
Mechanical Characterization and Modelling of Subcellular Components of Oocytes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
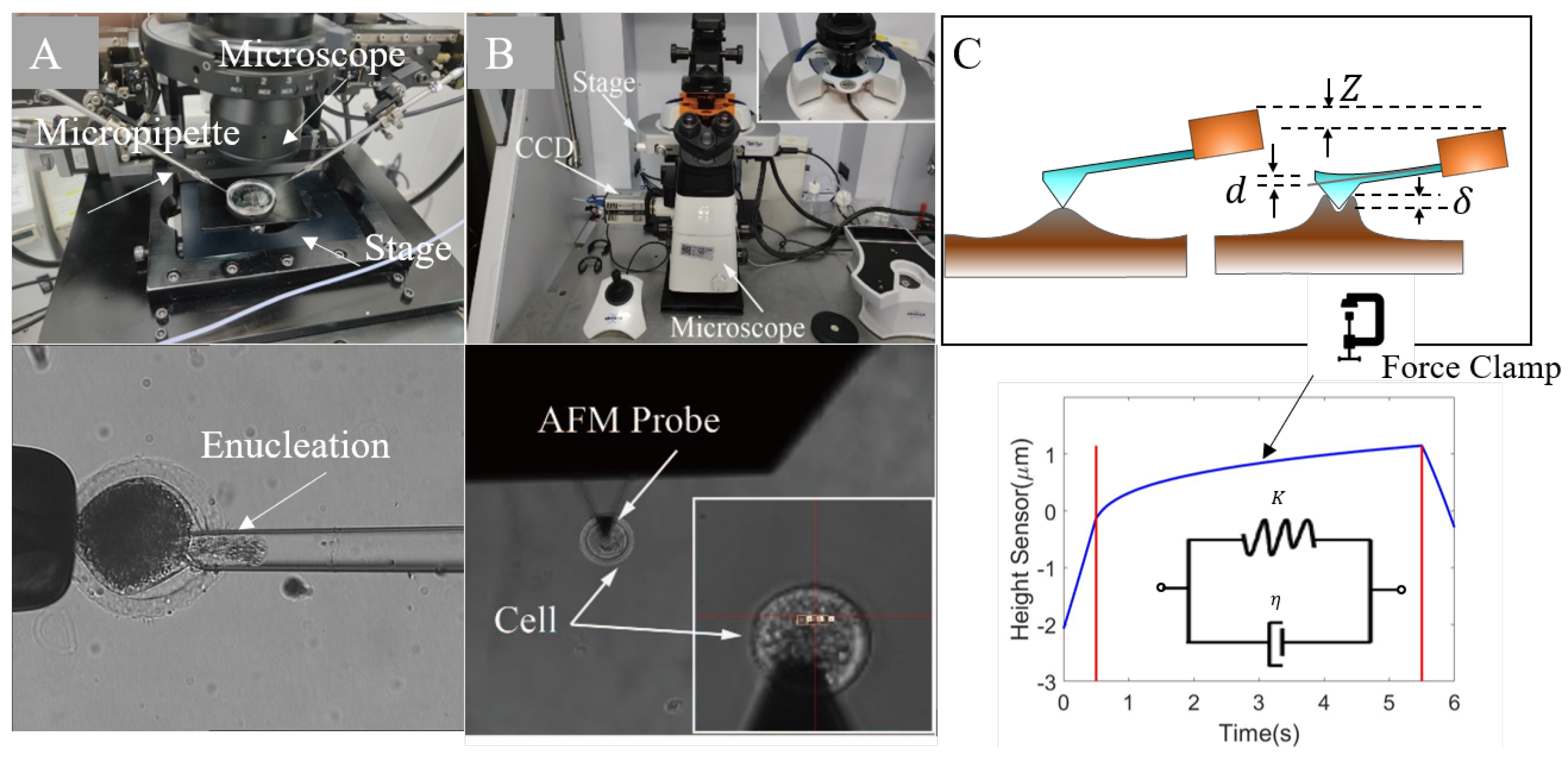
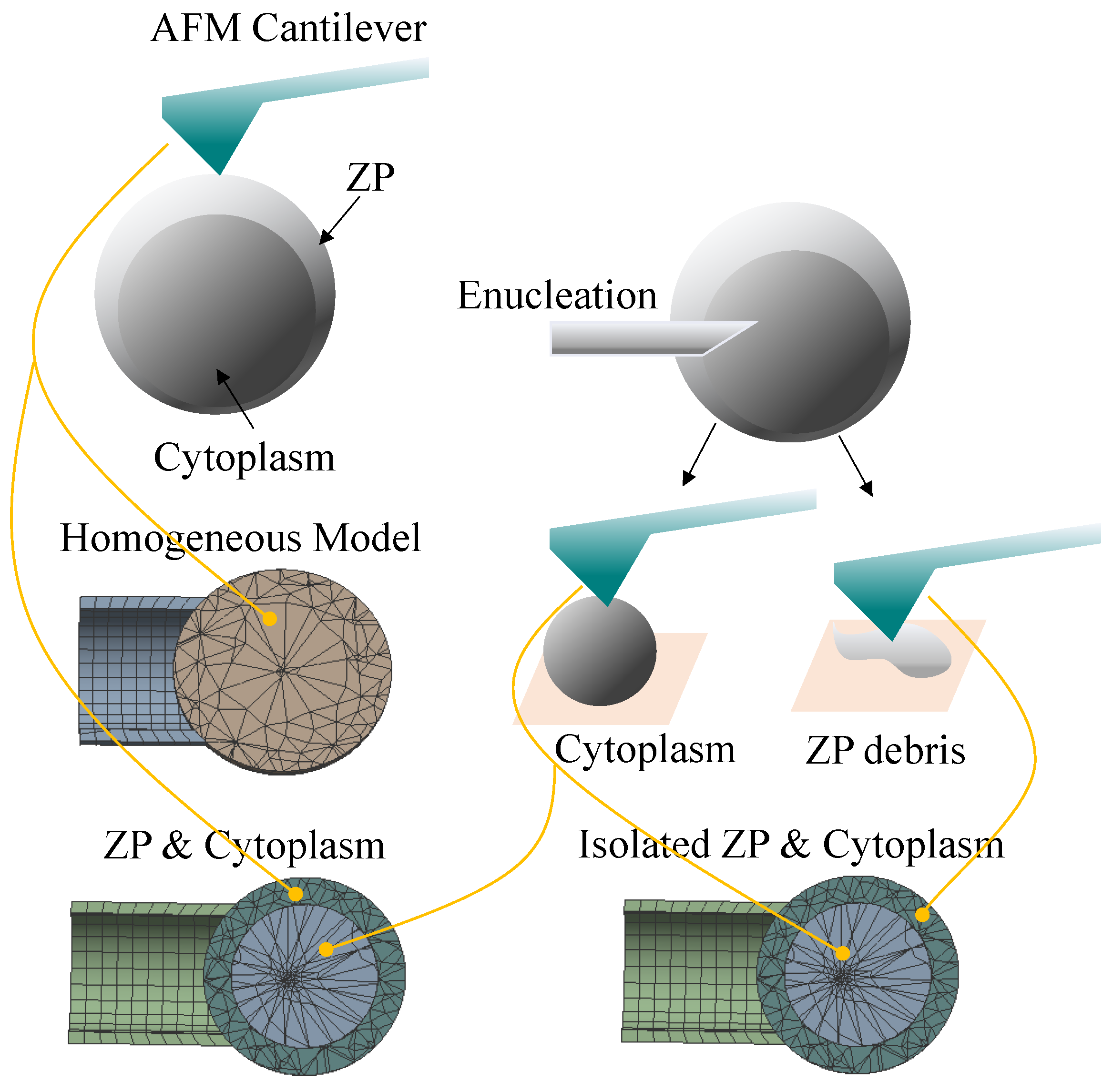
2.1. Preparation for Mechanical Characterization of Subcellular Components
2.2. The Setup of AFM Measurements
2.3. The FEM Oocyte Model
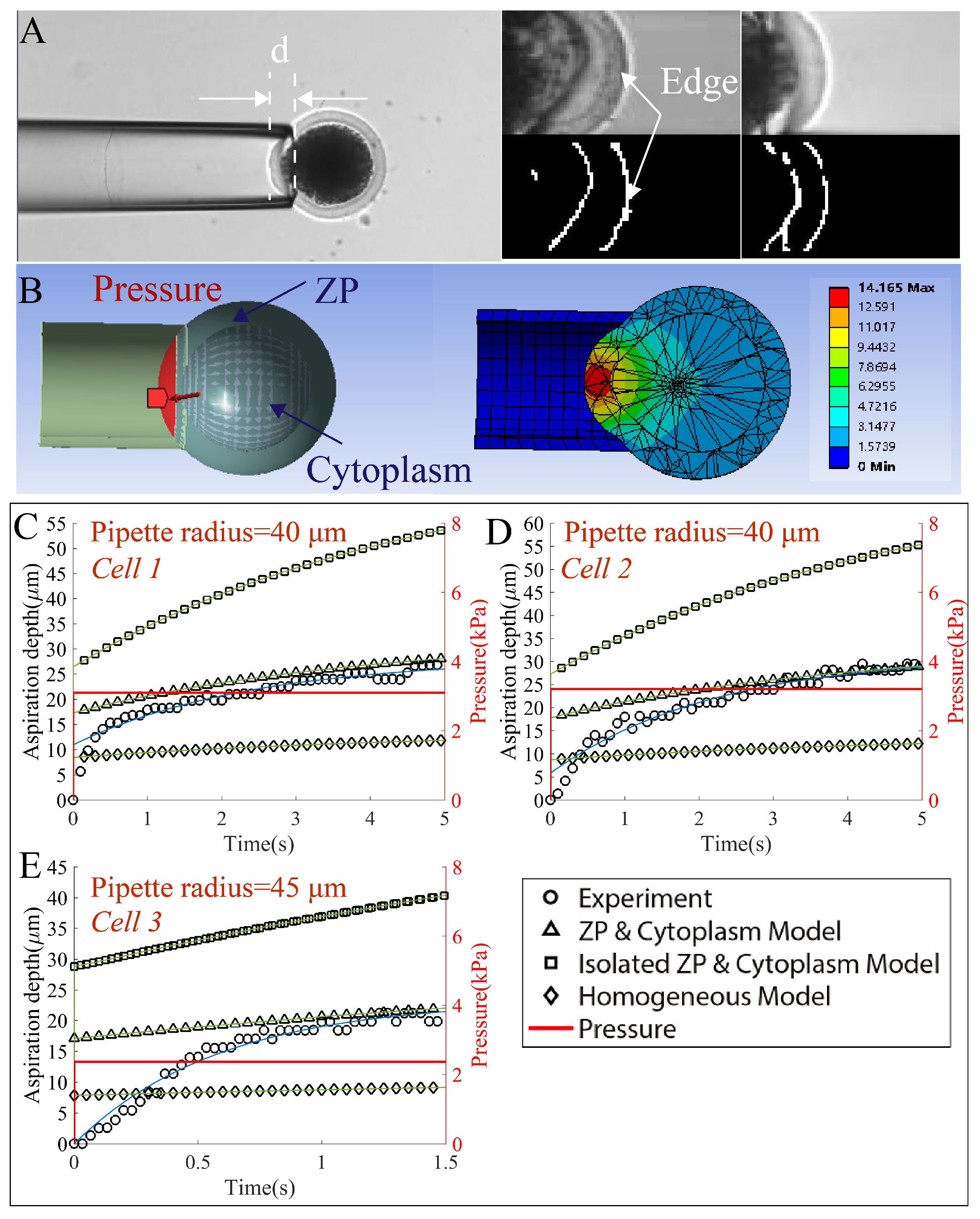
2.4. Validation Experiment I—Micropipette Aspiration Experiment
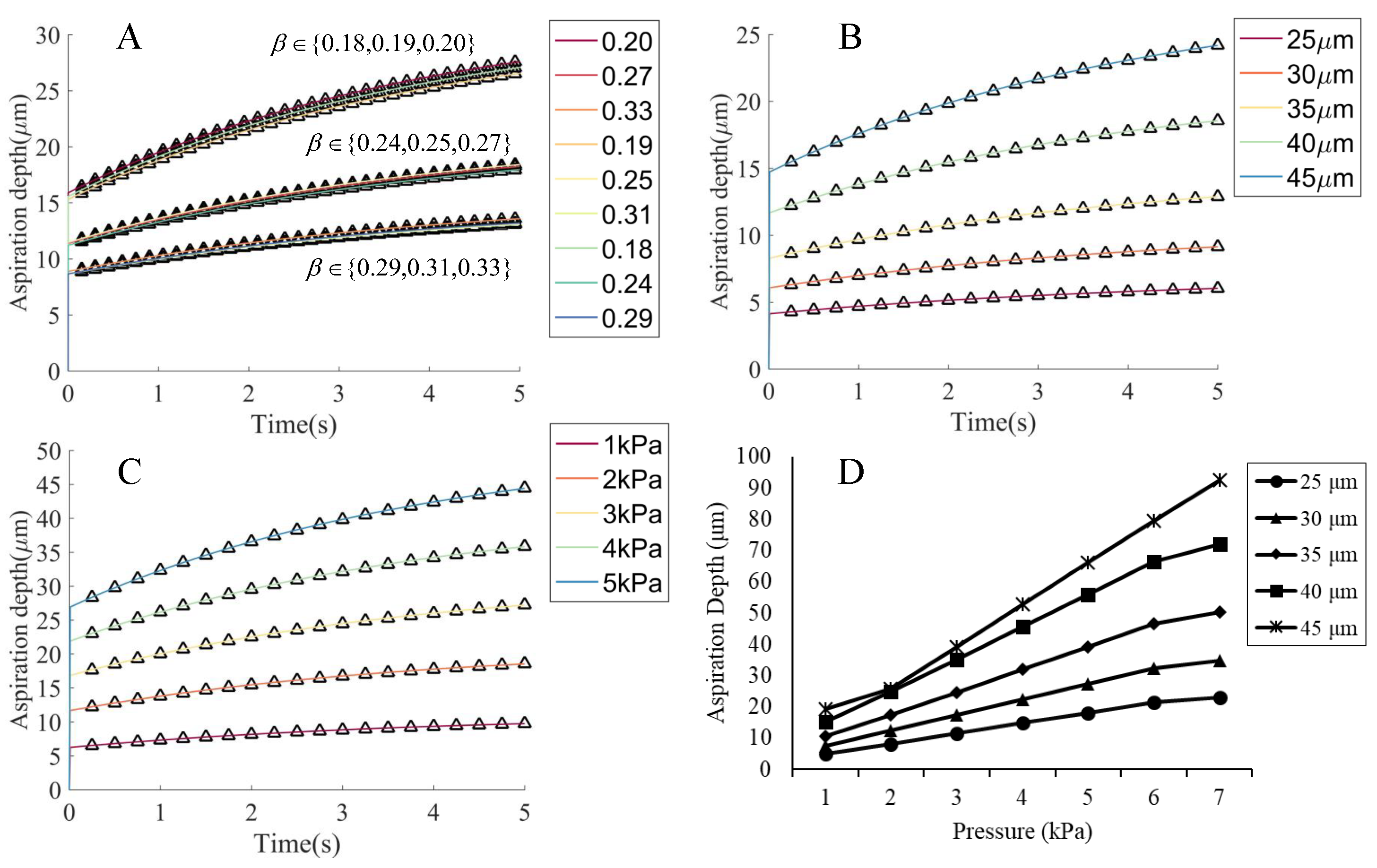
2.4.1. The Simulation Settings for Micropipette Aspiration
2.4.2. The Subpixel Detection of Cell Edge for Micropipette Aspiration
2.5. Validation Experiment II—Microinjection Experiment
2.5.1. The Simulation Settings for Microinjection Experiment
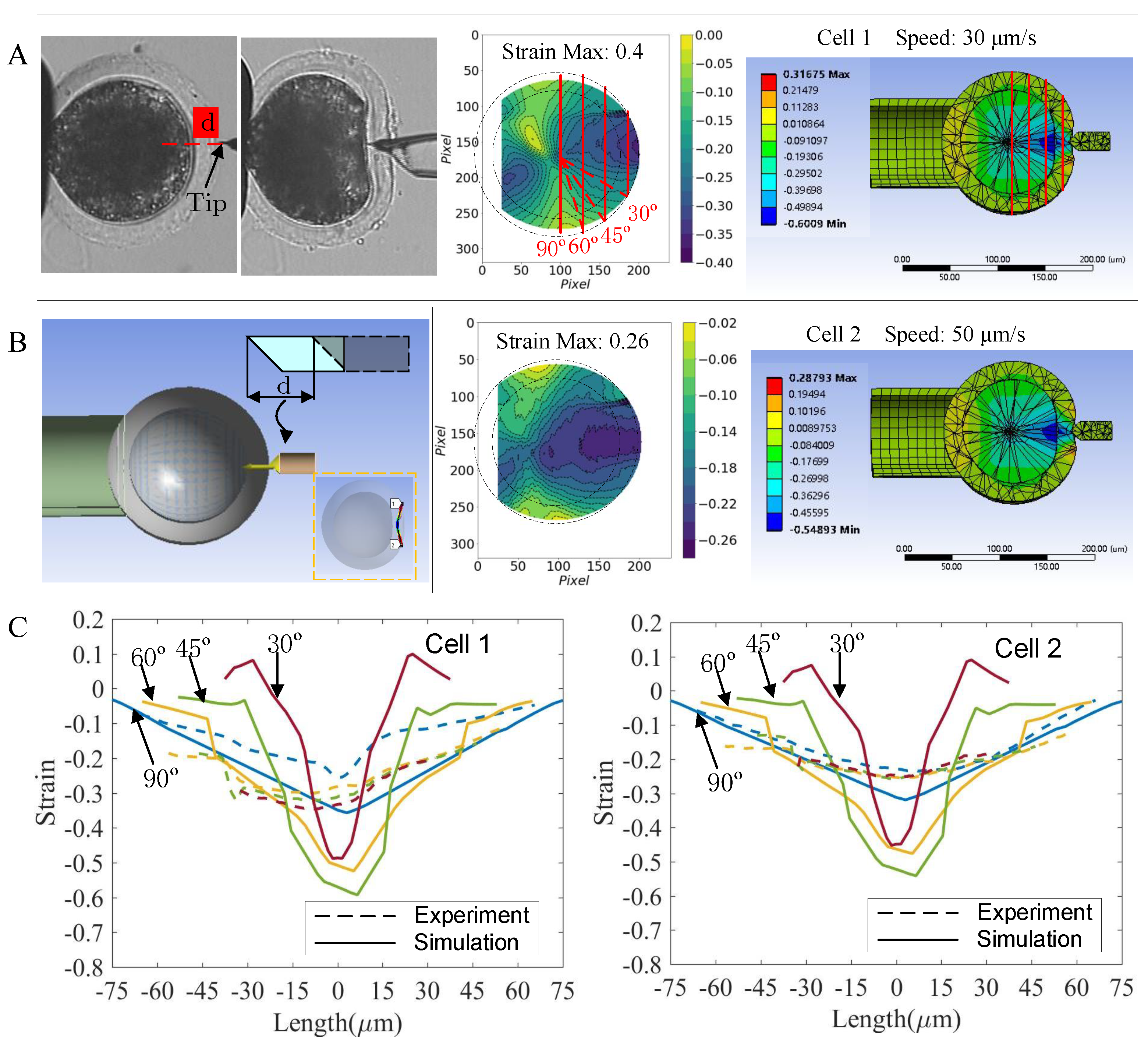
2.5.2. Detection of Microinjection-Induced Intracellular Strain from Microscopic Images
3. Experimental and Simulation Results
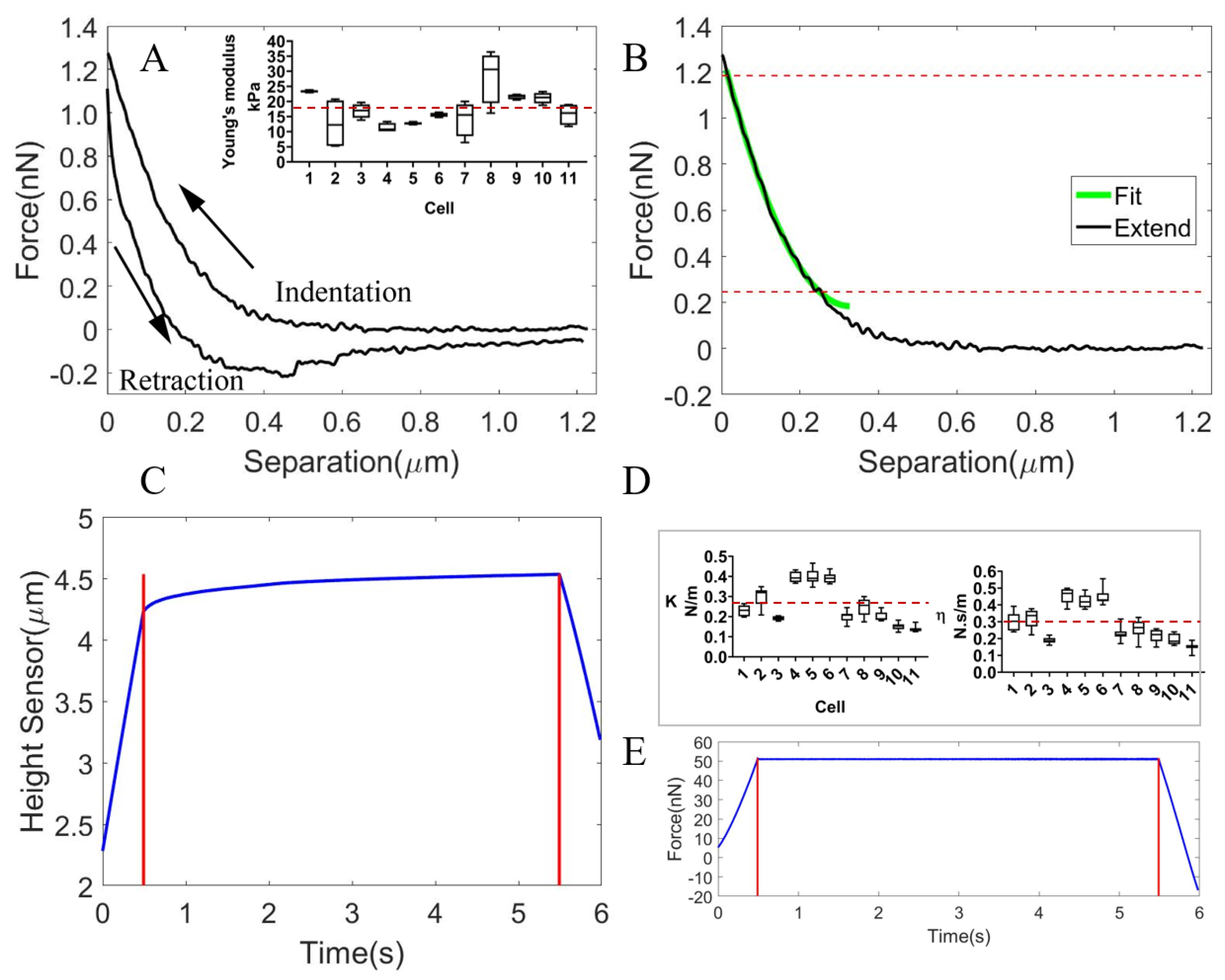
3.1. The Mechanical Characterization of the Subcellular Components of Oocytes
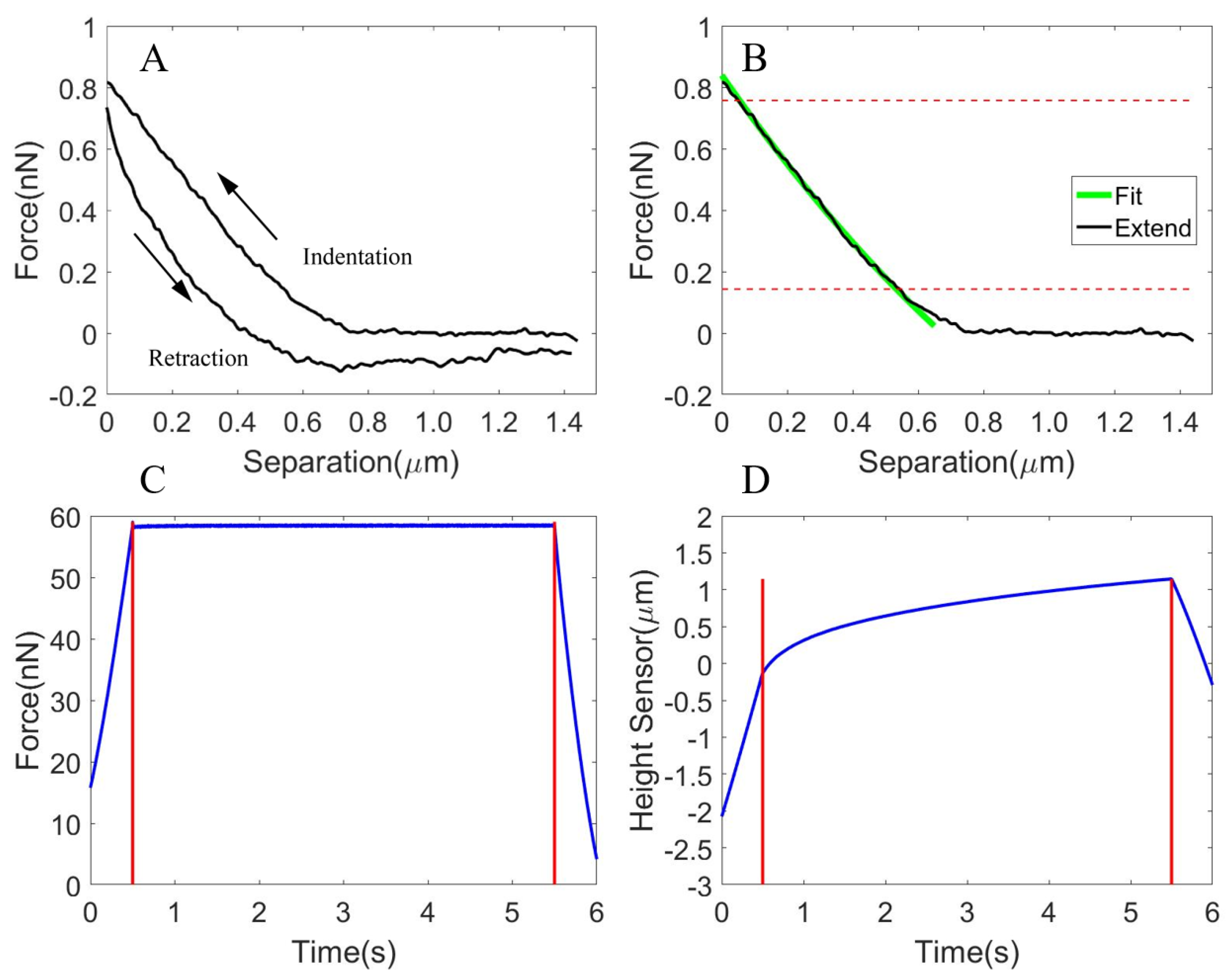
3.1.1. The Viscoelastic Properties of the Oocyte Cytoplasm
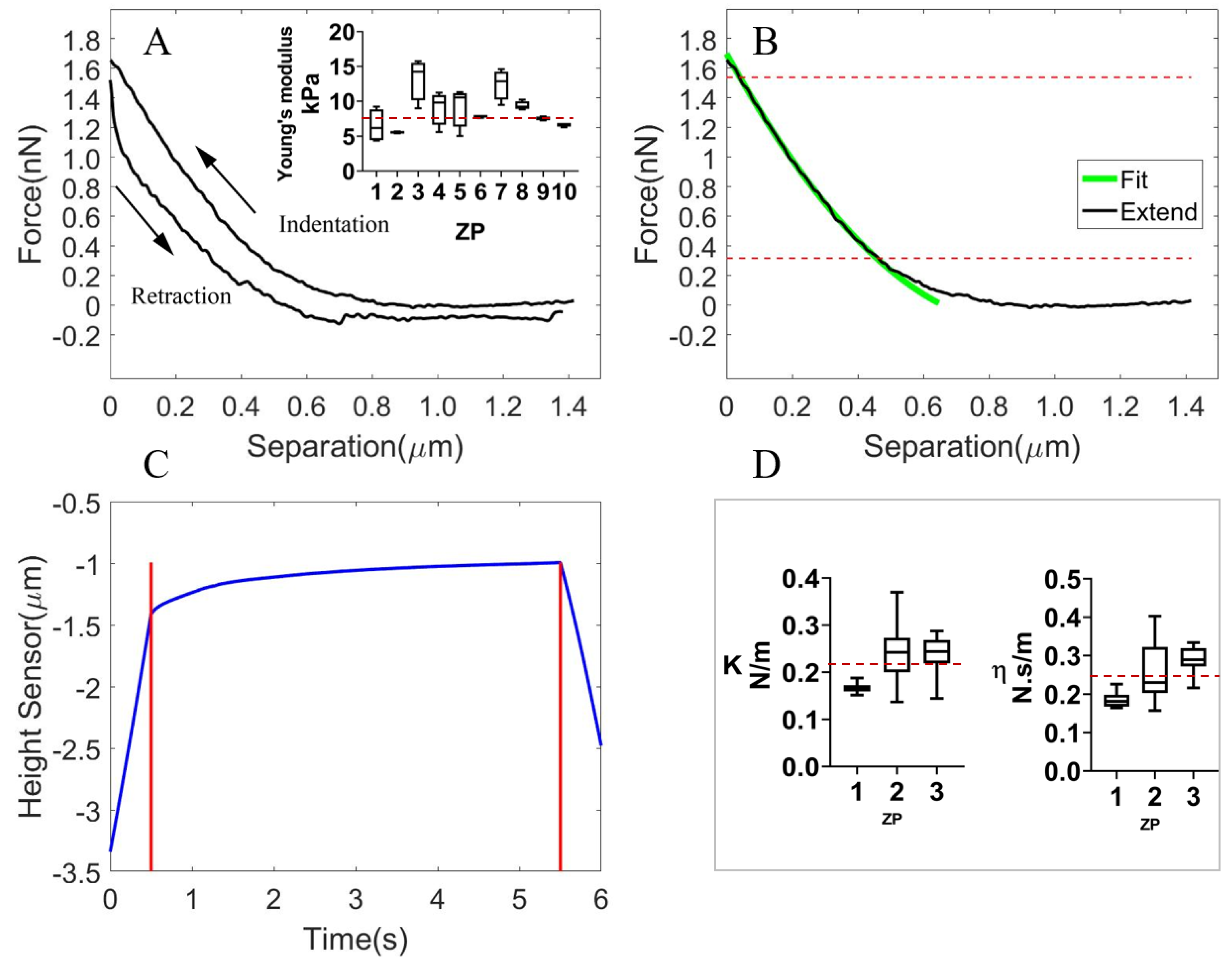
3.1.2. The Viscoelastic Properties of the Isolated ZP
3.1.3. The Viscoelastic Properties of the Whole Cell
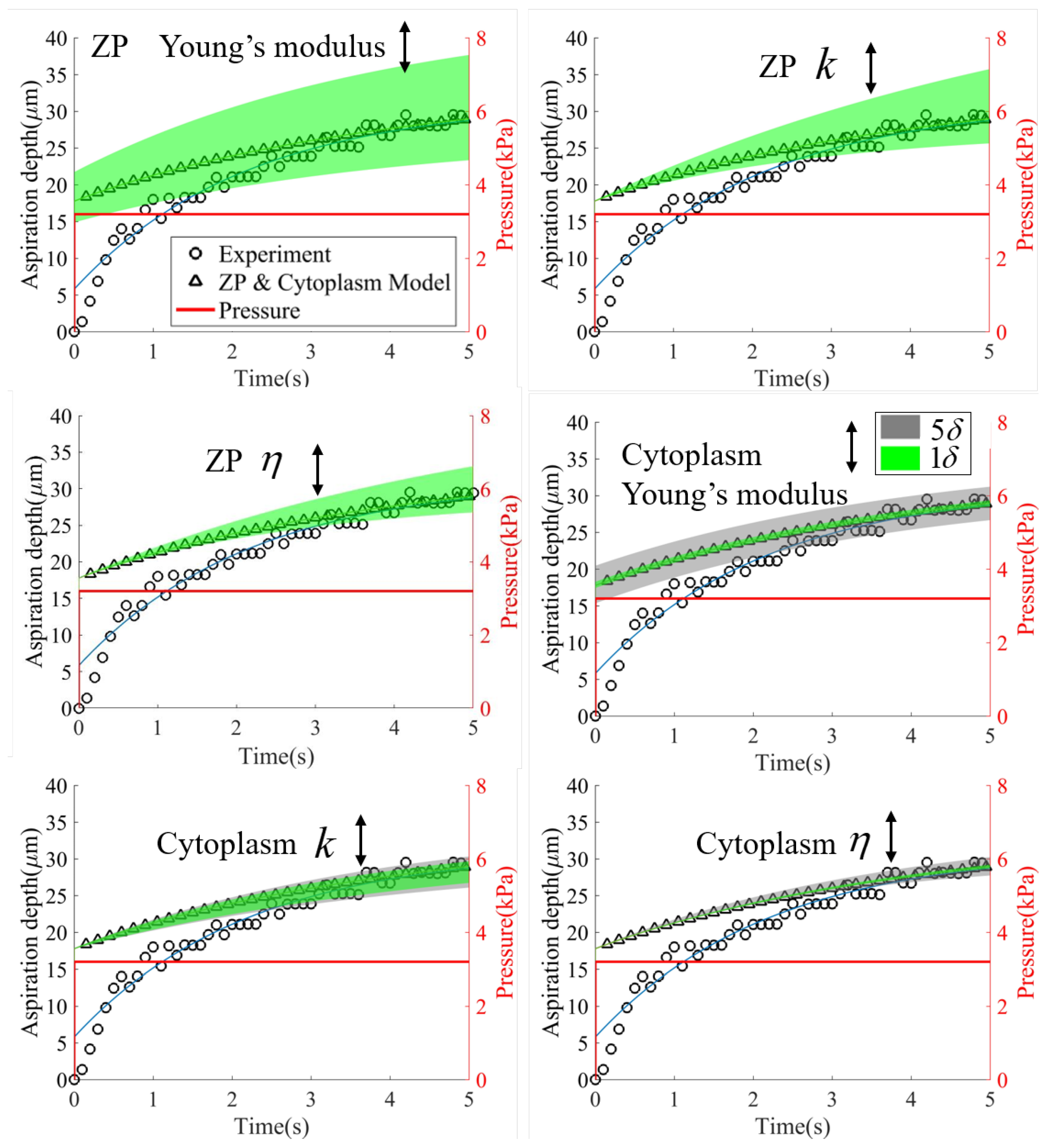
3.2. Validation of the Cell Model in the Micropipette Aspiration Experiment
