Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets



Abstract
:Simple Summary
Abstract
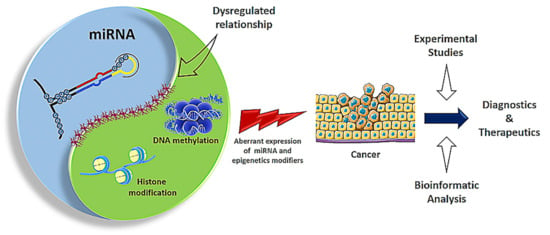
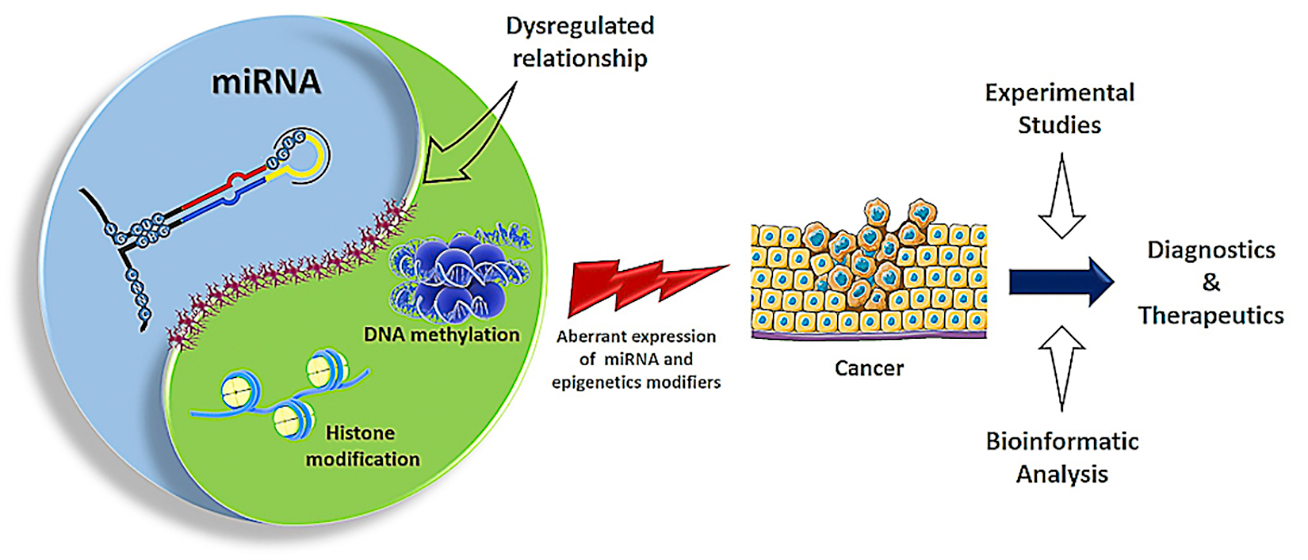
1. Introduction
2. Epigenetic Regulators of Cancer
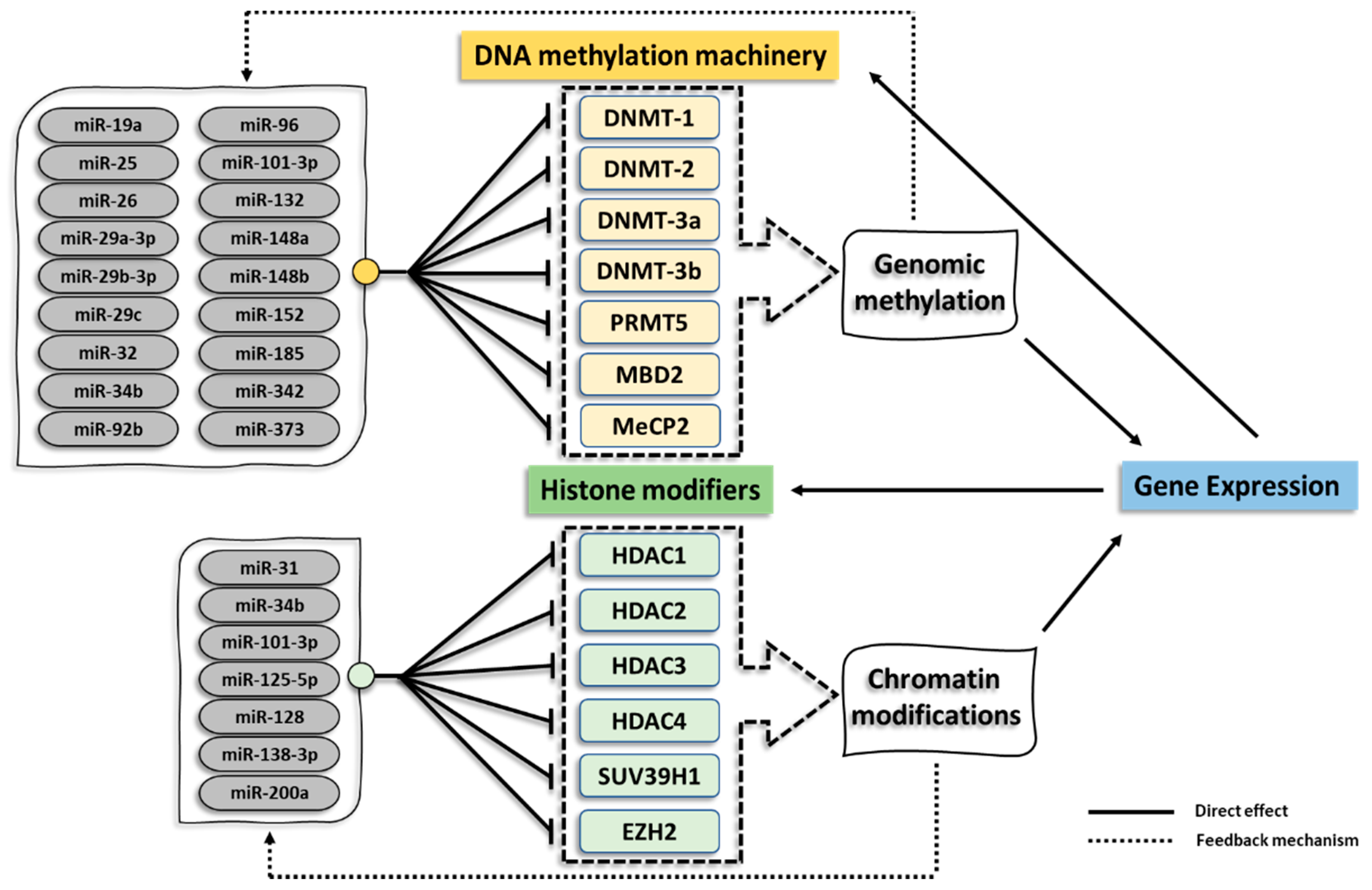
2.1. DNA Methylation and miRNA Regulation
2.2. Histone Acetylation and Deacetylation
3. Epigenetic Regulation by miRNAs in Cancer
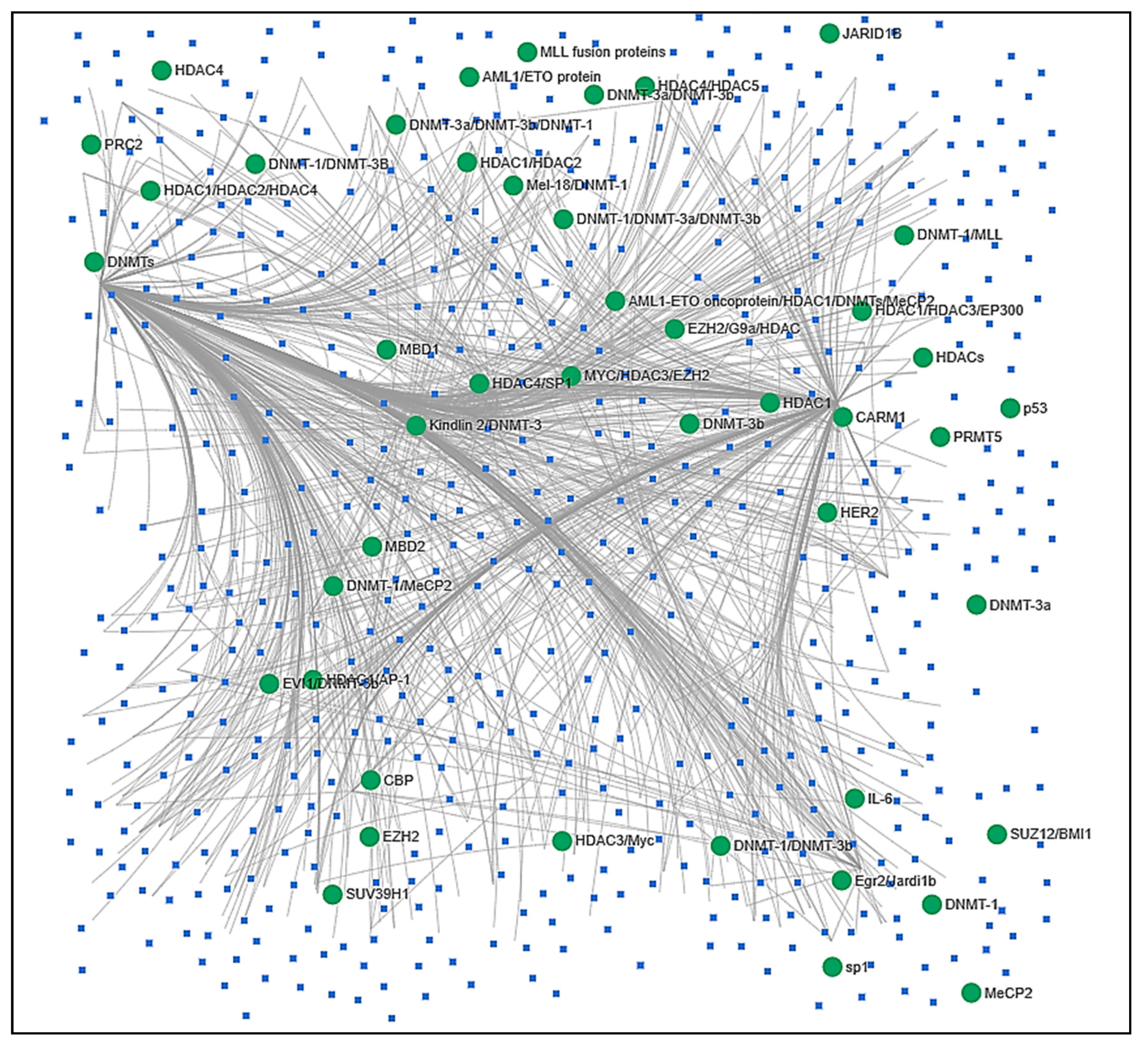
4. In Silico Analysis of Epi-miRNA Associations
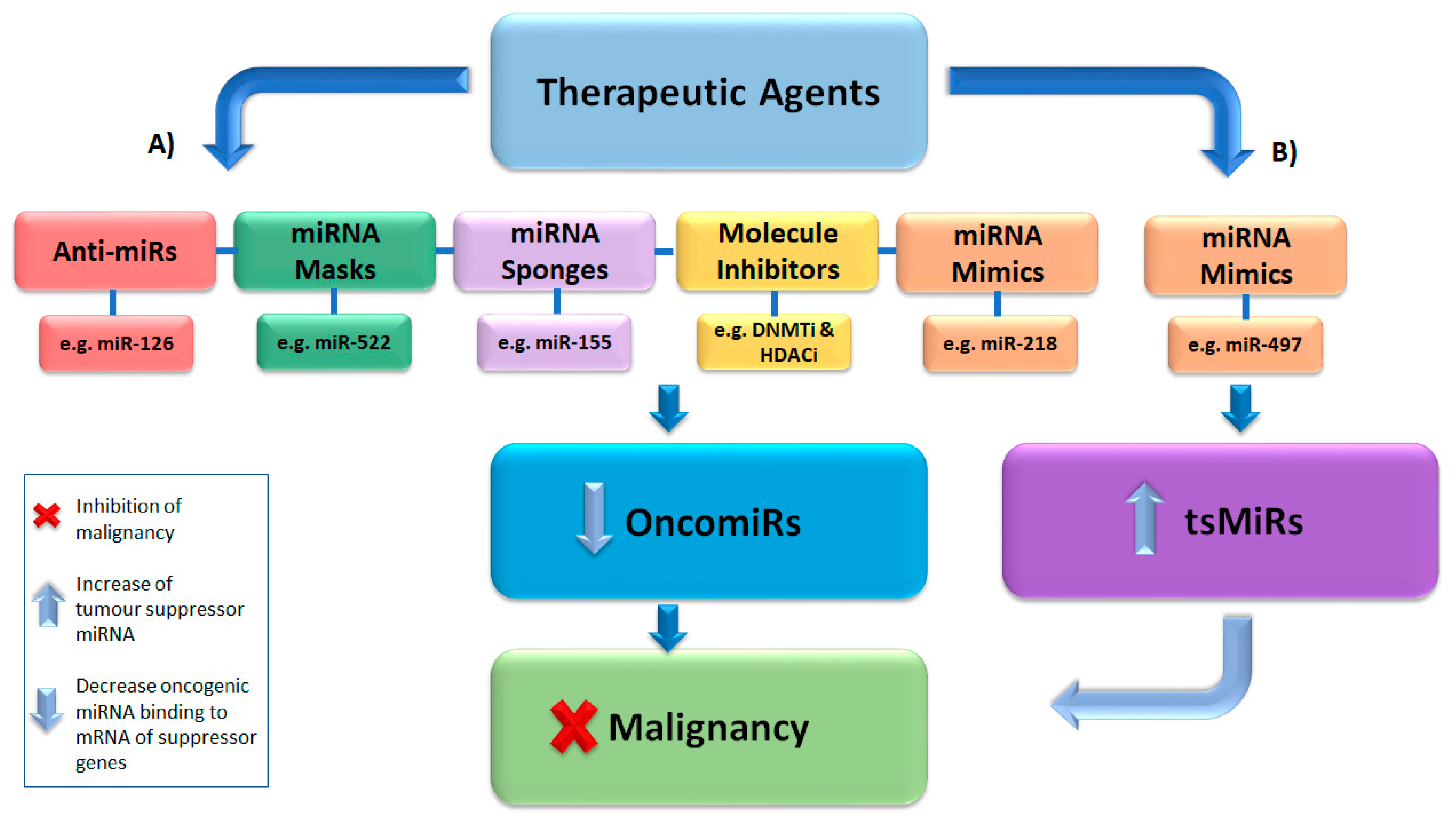
5. Epigenetic Strategies for Cancer Therapy
5.1. miRNA Mimics and Inhibitors
5.2. miRNA Sponges
5.3. miRNA Masking
5.4. Epigenetic Inhibitory Molecules
6. Epigenetic Therapeutics in Cancer Clinical Trials
7. Overcoming Limitations of miRNA Biomarkers and Therapeutic Agents
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- miRBase miRNA. Available online: http://www.mirbase.org/ (accessed on 1 August 2020).
- Di Lisio, L.; Sanchez-Beato, M.; Gomez-Lopez, G.; Rodriguez, M.E.; Montes-Moreno, S.; Mollejo, M.; Menarguez, J.; Martinez, M.A.; Alves, F.J.; Pisano, D.G.; et al. MicroRNA signatures in B-cell lymphomas. Blood Cancer J. 2012, 2, e57. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.Q.; Ahmed, E.I.; Elareer, N.R.; Junejo, K.; Steinhoff, M.; Uddin, S. Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies. Cells 2019, 8, 840. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Humphries, B.; Wang, Z.; Yang, C. MicroRNA Regulation of Epigenetic Modifiers in Breast Cancer. Cancers 2019, 11, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorio, M.V.; Piovan, C.; Croce, C.M. Interplay between microRNAs and the epigenetic machinery: An intricate network. Biochim. Biophys. Acta 2010, 1799, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Liu, X.; Li, D.; Wang, P.; Li, N.; Lu, L.; Cao, X. MicroRNA-466l upregulates IL-10 expression in TLR-triggered macrophages by antagonizing RNA-binding protein tristetraprolin-mediated IL-10 mRNA degradation. J. Immunol. 2010, 184, 6053–6059. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abba, M.L.; Patil, N.; Leupold, J.H.; Moniuszko, M.; Utikal, J.; Niklinski, J.; Allgayer, H. MicroRNAs as novel targets and tools in cancer therapy. Cancer Lett. 2017, 387, 84–94. [Google Scholar] [CrossRef]
- Guo, J.; Miao, Y.; Xiao, B.; Huan, R.; Jiang, Z.; Meng, D.; Wang, Y. Differential expression of microRNA species in human gastric cancer versus non-tumorous tissues. J. Gastroenterol. Hepatol. 2009, 24, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Getaneh, Z.; Asrie, F.; Melku, M. MicroRNA profiles in B-cell non-Hodgkin lymphoma. EJIFCC 2019, 30, 195–214. [Google Scholar] [PubMed]
- Bradshaw, G.; Sutherland, H.G.; Haupt, L.M.; Griffiths, L.R. Dysregulated MicroRNA Expression Profiles and Potential Cellular, Circulating and Polymorphic Biomarkers in Non-Hodgkin Lymphoma. Genes 2016, 7, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagawa, H.; Ikeda, S.; Sawada, K. Role of microRNA in the pathogenesis of malignant lymphoma. Cancer Sci. 2013, 104, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Caramuta, S.; Lee, L.; Ozata, D.M.; Akcakaya, P.; Georgii-Hemming, P.; Xie, H.; Amini, R.M.; Lawrie, C.H.; Enblad, G.; Larsson, C.; et al. Role of microRNAs and microRNA machinery in the pathogenesis of diffuse large B-cell lymphoma. Blood Cancer J. 2013, 3, e152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taslim, C.; Weng, D.Y.; Brasky, T.M.; Dumitrescu, R.G.; Huang, K.; Kallakury, B.V.; Krishnan, S.; Llanos, A.A.; Marian, C.; McElroy, J.; et al. Discovery and replication of microRNAs for breast cancer risk using genome-wide profiling. Oncotarget 2016, 7, 86457–86468. [Google Scholar] [CrossRef]
- Hwang, H.W.; Wentzel, E.A.; Mendell, J.T. Cell-cell contact globally activates microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 7016–7021. [Google Scholar] [CrossRef] [Green Version]
- Fuso, A.; Raia, T.; Orticello, M.; Lucarelli, M. The complex interplay between DNA methylation and miRNAs in gene expression regulation. Biochimie 2020, 173, 12–16. [Google Scholar] [CrossRef]
- Suzuki, H.; Takatsuka, S.; Akashi, H.; Yamamoto, E.; Nojima, M.; Maruyama, R.; Kai, M.; Yamano, H.O.; Sasaki, Y.; Tokino, T.; et al. Genome-wide profiling of chromatin signatures reveals epigenetic regulation of MicroRNA genes in colorectal cancer. Cancer Res. 2011, 71, 5646–5658. [Google Scholar] [CrossRef] [Green Version]
- Abba, M.; Patil, N.; Leupold, J.H.; Allgayer, H. MicroRNAs-from metastasis prediction to metastasis prevention? Mol. Cell. Oncol. 2016, 3, e1074336. [Google Scholar] [CrossRef] [Green Version]
- Ebert, M.S.; Sharp, P.A. MicroRNA sponges: Progress and possibilities. RNA 2010, 16, 2043–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Humphries, B.; Yang, C.; Wang, Z. Nanoparticle-Mediated Therapeutic Agent Delivery for Treating Metastatic Breast Cancer-Challenges and Opportunities. Nanomaterials 2018, 8, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, E.; Yu, X.; Zhang, Y.; Meng, F.; Wang, S.; Liu, X.; Liu, D.; Wang, J.; Li, X.; Jiang, W. EpimiR: A database of curated mutual regulation between miRNAs and epigenetic modifications. Database 2014, 2014, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Siklenka, K.; Arora, S.K.; Ribeiro, P.; Kimmins, S.; Xia, J. miRNet—Dissecting miRNA-target interactions and functional associations through network-based visual analysis. Nucleic Acids Res. 2016, 44, W135–W141. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Liu, B.; Qu, S.; Liang, G.; Luo, W.; Gong, C. MicroRNAs and cancer: Key paradigms in molecular therapy (Review). Oncol. Lett. 2018, 15, 2735–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. Epigenetic alteration and microRNA dysregulation in cancer. Front. Genet. 2013, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Coukos, G.; Zhang, L. MicroRNA epigenetic alterations in human cancer: One step forward in diagnosis and treatment. Int. J. Cancer 2008, 122, 963–968. [Google Scholar] [CrossRef]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. DNA methylation and microRNA dysregulation in cancer. Mol. Oncol. 2012, 6, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Morales, S.; Monzo, M.; Navarro, A. Epigenetic regulation mechanisms of microRNA expression. Biomol. Concepts 2017, 8, 203–212. [Google Scholar] [CrossRef]
