What Is the Role of Interleukins in Breast Cancer Bone Metastases? A Systematic Review of Preclinical and Clinical Evidence

 , and
, and
Abstract
:1. Introduction
2. Method
Literature Search
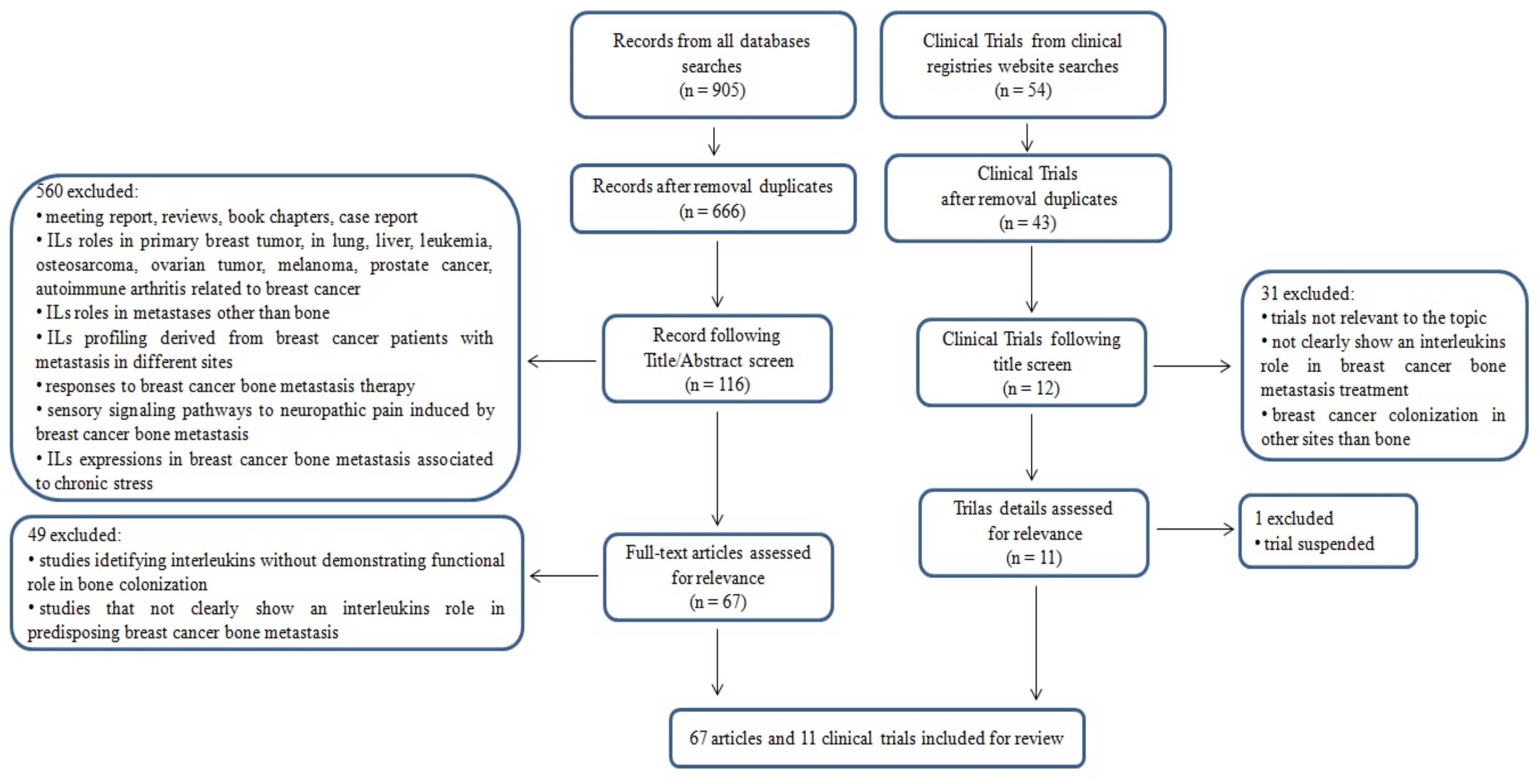
3. Results
3.1. Literature Search
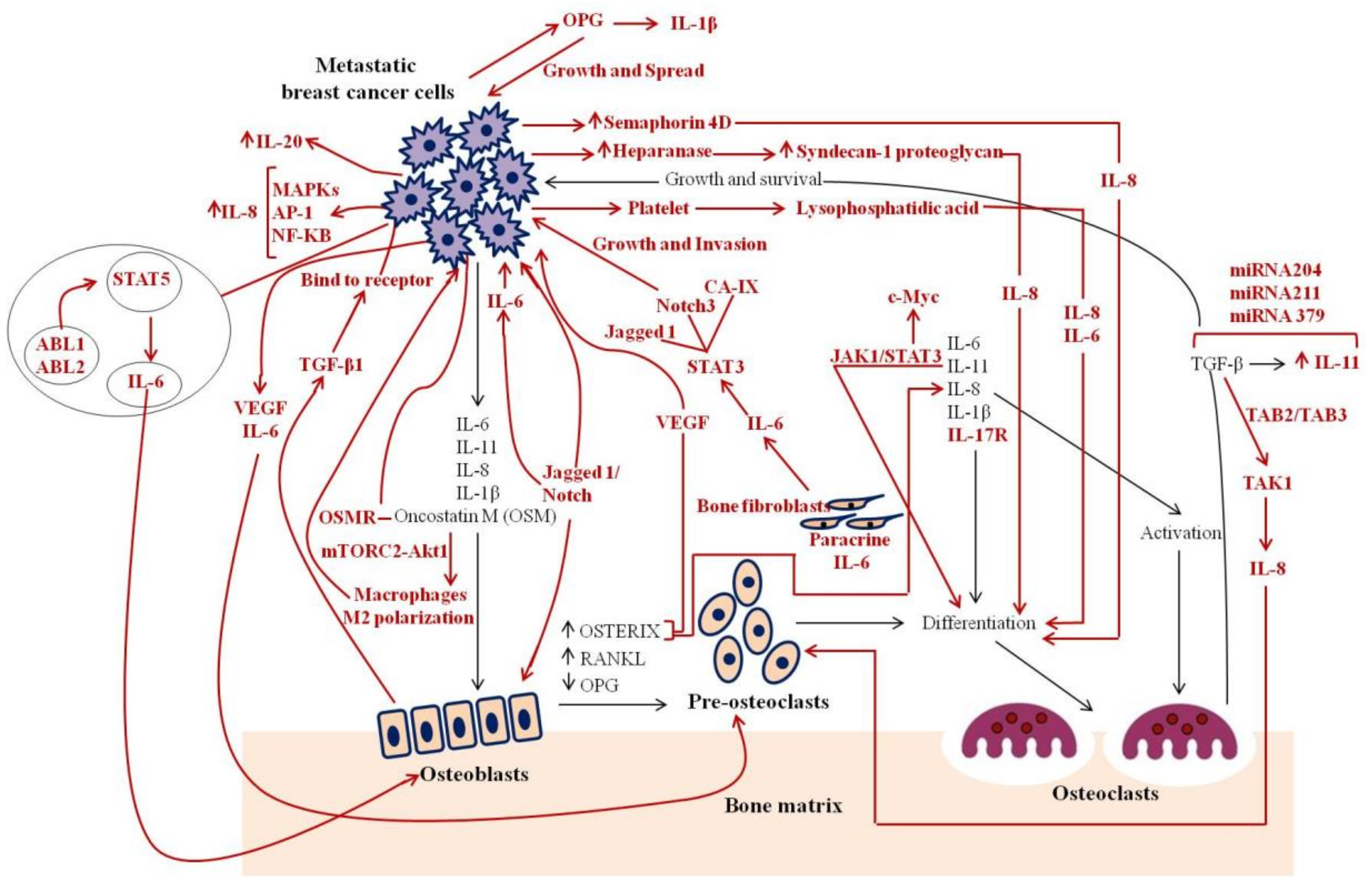
3.2. Findings from Preclinical Studies
3.2.1. IL-1
3.2.2. IL-2
3.2.3. IL-6
3.2.4. IL-8
3.2.5. IL-10
3.2.6. IL-11
3.2.7. IL-12
3.2.8. IL-15
3.2.9. IL-17
3.2.10. IL-18
3.2.11. IL-20
3.3. Findings from Clinical Trials
4. Discussion
5. Conclusions and Implications for Future Research
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Z.; Shi, A.; Lu, C.; Song, T.; Zhang, Z.; Zhao, J. Breast Cancer: Epidemiology and Etiology. Cell Biochem. Biophys. 2015, 72, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Kennecke, H.; Yerushalmi, R.; Woods, R.; Cheang, M.C.; Voduc, D.; Speers, C.H.; Nielsen, T.O.; Gelmon, K. Metastatic behavior of breast cancer subtypes. J. Clin. Oncol. 2010, 28, 3271–3277. [Google Scholar] [CrossRef] [PubMed]
- Liede, A.; Cai, M.; Crouter, T.F.; Niepel, D.; Callaghan, F.; Evans, D.G. Risk-reducing mastectomy rates in the US: A closer examination of the Angelina Jolie effect. Breast Cancer Res. Treat. 2018, 171, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Roodman, G.D. Mechanisms of bone metastasis. N. Engl. J. Med. 2004, 350, 1655–1664. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Lipton, A.; Uzzo, R.; Amato, R.J.; Ellis, G.K.; Hakimian, B.; Roodman, G.D.; Smith, M.R. The science and practice of bone health in oncology: Managing bone loss and metastasis in patients with solid tumors. J. Natl. Compr. Cancer Netw. 2009, 7 (Suppl. S7), S1–S29. [Google Scholar] [CrossRef] [Green Version]
- Selvaggi, G.; Scagliotti, G.V. Management of bone metastases in cancer: A review. Crit. Rev. Oncol. Hematol. 2005, 56, 365–367. [Google Scholar] [CrossRef]
- Weilbaecher, K.N.; Guise, T.A.; McCauley, L.K. Cancer to bone: A fatal attraction. Nat. Rev. Cancer 2011, 11, 411–425. [Google Scholar] [CrossRef]
- Phadke, P.A.; Mercer, R.R.; Harms, J.F.; Jia, Y.; Frost, A.R.; Jewell, J.L.; Bussard, K.M.; Nelson, S.; Moore, C.; Kappes, J.C.; et al. Kinetics of metastatic breast cancer cell trafficking in bone. Clin. Cancer Res. 2006, 12, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, R.A. Coming full circle-from endless complexity to simplicity and back again. Cell 2014, 157, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundy, G.R. Metastasis to bone: Causes, consequences and therapeutic opportunities. Nat. Rev. Cancer 2002, 2, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Chirgwin, J.M.; Mohammad, K.S.; Guise, T.A. Tumor-bone cellular interactions in skeletal metastases. J. Musculoskelet. Neuronal Interact. 2004, 4, 308–318. [Google Scholar] [PubMed]
- Guise, T.A.; Kozlow, W.M.; Heras-Herzig, A.; Padalecki, S.S.; Yin, J.J.; Chirgwin, J.M. Molecular mechanisms of breast cancer metastases to bone. Clin. Breast Cancer 2005, 5 (Suppl. S2), S46–S53. [Google Scholar] [CrossRef] [PubMed]
- Kan, C.; Vargas, G.; Pape, F.L.; Clézardin, P. Cancer Cell Colonisation in the Bone Microenvironment. Int. J. Mol. Sci. 2016, 17, E1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottewell, P.D. The role of osteoblasts in bone metastasis. J. Bone Oncol. 2016, 5, 124–127. [Google Scholar] [CrossRef] [Green Version]
- Virk, M.S.; Lieberman, J.R. Tumor metastasis to bone. Arthritis Res. Ther. 2007, 9 (Suppl. S1), S5. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, P.; Suva, L.J.; Welch, D.R.; Donahue, H.J. Osteoprotegrin and the bone homing and colonization potential of breast cancer cells. J. Cell Biochem. 2008, 103, 30–41. [Google Scholar] [CrossRef]
- Chung, S.T.; Geerts, D.; Roseman, K.; Renaud, A.; Connelly, L. Osteoprotegerin mediates tumor-promoting effects of Interleukin-1 beta in breast cancer cells. Mol. Cancer 2017, 16, 27. [Google Scholar] [CrossRef] [Green Version]
- Fong, Y.C.; Maa, M.C.; Tsai, F.J.; Chen, W.C.; Lin, J.G.; Jeng, L.B.; Yang, R.S.; Fu, W.M.; Tang, C.H. Osteoblast-derived TGF-beta1 stimulates IL-8 release through AP-1 and NF-kappaB in human cancer cells. J. Bone Min. Res. 2008, 23, 961–970. [Google Scholar] [CrossRef]
- Gupta, J.; Robbins, J.; Jilling, T.; Seth, P. TGFβ-dependent induction of interleukin-11 and interleukin-8 involves SMAD and p38 MAPK pathways in breast tumor models with varied bone metastases potential. Cancer Biol. Ther. 2011, 11, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollari, S.; Käkönen, R.S.; Mohammad, K.S.; Rissanen, J.P.; Halleen, J.M.; Wärri, A.; Nissinen, L.; Pihlavisto, M.; Marjamäki, A.; Perälä, M.; et al. Heparin-like polysaccharides reduce osteolytic bone destruction and tumor growth in a mouse model of breast cancer bone metastasis. Mol. Cancer Res. 2012, 10, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollari, S.; Leivonen, S.K.; Perälä, M.; Fey, V.; Käkönen, S.M.; Kallioniemi, O. Identification of microRNAs inhibiting TGF-β-induced IL-11 production in bone metastatic breast cancer cells. PLoS ONE 2012, 7, e37361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keklikoglou, I.; Koerner, C.; Schmidt, C.; Zhang, J.D.; Heckmann, D.; Shavinskaya, A.; Allgayer, H.; Gückel, B.; Fehm, T.; Schneeweiss, A.; et al. MicroRNA-520/373 family functions as a tumor suppressor in estrogen receptor negative breast cancer by targeting NF-κB and TGF-β signaling pathways. Oncogene 2012, 31, 4150–4163. [Google Scholar] [CrossRef] [PubMed]
- Studebaker, A.W.; Storci, G.; Werbeck, J.L.; Sansone, P.; Sasser, A.K.; Tavolari, S.; Huang, T.; Chan, M.W.; Marini, F.C.; Rosol, T.J.; et al. Fibroblasts isolated from common sites of breast cancer metastasis enhance cancer cell growth rates and invasiveness in an interleukin-6-dependent manner. Cancer Res. 2008, 68, 9087–9095. [Google Scholar] [CrossRef] [Green Version]
- Sansone, P.; Ceccarelli, C.; Berishaj, M.; Chang, Q.; Rajasekhar, V.K.; Perna, F.; Bowman, R.L.; Vidone, M.; Daly, L.; Nnoli, J.; et al. Self-renewal of CD133(hi) cells by IL6/Notch3 signalling regulates endocrine resistance in metastatic breast cancer. Nat. Commun. 2016, 7, 10442. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Bedollaì, M.A.; Carretero-Ortega, J.; Valadez-Sánchez, M.; Vázquez-Prado, J.; Reyes-Cruz, G. Chemotactic and proangiogenic role of calcium sensing receptor is linked to secretion of multiple cytokines and growth factors in breast cancer MDA-MB-231 cells. Biochim. Biophys. Acta 2015, 1853, 166–182. [Google Scholar] [CrossRef] [Green Version]