3.3. Validation of the Cell Model in the Microinjection Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yanez, L.Z.; Han, J.; Behr, B.B.; Pera, R.A.R.; Camarillo, D.B. Human oocyte developmental potential is predicted by mechanical properties within hours after fertilization. Nat. Commun. 2016, 7, 10809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andolfi, L.; Masiero, E.; Giolo, E.; Martinelli, M.; Luppi, S.; Zilio, S.D.; Delfino, I.; Bortul, R.; Zweyer, M.; Ricci, G.; et al. Investigating the mechanical properties of zona pellucida of whole human oocytes by atomic force spectroscopy. Integr. Biol. 2016, 8, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Litscher, E.S.; Wassarman, P.M. Zona pellucida proteins, fibrils, and matrix. Annu. Rev. Biochem. 2020, 89, 695–715. [Google Scholar] [CrossRef]
- Zarnescu, L.; Han, J.; Behr, B.; Pera, R.R.; Camarillo, D. Mechanical biomarkers of oocyte maturation. Fertil. Steril. 2014, 102, e86. [Google Scholar] [CrossRef]
- Wassarman, P.M. Zona pellucida glycoproteins. J. Biol. Chem. 2008, 283, 24285–24289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murayama, Y.; Yoshida, M.; Mizuno, J.; Nakamura, H.; Inoue, S.; Watanabe, Y.; Akaishi, K.; Inui, H.; Constantinou, C.E.; Omata, S. Elasticity measurement of zona pellucida using a micro tactile sensor to evaluate embryo quality. J. Mamm. Ova Res. 2008, 25, 8–16. [Google Scholar] [CrossRef]
