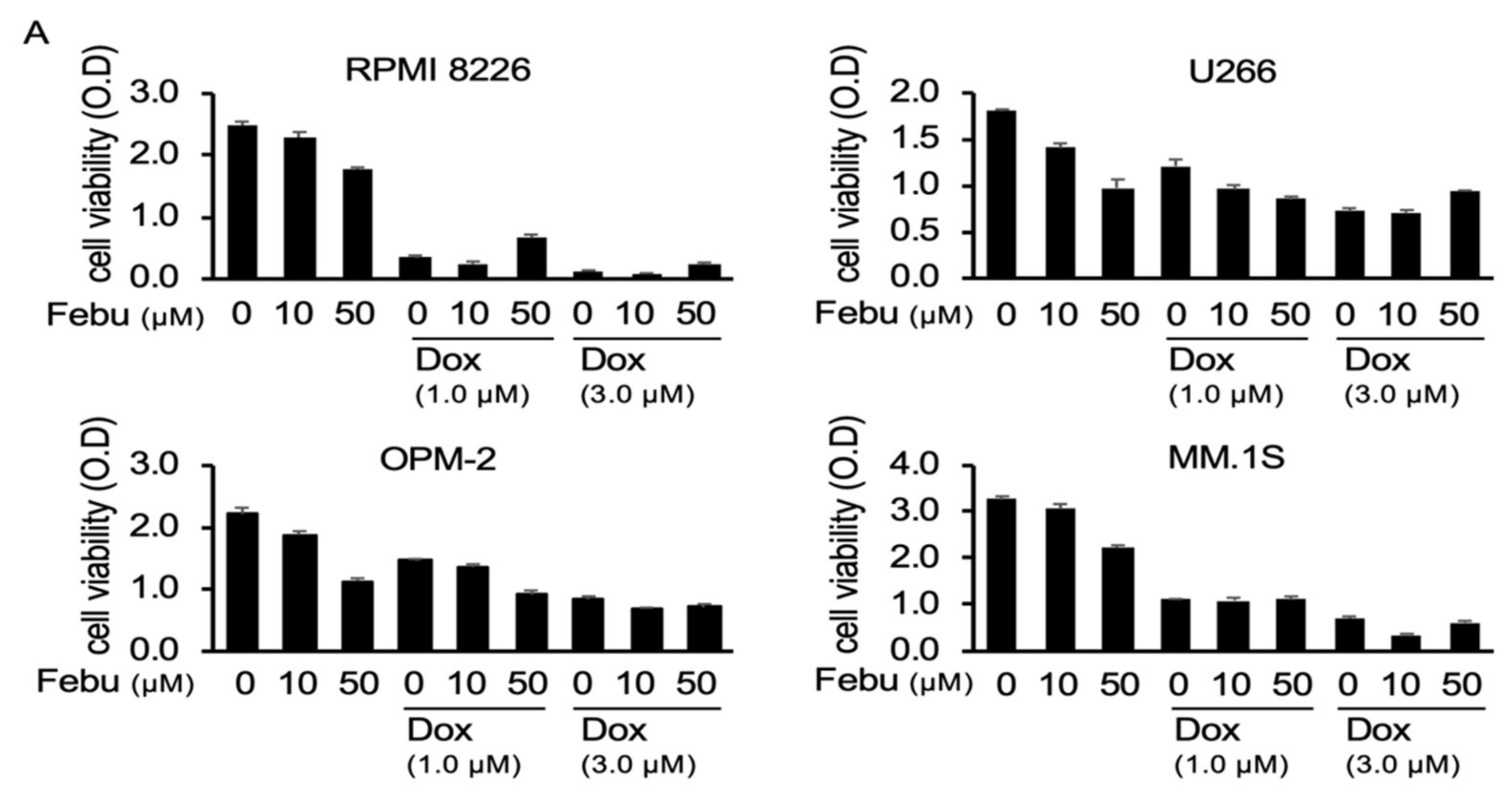
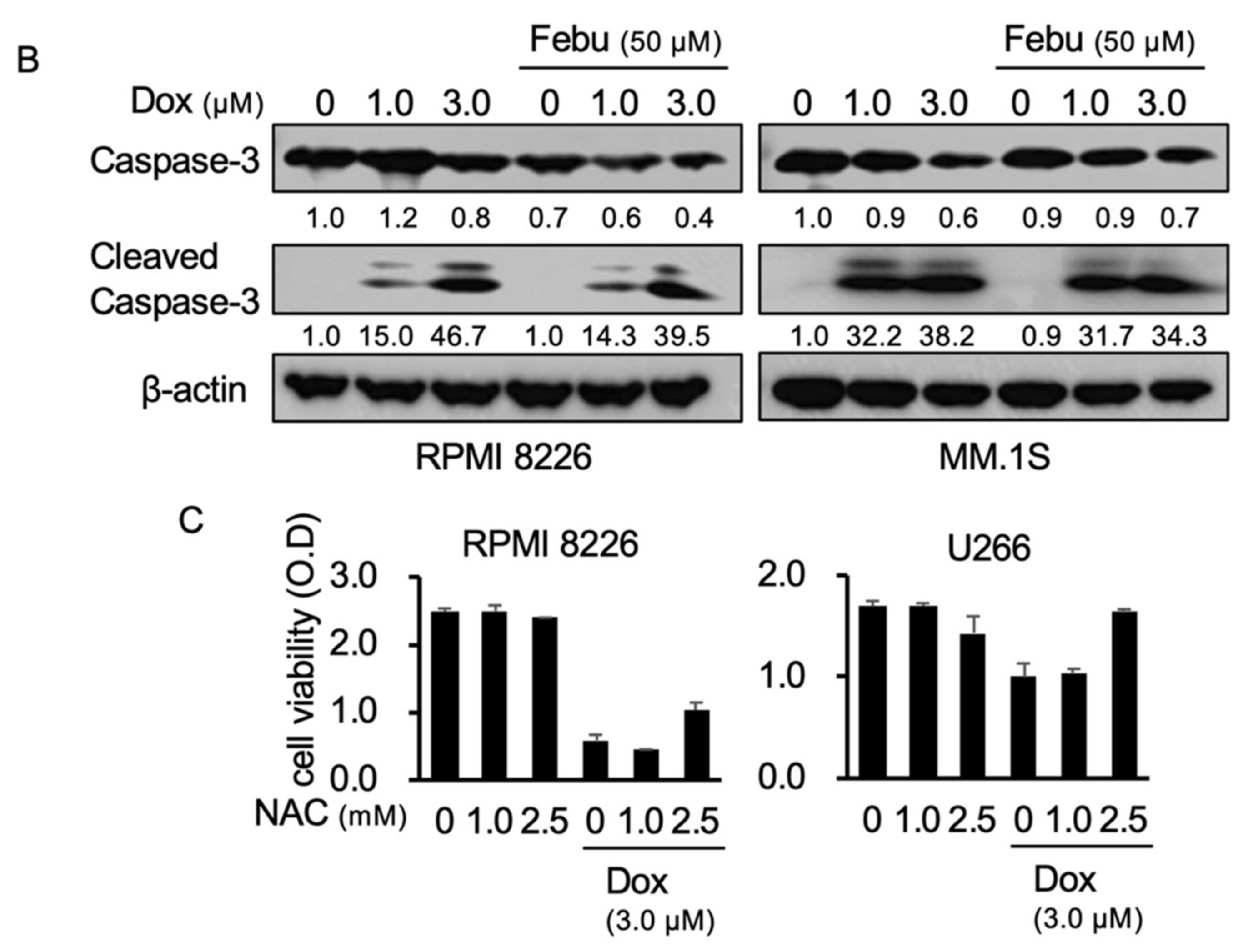
2.3. Febuxostat Does Not Compromise the Cytotoxic Effects of Dox on MM Cells
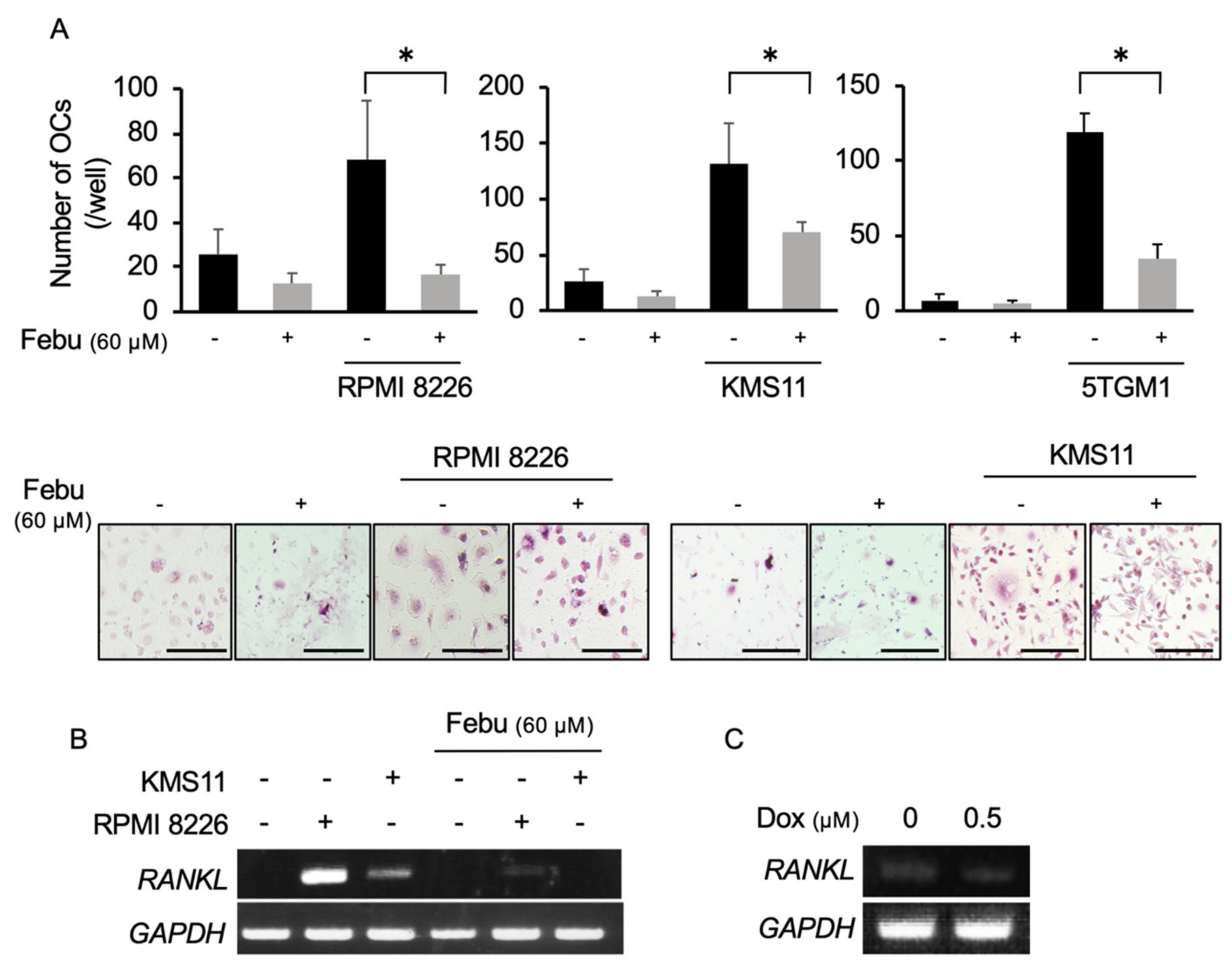
2.4. Febuxostat Suppresses Osteoclastogenesis by MM Cells
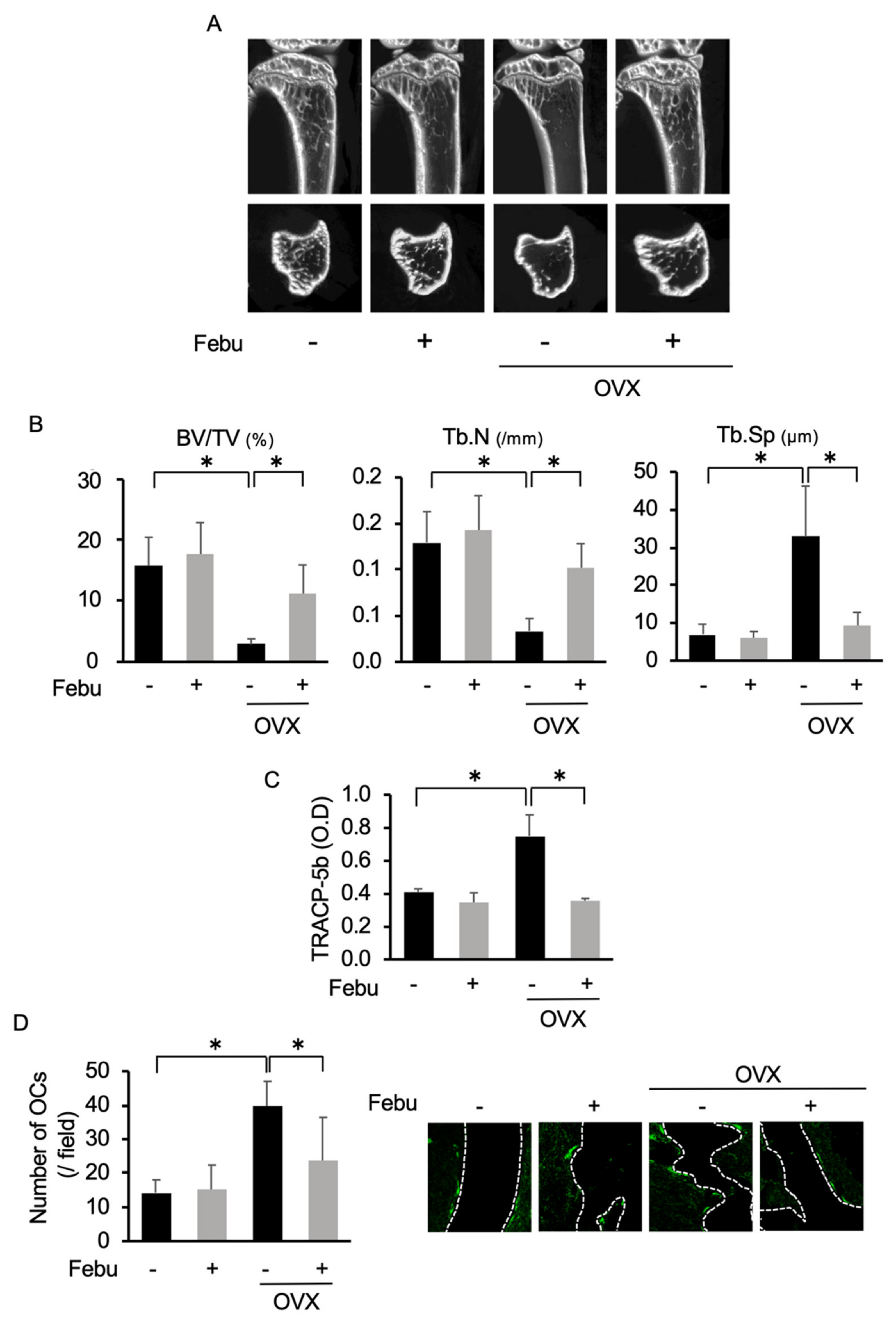
2.5. Febuxostat Reduces Osteoclastic Activity and Bone Loss in Ovariectomized (OVX) Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. OC Formation
4.4. Bone Resorption Assay
4.5. Coculture Experiment
4.6. Animal Experiment
4.7. Quantifying ROS
4.8. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Real-time PCR
4.9. Cell Viability
4.10. Western Blot
4.11. Actin Ring Staining
4.12. Immunofluorescence Staining
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Xu, J.; Li, H.; Zheng, Y.; He, J.; Liu, H.; Zhong, Y.; Lu, Y.; Hong, B.; Zhang, M. Bone marrow stromal cells derived MCP-1 reverses the inhibitory effects of multiple myeloma cells on osteoclastogenesis by upregulating the RANK expression. PLoS ONE. 2013, 8, e82453. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Gavriatopoulou, M.; Dimopoulos, M.A. Pathogenesis of bone disease in multiple myeloma: From bench to bedside. Blood Cancer J. 2018, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Abe, M. Bone destruction in multiple myeloma. Ann. N. Y. Acad. Sci. 2006, 1068, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, N.; Colla, S.; Morandi, F.V.R. The RANK/RANK ligand system is involved in interleukin-6 and interleukin-11 up-regulation by human myeloma cells in the bone marrow microenvironment. Hematologica 2004, 89, 1118–1123. [Google Scholar]