- Remeniuk, B.; King, T.; Sukhtankar, D.; Nippert, A.; Li, N.; Li, F.; Cheng, K.; Rice, K.C.; Porreca, F. Disease modifying actions of interleukin-6 blockade in a rat model of bone cancer pain. Pain 2018, 159, 684–698. [Google Scholar] [CrossRef]
- Yao, B.; Wang, J.; Qu, S.; Liu, Y.; Jin, Y.; Lu, J.; Bao, Q.; Li, L.; Yuan, H.; Ma, C. Upregulated osterix promotes invasion and bone metastasis and predicts for a poor prognosis in breast cancer. Cell Death Dis. 2019, 10, 28. [Google Scholar] [CrossRef]
- Safina, A.; Sotomayor, P.; Limoge, M.; Morrison, C.; Bakin, A.V. TAK1-TAB2 signaling contributes to bone destruction by breast carcinoma cells. Mol. Cancer Res. 2011, 9, 1042–1053. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.; Suva, L.J.; Nicks, K.M.; MacLeod, V.; Sanderson, R.D. Tumor-derived syndecan-1 mediates distal cross-talk with bone that enhances osteoclastogenesis. J. Bone Min. Res. 2010, 25, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Ma, Q.; Ding, N.; Luo, F.; Bai, Y.; Kang, F.; Gong, X.; Dong, R.; Dai, J.; Dai, Q.; et al. IL-11 is essential in promoting osteolysis in breast cancer bone metastasis via RANKL-independent activation of osteoclastogenesis. Cell Death Dis. 2019, 10, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, J.S.; Sharma, A.R.; Nguyen, L.T.; Jagga, S.; Lee, Y.H.; Sharma, G.; Lee, S.S. Lysophosphatidic acid enhances breast cancer cells-mediated osteoclastogenesis. Korean J. Physiol. Pharmacol. 2018, 22, 503–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucharaba, A.; Guillet, B.; Menaa, F.; Hneino, M.; van Wijnen, A.J.; Clézardin, P.; Peyruchaud, O. Bioactive lipids lysophosphatidic acid and sphingosine 1-phosphate mediate breast cancer cell biological functions through distinct mechanisms. Oncol. Res. 2009, 18, 173–184, Erratum in: Oncol. Res. 2009, 18, 357. [Google Scholar] [CrossRef] [PubMed]
- Bolin, C.; Tawara, K.; Sutherland, C.; Redshaw, J.; Aranda, P.; Moselhy, J.; Anderson, R.; Jorcyk, C.L. Oncostatin m promotes mammary tumor metastasis to bone and osteolytic bone degradation. Genes Cancer 2012, 3, 117–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Rouse, C.; Jasper, J.S.; Pendergast, A.M. ABL kinases promote breast cancer osteolytic metastasis by modulating tumor-bone interactions through TAZ and STAT5 signaling. Sci. Signal. 2016, 9, ra12. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.H.; Buhamrah, A.; Schneider, A.; Lin, Y.L.; Zhou, H.; Bugshan, A.; Basile, J.R. Semaphorin 4D Promotes Skeletal Metastasis in Breast Cancer. PLoS ONE 2016, 11, e0150151. [Google Scholar] [CrossRef]
- Shrivastava, R.; Asif, M.; Singh, V.; Dubey, P.; Ahmad Malik, S.; Lone, M.U.; Tewari, B.N.; Baghel, K.S.; Pal, S.; Nagar, G.K.; et al. M2 polarization of macrophages by Oncostatin M in hypoxic tumor microenvironment is mediated by mTORC2 and promotes tumor growth and metastasis. Cytokine 2019, 118, 130–143. [Google Scholar] [CrossRef]
- Nutter, F.; Holen, I.; Brown, H.K.; Cross, S.S.; Evans, C.A.; Walker, M.; Coleman, R.E.; Westbrook, J.A.; Selby, P.J.; Brown, J.E.; et al. Different molecular profiles are associated with breast cancer cell homing compared with colonisation of bone: Evidence using a novel bone-seeking cell line. Endocr. Relat. Cancer 2014, 21, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.C.; Leal, A.C.; Gonçalves-Silva, T.; Mercadante, A.C.; Kestelman, F.; Chaves, S.B.; Azevedo, R.B.; Monteiro, J.P.; Bonomo, A. T cells induce pre-metastatic osteolytic disease and help bone metastases establishment in a mouse model of metastatic breast cancer. PLoS ONE 2013, 8, e68171. [Google Scholar] [CrossRef] [Green Version]
- Templeton, Z.S.; Lie, W.R.; Wang, W.; Rosenberg-Hasson, Y.; Alluri, R.V.; Tamaresis, J.S.; Bachmann, M.H.; Lee, K.; Maloney, W.J.; Contag, C.H.; et al. Breast Cancer Cell Colonization of the Human Bone Marrow Adipose Tissue Niche. Neoplasia 2015, 17, 849–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holen, I.; Nutter, F.; Wilkinson, J.M.; Evans, C.A.; Avgoustou, P.; Ottewell, P.D. Human breast cancer bone metastasis in vitro and in vivo: A novel 3D model system for studies of tumour cell-bone cell interactions. Clin. Exp. Metastasis 2015, 32, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Borsari, V.; Brogini, S.; Torricelli, P.; Cepollaro, S.; Cadossi, M.; Fini, M. A Human 3D In Vitro Model to Assess the Relationship Between Osteoporosis and Dissemination to Bone of Breast Cancer Tumor Cells. J. Cell Physiol. 2017, 232, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Sosnoski, D.M.; Norgard, R.J.; Grove, C.D.; Foster, S.J.; Mastro, A.M. Dormancy and growth of metastatic breast cancer cells in a bone-like microenvironment. Clin. Exp. Metastasis 2015, 32, 335–344. [Google Scholar] [CrossRef]
- Holen, I.; Lefley, D.V.; Francis, S.E.; Rennicks, S.; Bradbury, S.; Coleman, R.E.; Ottewell, P. IL-1 drives breast cancer growth and bone metastasis in vivo. Oncotarget 2016, 7, 75571–75584. [Google Scholar] [CrossRef] [Green Version]
- Tulotta, C.; Lefley, D.V.; Freeman, K.; Gregory, W.M.; Hanby, A.M.; Heath, P.R.; Nutter, F.; Wilkinson, J.M.; Spicer-Hadlington, A.R.; Liu, X.; et al. Endogenous Production of IL1B by Breast Cancer Cells Drives Metastasis and Colonization of the Bone Microenvironment. Clin. Cancer Res. 2019, 25, 2769–2782. [Google Scholar] [CrossRef]
- Jewell, A.N.; Swamydas, M.; Castillo, C.I.; Wyan, H.; Allen, L.D.; McDermott, K.A.; Eddy, J.M.; Dréau, D. The endothelin axis stimulates the expression of pro-inflammatory cytokines and pro-migratory molecules in breast cancer. Cancer Investig. 2010, 28, 932–943. [Google Scholar] [CrossRef]
- Iakovou, I.; Doumas, A.; Badiavas, K.; Mpalaris, V.; Frangos, S.; Farmakis, G. Pain palliative therapy in women with breast cancer osseous metastatic disease and the role of specific serum cytokines as prognostic factors. Cancer Biother. Radiopharm. 2014, 29, 116–123. [Google Scholar] [CrossRef]
- Dean-Colomb, W.; Hess, K.R.; Young, E.; Gornet, T.G.; Handy, B.C.; Moulder, S.L.; Ibrahim, N.; Pusztai, L.; Booser, D.; Valero, V.; et al. Elevated serum P1NP predicts development of bone metastasis and survival in early-stage breast cancer. Breast Cancer Res. Treat. 2013, 137, 631–636. [Google Scholar] [CrossRef]
- Noman, A.S.; Uddin, M.; Chowdhury, A.A.; Nayeem, M.J.; Raihan, Z.; Rashid, M.I.; Azad, A.K.; Rahman, M.L.; Barua, D.; Sultana, A.; et al. Serum sonic hedgehog (SHH) and interleukin-(IL-6) as dual prognostic biomarkers in progressive metastatic breast cancer. Sci. Rep. 2017, 7, 1796. [Google Scholar] [CrossRef]