- Cora, D.; Re, A.; Caselle, M.; Bussolino, F. MicroRNA-mediated regulatory circuits: Outlook and perspectives. Phys. Biol. 2017, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osella, M.; Riba, A.; Testori, A.; Corá, D.; Caselle, M. Interplay of microRNA and epigenetic regulation in the human regulatory network. Front. Genet. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malumbres, M. MiRNAs and cancer: An epigenetics view. Mol. Aspects Med. 2013, 34, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Sato, F.; Tsuchiya, S.; Meltzer, S.J.; Shimizu, K. MicroRNAs and epigenetics. FEBS J. 2011, 278, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Pogribny, I.P.; Filkowski, J.N.; Tryndyak, V.P.; Golubov, A.; Shpyleva, S.I.; Kovalchuk, O. Alterations of microRNAs and their targets are associated with acquired resistance of MCF-7 breast cancer cells to cisplatin. Int. J. Cancer 2010, 127, 1785–1794. [Google Scholar] [CrossRef]
- Garzon, R.; Liu, S.; Fabbri, M.; Liu, Z.; Heaphy, C.E.A.; Callegari, E.; Schwind, S.; Pang, J.; Yu, J.; Muthusamy, N.; et al. MicroRNA-29b induces global DNA hypomethylation and tumor suppressor gene reexpression in acute myeloid leukemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood 2009, 113, 6411–6418. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.M.; Liang, G.; Liu, C.C.; Wolff, E.M.; Tsai, Y.C.; Ye, W.; Zhou, X.; Jones, P.A. The putative tumor suppressor microRNA-101 modulates the cancer epigenome by repressing the polycomb group protein EZH2. Cancer Res. 2009, 69, 2623–2629. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Zhang, B.; Wu, T.; Skogerbø, G.; Zhu, X.; Guo, X.; He, S.; Chen, R. Transcriptional inhibiton of Hoxd4 expression by miRNA-10a in human breast cancer cells. BMC Mol. Biol. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, R.; Rivenbark, A.G.; Coleman, W.B. Loss of post-transcriptional regulation of DNMT3b by microRNAs: A possible molecular mechanism for the hypermethylation defect observed in a subset of breast cancer cell lines. Int. J. Oncol. 2012, 41, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Jiang, Y.; Yin, Y.; Li, Q.; He, J.; Jing, Y.; Qi, Y.T.; Xu, Q.; Li, W.; Lu, B.; et al. A regulatory circuit of miR-148a/152 and DNMT1 in modulating cell transformation and tumor angiogenesis through IGF-IR and IRS1. J. Mol. Cell Biol. 2013, 5, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Baritaki, S.; Marathe, H.; Feng, J.; Park, S.; Beach, S.; Bazeley, P.S.; Beshir, A.B.; Fenteany, G.; Mehra, R.; et al. Polycomb protein EZH2 regulates tumor invasion via the transcriptional repression of the metastasis suppressor RKIP in breast and prostate cancer. Cancer Res. 2012, 72, 3091–3104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, E.K.O.; Tsang, W.P.; Ng, S.S.M.; Jin, H.C.; Yu, J.; Li, J.J.; Röcken, C.; Ebert, M.P.A.; Kwok, T.T.; Sung, J.J.Y. MicroRNA-143 targets DNA methyltransferases 3A in colorectal cancer. Br. J. Cancer 2009, 101, 699–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wu, J.; Meng, X.; Ying, X.; Zuo, Y.; Liu, R.; Pan, Z.; Kang, T.; Huang, W. Microrna-342 inhibits colorectal cancer cell proliferation and invasion by directly targeting dna methyltransferase 1. Carcinogenesis 2011, 32, 1033–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Kuo, C.; Nicholl, M.B.; Sim, M.S.; Turner, R.R.; Morton, D.L.; Hoon, D.S.B. Downregulation of microRNA-29c is associated with hypermethylation of tumor-related genes and disease outcome in cutaneous melanoma. Epigenetics 2011, 6, 388–394. [Google Scholar] [CrossRef]
- Tsuruta, T.; Kozaki, K.I.; Uesugi, A.; Furuta, M.; Hirasawa, A.; Imoto, I.; Susumu, N.; Aoki, D.; Inazawa, J. miR-152 is a tumor suppressor microRNA that is silenced by DNA hypermethylation in endometrial cancer. Cancer Res. 2011, 71, 6450–6462. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Xia, J.; Zuo, J.; Jin, S.; Zhou, H.; Yao, L.; Huang, H.; Han, Z. MicroRNA-148a is silenced by hypermethylation and interacts with DNA methyltransferase 1 in gastric cancer. Med. Oncol. 2012, 29, 2701–2709. [Google Scholar] [CrossRef]
- Smits, M.; Nilsson, J.; Mir, S.E.; Van Der Stoop, P.M.; Hulleman, E.; Niers, J.M.; Hamer, P.C.D.W.; Marquez, V.E.; Cloos, J.; Krichevsky, A.M.; et al. miR-101 is down-regulated in glioblastoma resulting in EZH2- induced proliferation, migration, and angiogenesis AbstrAct: Background: Glioblastoma (GBM) is a malignant brain tumor with dismal. Oncotarget 2010, 1, 710–720. [Google Scholar] [CrossRef] [Green Version]