- Abe, M.; Hiura, K.; Wilde, J.; Shioyasono, A.; Moriyama, K.; Hashimoto, T.; Kido, S.; Oshima, T.; Shibata, H.; Ozaki, S.; et al. Osteoclasts enhance myeloma cell growth and survival via cell-cell contact: A vicious cycle between bone destruction and myeloma expansion. Blood 2004, 104, 2484–2491. [Google Scholar] [CrossRef]
- Roodman, G.D.W.C.D. RANK ligand as a therapeutic target for bone metastases and multiple myeloma. Cancer Treat. Rev. 2008, 34, 92–101. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Mitra, S.; Nguyen, L.N.; Akter, M.; Park, G.; Choi, E.H.; Kaushik, N.K. Impact of ROS Generated by Chemical, Physical, and Plasma Techniques on Cancer Attenuation. Cancers. 2019, 11, 1030. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell. Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Agidigbi, T.S.; Kim, C. Reactive Oxygen Species in Osteoclast Differentiation and Possible Pharmaceutical Targets of ROS-Mediated Osteoclast Diseases. Int. J. Mol. Sci. 2019, 20, 3576. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, D.A.; Jiang, J.X. Reactive oxygen species and oxidative stress in osteoclastogenesis, skeletal aging and bone diseases. J. Bone Miner. Metab. 2015, 33, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell. Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, S.; Koenigstein, A.; Joseph, J.; Sun, L.; Kalyanaraman, B.; Zaidi, M.; Avadhani, N.G. Role of mitochondrial reactive oxygen species in osteoclast differentiation. Ann. N. Y. Acad. Sci. 2010, 1192, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Asensio-Lopez, M.C.; Soler, F.; Pascual-Figal, D.; Fernandez-Belda, F.; Lax, A. Doxorubicin-induced oxidative stress: The protective effect of nicorandil on HL-1 cardiomyocytes. PLoS ONE 2017, 12, e0172803. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Kim, S.J.; Kim, B.J.; Rah, S.Y.; Chung, S.M.; Im, M.J.; Kim, U.H. Doxorubicin-induced reactive oxygen species generation and intracellular Ca2+ increase are reciprocally modulated in rat cardiomyocytes. Exp. Mol. Med. 2006, 38, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Rana, T.; Chakrabarti, A.; Freeman, M.; Biswas, S. Doxorubicin-mediated bone loss in breast cancer bone metastases is driven by an interplay between oxidative stress and induction of TGFbeta. PLoS ONE 2013, 8, e78043. [Google Scholar] [CrossRef]
- Takai, M.; Yamauchi, T.; Fujita, K.; Lee, S.; Ookura, M.; Kishi, S.; Urasaki, Y.; Yoshida, A.; Iwasaki, H.; Ueda, T. Controlling serum uric acid using febuxostat in cancer patients at risk of tumor lysis syndrome. Oncol. Lett. 2014, 8, 1523–1527. [Google Scholar] [CrossRef] [Green Version]
- Nomura, J.; Busso, N.; Ives, A.; Tsujimoto, S.; Tamura, M.; So, A.; Yamanaka, Y. Febuxostat, an inhibitor of xanthine oxidase, suppresses lipopolysaccharide-induced MCP-1 production via MAPK phosphatase-1-mediated inactivation of JNK. PLoS ONE 2013, 8, e75527. [Google Scholar] [CrossRef]
- Yisireyili, M.; Hayashi, M.; Wu, H.; Uchida, Y.; Yamamoto, K.; Kikuchi, R.; Shoaib Hamrah, M.; Nakayama, T.; Wu Cheng, X.; Matsushita, T.; et al. Xanthine oxidase inhibition by febuxostat attenuates stress-induced hyperuricemia, glucose dysmetabolism, and prothrombotic state in mice. Sci. Rep. 2017, 7, 1266. [Google Scholar] [CrossRef]