- Ferreira, A.R.; Bettencourt, M.; Alho, I.; Costa, A.L.; Sousa, A.R.; Mansinho, A.; Abreu, C.; Pulido, C.; Macedo, D.; Vendrell, I.; et al. Serum YB-1 (Y-box binding protein 1) as a biomarker of bone disease progression in patients with breast cancer and bone metastases. J. Bone Oncol. 2017, 6, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Tumminello, F.M.; Badalamenti, G.; Incorvaia, L.; Fulfaro, F.; D’Amico, C.; Leto, G. Serum interleukin-6 in patients with metastatic bone disease: Correlation with cystatin C. Med. Oncol. 2009, 26, 10–15. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, L.; Patanè, S.; Grange, C.; Bussolati, B.; Isella, C.; Fontani, L.; Godio, L.; Cilli, M.; D’Amelio, P.; Isaia, G.; et al. Primary breast cancer stem-like cells metastasise to bone, switch phenotype and acquire a bone tropism signature. Br. J. Cancer 2013, 108, 2525–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinder, M.; Chislock, E.; Bussard, K.M.; Shuman, L.; Mastro, A.M. Metastatic breast cancer induces an osteoblast inflammatory response. Exp. Cell Res. 2008, 314, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Dhurjati, R.; Krishnan, V.; Shuman, L.A.; Mastro, A.M.; Vogler, E.A. Metastatic breast cancer cells colonize and degrade three-dimensional osteoblastic tissue in vitro. Clin. Exp. Metastasis 2008, 25, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Bussard, K.M.; Venzon, D.J.; Mastro, A.M. Osteoblasts are a major source of inflammatory cytokines in the tumor microenvironment of bone metastatic breast cancer. J. Cell Biochem. 2010, 111, 1138–1148. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, V.; Shuman, L.A.; Sosnoski, D.M.; Dhurjati, R.; Vogler, E.A.; Mastro, A.M. Dynamic interaction between breast cancer cells and osteoblastic tissue: Comparison of two- and three-dimensional cultures. J. Cell Physiol. 2011, 226, 2150–2158. [Google Scholar] [CrossRef]
- Sethi, N.; Dai, X.; Winter, C.G.; Kang, Y. Tumor-derived JAGGED1 promotes osteolytic bone metastasis of breast cancer by engaging notch signaling in bone cells. Cancer Cell 2011, 19, 192–205. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, V.; Vogler, E.A.; Sosnoski, D.M.; Mastro, A.M. In vitro mimics of bone remodeling and the vicious cycle of cancer in bone. J. Cell Physiol. 2014, 229, 453–462. [Google Scholar] [CrossRef]
- Bussard, K.M.; Okita, N.; Sharkey, N.; Neuberger, T.; Webb, A.; Mastro, A.M. Localization of osteoblast inflammatory cytokines MCP-1 and VEGF to the matrix of the trabecula of the femur, a target area for metastatic breast cancer cell colonization. Clin. Exp. Metastasis 2010, 27, 331–340. [Google Scholar] [CrossRef]
- Rajski, M.; Vogel, B.; Baty, F.; Rochlitz, C.; Buess, M. Global gene expression analysis of the interaction between cancer cells and osteoblasts to predict bone metastasis in breast cancer. PLoS ONE 2012, 7, e29743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liverani, C.; Mercatali, L.; Spadazzi, C.; La Manna, F.; De Vita, A.; Riva, N.; Calpona, S.; Ricci, M.; Bongiovanni, A.; Gunelli, E.; et al. CSF-1 blockade impairs breast cancer osteoclastogenic potential in co-culture systems. Bone 2014, 66, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Mazaki, A.; Orita, S.; Inage, K.; Suzuki, M.; Abe, K.; Shiga, Y.; Inoue, M.; Norimoto, M.; Umimura, T.; Ohtori, S.; et al. Tumor Necrosis Factor-α Produced by Osteoclasts Might Induce Intractable Pain in a Rat Spinal Metastasis Model of Breast Cancer. Spine Surg. Relat. Res. 2019, 3, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Chow, S.O.; Boernert, K.; Basel, D.; Mikuscheva, A.; Kim, S.; Fong-Yee, C.; Trivedi, T.; Buttgereit, F.; Sutherland, R.L.; et al. Direct crosstalk between cancer and osteoblast lineage cells fuels metastatic growth in bone via auto-amplification of IL-6 and RANKL signaling pathways. J. Bone Min. Res. 2014, 29, 1938–1949. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Hamaguchi, T.; Nagao, N.; Kato, S.; Iino, T.; Nakamura, T.; Sudo, A. Interleukin-6 receptor inhibitor suppresses bone metastases in a breast cancer cell line. Breast Cancer 2018, 25, 566–574. [Google Scholar] [CrossRef]
- Luo, X.; Fu, Y.; Loza, A.J.; Murali, B.; Leahy, K.M.; Ruhland, M.K.; Gang, M.; Su, X.; Zamani, A.; Shi, Y.; et al. Stromal-Initiated Changes in the Bone Promote Metastatic Niche Development. Cell Rep. 2016, 14, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Alho, I.; Costa, L.; Bicho, M.; Coelho, C. Low Molecular Weight Protein Tyrosine Phosphatase Slow Isoform Knockdown in MDA-MB-435 Cells Decreases RAW 264.7 Osteoclastic Differentiation. Anticancer Res. 2016, 36, 2227–2232. [Google Scholar]
- Schwaninger, R.; Rentsch, C.A.; Wetterwald, A.; van der Horst, G.; van Bezooijen, R.L.; van der Pluijm, G.; Löwik, C.W.; Ackermann, K.; Pyerin, W.; Hamdy, F.C.; et al. Lack of noggin expression by cancer cells is a determinant of the osteoblast response in bone metastases. Am. J. Pathol. 2007, 170, 160–175. [Google Scholar] [CrossRef] [Green Version]
- Kamalakar, A.; Bendre, M.S.; Washam, C.L.; Fowler, T.W.; Carver, A.; Dilley, J.D.; Bracey, J.W.; Akel, N.S.; Margulies, A.G.; Skinner, R.A.; et al. Circulating interleukin-8 levels explain breast cancer osteolysis in mice and humans. Bone 2014, 61, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Coonrod, S.; Estroff, L.; Fischbach, C. Chemical and physical properties of carbonated hydroxyapatite affect breast cancer cell behavior. Acta Biomater. 2015, 24, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Pagani, S.; Fini, M.; Giavaresi, G.; Salamanna, F.; Borsari, V. The active role of osteoporosis in the interaction between osteoblasts and bone metastases. Bone 2015, 79, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, J.; Ji, Y.; Hong, A.; Xie, Q. Cytokines in osteoblast-conditioned medium promote the migration of breast cancer cells. Tumour Biol. 2014, 35, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Castro, N.J.; Cui, H.; Zhou, X.; Boualam, B.; McGrane, R.; Glazer, R.I.; Zhang, L.G. A 3D printed nano bone matrix for characterization of breast cancer cell and osteoblast interactions. Nanotechnology 2016, 27, 315103. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Villanueva, D.; Zeef, L.; Shore, P. Metastatic breast cancer cells inhibit osteoblast differentiation through the Runx2/CBFβ-dependent expression of the Wnt antagonist, sclerostin. Breast Cancer Res. 2011, 13, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Hu, Z.; Gupta, J.; Krimmel, J.D.; Gerseny, H.M.; Berg, A.F.; Robbins, J.S.; Du, H.; Prabhakar, B.; Seth, P. Intravenous administration of adenoviruses targeting transforming growth factor beta signaling inhibits established bone metastases in 4T1 mouse mammary tumor model in an immunocompetent syngeneic host. Cancer Gene Ther. 2012, 19, 630–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.L.; Huang, W.D.; Li, B.; Chen, T.R.; Li, Z.X.; Zhao, C.L.; Li, H.Y.; Wu, Y.M.; Yan, W.J.; Xiao, J.R. microRNA-124 inhibits bone metastasis of breast cancer by repressing Interleukin-11. Mol. Cancer 2018, 17, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.; Wang, X.; Dong, Z.; Liu, J.; Zhang, S. Bone metastasis from breast cancer involves elevated IL-11 expression and the gp130/STAT3 pathway. Med. Oncol. 2013, 30, 634. [Google Scholar] [CrossRef]