- Smits, M.; Mir, S.E.; Nilsson, J.R.A.; van der Stoop, P.M.; Niers, J.M.; Marquez, V.E.; Cloos, J.; Breakefield, X.O.; Krichevsky, A.M.; Noske, D.P.; et al. Down-regulation of miR-101 in endothelial cells promotes blood vessel formation through reduced repression of EZH2. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, H.; Wang, Z.; Zhang, B.; Liu, W.; Lu, H.; Xiao, L.; Liu, X.; Wang, R.; Li, X.; et al. MiR-185 Targets the DNA Methyltransferases 1 and Regulates Global DNA Methylation in human glioma. Mol. Cancer 2011, 10, 124. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Xiang, T.; Ren, G.; Tan, C.; Liu, R.; Xu, X.; Wu, Z. MiR-101 is down-regulated by the hepatitis B virus x protein and induces aberrant DNA methylation by targeting DNA methyltransferase 3A. Cell. Signal. 2013, 25, 439–446. [Google Scholar] [CrossRef]
- Yuan, J.H.; Yang, F.; Chen, B.F.; Lu, Z.; Huo, X.S.; Zhou, W.P.; Wang, F.; Sun, S.H. The histone deacetylase 4/SP1/microrna-200a regulatory network contributes to aberrant histone acetylation in hepatocellular carcinoma. Hepatology 2011, 54, 2025–2035. [Google Scholar] [CrossRef] [PubMed]
- Ngo-Yin Fan, D.; Ho-Ching Tsang, F.; Hoi-Kam Tam, A.; Leung-Kuen Au, S.; Chak-Lui Wong, C.; Wei, L.; Man-Fong Lee, J.; He, X.; Oi-Lin Ng, I.; Wong, C.M. Histone lysine methyltransferase, suppressor of variegation 3-9 homolog 1, promotes hepatocellular carcinoma progression and is negatively regulated by microRNA-125b. Hepatology 2013, 57, 637–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wang, Y.; Guo, Y.; Sun, S. Down-regulated microRNA-152 induces aberrant DNA methylation in hepatitis B virus-related hepatocellular carcinoma by targeting DNA methyltransferase 1. Hepatology 2010, 52, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Luo, J.; Tian, R.; Sun, H.; Zou, S. miR-373 negatively regulates methyl-CpG-binding domain protein 2 (MBD2) in hilar cholangiocarcinoma. Dig. Dis. Sci. 2011, 56, 1693–1701. [Google Scholar] [CrossRef]
- Chen, Y.J.; Luo, J.; Yang, G.Y.; Yang, K.; Wen, S.Q.; Zou, S.Q. Mutual regulation between microRNA-373 and methyl-CpGbinding domain protein 2 in hilar cholangiocarcinoma. World J. Gastroenterol. 2012, 18, 3849–3861. [Google Scholar] [CrossRef]
- Braconi, C.; Huang, N.; Patel, T. Microrna-dependent regulation of DNA methyltransferase-1 and tumor suppressor gene expression by interleukin-6 in human malignant cholangiocytes. Hepatology 2010, 51, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Pal, S.; Sif, S. Protein Arginine Methyltransferase 5 Suppresses the Transcription of the RB Family of Tumor Suppressors in Leukemia and Lymphoma Cells. Mol. Cell. Biol. 2008, 28, 6262–6277. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.M.; Jeon, H.S.; Lee, S.Y.; Jeong, K.J.; Park, S.Y.; Lee, H.Y.; Lee, J.U.; Kim, J.H.; Kwon, S.J.; Choi, E.; et al. microRNA-101 inhibits lung cancer invasion through the regulation of enhancer of zeste homolog 2. Exp. Ther. Med. 2011, 2, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Asangani, I.A.; Harms, P.W.; Dodson, L.; Pandhi, M.; Kunju, L.P.; Maher, C.A.; Fullen, D.R.; Johnson, T.M.; Giordano, T.J.; Palanisamy, N.; et al. Genetic and epigenetic loss of microRNA-31 leads to feed-forward expression of EZH2 in melanoma. Oncotarget 2012, 3, 1011–1025. [Google Scholar] [CrossRef] [Green Version]
- Amodio, N.; Leotta, M.; Bellizzi, D.; Di Martino, M.T.; D’Aquila, P.; Lionetti, M.; Fabiani, F.; Leone, E.; Gullà, A.M.; Passarino, G.; et al. DNA-demethylating and anti-tumor activity of synthetic miR-29b mimics in multiple myeloma. Oncotarget 2012, 3, 1246–1258. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc. Natl. Acad. Sci. USA. 2007, 104, 15805–15810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Zhao, J.; Peng, X.; Liang, J.; Deng, X.; Chen, Y. The mechanism involved in the loss of PTEN expression in NSCLC tumor cells. Biochem. Biophys. Res. Commun. 2012, 418, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Samakoglu, S.; Deevi, D.S.; Li, H.; Wang, S.; Murphy, M.; Bao, C.; Bassi, R.; Prewett, M.; Tonra, J.R. Preclinical rationale for combining an EGFR antibody with cisplatin/gemcitabine for the treatment of NSCLC. Cancer Genom. Proteom. 2012, 9, 77–92. [Google Scholar] [PubMed]
- Xiang, Y.; Ma, N.; Wang, D.; Zhang, Y.; Zhou, J.; Wu, G.; Zhao, R.; Huang, H.; Wang, X.; Qiao, Y.; et al. MiR-152 and miR-185 co-contribute to ovarian cancer cells cisplatin sensitivity by targeting DNMT1 directly: A novel epigenetic therapy independent of decitabine. Oncogene 2014, 33, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Majid, S.; Dar, A.A.; Saini, S.; Shahryari, V.; Arora, S.; Zaman, M.S.; Chang, I.; Yamamura, S.; Tanaka, Y.; Chiyomaru, T.; et al. miRNA-34b inhibits prostate cancer through demethylation, active chromatin modifications, and AKT pathways. Clin. Cancer Res. 2013, 19, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Noonan, E.J.; Place, R.F.; Pookot, D.; Basak, S.; Whitson, J.M.; Hirata, H.; Giardina, C.; Dahiya, R. MiR-449a targets HDAC-1 and induces growth arrest in prostate cancer. Oncogene 2009, 28, 1714–1724. [Google Scholar] [CrossRef] [Green Version]