- Khalilian, M.; Navidbakhsh, M.; Valojerdi, M.R.; Chizari, M.; Yazdi, P.E. Estimating Young’s modulus of zona pellucida by micropipette aspiration in combination with theoretical models of ovum. J. R. Soc. Interface 2010, 7, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Khalilian, M.; Navidbakhsh, M.; Rezazadeh Valojerdi, M.; Chizari, M.; Eftekhari Yazdi, P. Alteration in the mechanical properties of human ovum zona pellucida following fertilization: Experimental and analytical studies. Exp. Mech. 2011, 51, 175–182. [Google Scholar] [CrossRef]
- Sun, Y.; Wan, K.T.; Roberts, K.P.; Bischof, J.C.; Nelson, B.J. Mechanical property characterization of mouse zona pellucida. IEEE Trans. Nanobiosci. 2003, 2, 279–286. [Google Scholar] [CrossRef]
- Papi, M.; Maiorana, A.; Douet, C.; Maulucci, G.; Parasassi, T.; Brunelli, R.; Goudet, G.; De Spirito, M. Viscous forces are predominant in the zona pellucida mechanical resistance. Appl. Phys. Lett. 2013, 102, 043703. [Google Scholar] [CrossRef]
- Boccaccio, A.; Lamberti, L.; Papi, M.; De Spirito, M.; Douet, C.; Goudet, G.; Pappalettere, C. A hybrid characterization framework to determine the visco-hyperelastic properties of a porcine zona pellucida. Interface Focus 2014, 4, 20130066. [Google Scholar] [CrossRef] [PubMed]
- Valberg, P.A.; Feldman, H.A. Magnetic particle motions within living cells. Measurement of cytoplasmic viscosity and motile activity. Biophys. J. 1987, 52, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.; Yeung, A. Apparent viscosity and cortical tension of blood granulocytes