- Kim, H.; Lee, Y.D.; Kim, H.J.; Lee, Z.H.; Kim, H.H. SOD2 and Sirt3 Control Osteoclastogenesis by Regulating Mitochondrial ROS. J. Bone Miner. Res. 2017, 32, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.Y.H.; Li, Z.; Jones, M.M.; Yang, S.; Li, C.; Fu, C.; Tu, C.; Oursler, M.J.; Qu, J.; Yang, S. Regulator of G protein signaling 12 enhances osteoclastogenesis by suppressing Nrf2-dependent antioxidant proteins to promote the generation of reactive oxygen species. Elife 2019, 8, e42951. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Z.; Maimon, Y.; Samuels, N.; Berger, R. Role of reactive oxygen species in the anticancer activity of botanicals: Comparing sensitivity profiles. Oncol. Lett. 2017, 13, 2642–2648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ri, M. Mechanism of action of bortezomib in multiple myeloma therapy. Int. J. Myeloma 2016, 6, 1–6. [Google Scholar]
- Hussein, M. Pegylated liposomal doxorubicin, vincristine, and reduced-dose dexamethasone as first-line therapy for multiple myeloma. Clin. Lymphoma. 2003, 4, S18–S22. [Google Scholar] [CrossRef] [PubMed]
- Fatfat, M.; Fakhoury, I.; Habli, Z.; Mismar, R.; HalaGali, M. Thymoquinone enhances the anticancer activity of doxorubicin against adult T-cell leukemia in vitro and in vivo through ROS-dependent mechanisms. Life Sci. 2019, 232, 116628. [Google Scholar] [CrossRef]
- Teramachi, J.; Hiasa, M.; Oda, A.; Harada, T.; Nakamura, S.; Amachi, R.; Tenshin, H.; Iwasa, M.; Fujii, S.; Kagawa, K.; et al. Pim-2 is a critical target for treatment of osteoclastogenesis enhanced in myeloma. Br. J. Haematol. 2018, 180, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Hiasa, M.; Teramachi, J.; Oda, A.; Amachi, R.; Harada, T.; Nakamura, S.; Miki, H.; Fujii, S.; Kagawa, K.; Watanabe, K.; et al. Pim-2 kinase is an important target of treatment for tumor progression and bone loss in myeloma. Leukemia 2015, 29, 207–217. [Google Scholar] [CrossRef]