- Cao, P.; Deng, Z.; Wan, M.; Huang, W.; Cramer, S.D.; Xu, J.; Lei, M.; Sui, G. MicroRNA-101 negatively regulates Ezh2 and its expression is modulated by androgen receptor and HIF-1α/HIF-1β. Mol. Cancer 2010, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, C.; Chen, Z.; Jin, Y.; Wang, Y.; Kolokythas, A.; Dai, Y.; Zhou, X. MicroRNA-138 suppresses epithelial-mesenchymal transition in squamous cell carcinoma cell lines. Biochem. J. 2011, 440, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, D.; Imaizumi, Y.; Hasegawa, H.; Osaka, A.; Tsukasaki, K.; Lim Choi, Y.; Mano, H.; Marquez, V.E.; Hayashi, T.; Yanagihara, K.; et al. Overexpression of enhancer of zeste homolog 2 with trimethylation of lysine 27 on histone H3 in adult T-cell leukemia/lymphoma as a target for epigenetic therapy. Haematologica 2011, 96, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Roscigno, G.; Quintavalle, C.; Donnarumma, E.; Puoti, I.; Diaz-Lagares, A.; Iaboni, M.; Fiore, D.; Russo, V.; Todaro, M.; Romano, G.; et al. MiR-221 promotes stemness of breast cancer cells by targeting DNMT3b. Oncotarget 2016, 7, 580–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Liu, J.; Li, X.; Xiao, G.; Wang, H.; Yang, G.; Li, Y.; Tang, S.C.; Qin, S.; Du, N.; et al. MYC and DNMT3A-mediated DNA methylation represses microRNA-200b in triple negative breast cancer. J. Cell Mol. Med. 2018, 22, 6262–6274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Young, J.; Prabhala, H.; Pan, E.; Mestdagh, P.; Muth, D.; Teruya-Feldstein, J.; Reinhardt, F.; Onder, T.T.; Valastyan, S.; et al. miR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat. Cell. Biol. 2010, 12, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selcuklu, S.D.; Donoghue, M.T.; Rehmet, K.; de Souza Gomes, M.; Fort, A.; Kovvuru, P.; Muniyappa, M.K.; Kerin, M.J.; Enright, A.J.; Spillane, C. MicroRNA-9 inhibition of cell proliferation and identification of novel miR-9 targets by transcriptome profiling in breast cancer cells. J. Biol. Chem. 2012, 287, 29516–29528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lujambio, A.; Ropero, S.; Ballestar, E.; Fraga, M.F.; Cerrato, C.; Setien, F.; Casado, S.; Suarez-Gauthier, A.; Sanchez-Cespedes, M.; Git, A.; et al. Genetic unmasking of an epigenetically silenced microRNA in human cancer cells. Cancer Res. 2007, 67, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, U.; Hasemeier, B.; Christgen, M.; Muller, M.; Romermann, D.; Langer, F.; Kreipe, H. Epigenetic inactivation of microRNA gene hsa-mir-9-1 in human breast cancer. J. Pathol. 2008, 214, 17–24. [Google Scholar] [CrossRef]
- Bandres, E.; Agirre, X.; Bitarte, N.; Ramirez, N.; Zarate, R.; Roman-Gomez, J.; Prosper, F.; Garcia-Foncillas, J. Epigenetic regulation of microRNA expression in colorectal cancer. Int. J. Cancer 2009, 125, 2737–2743. [Google Scholar] [CrossRef]
- Chen, P.; Price, C.; Li, Z.; Li, Y.; Cao, D.; Wiley, A.; He, C.; Gurbuxani, S.; Kunjamma, R.B.; Huang, H.; et al. miR-9 is an essential oncogenic microRNA specifically overexpressed in mixed lineage leukemia-rearranged leukemia. Proc. Natl. Acad. Sci. USA 2013, 110, 11511–11516. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; An, X.; Yu, H.; Zhang, S.; Tang, B.; Zhang, X.; Li, Z. MiR-29b/TET1/ZEB2 signaling axis regulates metastatic properties and epithelial-mesenchymal transition in breast cancer cells. Oncotarget 2017, 8, 102119–102133. [Google Scholar] [CrossRef]
- Asmar, F.; Punj, V.; Christensen, J.; Pedersen, M.T.; Pedersen, A.; Nielsen, A.B.; Hother, C.; Ralfkiaer, U.; Brown, P.; Ralfkiaer, E.; et al. Genome-wide profiling identifies a DNA methylation signature that associates with TET2 mutations in diffuse large B-cell lymphoma. Haematologica 2013, 98, 1912–1920. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S. Dysregulation of TET2 in hematologic malignancies. Int. J. Hematol. 2017, 105, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inomata, M.; Tagawa, H.; Guo, Y.M.; Kameoka, Y.; Takahashi, N.; Sawada, K. MicroRNA-17-92 down-regulates expression of distinct targets in different B-cell lymphoma subtypes. Blood 2009, 113, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Paik, J.H.; Jang, J.Y.; Jeon, Y.K.; Kim, W.Y.; Kim, T.M.; Heo, D.S.; Kim, C.W. MicroRNA-146a downregulates NFκB activity via targeting TRAF6 and functions as a tumor suppressor having strong prognostic implications in NK/T cell lymphoma. Clin. Cancer Res. 2011, 17, 4761–4771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, A.E.; Zheng, T.; Yi, C.; Leaderer, D.; Weidhaas, J.; Slack, F.; Zhang, Y.; Paranjape, T.; Zhu, Y. microRNA miR-196a-2 and breast cancer: A genetic and epigenetic association study and functional analysis. Cancer Res. 2009, 69, 5970–5977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, R.; Scheel, C.; Weinhold, S.; Honisch, E.; Iwaniuk, K.M.; Trompeter, H.I.; Niederacher, D.; Wernet, P.; Santourlidis, S.; Uhrberg, M. Role of DNA methylation in miR-200c/141 cluster silencing in invasive breast cancer cells. BMC Res. Notes. 2010, 3, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tryndyak, V.P.; Beland, F.A.; Pogribny, I.P. E-cadherin transcriptional down-regulation by epigenetic and microRNA-200 family alterations is related to mesenchymal and drug-resistant phenotypes in human breast cancer cells. Int. J. Cancer 2010, 126, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Wences, H.; Martínez-Carrillo, D.N.; Peralta-Zaragoza, O.; Campos-Viguri, G.E.; Hernández-Sotelo, D.; Jiménez-López, M.A.; Muñoz-Camacho, J.G.; Garzón-Barrientos, V.H.; Illades-Aguiar, B.; Fernández-Tilapa, G. Methylation and expression of miRNAs in precancerous lesions and cervical cancer with HPV16 infection. Oncol. Rep. 2016, 35, 2297–2305. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, H.; Sun, L.; Shen, S.; Zhou, Q.; Yuan, Y.; Xing, C. Epigenetic Alternations of MicroRNAs and DNA Methylation Contribute to Liver Metastasis of Colorectal Cancer. Dig. Dis. Sci. 2019, 64, 1523–1534. [Google Scholar] [CrossRef]
- Deng, H.; Guo, Y.; Song, H.; Xiao, B.; Sun, W.; Liu, Z.; Yu, X.; Xia, T.; Cui, L.; Guo, J. MicroRNA-195 and microRNA-378 mediate tumor growth suppression by epigenetical regulation in gastric cancer. Gene 2013, 518, 351–359. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.W.; Wu, C.W.; Hu, L.Y.; Li, S.C.; Liao, Y.L.; Lai, C.H.; Kao, H.W.; Fang, W.L.; Huang, K.H.; Chan, W.C.; et al. Epigenetic regulation of miR-34b and miR-129 expression in gastric cancer. Int. J. Cancer 2011, 129, 2600–2610. [Google Scholar] [CrossRef]