determined by micropipet aspiration. Biophys. J. 1989, 56, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Ragsdale, G.K.; Phelps, J.; Luby-Phelps, K. Viscoelastic response of fibroblasts to tension transmitted through adherens junctions. Biophys. J. 1997, 73, 2798–2808. [Google Scholar] [CrossRef] [Green Version]
- Bausch, A.R.; Ziemann, F.; Boulbitch, A.A.; Jacobson, K.; Sackmann, E. Local measurements of viscoelastic parameters of adherent cell surfaces by magnetic bead microrheometry. Biophys. J. 1998, 75, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Bausch, A.R.; Möller, W.; Sackmann, E. Measurement of local viscoelasticity and forces in living cells by magnetic tweezers. Biophys. J. 1999, 76, 573–579. [Google Scholar] [CrossRef] [Green Version]
- Daniels, B.R.; Masi, B.C.; Wirtz, D. Probing single-cell micromechanics in vivo: The microrheology of C. elegans developing embryos. Biophys. J. 2006, 90, 4712–4719. [Google Scholar] [CrossRef] [Green Version]
- Margraves, C.; Kihm, K.; Yoon, S.Y.; Choi, C.K.; Lee, S.H.; Liggett, J.; Baek, S.J. Simultaneous measurements of cytoplasmic viscosity and intracellular vesicle sizes for live human brain cancer cells. Biotechnol. Bioeng. 2011, 108, 2504–2508. [Google Scholar] [CrossRef]