- McCoy, E.M.; Hong, H.; Pruitt, H.C.; Feng, X. IL-11 produced by breast cancer cells augments osteoclastogenesis by sustaining the pool of osteoclast progenitor cells. BMC Cancer 2013, 13, 16. [Google Scholar] [CrossRef] [Green Version]
- Irawan, C.; Atmakusumah, D.; Siregar, N.C.; Tean, T.B.; Kong, L.W.; Kiat, O.C.; Abdulmuthalib, A.; Harahap, A.; Mansyur, M. Expression of Biomarkers CXCR4, IL11-RA, TFF1, MLF1P in Advanced Breast Cancer Patients with Bone Metastatic: A Diagnostic Study. Acta Med. Indones. 2016, 48, 261–268. [Google Scholar]
- Bottos, A.; Gotthardt, D.; Gill, J.W.; Gattelli, A.; Frei, A.; Tzankov, A.; Sexl, V.; Wodnar-Filipowicz, A.; Hynes, N.E. Decreased NK-cell tumour immunosurveillance consequent to JAK inhibition enhances metastasis in breast cancer models. Nat. Commun. 2016, 7, 12258. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, R.H.; Reagan, M.R.; Anderson, K.; Kaplan, D.L.; Rosenblatt, M. Human bone marrow-derived MSCs can home to orthotopic breast cancer tumors and promote bone metastasis. Cancer Res. 2010, 70, 10044–10050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayir, E.; Aygün, S.; Ata, A.; Arican, A. Relationship Between IL-18 and Bone Metastasis in Female Breast Cancer. Turk. J. Oncol. 2016, 31, 10–14. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Hsing, C.H.; Li, C.F.; Chan, C.H.; Chang, M.C.; Yan, J.J.; Chang, M.S. Anti-IL-20 monoclonal antibody suppresses breast cancer progression and bone osteolysis in murine models. J. Immunol. 2012, 188, 1981–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunda, M.J.; Luistro, L.; Warrier, R.R.; Wright, R.B.; Hubbard, B.R.; Murphy, M.; Wolf, S.F.; Gately, M.K. Antitumor and antimetastatic activity of interleukin 12 against murine tumors. J. Exp. Med. 1993, 178, 1223–1230. [Google Scholar] [CrossRef]
- Setrerrahmane, S.; Xu, H. Tumor-related interleukins: Old validated targets for new anti-cancer drug development. Mol. Cancer 2017, 16, 153. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| References | Interleukin(s) Identified | Interleukin Source | Aims | Experimental Approaches |
|---|---|---|---|---|
| [68] | IL-8 | Breast cancer cells | Evaluate the influence of the expression of IL-8 on bone resorption in osteolytic breast cancer cell lines. | In vitro MDA-MB-231, MDA-231B, T-47D, ZR-75-1 culture. |
| [54] | IL-6 IL-8 | Osteoblasts | Evaluate osteoblasts inflammatory stress response (IL-6, IL-8) in presence of metastatic breast cancer cells. | In vitro hFOB 1.19 co-culture with MDA-MB-231 CM; hFOB 1.19 and MDA-MB-231 co-culture. |
| [18] | IL-1β | Osteoblasts | Evaluate if OPG production by breast cancer cells, pretreated or not with IL-1β, correlates with bone colonization. | In vitro hFOB 1.19 and 435/BRMS1, 231/BRMS1, 231/K and MDA-MB-435 derived from the pleural effusion of a female patient with an infiltrating ductal carcinoma or their CM (pretreated or not with IL-1β) co-culture. |
| [25] | IL-6 | Breast cancer cells | Evaluate fibroblasts isolated from breast cancer metastasis and the link with tumor growth rates. | In vivo and in vitro MCF-7 and BT474 co-cultured with Ped300 and P162 fibroblast cells CM. MCF-7 cells injected s.c. (± fibroblasts) in the dorsal area of BALB/c nude mice |
| [20] | IL-8 | Osteoblasts | Evaluate osteoblast-derived TGF association with osteolytic bone diseases and ILs regulation. | In vitro MDA-231 and MDA-435 co-cultured with MG-63 CM. |
| [55] | IL-6 | Osteoblasts | Evaluate colonization and degradation of osteoblastic tissue by breast cancer cells. | In vitro MDA-MB-231 CM co-cultured with osteoblast tissue. |
| [52] | IL-6 | Peripheral venous blood | Identify clinical significance of serum IL-6 and its correlation with Cyst C in patients. | In vitro Peripheral venous blood from 25 healthy donors,10 patients with age-related osteoporosis, 16 patients with localized breast cancer, 10 patients with breast cancer bone metastases. |
| [34] | IL-8 IL-6 | Breast cancer cells | Evaluate the activity of S1P and LPA on breast cancer cells to stimulate osteoclasts. | In vitro MDA-MB-231/BO2 (MDA- BO2) CM untreated or treated with S1P and LPA. Mice bone marrow cells cultured with MDA-BO2 CM treated with S1P and LPA. |
| [31] | IL-8 | Breast cancer cells | Evaluate the role of Syndecan-1 in osteoclastogenesis. | In vitro Human PBMC: mCSF present in all groups; RANKL or IL-8 used as positive controls. Human PBMCs that received mCSF cultured with CM from MDA MET-derived cell lines HPSE-high, HPSE-low, M225, and M343. For same cultures, Syndecan-1 immunodepleted from CM using monoclonal antibody BB4 and protein-G sepharose beads. |
| [60] | IL-6 | Osteoblasts | Determine the localization of osteoblast-derived IL-6. | In vivo MDA-MB-231 expressing GFP variant injected into the left cardiac ventricle of athymic mice. |
| [56] | IL-6 IL-8 | Osteoblasts | Identify key cytokines expressed by osteoblasts in metastatic breast cancer cells. | In vitro and in vivo MC3T3-E1 co-cultured with MDA-MB-231 variants CM; MDA-MB-231 variants injected into the left cardiac ventricle of athymic mice. |
| [47] | IL-1α IL-17 IL-12 | Breast cancer cells | Evaluate if limiting ET-1 expression and activity (blocking ETAR and ETBR) reduced breast cancer growth and development of tumor infiltration in bone. | In vitro and vivo 4T1 cells injected in C57Bl/6× Balb/C male mice and treated with vehicle) or with ETAR/ETBR dual selective antagonist. 4T1 cells CM treated with ET-1. |