- Alpini, G.; Glaser, S.S.; Zhang, J.P.; Francis, H.; Han, Y.; Gong, J.; Stokes, A.; Francis, T.; Hughart, N.; Hubble, L.; et al. Regulation of placenta growth factor by microRNA-125b in hepatocellular cancer. J. Hepatol. 2011, 55, 1339–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tivnan, A.; Zhao, J.; Johns, T.G.; Day, B.W.; Stringer, B.W.; Boyd, A.W.; Tiwari, S.; Giles, K.M.; Teo, C.; McDonald, K.L. The tumor suppressor microRNA, miR-124a, is regulated by epigenetic silencing and by the transcriptional factor, REST in glioblastoma. Tumor. Biol. 2014, 35, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Braconi, C.; Kogure, T.; Valeri, N.; Huang, N.; Nuovo, G.; Costinean, S.; Negrini, M.; Miotto, E.; Croce, C.M.; Patel, T. MicroRNA-29 can regulate expression of the long non-coding RNA gene MEG3 in hepatocellular cancer. Oncogene 2011, 30, 4750–4756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, H.Y.; Wan, T.S.; So, C.C.; Chim, C.S. Epigenetic inactivation of DAPK1, p14ARF, mir-34a and -34b/c in acute promyelocytic leukaemia. J. Clin. Pathol. 2014, 67, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Chim, C.S.; Wong, K.Y.; Leung, C.Y.; Chung, L.P.; Hui, P.K.; Chan, S.Y.; Yu, L. Epigenetic inactivation of the hsa-miR-203 in haematological malignancies. J. Cell. Mol. Med. 2011, 15, 2760–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.Y.; So, C.C.; Loong, F.; Chung, L.P.; Lam, W.W.L.; Liang, R.; Li, G.K.H.; Jin, D.Y.; Chim, C.S. Epigenetic inactivation of the miR-124-1 in haematological malignancies. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.Y.; Yim, R.L.H.; Kwong, Y.L.; Leung, C.Y.; Hui, P.K.; Cheung, F.; Liang, R.; Jin, D.Y.; Chim, C.S. Epigenetic inactivation of the MIR129-2 in hematological malignancies. J. Hematol. Oncol. 2013, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rusek, M.M.; Abba, M.; Eljaszewicz, A.; Moniuszko, M.; Niklinski, J.; Allgayer, H. MicroRNA modulators of epigenetic regulation, the tumor microenvironment and the immune system in lung cancer. Mol. Cancer 2015, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mazar, J.; DeBlasio, D.; Govindarajan, S.S.; Zhang, S.; Perera, R.J. Epigenetic regulation of microRNA-375 and its role in melanoma development in humans. FEBS Lett. 2011, 585, 2467–2476. [Google Scholar] [CrossRef] [Green Version]
- Azizi, M.; Teimoori-Toolabi, L.; Arzanani, M.K.; Azadmanesh, K.; Fard-Esfahani, P.; Zeinali, S. MicroRNA-148b and microRNA-152 reactivate tumor suppressor genes through suppression of DNA methyltransferase-1 gene in pancreatic cancer cell lines. Cancer Biol. Ther. 2014, 15, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulf, T.; Sibbritt, T.; Wiklund, E.D.; Patterson, K.; Song, J.Z.; Stirzaker, C.; Qu, W.; Nair, S.; Horvath, L.G.; Armstrong, N.J.; et al. Epigenetic-induced repression of microRNA-205 is associated with MED1 activation and a poorer prognosis in localized prostate cancer. Oncogene 2013, 32, 2891–2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrba, L.; Garbe, J.C.; Stampfer, M.R.; Futscher, B.W. Epigenetic regulation of normal human mammary cell type-specific miRNAs. Genome Res. 2011, 21, 2026–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, S.; Oncology, R.; General, M. Breast Cancer Breast Cancer. J. R. Soc. Med. 2016, 70, 515–517. [Google Scholar] [CrossRef]
- Veeck, J.; Esteller, M. Breast cancer epigenetics: From DNA methylation to microRNAs. J. Mammary Gland Biol. Neoplasia 2010, 15, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, R.; Pilato, B.; Ottini, L.; Lambo, R.; Simone, G.; Paradiso, A.; Tommasi, S. Different methylation and MicroRNA expression pattern in male and female familial breast cancer. J. Cell. Physiol. 2013, 228, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Duru, N.; Gernapudi, R.; Eades, G.; Eckert, R.; Zhou, Q. Epigenetic Regulation of miRNAs and Breast Cancer Stem Cells. Curr. Pharmacol. Rep. 2015, 1, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Rocha Simonini, P.D.S.; Breiling, A.; Gupta, N.; Malekpour, M.; Youns, M.; Omranipour, R.; Malekpour, F.; Volinia, S.; Croce, C.M.; Najmabadi, H.; et al. Epigenetically deregulated microRNA-375 is involved in a positive feedback loop with estrogen receptor α in breast cancer cells. Cancer Res. 2010, 70, 9175–9184. [Google Scholar] [CrossRef] [Green Version]
- de Brot, S.; Rutland, C.S.; Mongan, N.P.; James, V. Epigenetic Control. of MicroRNA Expression and Cancer; Elsevier Inc.: Atlanta, GA, USA, 2018; ISBN 9780128110225. [Google Scholar]
- Li, Y.; Kong, D.; Ahmad, A.; Bao, B.; Dyson, G.; Sarkar, F.H. Epigenetic deregulation of miR-29a and miR-1256 by isoflavone contributes to the inhibition of prostate cancer cell growth and invasion. Epigenetics 2012, 7, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sarkar, F.H. Targeting Epigenetically Deregulated miRNA by Nutraceuticals: Focusing on Cancer Prevention and Treatment. Curr. Pharmacol. Rep. 2015, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Guil, S.; Esteller, M. DNA methylomes, histone codes and miRNAs: Tying it all together. Int. J. Biochem. Cell. Biol. 2009, 41, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Sheervalilou, R.; Shirvaliloo, S.; Aval, S.F.; Khamaneh, A.M.; Sharifi, A.; Ansarin, K.; Zarghami, N. A new insight on reciprocal relationship between microRNA expression and epigenetic modifications in human lung cancer. Tumor. Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He-Yuan Qi; Zhao-Jun Zhang; Ya-Juan Li; Xian-Dong Fang No Title. Hereditas 2011, 33, 1291–1299.