- Moeendarbary, E.; Valon, L.; Fritzsche, M.; Harris, A.R.; Moulding, D.A.; Thrasher, A.J.; Stride, E.; Mahadevan, L.; Charras, G.T. The cytoplasm of living cells behaves as a poroelastic material. Nat. Mater. 2013, 12, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Brugués, J.; Maugis, B.; Casademunt, J.; Nassoy, P.; Amblard, F.; Sens, P. Dynamical organization of the cytoskeletal cortex probed by micropipette aspiration. Proc. Natl. Acad. Sci. USA 2010, 107, 15415–15420. [Google Scholar] [CrossRef] [Green Version]
- Guevorkian, K.; Maître, J.L. Micropipette aspiration: A unique tool for exploring cell and tissue mechanics in vivo. Methods Cell Biol. 2017, 139, 187–201. [Google Scholar] [PubMed]
- Pu, H.; Liu, N.; Yu, J.; Yang, Y.; Sun, Y.; Peng, Y.; Xie, S.; Luo, J.; Dong, L.; Chen, H.; et al. Micropipette aspiration of single cells for both mechanical and electrical characterization. IEEE Trans. Biomed. Eng. 2019, 66, 3185–3191. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ho, C.; Tsatskis, Y.; Law, J.; Zhang, Z.; Zhu, M.; Dai, C.; Wang, F.; Tan, M.; Hopyan, S.; et al. Intracellular manipulation and measurement with multipole magnetic tweezers. Sci. Robot. 2019, 4, eaav6180. [Google Scholar] [CrossRef] [PubMed]
- Guilluy, C.; Osborne, L.D.; Van Landeghem, L.; Sharek, L.; Superfine, R.; Garcia-Mata, R.; Burridge, K. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nat. Cell Biol. 2014, 16, 376–381. [Google Scholar] [CrossRef]
- Sakuma, S.; Nakahara, K.; Arai, F. Continuous mechanical indexing of single-cell spheroids using a robot-integrated microfluidic chip. IEEE Robot. Autom. Lett. 2019, 4, 2973–2980. [Google Scholar] [CrossRef]