| [57] | IL-6 | Osteoblasts | Evaluate the dynamic interaction between breast cancer cells and osteoblastic tissue. | In vitro MDA-MB-231 and MC3T3-E1 co-culture; MDA-MB-231 inoculated into bioreactor containing osteoblastic tissue. |
| [81] | IL-17B | Bone marrow mesenchymal cells | Evaluate the role of hBMSC in metastatic breast cancer cells. | In vitro and in vivo Migration assay of hBMSC to human BCC (SUM 1315, SUM 1315-BP2, MDA-MB-231 and MCF7), wild type (wt) or over-expressed for IL-17 br, or treated with IL-17B. In vivo metastasis model of hBMSC and breast cancer cells(SUM 1315, SUM 1315-BP2, MDA-MB-231 and MCF7) wt or over-expressed for IL-17 br, co-injected in mice mammary fat pad. |
| [58] | IL-6 | Osteoblasts | Identify if Notch-dependent signaling proteins secreted by osteoblasts may stimulate tumor growth. | In vitro MC3T3-E1 and JAG1 OE breast cancer cells co-culture. |
| [74] | IL-11 | Breast cancer cells | Determine how Runx2 mediates the ability of metastatic breast cancer cells to modulate the activity of bone cells. | In vitro MC3T3-E1 co-cultured with MDA-MB-231 CM or sclerostin-free CM; Mouse BMSCs co-cultured with MDA-MB-231 CM or sclerostin-free CM. |
| [21] | IL-8 IL-11 | Breast cancer cells | Delineate TGF-β signaling pathways in the production of IL-8 and IL-11 in breast cancer cells with different bone metastatic potential. | In vitro MDA-MB-231, MDA-MB-468, and MCF-7 cultured with TGF-β. MDA-MB-231 treated with SMAD4 RNAis or MAPK14 RNAis and low GC content negative control RNAi and treated with TGFβ-1. |
| [30] | IL-1β IL-8 | Breast cancer cells | Identify if TAK1 (TGF-β-activated protein kinase 1) effectors contribute to bone metastatic potential of breast cancer cells. | In vitro Suppression of TAK1 signaling in MDA-MB-231 cells by expressing the dominant-negative TAK1-K63W mutant allele or by siRNA. Control: empty-vector. CM from control and dn-TAK1-expressing MDA-MB-231 cells treated with TGF-β1 or/and IL1-β |
| [61] | IL-6 | Evaluate the clinical relevance of gene expression signatures induced by heterotypic interaction of breast cancer cells and osteoblasts. | In vitro Co-culture of HMECs; MCF-7, T47D, MDA-MB-231, SKBR-3 and Hs578T co-culture with primary human osteoblasts. | |
| [22] | IL-11 | Breast cancer cells | Identify and characterize miRNAs that regulate the TGF-β induction of IL-11 in metastatic breast cancer cells. | In vitro MDA-MB-231(SA) transfected with precursors or miRNA inhibitors and cultured with TGF-β. |
| [23] | IL-11 | Breast cancer cells | Evaluate gene for TGF-β-induced IL-11 production in highly bone metastatic MDA-MB-231(SA) by a cell-based siRNA screen. | In vitro MDA-MB-231(SA) transfected with siRNAs cultured with heparin, fragmin, or K5-NSOS and TGF-β. |
| [24] | IL-6 IL-8 | Breast cancer cells | Evaluate an unbiased genome-wide miRNA screen to identify miRNAs modulating NF-κB signaling in metastatic breast cancer cells. | In vitro MDA-MB-231 transfected with miR-373 or miR-520c and treated with TNF-α or TGF-β. |
| [83] | IL-20 | Tumor tissue Breast cancer cells | Evaluate the function of IL-20 in tumor growth, metastasis, and clinical outcome. | In vitro and in vivo IL-20, IL-20 receptors, and anti–IL-20 mAb 7E immunohistochemical staining of tumor tissue samples with primary intraductal carcinoma of the breast, with breast cancer, with tumorous and non-tumorous breast tissue with breast cancer bone metastases, and breast cancer cell lines (MDA-MB231 and MCF-7); Intracardiac and intratibial injection of 4T1-Luc in BALB/c mice. |
| [49] | IL-6 | Serum | Determine serum level of IL-6 protein clinical utility in patients with breast cancer bone metastasis. | In vitro Blood samples from 164 patients with stage I–III breast cancer. |
| [75] | IL-11 | Breast cancer cells | Evaluate the effect of TGFβ-1 on IL-11 production. | In vitro 4T1 cells exposed to different doses TGFβ-1. |
| [35] | IL-6 (Oncostatin M-OSM) | Breast cancer cells | Evaluate the role of OSM in the formation of bone metastases. | In vitro and in vivo 4T1.2-OSM (OSM expression knocked down using shRNA) and 4T1.2-LacZ (control) injected into the mammary fat pad or intra-tibial in Balb/c mice. RAW 264.7 co-cultured with 4T1.2. |
| [40] | IL-17F IL-1β IL-6 | Breast cancer cells T-cells | Evaluate how the pre-metastatic niches arise in the bone tissue. | In vitro and in vivo Inoculation of non- metastatic 67 NR or metastatic 4T1 BCC in the mammary fat pad of BALB/c mice. T-cells from 4T1 tumor-bearing mice KD for RANKL and IL-17F, and transferred into Balb/c nude mice. |
| [77] | IL-11 | Serum Tumor tissue | Evaluate the relationship between IL-11 and breast cancer bone metastasis in patients. | In vitro Blood samples from patients with bone metastasis, with primary cancer and healthy patients; formalin-fixed, paraffin-embedded tumor tissue (from patients with bone metastasis and from patients with primary cancer); fresh tumor tissue from patients (with primary cancer and with breast cancer metastasis). |
| [78] | IL-11 | Breast cancer cells | Investigate the role of IL-11 in metastasis-induced osteolysis. | In vitro Bone marrow cells from femur and tibia of C56BL/6 mice co-cultured with MDA-MB-231 CM; IL-11 neutralizing antibody added to bone marrow in 20% MDA-MB-231 CM. Bone marrow cultured with IL-11 to generate osteoclasts from IL-11-dependant precursors. |
| [53] | IL-6 IL-8 | Cancer stem cells | Investigate the ability of non-metastatic human breast cancer stem cells to metastasize to bone. | In vivo Primary CD44+ CD24− breast CSCs-like transduced by luciferase-lentiviral vector injected s.c. and intra-cardiac in immunodeficient mice carrying s.c. human bone implants. Mammospheres derived from patient tumor specimens. |
| [59] | IL-6 | Osteoblasts | Determine whether 3D mineralizing tissue would be a bone surrogate for studying the early stages of breast cancer colonization to bone. | In vitro MC3T3-E1 (grown for 60 days in bioreactor to form a 3D collagenous osteoblastic tissue), pre-osteoclasts (obtained from bone marrow cells harvested from 6- to 9-week-old GFP mice) and MDA-MB-231-GFP. |
| [64] | IL-6 | Breast cancer cells | Investigate the effects of RANKL on cancer cells and the role of tumor-derived IL-6 within the bone microenvironment. | In vitro and in vivo. Normal or modified (silenced IL-6 and RANK via a lentiviral-based system) MDA-MB-231 injected intra-tibial or s.c. in e BALB/c nu/nu mice. |