- Li, B.; Carey, M.; Workman, J.L. The Role of Chromatin during Transcription. Cell 2007, 128, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-resolution profiling of histone methylations in the human genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Tao, B.B.; Liu, X.Q.; Zhang, W.; Li, S.; Dong, D.; Xiao, M.; Zhong, J. Evidence for the association of chromatin and microRNA regulation in the human genome. Oncotarget 2017, 8, 70958–70966. [Google Scholar] [CrossRef]
- Linares, A.; Dalenc, F.; Balaguer, P.; Boulle, N.; Cavailles, V. Manipulating protein acetylation in breast cancer: A promising approach in combination with hormonal therapies? J. Biomed. Biotechnol. 2011, 2011, 856985. [Google Scholar] [CrossRef] [Green Version]
- Tessarz, P.; Kouzarides, T. Histone core modifications regulating nucleosome structure and dynamics. Nat. Rev. Mol. Cell. Biol. 2014, 15, 703–708. [Google Scholar] [CrossRef]
- Zentner, G.E.; Henikoff, S. Regulation of nucleosome dynamics by histone modifications. Nat. Struct. Mol. Biol. 2013, 20, 259–266. [Google Scholar] [CrossRef]
- Shen, T.; Sanchez, H.N.; Zan, H.; Casali, P. Genome-Wide Analysis Reveals Selective Modulation of microRNAs and mRNAs by Histone Deacetylase Inhibitor in B Cells Induced to Undergo Class-Switch DNA Recombination and Plasma Cell Differentiation. Front. Immunol. 2015, 6, 627. [Google Scholar] [CrossRef] [Green Version]
- Bamodu, O.A.; Huang, W.C.; Lee, W.H.; Wu, A.; Wang, L.S.; Hsiao, M.; Yeh, C.T.; Chao, T.Y. Aberrant KDM5B expression promotes aggressive breast cancer through MALAT1 overexpression and downregulation of hsa-miR-448. BMC Cancer 2016, 16, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wu, W.; Claret, F.X. Mutual regulation of microRNAs and DNA methylation in human cancers. Epigenetics 2017, 12, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, J.C.; Jones, P.A. Epigenetics and microRNAs. Pediatr. Res. 2007, 61, 24–29. [Google Scholar] [CrossRef]
- Nowacka-Zawisza, M.; Wiśnik, E. DNA methylation and histone modifications as epigenetic regulation in prostate cancer (review). Oncol. Rep. 2017, 38, 2587–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari Moqadam, F.; Pieters, R.; Den Boer, M.L. The hunting of targets: Challenge in miRNA research. Leukemia 2013, 27, 16–23. [Google Scholar] [CrossRef]
- Tarang, S.; Weston, M.D. Macros in microRNA target identification: A comparative analysis of in silico, in vitro, and in vivo approaches to microRNA target identification. RNA Biol. 2014, 11, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.P.; Goodall, G.J.; Liu, B. Understanding principles of miRNA target recognition and function through integrated biological and bioinformatics approaches. Wiley Interdiscip. Rev. RNA 2014, 5, 361–379. [Google Scholar] [CrossRef]
- Akhtar, M.M.; Micolucci, L.; Islam, M.S.; Olivieri, F.; Procopio, A.D. Bioinformatic tools for microRNA dissection. Nucleic Acids Res. 2016, 44, 24–44. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.C.; Bovolenta, L.A.; Nachtigall, P.G.; Herkenhoff, M.E.; Lemke, N.; Pinhal, D. Combining results from distinct microRNA target prediction tools enhances the performance of analyses. Front. Genet. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Shi, J.; Gao, Y.; Cui, C.; Zhang, S.; Li, J.; Zhou, Y.; Cui, Q. HMDD v3.0: A database for experimentally supported human microRNA-disease associations. Nucleic Acids Res. 2019, 47, D1013–D1017. [Google Scholar] [CrossRef] [Green Version]
- Xie, B.; Ding, Q.; Han, H.; Wu, D. MiRCancer: A microRNA-cancer association database constructed by text mining on literature. Bioinformatics 2013, 29, 638–644. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chang, S.; Zhang, J.; Zhao, Q.; Xiang, H.; Kusonmano, K.; Yang, L.; Sun, Z.S.; Yang, H.; Wang, J. MethyCancer: The database of human DNA methylation and cancer. Nucleic Acids Res. 2008, 36, 836–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, S.; Lu, J.; Sun, M.; Li, Z.; Zhang, H.; Neilly, M.B.; Wang, Y.; Qian, Z.; Jin, J.; Zhang, Y.; et al. MicroRNA expression signatures accurately discriminate acute lymphoblastic leukemia from acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2007, 104, 19971–19976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotte, D.; Chau, J.C.; Sylvester, G.; Liu, G.; Chen, C.; van der Velden, V.H.; Broekhuis, M.J.; Peters, T.C.; Pieters, R.; den Boer, M.L. Identification of new microRNA genes and aberrant microRNA profiles in childhood acute lymphoblastic leukemia. Leukemia 2009, 23, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Schotte, D.; De Menezes, R.X.; Akbari Moqadam, F.; Khankahdani, L.M.; Lange-Turenhout, E.; Chen, C.; Pieters, R.; Den Boer, M.L. MicroRNA characterize genetic diversity and drug resistance in pediatric acute lymphoblastic leukemia. Haematologica 2011, 96, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Martin-Palanco, V.; Rodriguez, G.; Martin, C.; Rojas, R.; Torres, A.; Roman-Gomez, J. microRNA methylation profile has prognosis impact in acute lymphoblastic leukemia patients undergoing stem cell transplantation. Biol. Blood Marrow Transpl. 2011, 17, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Esau, C.C.; Monia, B.P. Therapeutic potential for microRNAs. Adv. Drug Deliv. Rev. 2007, 59, 101–114. [Google Scholar] [CrossRef]
- Wang, V.; Wu, W. MicroRNA-based therapeutics for cancer. BioDrugs 2009, 23, 15–23. [Google Scholar] [CrossRef]
- Janssen, E.A.; Slewa, A.; Gudlaugsson, E.; Jonsdottir, K.; Skaland, I.; Soiland, H.; Baak, J.P. Biologic profiling of lymph node negative breast cancers by means of microRNA expression. Mod. Pathol. 2010, 23, 1567–1576. [Google Scholar] [CrossRef] [Green Version]
- Rio-Machin, A.; Ferreira, B.I.; Henry, T.; Gomez-Lopez, G.; Agirre, X.; Alvarez, S.; Rodriguez-Perales, S.; Prosper, F.; Calasanz, M.J.; Martinez, J.; et al. Downregulation of specific miRNAs in hyperdiploid multiple myeloma mimics the oncogenic effect of IgH translocations occurring in the non-hyperdiploid subtype. Leukemia 2013, 27, 925–931. [Google Scholar] [CrossRef] [Green Version]
- Otoukesh, B.; Abbasi, M.; Gorgani, H.O.; Farahini, H.; Moghtadaei, M.; Boddouhi, B.; Kaghazian, P.; Hosseinzadeh, S.; Alaee, A. MicroRNAs signatures, bioinformatics analysis of miRNAs, miRNA mimics and antagonists, and miRNA therapeutics in osteosarcoma. Cancer Cell Int. 2020, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, D.Y.; Huang, L. In vivo delivery of miRNAs for cancer therapy: Challenges and strategies. Adv. Drug Deliv. Rev. 2015, 81, 128–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.; Gao, Y.; Long, Y.; Chen, L.; Huang, Y.; Fang, L. MiRNA-497 regulates cell growth and invasion by targeting cyclin E1 in breast cancer. Cancer Cell Int. 2013, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sun, H.H.; Sui, M.H.; Ma, J.J. miR-218 inhibits acute promyelocytic leukemia cell growth by targeting BMI-1. Oncol. Lett. 2017, 14, 8078–8083. [Google Scholar] [CrossRef] [PubMed]
- Dorrance, A.M.; Neviani, P.; Ferenchak, G.J.; Huang, X.; Nicolet, D.; Maharry, K.S.; Ozer, H.G.; Hoellarbauer, P.; Khalife, J.; Hill, E.B.; et al. Targeting leukemia stem cells in vivo with antagomiR-126 nanoparticles in acute myeloid leukemia. Leukemia 2015, 29, 2143–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ors-Kumoglu, G.; Gulce-Iz, S.; Biray-Avci, C. Therapeutic microRNAs in human cancer. Cytotechnology 2019, 71, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Mercatelli, N.; Fortini, D.; Palombo, R.; Paronetto, M.P. Small molecule inhibition of Ewing sarcoma cell growth via targeting the long non coding RNA HULC. Cancer Lett. 2020, 469, 111–123. [Google Scholar] [CrossRef]
- Jung, J.; Yeom, C.; Choi, Y.S.; Kim, S.; Lee, E.J.; Park, M.J.; Kang, S.W.; Kim, S.B.; Chang, S. Simultaneous inhibition of multiple oncogenic miRNAs by a multi-potent microRNA sponge. Oncotarget 2015, 6, 20370–20387. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Ju, J.; Hou, L.; Li, Z.; Xiao, D.; Li, Y.; Yao, J.; Wang, C.; Zhang, Y.; et al. Downregulation of miR-522 suppresses proliferation and metastasis of non-small cell lung cancer cells by directly targeting DENN/MADD domain containing 2D. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Yang, B.; Lin, H.; Lu, Y.; Luo, X.; Wang, Z. Novel approaches for gene-specific interference via manipulating actions of microRNAs: Examination on the pacemaker channel genes HCN2 and HCN4. J. Cell. Physiol. 2007, 212, 285–292. [Google Scholar] [CrossRef]
- Saito, Y.; Liang, G.; Egger, G.; Friedman, J.M.; Chuang, J.C.; Coetzee, G.A.; Jones, P.A. Specific activation of microRNA-127 with downregulation of the proto-oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell 2006, 9, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldivar-Gonzalez, F.I.; Gomez-Garcia, A.; Chavez-Ponce de Leon, D.E.; Sanchez-Cruz, N.; Ruiz-Rios, J.; Pilon-Jimenez, B.A.; Medina-Franco, J.L. Inhibitors of DNA Methyltransferases From Natural Sources: A Computational Perspective. Front. Pharmacol. 2018, 9, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, A.C.; Johnstone, R.W.; West, A.C.; Johnstone, R.W. New and emerging HDAC inhibitors for cancer treatment Find the latest version: Review series New and emerging HDAC inhibitors for cancer treatment. J. Clin. Investig. 2014, 124, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.C.; Li, H.; Van Neste, L.; Cai, Y.; Robert, C.; Rassool, F.V.; Shin, J.J.; Harbom, K.M.; Beaty, R.; Pappou, E.; et al. Transient Low Doses of DNA-Demethylating Agents Exert Durable Antitumor Effects on Hematological and Epithelial Tumor Cells. Cancer Cell 2012, 21, 430–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, K.; Das, V.; Vyas, P.; Hajduch, M. Nucleosidic DNA demethylating epigenetic drugs—A comprehensive review from discovery to clinic. Pharmacol. Ther. 2018, 188, 45–79. [Google Scholar] [CrossRef]
- Azad, N.; Zahnow, C.A.; Rudin, C.M.; Baylin, S.B. The future of epigenetic therapy in solid tumours—Lessons from the past. Nat. Rev. Clin. Oncol. 2013, 10, 256–266. [Google Scholar] [CrossRef]
- Reid, G.; Kao, S.C.; Pavlakis, N.; Brahmbhatt, H.; MacDiarmid, J.; Clarke, S.; Boyer, M.; van Zandwijk, N. Clinical development of TargomiRs, a miRNA mimic-based treatment for patients with recurrent thoracic cancer. Epigenomics 2016, 8, 1079–1085. [Google Scholar] [CrossRef] [Green Version]
- van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet. Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- de Gooijer, C.J.; Baas, P.; Burgers, J.A. Current chemotherapy strategies in malignant pleural mesothelioma. Transl. Lung Cancer Res. 2018, 7, 574–583. [Google Scholar] [CrossRef]
- National Institute of Health. U.S National Library of Medicine Clinical Trials. Available online: https://clinicaltrials.gov/ct2 (accessed on 10 August 2020).