- Hu, S.; Liu, G.; Chen, W.; Li, X.; Lu, W.; Lam, R.H.; Fu, J. Multiparametric biomechanical and biochemical phenotypic profiling of single cancer cells using an elasticity microcytometer. Small 2016, 12, 2300–2311. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Meng, X.; Zhang, H.; Zhao, K.; Hu, Y.; Xie, H. Probing Multidimensional Mechanical Phenotyping of Intracellular Structures by Viscoelastic Spectroscopy. ACS Appl. Mater. Interfaces 2020, 12, 1913–1923. [Google Scholar] [CrossRef]
- Liu, H.; Wen, J.; Xiao, Y.; Liu, J.; Hopyan, S.; Radisic, M.; Simmons, C.A.; Sun, Y. In situ mechanical characterization of the cell nucleus by atomic force microscopy. ACS Nano 2014, 8, 3821–3828. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Zhu, M.; Cao, C.; Xu, Z.; Tsatskis, Y.; Lau, K.; Kuok, C.; Filleter, T.; McNeill, H.; et al. Mechanical stability of the cell nucleus–roles played by the cytoskeleton in nuclear deformation and strain recovery. J. Cell. Sci. 2018, 131, jcs209627. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, X.; Zhao, Q.; Zhao, X.; Sun, M. Robotic batch somatic cell nuclear transfer based on microfluidic groove. IEEE Trans. Autom. Sci. Eng. 2020, 17, 2097–2106. [Google Scholar] [CrossRef]