| [39] | IL-1β | Breast cancer cells | Identify changes in gene and protein expression associated with bone-homing or colonization, developing a novel bone-seeking clone of MDA-MB-231 cells. | In vitro and in vivo Intra-cardiac injection of MDA-P cells in female Balb/c nude mice. Cells that formed colonies in mouse tibiae were extracted and pooled before culture in vitro. Following 1 week of culture, cells were re-injected into the mice left cardiac ventricle or tail vein. Procedure repeated 7 times until tumor growth in mouse long bones was detected following intravenous injection (MDA-IV). |
| [62] | IL-6 | Breast cancer cells | Assess the effect of soluble mediators produced by breast cancer cells on human osteoclast maturation. | In vitro Differentiation toward osteoclasts (from healthy volunteer) induced by α-MEM supplemented with MCS-F and RANKL or by α-MEM supplemented with 10% MDA-MB-231 (CM). |
| [69] | IL-8 | Plasma | Identify IL-8 association with increased bone resorption and breast cancer bone metastasis. | In vitro and in vivo Plasma from patients with or without bone metastasis from breast cancer; MDA-MET injected into the tibia of nude mice and 7 days later treated daily with a neutralizing IL-8 monoclonal antibody. |
| [48] | IL-2 IL-6 | Blood | Evaluate the possible relationship between RPT efficacy and cytokines levels. | In vitro Blood samples from patients with bone metastases. |
| [72] | IL-8 | Osteoblasts | Test the cell migration stimulated by OCM. | In vitro MCF-7 cultured with hFOB1.19 human osteoblasts CM. |
| [44] | IL-1β | Osteoblasts | Test if bone remodeling cytokines could stimulate dormant cells to grow. | In vitro MDA-MB-231BRMS1 and MC3T3-E1 co-cultured in a long term 3D system. |
| [71] | IL-8 | Osteoblasts | Evaluate the link between osteoblasts and breast cancer cells in healthy and osteoporotic conditions. | In vitro Osteoblasts isolated from trabecular bone of iliac crest of SHAM and OVX rats co-cultured with MRMT-1 rat breast cancer cells CM. |
| [41] | IL-1β | Breast cancer cells | Identify parameters of human bone tissue associated with breast cancer cell osteotropism and colonization. | In vitro MDA-MB-231-fLuc-EGFP and MCF-7-fLuc-EGFP co-cultured with cancellous bone tissue fragments from patients. |
| [26] | IL-6 | Breast cancer cells | Evaluate the association between HT and breast cancer bone metastases. | In vitro and in vivo Tumor-derived cancer cells from xenografts and primary human ductal carcinoma tissues. Xenografts from tumorigenic MCF7 clones administered with tamoxifen citrate implants, fulvestrant and tocilizumab. Cancer cells injected bilaterally in the mammary fat pads of NOD/SCID mice. |
| [27] | IL-6 | Breast cancer cells | Evaluate the ability of CaSR to play a chemotactic and pro-angiogenic role in MDA-MB-231 breast cancer cells by cytokines secretion. | In vitro MDA-MB-231 and MCF-12A CM stimulated with different Ca2+ concentrations. |
| [42] | IL-1β | Breast cancer cells | Generate a clinically relevant model for the study of breast cancer tumor cell-bone interactions. | In vitro and in vivo Human bone discs from patients and DiD labeled MDA-MB-231-luc2 co-culture; tumor-cell bearing bone discs implanted s.c. into the flanks of NOD SCID nude mice; MDA-MB-231 luc2 injected into the hind mammary fat pads of NOD SCID nude mice. |
| [70] | IL-8 | Breast cancer cells | Investigate breast cancer cell IL-8 expression in response to systematic changes of HA. | In vitro MDA-MB-231, MCF-7, MCF10A, MCF10AT1, and MCFDCIS breast cell seeded onto mineral coatings Poly (d,l-lactide-co-glycolide) PLG. |
| [66] | IL-6 | Osteoblasts | Study reactive senescent osteoblasts and evaluate if they increased breast cancer colonization to bone. | In vitro and in vivo NT2.5 and BMMs co-cultured with senescent or non-senescent FASST osteoblasts CM + IL-6 neutralization antibody or IgG control; senescent vs. control osteoblasts co-cultured with mice BMMs. NT2.5luc injected into the left cardiac ventricle of FASST or littermate control mice that received either anti-murine IL-6 antibody or anti-murine IgG2a antibody. |
| [67] | IL-6 IL-8 | Breast cancer cells | Evaluate the effect of LMW-PTP slow isoform in tumor cell induced osteoclastogenesis | In vitro MDA-MB-435 KD for LMW-PTP co-cultured with Raw 264.7. |
| [82] | IL-18 | Serum | Compare serum IL-18 levels in breast cancer patients with and without bone metastases | In vitro Blood samples from female breast cancer patients with or without bone metastases and from healthy subjects. |
| [79] | IL-11RA | Tumor tissue | Analyze expression of IL11-RA in advanced breast cancer patients with or without bone metastasis. | In vitro Human tissue samples from breast cancer patients with or without bone metastases. |
| [80] | IL-15 | Tumor tissue Breast cancer cells | Study JAK/STAT pathway in the bone metastasis from breast cancer. | In vitro and in vivo Breast cancer and bone metastases biopsies from patients. Intra-mammary and intra-cardiac injection of MDA-MB-231 scp1833 and EO771 cells in nude mice. |
| [45] | IL-1 | Breast cancer cells | Investigate the blocking IL-1R signaling with the clinically licensed antagonist, i.e., anakinra. | In vitro and in vivo Co-culture of MDA-MB-231-IV (eGFP-expressing bone-homing derivative of MDA-MB-231) or MCF7 with HS5 human bone marrow cells. Mice pretreated with anakinra or placebo starting 3 days before injection of MCF7 or MDAMB- 231-IV cells s.c. or i.v. via the lateral tail vein; MDA-MB-231-IV cells injected s.c. or i.v. 7 days prior to commencement of anakinra treatment. Mice injected with MCF7 cells were supplemented with B-estradiol. |
| [50] | IL-6 | Serum | Evaluate serum levels of IL-6 in breast cancer patients with or without metastasis. | In vitro Blood samples from patients with breast cancer, with metastatic breast cancer and from healthy subjects. |
| [43] | IL-6 IL-1β IL-10 IL-8 | Breast cancer cells | Evaluate the relationship between osteoporosis and breast cancer-derived bone metastases in a humanized 3D model. | In vitro Bone tissue samples from healthy and osteoporotic patients co-cultured with MCF-7. |
| [36] | IL-6 | Breast cancer cells | Identify tumor-secreted cytokines regulated by the ABL kinases that promote breast cancer metastasis to bone. | In vitro CM of 1833 (bone metastasis sub-line derived from MDA-MB-231 breast cancer cells) cells transduced with control shRNA (Scr) or shRNAs against ABL1 and ABL2 (shAA). The Cancer Genome Atlas dataset analyses. |