- Huang, Z.; Zhang, L.; Zhu, D.; Shan, X.; Zhou, X.; Qi, L.W.; Wu, L.; Zhu, J.; Cheng, W.; Zhang, H.; et al. A novel serum microRNA signature to screen esophageal squamous cell carcinoma. Cancer Med. 2017, 6, 109–119. [Google Scholar] [CrossRef]
- Zhou, X.; Wen, W.; Shan, X.; Zhu, W.; Xu, J.; Guo, R.; Cheng, W.; Wang, F.; Qi, L.W.; Chen, Y.; et al. A six-microRNA panel in plasma was identified as a potential biomarker for lung adenocarcinoma diagnosis. Oncotarget 2017, 8, 6513–6525. [Google Scholar] [CrossRef] [PubMed]
- Monzo, M.; Santasusagna, S.; Moreno, I.; Martinez, F.; Hernández, R.; Muñoz, C.; Castellano, J.J.; Moreno, J.; Navarro, A. Exosomal microRNAs isolated from plasma of mesenteric veins linked to liver metastases in resected patients with colon cancer. Oncotarget 2017, 8, 30859–30869. [Google Scholar] [CrossRef] [PubMed]
- Dirin, M.; Winkler, J. Influence of diverse chemical modifications on the ADME characteristics and toxicology of antisense oligonucleotides. Expert. Opin. Biol. Ther. 2013, 13, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Velpurisiva, P.; Gad, A.; Piel, B.; Jadia, R.; Rai, P. Nanoparticle Design Strategies for Effective Cancer Immunotherapy. J. Biomed. 2017, 2, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Wei, J.; Yu, C.; Han, X.; Qin, X.; Zhang, C.; Liao, W.; Li, L.; Huang, W. Non-viral nanocarriers for intracellular delivery of microRNA therapeutics. J. Mater. Chem. B 2019, 7, 1209–1225. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control. Release 2013, 172, 962–974. [Google Scholar] [CrossRef] [Green Version]
- Mercatelli, N.; Coppola, V.; Bonci, D.; Miele, F.; Costantini, A.; Guadagnoli, M.; Bonanno, E.; Muto, G.; Frajese, G.V.; De Maria, R.; et al. The inhibition of the highly expressed miR-221 and miR-222 impairs the growth of prostate carcinoma xenografts in mice. PLoS ONE 2008, 3, e4029. [Google Scholar] [CrossRef]
- Xu, Y.; So, C.; Lam, H.M.; Fung, M.C.; Tsang, S.Y. Apoptosis Reversal Promotes Cancer Stem Cell-Like Cell Formation. Neoplasia 2018, 20, 295–303. [Google Scholar] [CrossRef]
- Coward, J.; Harding, A. Size does matter: Why polyploid tumor cells are critical drug targets in the war on cancer. Front. Oncol. 2014, 4, 1–15. [Google Scholar] [CrossRef]
- Liu, J. The dualistic origin of human tumors. Semin. Cancer Biol. 2018, 53, 1–16. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, P.; Lv, H.; Zhang, D.; Liu, G.; Yang, Z.; Li, Y.; Liu, J.; Zhang, S. Number of polyploid giant cancer cells and expression of EZH2 are associated with VM formation and tumor grade in human ovarian tumor. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzayans, R.; Andrais, B.; Murray, D. Roles of polyploid/multinucleated giant cancer cells in metastasis and disease relapse following anticancer treatment. Cancers 2018, 10, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.A.; Piao, M.J.; Kim, K.C.; Kang, H.K.; Chang, W.Y.; Park, I.C.; Keum, Y.S.; Surh, Y.J.; Hyun, J.W. Epigenetic modification of Nrf2 in 5-fluorouracil-resistant colon cancer cells: Involvement of TET-dependent DNA demethylation. Cell Death Dis. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | miRNA | Expression | Epi Regulator | Epi Modification | Epi Target | Ref |
|---|---|---|---|---|---|---|
| Acquired resistance of breast cancer | miR-29a miR-29b-3p | High | DNMT-3a | DNA Methylation | global DNA methylation | [36] |
| miR-132 | High | MeCP2 | DNA Methylation | global DNA methylation | ||
| Acute myeloid leukaemia | miR-29b-3p | High | DNMT-3a/DNMT-3b/DNMT-1 | DNA Methylation | ESR1/cyclin-dependent kinase inhibitor 2B | [37] |
| Bladder transitional cell carcinoma | miR-101-3p | Low | EZH2 | H3K27me3 | - | [38] |
| Breast cancer | miR-10a | Low | - | H3K27me3/DNA Methylation | HOXD4 | [39] |
| miR-29c miR-26b miR-148b | High | DNMT-3b | DNA Methylation | CEACAM6/CST6/SCNN1A | [40] | |
| miR-148a | High | DNMT-1 | DNA Methylation | miR-148a | [41] | |
| miR-152 | High | DNMT-1 | DNA Methylation | miR-152 | ||
| Breast/Prostate | miR-101-3p | High | EZH2 | H3K27me3 | miR-101 | [42] |
| Colorectal cancer | miR-143 | Low | DNMT-3a | DNA Methylation | - | [43] |
| miR-342 | High | DNMT-1 | DNA Methylation | ADAM23/Hint1/RASSF1A/RECK | [44] | |
| Cutaneous melanoma | miR-29a-3p | High | DNMT-3a/DNMT-3b | DNA Methylation | RASSF1A/TFPI-2/RAR-/SOCS/GATA4 | [45] |
| Endometrial cancer | miR-152 | Low | DNMT-1 | DNA Methylation | - | [46] |
| Gastric cancer | miR-148a | High | DNMT-1 | DNA Methylation | miR-148a | [47] |
| Glioblastoma | miR-101-3p | Low | EZH2 | H3K27me3 | - | [48] |
| miR-152 | High | DNMT-1 | DNA Methylation | miR-152 | [49] | |
| Glioma | miR-185 | High | DNMT-1 | DNA Methylation | ANKDD1A/GAD1/HIST1H3E/PCDHA8/PCDHA13/PHOX2B/SIX3/SST | [50] |
| Hepatitis B virus (HBV)-related hepatocellular carcinoma | miR-101-3p | High | DNMT-3a | DNA Methylation | RASSF1/PRDM2/GSTP1/RUNX3 | [51] |
| Hepatocellular carcinoma | miR-200a | High | HDAC4 | H3ac | miR-200a | [52] |
| miR-125b-5p | High | SUV39H1 | H3K9me3 | Ki67 | [53] | |
| miR-152 | High | DNMT-1 | DNA Methylation | GSTP1/CDH1 | [54] | |
| Hilar cholangiocarcinoma | miR-373 | High | MBD2 | DNA Methylation | MBD2 regulate RASSF1A | [55] |
| High | MBD2 | DNA Methylation | miR-373 | [56] | ||
| Human malignant cholangiocytes | miR-148a miR-152 | High | DNMT-1 | DNA Methylation | Rassf1a/p16INK4a | [57] |
| Leukaemia and lymphoma cells | miR-19a miR-25 miR-32 miR-92b miR-96 | High | PRMT5 | DNA Methylation | H3R8/H4R3 | [58] |
| Lung cancer | miR-101-1 | High | EZH2 | H3K27me3 | CDH1 | [59] |
| Melanoma | miR-31 | High | EZH2 | DNA Methylation | miR-31 | [60] |
| Multiple myeloma | miR-29b-3p | Low | DNMT-3a/DNMT-3b | DNA Methylation | - | [61] |
| Non-small-cell lung cancer | miR-29a miR-29b-3p miR-29c | High | DNMT-3a/DNMT-3b | DNA Methylation | FHIT/WWOX | [62] |
| miR-29b-3p | High | DNMT-1/DNMT-3a/DNMT-3b | DNA Methylation | PTEN | [63] | |
| miR-29b-3p | High | DNMT-3b | DNA Methylation | CADM1/RASSF1/FHIT | [64] | |
| Ovarian cancer | miR-185 miR-152 | Low | DNMT-1 | DNA Methylation | - | [65] |
| Prostate cancer | miR-34b | High | HDAC1/HDAC2/HDAC4 | H3K4me3 | miR-34b | [66] |
| DNMT-1 | DNA Methylation | |||||
| miR-449a | Low | HDAC1 | Histone Acetylation | - | [67] | |