- Gong, H.; Li, L.; Qiu, J.; Yao, Y.; Liu, Y.; Cui, M.; Zhao, Q.; Zhao, X.; Sun, M. Automatic cell rotation based on real-time detection and tracking. IEEE Robot. Autom. Lett. 2021, 6, 7909–7916. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, M.; Sun, Y.; Feng, Z.; Bai, Y.; Sun, M.; Zhao, Q.; Zhao, X. Oocyte orientation selection method based on the minimum strain position in the penetration process. J. Appl. Phys. 2019, 125, 154701. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Y.; Zhao, Q.; Sun, M.; Zhao, X. Modeling and measuring intracellular displacement during cell penetration. J. Appl. Phys. 2020, 127, 044701. [Google Scholar] [CrossRef]
- Grasland-Mongrain, P.; Zorgani, A.; Nakagawa, S.; Bernard, S.; Paim, L.G.; Fitzharris, G.; Catheline, S.; Cloutier, G. Ultrafast imaging of cell elasticity with optical microelastography. Proc. Natl. Acad. Sci. USA 2018, 115, 861–866. [Google Scholar] [CrossRef] [Green Version]
- Papi, M.; Brunelli, R.; Sylla, L.; Parasassi, T.; Monaci, M.; Maulucci, G.; Missori, M.; Arcovito, G.; Ursini, F.; De Spirito, M. Mechanical properties of zona pellucida hardening. Eur. Biophys. J. 2010, 39, 987–992. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, M.; Huang, J.; Sun, M.; Zhao, X.; Zhao, Q. Robotic micropipette aspiration for multiple cells. Micromachines 2019, 10, 348. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.H.; Abeydeera, L.R.; Prather, R.S.; Day, B.N. Morphologic comparison of ovulated and in vitro–matured porcine oocytes, with particular reference to polyspermy after in vitro fertilization. Mol. Reprod. Dev. 1998, 49, 308–316. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Simulation | |