- Giuliani, N.; Colla, S.; Rizzoli, V. New insight in the mechanism of osteoclast activation and formation in multiple myeloma: Focus on the receptor activator of NF-κB ligand (RANKL). Exp. Hematol. 2004, 32, 685–691. [Google Scholar] [CrossRef]
- Muthusami, S.; Ramachandran, I.; Muthusamy, B.; Vasudevan, G.; Prabhu, V.; Subramaniam, V.; Jagadeesan, A.; Narasimhan, S. Ovariectomy induces oxidative stress and impairs bone antioxidant system in adult rats. Clin. Chim. Acta. 2005, 360, 81–86. [Google Scholar] [CrossRef]
- Li, H.; Huang, C.; Zhu, J.; Gao, K.; Fang, J.; Li, H. Lutein Suppresses Oxidative Stress and Inflammation by Nrf2 Activation in an Osteoporosis Rat Model. Med. Sci. Monit. 2018, 24, 5071–5075. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Gu, Y.; Tan, X.; Deng, Y.; Wu, Z.T.; Xu, M.J.; Wang, W.Z. Estrogen Replacement Reduces Oxidative Stress in the Rostral Ventrolateral Medulla of Ovariectomized Rats. Oxid. Med. Cell. Longev. 2016, 2016, 2158971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, M.A.; Schumacher, H.R.; Espinoza, L.R.; Wells, A.F.; MacDonald, P.; Lloyd, E.; Lademacher, C. The urate-lowering efficacy and safety of febuxostat in the treatment of the hyperuricemia of gout: The CONFIRMS trial. Arthritis Res. Ther. 2010, 12, R63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Kawai, Y.; Kiguchi, T.; Okamoto, M.; Kaneko, M.; Maemondo, M.; Gemba, K.; Fujimaki, K.; Kirito, K.; Goto, T.; et