| [37] | IL-8 | Osteoblasts | Evaluate Sema4D-mediated induction of IL-8 and LIX/CXCL5. | In vitro and in vivo HOB and MC3T3 CM cultured with Sema4D. HOB cultured with CM by MDA-MB-231 or cells silenced for Sema4D, with and without Clostridium botulinum toxin C3. RAW264.7 cultured with RANKL, IL-8, or CM by HOB treated with sSema4D or empty vector transfected controls and anti-IL-8 antibody. MDA-MB-231 infected ex vivo with control virus or lentivirus coding for Sema4D shRNA injected into the left cardiac ventricle of nude mice. |
| [73] | IL-8 | Breast cancer cells | Investigate the interaction between breast cancer cell and osteoblasts in a 3D printed bone matrices. | In vitro hFOB and MDA-MB-231 cells co-cultured on bone matrix. |
| [51] | IL-6 | Blood | Assess the correlation between sYB-1 and serum IL-6 in patients with breast cancer bone metastasis. | In vitro Peripheral blood from patients with breast cancer bone metastasis. |
| [19] | IL-1β | Breast cancer cells Macrophages | Evaluate the link between OPG, macrophages, and IL-1β. | In vitro MDA-MB-231, MDAMB-436, BT549, SKBR3, ZR75-1, HCC1954 human breast cancer cells transfected with OPG Stealth RNA siRNA; Breast cancer cells and THP-1 monocyte cells (previously treated with PMA to induce macrophage differentiation) co-culture. |
| [65] | IL-6 | Breast cancer cells | Study the effects of IL-6 receptor on breast cancer aggressiveness and bone metastases. | In vitro and in vivo MDA-MB-231 cultured in presence or absence of tocilizumab (anti-human IL-6 receptor (IL-6R). Intracardiac injection of MDA-MB-231 in BALB/c nu/nu mice. |
| [28] | IL-6 | Breast cancer cells | Evaluate the inhibition of IL-6 signaling using a molecule antagonist, TB-2-081, on bone integrity, tumor progression, and pain. | In vitro and in vivo MAT B III cells pretreated with either vehicle or TB-2-081. Intratibial injection of MAT B III in Fisher F344/NhSD rats and treatment with either vehicle or TB-2-08. Osteoblasts harvested from 24- to 48-hour-old neonatal Fischer F344/NhSD pups exposed to vehicle or TB-2-081 pretreatment before challenge of IL-6 recombinant protein. |
| [76] | IL-11 | Breast cancer cells | Investigate miR-124/IL-11 in the prognosis of breast cancer patients with bone metastasis. | In vitro and in vivo MDA-MB-231 cells transfected with miR-124 mimic (miR-124) and MCF7 cells transfected with miR-124 inhibitor (in-miR-124). MCF7 stably expressing miR-124 inhibitor or NC inoculated into left ventricle of nude mice. |
| [38] | IL-6 (Oncostatin M-OSM) | Macrophage | Evaluate OSM induced macrophage M2 polarization during breast cancer bone metastasis. | In vitro and in vivo MDA-MB-231 CM cultured under hypoxia or normoxia with THP-1 cells differentiated to macrophages. BALB/c mice s.c. injected with 2 doses of 4T1 cells in the T4 of mammary fat pad. Mouse PBMCs transfected with the siRNA targeted against Rictor and quantum dots (Q-Dots) before being reintroduced in mice. Macrophages isolated from tumor tissue. |
| [33] | IL-8 IL-11 | Breast cancer cells | Evaluate the role of LPA in the regulation of osteoclastogenic cytokines from breast cancer cells. | In vitro MDA-MB-231 CM and MDA-MB-468 containing LPA co-cultured with RAW264.7 cells |
| [32] | IL-11 | Breast cancer cells | Evaluate the role of IL-11 in bone metastasis from breast cancer. | In vitro and in vivo MCF-7, MDA-MB-231 and BoM-1833 CMs (infected with UBI-MCS-EGFP-SV40-Firefly-Luciferase-IRES Puromycin) co-cultured with BMMs (transfected with siRNA against mouse STAT3, c-Myc, and control si-RNA). Block of osteolytic factors in BoM-1833 cells CM through monoclonal antibodies against VEGF, PTHrP, IL-11, and CTGF. MCF-7, MDA-MB-231, and BoM-1833 injected into femur of BALB/c-nu/nu nude mice. STAT3 inhibitor AG-490 i.v. injected every 2 days for 3 weeks. |
| [63] | IL-6 | Breast cancer cells | Evaluate TNF-α and IL-6 in the pathophysiology of pain syndrome in breast cancer bone metastasis. | In vivo Tumor section, obtained from Sprague-Dawley rats injected s.c. with CRL-1666 breast cancer cells, implanted in L6 vertebra bone defect. |
| [46] | IL-1β | Breast cancer cells | Identify how tumor cell derived IL-1β drives breast cancer progression and bone metastasis; effect of targeting the IL-1β pathway using the anti-human IL-1β antibody, canakinumab, and IL-1R antagonist, anakinra. | In vitro and in vivo Patient tissues with stage II/III breast cancer. MDA-MB-231, MCF-7, and T47D cells (transfected to over-express IL-1β or IL-1R1) co-cultured with OB1. MDA-MB-231 or T47D cells co-cultured with human bone discs. Human bone discs implanted s.c. into NOD SCID mice; after 4 weeks MDA-MB-231 Luc2-TdTomato, MCF7 Luc2, or T47D Luc2 injected into hind mammary fat pads. IL-1Ra or canakinumab s.c. in NOD SCID mice, starting 7 days after injection of tumor cells. IL-1Ra administered for 21 or 31 days or canakinumab administered as a single s.c. injection in BALB/c and C57BL/6 mice. Injection of MDA-MB-231 GFP, MDA-MB-231 IV, MDA-MB-231 IL-1B+, or MDA-MB-231 IL-1R1+ cells into tail vein of BALB/c nude mice or following intra-ductal injection of E0771 into mammary ducts of IL-1B-KO or fl/fl. |
| [29] | IL-8 | Breast cancer cells | Evaluate the knockdown of Osx in breast cancer bone metastasis. | In vivo Transfected MDA-MB 231 cells with stable Osx knockdown or Osx over-expression, injected into the tibiae of nude mice. |
| Trial Number and Status | Aim of the Trial | Trial Arms | Interleukin(s) Identified and Dosage | Function of Interleukin(s) | Summary of Main Trial Findings |
|---|---|---|---|---|---|
| NCT00006228 Completed | Study the effectiveness of trastuzumab + IL-2 in patients with metastatic breast cancer that did not respond to trastuzumab therapy alone. | Trastuzumab IV and aldesleukin SC. | IL-2 Trastuzumab over 30–90 min on days 1 and 8 and aldesleukin on days 2–7 and 9–21. Beginning on day 22, trastuzumab over 30 min every 14 days and aldesleukin daily on days 1–14. Treatment continues for 1 year. | IL-2 may stimulate patients white blood cells to kill breast cancer cells. | No results posted |
| NCT00001269 Completed | Determine the maximal tolerated dose of IL-3 given alone or sequentially with GM-CSF following FLAC CT in metastatic breast cancer patients. | IL-3 alone or sequentially with GM-CSF. | IL-3 Dosage not reported. | IL-3 and GM-CSF may ameliorate cumulative thrombocytopenia after FLAC CT. | No results posted |
