Microfluidic Transportation Control of Larval Zebrafish through Optomotor Regulations under a Pressure-Driven Flow


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Maintenance and Larvae Preparation
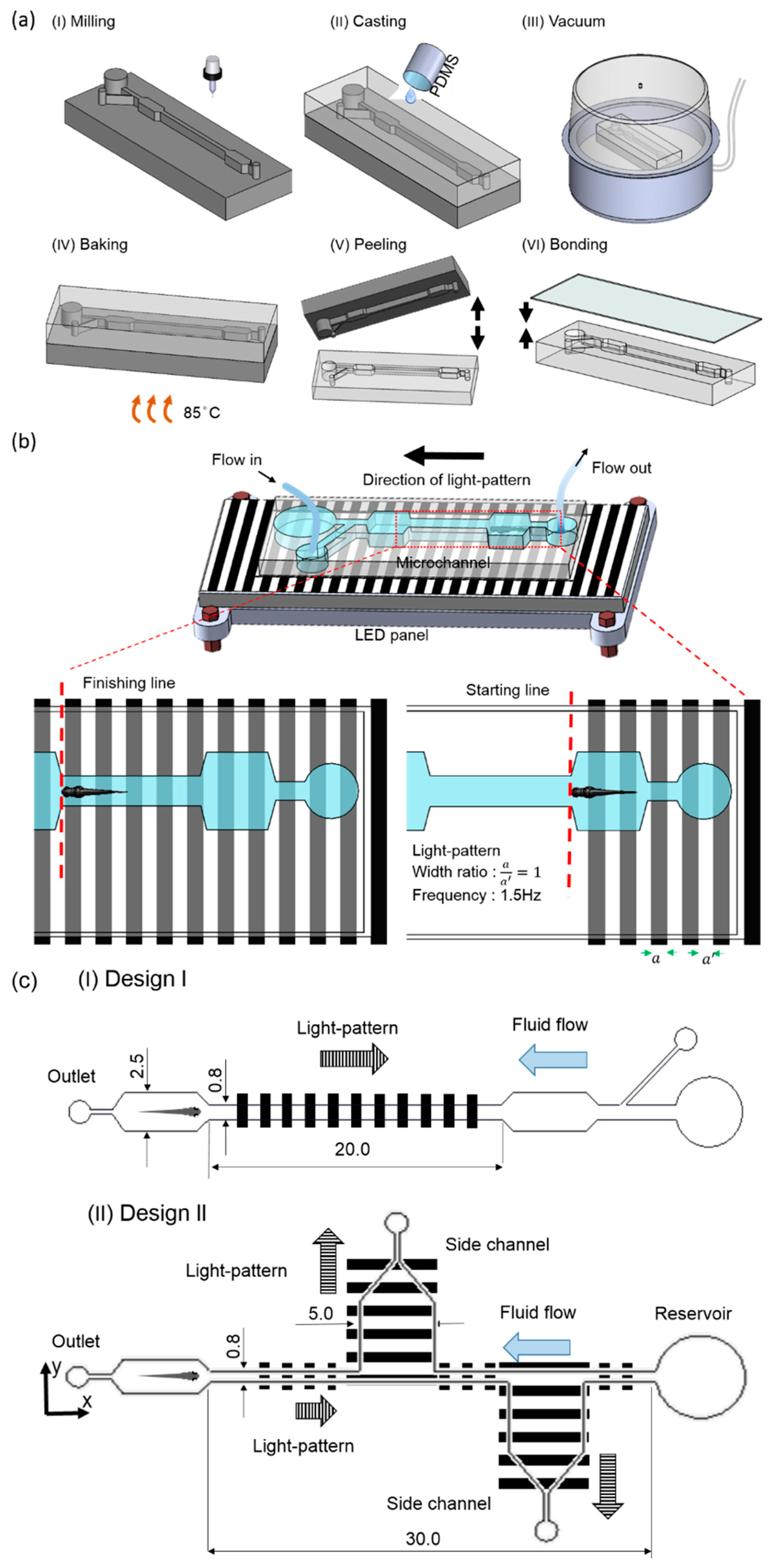
2.2. Design Details and Experimental Platform
2.3. Data Analysis
3. Results
3.1. Behavioral Responses Corresponding to Optical and Hydrodynamic Cues
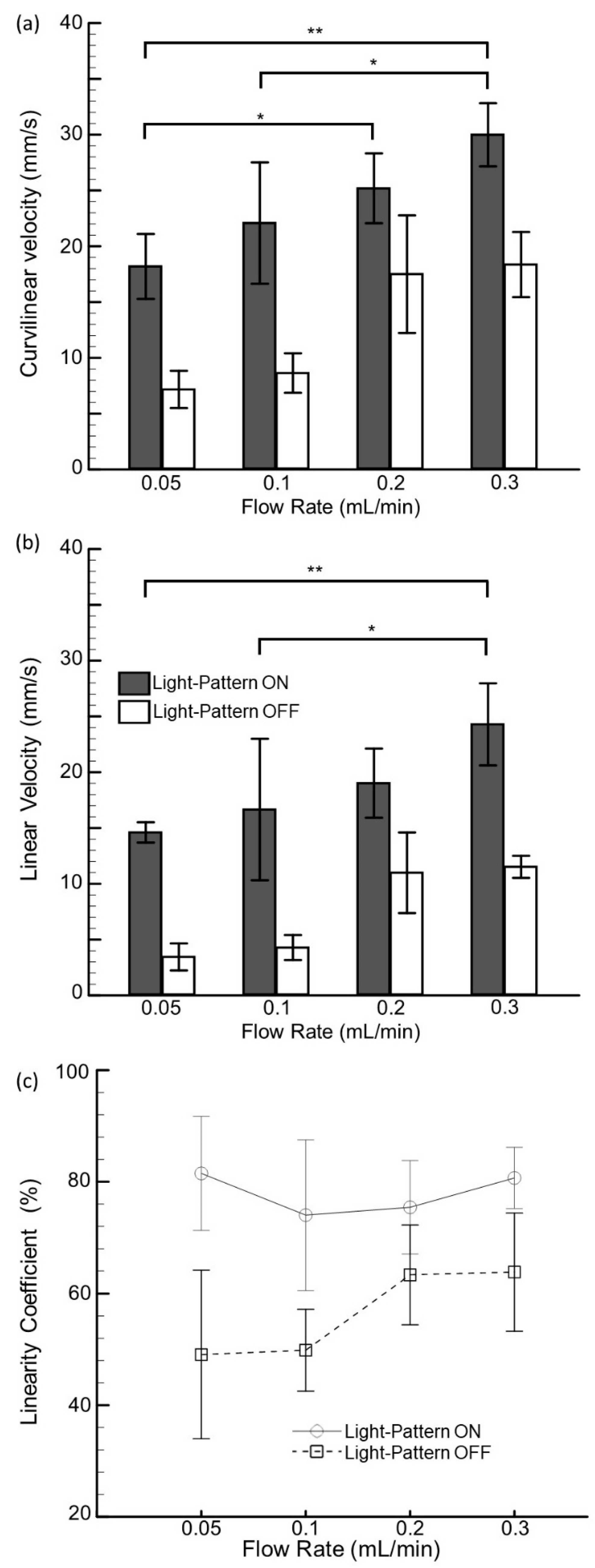