al. Efficacy and safety of febuxostat for prevention of tumor lysis syndrome in patients with malignant tumors receiving chemotherapy: A phase III, randomized, multi-center trial comparing febuxostat and allopurinol. Int. J. Clin. Oncol. 2016, 21, 996–1003. [Google Scholar] [CrossRef]
- Bellos, I.; Kontzoglou, K.; Psyrri, A.; Pergialiotis, V. Febuxostat administration for the prevention of tumour lysis syndrome: A meta-analysis. J. Clin. Pharm. Ther. 2019, 44, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- D’Oronzo, S.; Stucci, S.; Tucci, M.; Silvestris, F. Cancer treatment-induced bone loss (CTIBL): Pathogenesis and clinical implications. Cancer Treat. Rev. 2015, 41, 798–808. [Google Scholar] [CrossRef]
- Guenther, G.; Steger, R.B. Denosumab for the treatment of bone metastases in breast cancer: Evidence and opinion. Ther. Adv. Med. Oncol. 2011, 3, 233–243. [Google Scholar]
- Gnant, M.; Blaha, P.; Dubsky, P.; Exner, R.; Fitzal, F.; Sporn, E.; Panhofer, P.; Borgo, A.D.; Bigenzahn, S.; Steger, G.; et al. Adjuvant bisphosphonates in endocrine-responsive breast cancer: What is their place in therapy? Ther. Adv. Med. Oncol. 2009, 1, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Tenshin, H.; Teramachi, J.; Oda, A.; Amachi, R.; Hiasa, M.; Bat-Erdene, A.; Watanabe, K.; Iwasa, M.; Harada, T.; Fujii, S.; et al. TAK1 inhibition subverts the osteoclastogenic action of TRAIL while potentiating its antimyeloma effects. Blood advances 2017, 1, 2124–2137. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, H.; Teramachi, J.; Okamura, H.; Yang, D.; Nagata, T.; Haneji, T. Double Stranded RNA-Dependent Protein Kinase is Necessary for TNF-alpha-Induced Osteoclast Formation In Vitro and In Vivo. J. Cell. Biochem. 