| miR-101-3p | Low | EZH2 | H3K27me3 | - | [68] | |
| Squamous cell carcinoma | miR-138-5p | High | EZH2 | H3K27me3 | E-cad | [69] |
| T-cell leukaemia | miR-101-3p miR-128 | Low | EZH2 | H3K27me3 | - | [70] |
| Database | Functions | Link | Reference |
|---|---|---|---|
| EpimiR | Contains 1974 regulatory relationships between 19 different epigenetic modifications and 617 miRNAs across Homo sapiens and 6 more species. The records are divided into two sections: Epi2miR and miR2Epi. | http://www.jianglab.cn/EpimiR/ | [24] |
| HMDD | The Human microRNA Disease Database (HMDD) collects curated experiment-supported evidence for disease-associated human miRNAs classified into 6 evidence classes (genetics, epigenetics, target, circulation, tissue and other) and 20 evidence codes. It also provides a disease-associated miRNA-target network visualisation function. | https://www.cuilab.cn/hmdd | [132] |
| miRCancer | The database presently records 878 interactions between 236 miRNAs and 79 human cancers through the processing of more than 26,000 published articles. | http://mircancer.ecu.edu/browse.jsp | [133] |
| MethyCancer | MethyCancer introduces highly integrated DNA methylation data, cancer-related gene, mutation and cancer evidence from numerous resources, and the CpG island (CGI) clones derived from large-scale sequencing. | http://methycancer.psych.ac.cn/ | [134] |
| miRNet | miRNet enables statistical analysis and functional interpretation of a variety of data produced from existing miRNA studies. The key features include: (i) integration of high-quality miRNA-target interaction data from 11 databases; (ii) differential expression analysis of data from microarray, RNA-seq and quantitative PCR; (iii) flexible options for data filtering, refinement and customisation during network creation; and (iv) a network visualisation system coupled with enrichment analysis. | https://www.mirnet.ca/miRNet/home.xhtml | [25] |
| miRNA | Hits | p-Value | Adj p-Value |
|---|---|---|---|
| mir-29 | 14 | 0 | 0 |
| mir-515 | 58 | 1.275 × 10−9 | 2.55 × 10−11 |
| mir-17 | 20 | 1.786667 × 10−8 | 5.36 × 10−10 |
| mir-26 | 9 | 0.00004275 | 0.00000171 |
| mir-30 | 13 | 0.00024 | 0.000012 |
| mir-25 | 10 | 0.00027 | 0.0000162 |
| mir-19 | 8 | 0.0007728571 | 0.0000541 |
| mir-193 | 6 | 0.0018125 | 0.000145 |
| mir-290 | 7 | 0.006945455 | 0.000764 |
| mir-9 | 7 | 0.006945455 | 0.000764 |
| mir-34 | 7 | 0.006945455 | 0.000764 |
| mir-196 | 6 | 0.02191667 | 0.00263 |
| mir-664 | 5 | 0.02207143 | 0.00309 |
| mir-129 | 5 | 0.02207143 | 0.00309 |
| mir-10 | 12 | 0.02226667 | 0.00334 |
| mir-15 | 8 | 0.03558824 | 0.0101 |
| mir-101 | 4 | 0.03558824 | 0.0113 |
| mir-1228 | 3 | 0.03558824 | 0.0121 |
| mir-454 | 3 | 0.03558824 | 0.0121 |
| mir-432 | 3 | 0.03558824 | 0.0121 |
| mir-483 | 3 | 0.03558824 | 0.0121 |
| mir-425 | 3 | 0.03558824 | 0.0121 |
| mir-322 | 3 | 0.03558824 | 0.0121 |
| mir-373 | 3 | 0.03558824 | 0.0121 |
| mir-127 | 3 | 0.03558824 | 0.0121 |
| mir-126 | 3 | 0.03558824 | 0.0121 |
| mir-191 | 3 | 0.03558824 | 0.0121 |
| mir-142 | 3 | 0.03558824 | 0.0121 |
| mir-140 | 3 | 0.03558824 | 0.0121 |
| mir-183 | 3 | 0.03558824 | 0.0121 |
| mir-139 | 3 | 0.03558824 | 0.0121 |
| mir-31 | 3 | 0.03558824 | 0.0121 |
| mir-22 | 3 | 0.03558824 | 0.0121 |
| mir-21 | 3 | 0.03558824 | 0.0121 |
| mir-28 | 5 | 0.05428571 | 0.019 |
| mir-365 | 4 | 0.06642857 | 0.0279 |
| mir-146 | 4 | 0.06642857 | 0.0279 |
| mir-221 | 4 | 0.06642857 | 0.0279 |
| mir-218 | 4 | 0.06642857 | 0.0279 |
| mir-132 | 4 | 0.06642857 | 0.0279 |
| mir-27 | 4 | 0.06642857 | 0.0279 |
| mir-24 | 4 | 0.06642857 | 0.0279 |
| mir-124 | 5 | 0.08069767 | 0.0347 |
| mir-663 | 3 | 0.09136364 | 0.0402 |
| let-7 | 13 | 0.1133333 | 0.051 |
| mir-1260a | 2 | 0.1147826 | 0.0528 |
| mir-135 | 4 | 0.1865957 | 0.0877 |
| mir-130 | 5 | 0.2350877 | 0.119 |
| mir-199 | 4 | 0.2350877 | 0.13 |
| mir-876 | 2 | 0.2350877 | 0.134 |
| mir-542 | 2 | 0.2350877 | 0.134 |
| mir-574 | 2 | 0.2350877 | 0.134 |
| mir-202 | 2 | 0.2350877 | 0.134 |
| mir-324 | 2 | 0.2350877 | 0.134 |
| mir-342 | 2 | 0.2350877 | 0.134 |
| mir-330 | 2 | 0.2350877 | 0.134 |
| mir-361 | 2 | 0.2350877 | 0.134 |
| mir-8 | 5 | 0.3482759 | 0.202 |
| mir-544 | 2 | 0.38 | 0.228 |
| mir-147 | 2 | 0.38 | 0.228 |
| mir-103 | 4 | 0.3819672 | 0.233 |
| mir-7 | 3 | 0.4163265 | 0.272 |
| mir-500 | 4 | 0.4163265 | 0.29 |
| mir-368 | 4 | 0.4163265 | 0.29 |
| mir-194 | 2 | 0.4163265 | 0.325 |
| mir-190 | 2 | 0.4163265 | 0.325 |
| mir-153 | 2 | 0.4163265 | 0.325 |
| mir-219 | 3 | 0.4163265 | 0.343 |
| mir-148 | 3 | 0.4163265 | 0.343 |
| mir-1256 | 1 | 0.4163265 | 0.407 |
| mir-1205 | 1 | 0.4163265 | 0.407 |
| mir-1204 | 1 | 0.4163265 | 0.407 |
| mir-1203 | 1 | 0.4163265 | 0.407 |
| mir-1202 | 1 | 0.4163265 | 0.407 |
| mir-944 | 1 | 0.4163265 | 0.407 |
| mir-943 | 1 | 0.4163265 | 0.407 |
| mir-935 | 1 | 0.4163265 | 0.407 |
| mir-760 | 1 | 0.4163265 | 0.407 |
| mir-765 | 1 | 0.4163265 | 0.407 |
| mir-1323 | 1 | 0.4163265 | 0.407 |
| mir-641 | 1 | 0.4163265 | 0.407 |
| mir-638 | 1 | 0.4163265 | 0.407 |
| mir-636 | 1 | 0.4163265 | 0.407 |
| mir-630 | 1 | 0.4163265 | 0.407 |
| mir-602 | 1 | 0.4163265 | 0.407 |
| mir-601 | 1 | 0.4163265 | 0.407 |
| mir-596 | 1 | 0.4163265 | 0.407 |
| mir-572 | 1 | 0.4163265 | 0.407 |
| mir-568 | 1 | 0.4163265 | 0.407 |
| mir-564 | 1 | 0.4163265 | 0.407 |
| mir-559 | 1 | 0.4163265 | 0.407 |
| mir-498 | 1 | 0.4163265 | 0.407 |
| mir-448 | 1 | 0.4163265 | 0.407 |
| mir-326 | 1 | 0.4163265 | 0.407 |
| mir-375 | 1 | 0.4163265 | 0.407 |
| mir-184 | 1 | 0.4163265 | 0.407 |
| mir-137 | 1 | 0.4163265 | 0.407 |
| mir-302 | 4 | 0.4163265 | 0.408 |
| mir-941 | 3 | 0.418 | 0.414 |
| mir-642 | 2 | 0.418 | 0.418 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arif, K.M.T.; Elliott, E.K.; Haupt, L.M.; Griffiths, L.R. Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets. Cancers 2020, 12, 2922. https://doi.org/10.3390/cancers12102922
Arif KMT, Elliott EK, Haupt LM, Griffiths LR. Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets. Cancers. 2020; 12(10):2922. https://doi.org/10.3390/cancers12102922
Chicago/Turabian StyleArif, K. M. Taufiqul, Esther K. Elliott, Larisa M. Haupt, and Lyn R. Griffiths. 2020. "Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets" Cancers 12, no. 10: 2922. https://doi.org/10.3390/cancers12102922