|---|---|---|
| ZP | Radius | 75 m |
| Young’s Modulus | 17.63 ± 5.10 kPa | |
| Poisson’s ratio | 0.5 | |
| k | 0.259 ± 0.093 N/m | |
| 0.290 ± 0.105 N·s/m | ||
| Isolated ZP | Thickness | 20 m |
| Young’s Modulus | 8.77 ± 2.38 kPa | |
| Poisson’s ratio | 0.5 | |
| k | 0.216 ± 0.034 N/m | |
| 0.245 ± 0.044 N·s/m | ||
| Cytoplasm | Radius | 55 m |
| Young’s Modulus | 3.16 ± 0.35 kPa | |
| Poisson’s ratio | 0.5 | |
| k | 0.052 ± 0.005 N/m | |
| 0.082 ± 0.009 N·s/m | ||
| Pipette | Inner Diameter | 90 m |
| External Diameter | 108 m | |
| Young’s Modulus | 5.5 × kPa | |
| Contact mode | ZP and Cytoplasm | Bonded |
| Pipette and ZP | Frictional | |
| Friction Coefficient 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Chen, Y.; Zhang, S.; Cheng, D.; Liu, Y.; Zhao, Q.; Sun, M.; Cui, M.; Zhao, X. Mechanical Characterization and Modelling of Subcellular Components of Oocytes. Micromachines 2022, 13, 1087. https://doi.org/10.3390/mi13071087
Du Y, Chen Y, Zhang S, Cheng D, Liu Y, Zhao Q, Sun M, Cui M, Zhao X. Mechanical Characterization and Modelling of Subcellular Components of Oocytes. Micromachines. 2022; 13(7):1087. https://doi.org/10.3390/mi13071087
Chicago/Turabian StyleDu, Yue, Yizhe Chen, Shuai Zhang, Dai Cheng, Yaowei Liu, Qili Zhao, Mingzhu Sun, Maosheng Cui, and Xin Zhao. 2022. "Mechanical Characterization and Modelling of Subcellular Components of Oocytes" Micromachines 13, no. 7: 1087. https://doi.org/10.3390/mi13071087