2015, 116, 1957–1967. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, C.; Wang, G.; Sun, Y.; Deng, Z.; Chen, L.; Chen, K.; Tickner, J.; Kenny, J.; Song, D.; et al. Loureirin B suppresses RANKL-induced osteoclastogenesis and ovariectomized osteoporosis via attenuating NFATc1 and ROS activities. Theranostics 2019, 9, 4648–4662. [Google Scholar] [CrossRef] [PubMed]
- Bouxsein, M.L.; Myers, K.S.; Shultz, K.L.; Donahue, L.R.; Rosen, C.J.; Beamer, W.G. Ovariectomy-induced bone loss varies among inbred strains of mice. J. Bone Miner. Res. 2005, 20, 1085–1092. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashtar, M.; Tenshin, H.; Teramachi, J.; Bat-Erdene, A.; Hiasa, M.; Oda, A.; Tanimoto, K.; Shimizu, S.; Higa, Y.; Harada, T.; et al. The Roles of ROS Generation in RANKL-Induced Osteoclastogenesis: Suppressive Effects of Febuxostat. Cancers 2020, 12, 929. https://doi.org/10.3390/cancers12040929
Ashtar M, Tenshin H, Teramachi J, Bat-Erdene A, Hiasa M, Oda A, Tanimoto K, Shimizu S, Higa Y, Harada T, et al. The Roles of ROS Generation in RANKL-Induced Osteoclastogenesis: Suppressive Effects of Febuxostat. Cancers. 2020; 12(4):929. https://doi.org/10.3390/cancers12040929
Chicago/Turabian StyleAshtar, Mohannad, Hirofumi Tenshin, Jumpei Teramachi, Ariunzaya Bat-Erdene, Masahiro Hiasa, Asuka Oda, Kotaro Tanimoto, So Shimizu, Yoshiki Higa, Takeshi Harada, and et al. 2020. "The Roles of ROS Generation in RANKL-Induced Osteoclastogenesis: Suppressive Effects of Febuxostat" Cancers 12, no. 4: 929. https://doi.org/10.3390/cancers12040929