| NCT00004893 Completed | Determine the activity of IL-12 as defined by the % of patients who did not progress after 6 months of therapy. | Arm I: Patients begin therapy no sooner than 3 and no later than 6 weeks since last CT dose and receive IL-12 SC twice a week. Patients are followed every 3 months for 1 year, and, if no progression, for 5 yrs. Arm II: Patients are observed for 6 months and, if disease progresses or not, they may receive IL-12 as in arm I. Patients are followed for toxicity until IL-12 is discontinued. | IL-12 Dosage not reported. | IL-12 may kill tumor cells by stopping blood flow to the tumor and by stimulating patients white blood cells to kill breast cancer cells. | No results posted |
| NCT00003412 Study completion date passed, and status not verified in more than 2 years | Study the effectiveness of IL-12 in women with metastatic breast cancer who have received high-dose CT, and peripheral stem cell transplantation. | Arms not reported. | IL-12 Dosage not reported. | IL-12 may kill tumor cells by stopping blood flow to the tumor and by stimulating patients white blood cells to kill breast cancer cells. | No results posted |
| NCT00849459 Completed | Determine toxicity and maximum tolerated dose of intratumoral injection of ADV-hIL12 gene in women with metastatic breast cancer. | ADV-hIL12 | IL-12 Starting dose of ADV-hIL12 to 1 × 10 to the 10th power vp per patient, escalation in half-log increments up to 1 × 10 to the 13th power vp per patient, dose escalation at lower increments of 2 × 10 to a maximum of 3.0 × 10 to the 13th power vp per patient. | Placing the gene for IL-12 into breast cancer cells may help the body build an immune response to kill tumor cells. | No results posted |
| NCT00002616 Study completion date passed, and status not verified in more than 2 years | Study the effectiveness of IL-2 + G-CSF to stimulate cell production in patients with stage IIIB, stage IV, metastatic, or recurrent breast cancer to undergo peripheral stem-cell transplantation. | Arm I: G-CSF for priming and following stem cell transplant. Arm II: G-CSF for priming and G-CSF + IL-2 following transplant. Arm III: G-CSF + IL-2 for priming and G-CSF following transplant. Arm IV: G-CSF + IL-2 for priming and following transplant. Cohorts of 3–6 patients each treated on each arm and at escalating doses of IL-2. | IL-2 Dosage not specified. | Estimate the maximum tolerated dose of continuous infusion IL-2 combined with a dose of G-CSF to stimulate PBSC for harvest in patients with advanced breast cancer. | No results posted |
| NCT00001270 Completed | Define the toxicity of IL-1 administered for 7 days prior to ICE CT. | Arms not reported. | IL-1 Dosage not specified. | IL-1 toxicity. | No results posted |
| NCT01368107 Completed | Evaluate the impact of an immunotherapy by IL-7 on CD4 lymphopenia (CYT107), risks of severe hematological toxicity and tumor progression in metastatic breast cancer patients. | Arm I: Placebo comparator; Arm II: CYT107 SC before CT; Arm III: CYT107 during CT; Arm IV: CYT107 before and during CT. | IL-7 Arm I: Placebo before the 1st (D0, D7, D14) and during the 3rd CT cycle (D57, D64, D71); Arm II: CYT107 (10 µg/kg/week for 3 weeks) before the 1st cycle and placebo during the 3rd cycle; Arm III: placebo before the 1st cycle and CYT107 during the 3rd cycle; Arm IV: CYT107 before the 1st cycle and IL-7 (10µg/kg/week SC for 3 weeks) during the 3rd cycle. | Immunotherapy by IL-7 on CD4 lymphopenia. | No results posted |
| NCT02627274 Recruiting | Evaluate safety, tolerability, pharmacokinetics, pharmacodynamics, and preliminary antitumor activity of RO6874281, an immunocytokine consisting of IL-2v targeting FAP, as single agent or in combination with trastuzumab or cetuximab. | Arm I: RO6874281 monotherapy; Arm II: RO6874281 + trastuzumab; Arm III: RO6874281 + cetuximab. | IL-2v Arm I: 5 mg of RO6874281 as single agent OW; Arm II: 10 mg of RO6874281 + trastuzumab (loading dose 4 mg/kg and maintenance dose 2 mg/kg from cycle 2) OW; Arm III: 5 mg of RO6874281 + cetuximab (loading dose 400 mg/m2 and maintenance dose 250 mg/m2 from cycle 2) OW. | Not specified | Not applicable |
| NCT00002780 Unknown | Evaluate the toxicities of low-dose IL-2 and GM-CSF, and multiple doses of activated T-cells following PBSC transplantation in women with stage IIIB or metastatic adenocarcinoma of the breast. | Patients receive GM-CSF SC daily for 5 days prior to PBSC collection or for 2 days prior to bone marrow harvest. Following collection, patients receive high dose CT on days −6 and −3. Patients undergo PBSC transplantation on day 0. Following transplantation, patients receive multiple doses of monoclonal antibody OKT3 activated T lymphocytes IV between days 1 and 65, continuous low dose IL-2 IV over days 1–65, and GM-CSF on days 5–21. | IL-2 Dosage not specified. | Evaluate if T-cells and IL-2 combined with PBSC transplantation or bone marrow transplantation in women who have stage IIIB or metastatic breast cancer may kill more tumor cells. | No results posted |
| NCT03135171 Recruiting | Determine the highest dose level of anti-IL-6R (tocilizumab) that, when given in combination with trastuzumab and pertuzumab every 3 weeks in subjects with HER2-positive metastatic breast cancer, results in less than 25% incidence of dose limiting toxicity. | Trastuzumab, pertuzumab, and tocilizumab. | Anti-IL-6R Trastuzumab: all dose levels receive 8 mg/kg loading dose for cycle 1 and 6 mg/kg in subsequent cycles, every 3 weeks. Pertuzumab: dose levels 2 and 3 receive 840 mg loading dose for cycle 1 and 420 mg in subsequent cycles, every 3 weeks. Tocilizumab: 4–8 mg/kg administered IV every 3 weeks. | Safety, tolerability, and recommended phase 2 dose of tocilizumab. | No results posted |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamanna, F.; Borsari, V.; Contartese, D.; Costa, V.; Giavaresi, G.; Fini, M. What Is the Role of Interleukins in Breast Cancer Bone Metastases? A Systematic Review of Preclinical and Clinical Evidence. Cancers 2019, 11, 2018. https://doi.org/10.3390/cancers11122018
Salamanna F, Borsari V, Contartese D, Costa V, Giavaresi G, Fini M. What Is the Role of Interleukins in Breast Cancer Bone Metastases? A Systematic Review of Preclinical and Clinical Evidence. Cancers. 2019; 11(12):2018. https://doi.org/10.3390/cancers11122018
Chicago/Turabian StyleSalamanna, Francesca, Veronica Borsari, Deyanira Contartese, Viviana Costa, Gianluca Giavaresi, and Milena Fini. 2019. "What Is the Role of Interleukins in Breast Cancer Bone Metastases? A Systematic Review of Preclinical and Clinical Evidence" Cancers 11, no. 12: 2018. https://doi.org/10.3390/cancers11122018