3.2. Kinetic Parameters of Swimming Larvae
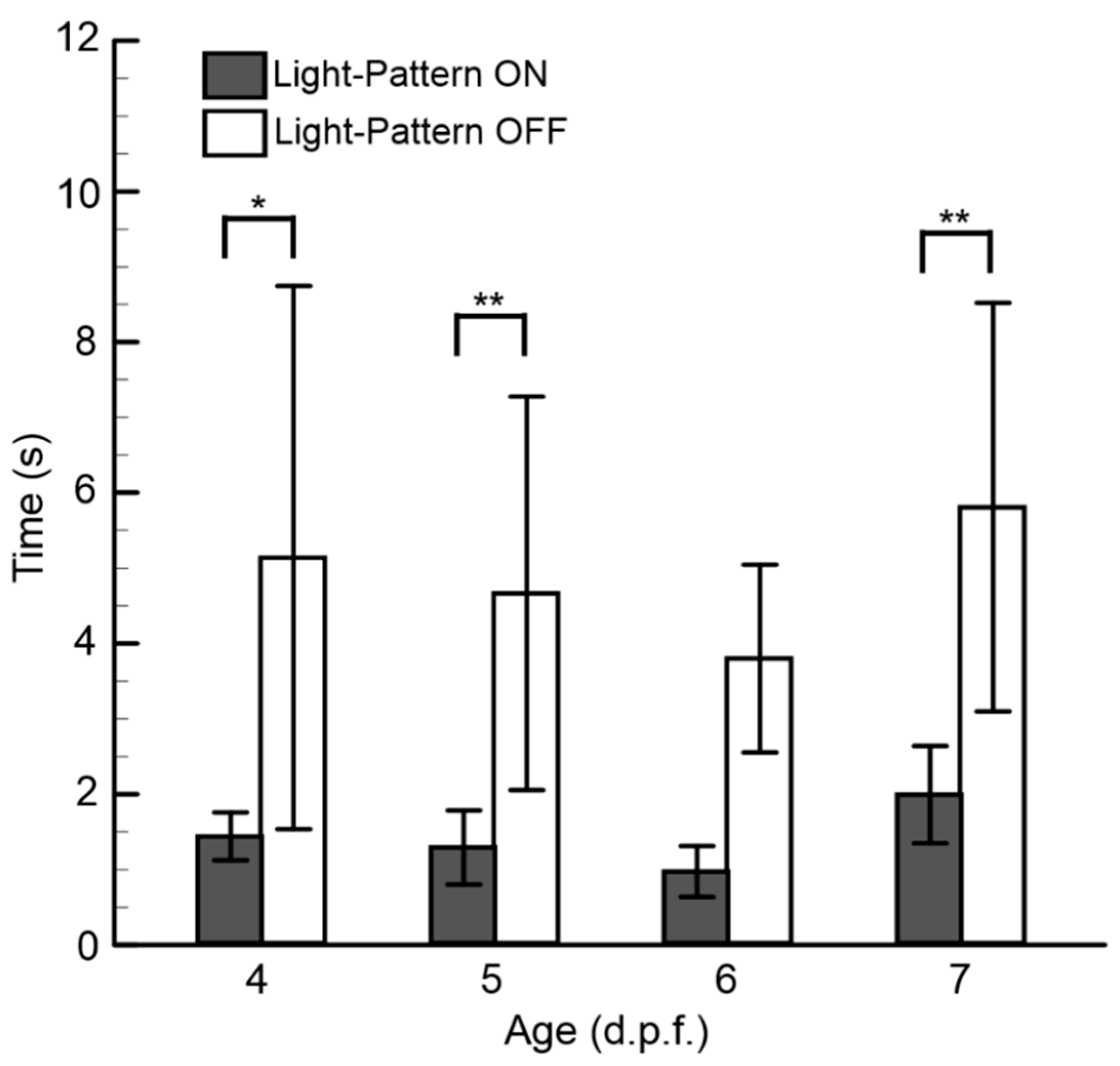
3.3. Behavioral Responses Corresponding to Larval Age
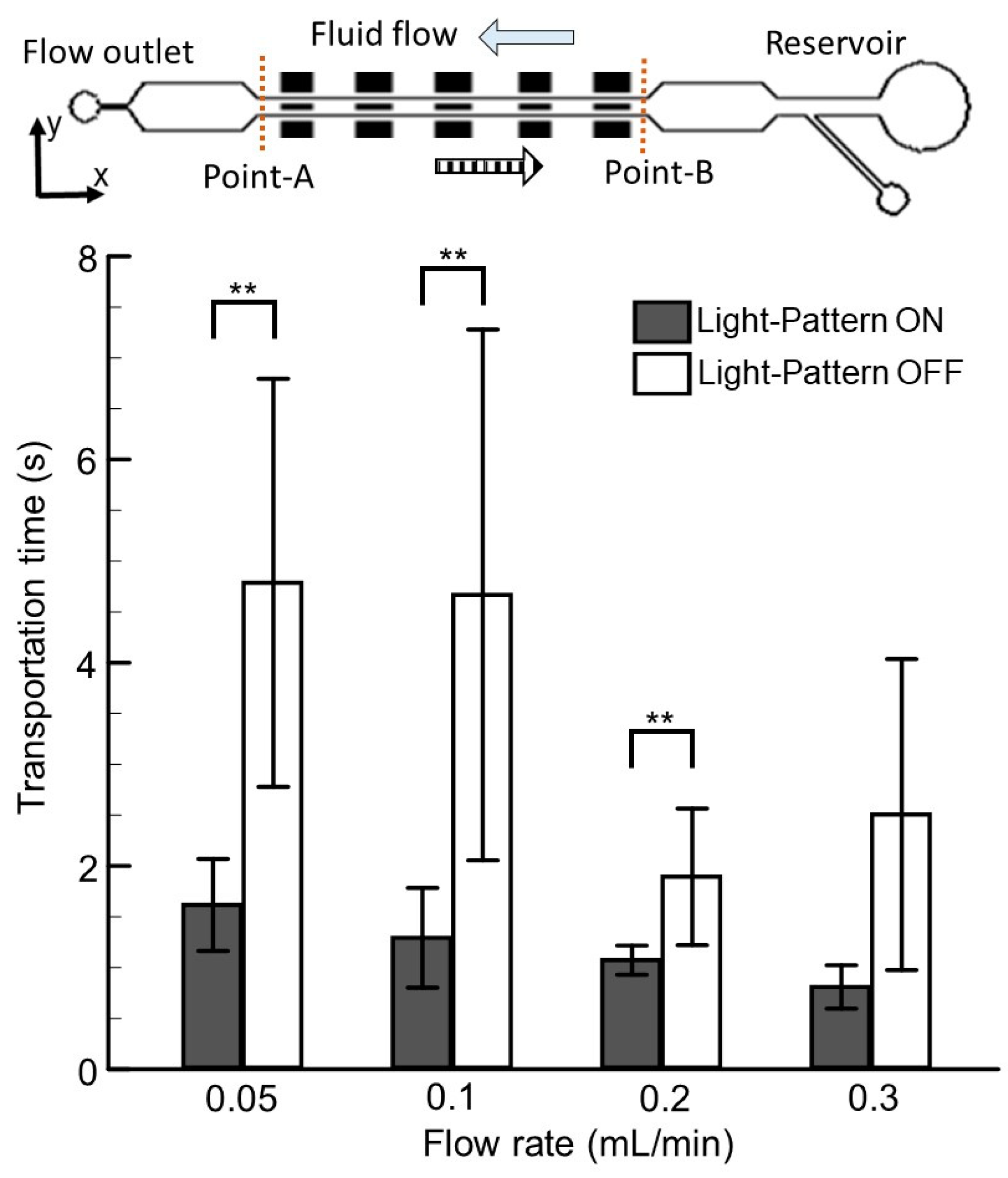
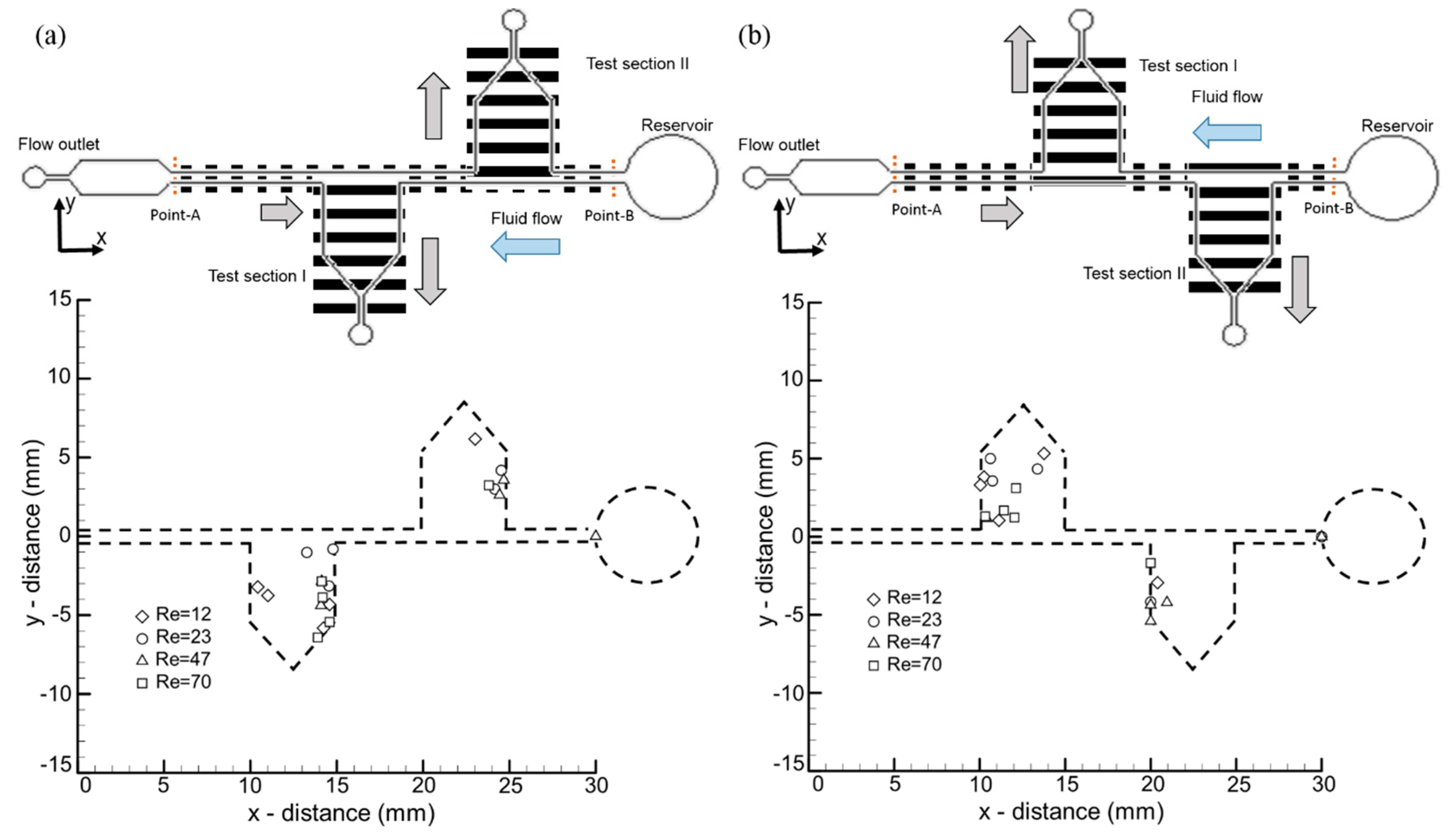
3.4. The Effects of Visual and Hydromechanical Stimuli on Larvae Transportation in a Microfluidic Network
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Tierney, K.B. Behavioural assessments of neurotoxic effects and neurodegeneration in zebrafish. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2011, 1812, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakkers, J. Zebrafish as a model to study cardiac development and human cardiac disease. Cardiovasc. Res. 2011, 91, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corti, P.; Young, S.; Chen, C.-Y.; Patrick, M.J.; Rochon, E.R.; Pekkan, K.; Roman, B.L. Interaction between alk1 and blood flow in the development of arteriovenous malformations. Development 2011, 138, 1573–1582. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Patrick, M.J.; Corti, P.; Kowalski, W.; Roman, B.L.; Pekkan, K. Analysis of early embryonic great-vessel microcirculation in zebrafish using high-speed confocal uPIV. Biorheology 2011, 48, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Cheng, C.M. Microfluidics expands the zebrafish potentials in pharmaceutically relevant screening. Adv. Healthc. Mater. 2014, 3, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Ekker, S.C. Zebrafish as a genomics research model. Curr. Pharm. Biotechnol. 2004, 5, 409–413. [Google Scholar] [CrossRef]
- Wang, K.; Ma, J.; He, M.; Gao, G.; Xu, H.; Sang, J.; Wang, Y.; Zhao, B.; Cui, D. Toxicity assessments of near-infrared upconversion luminescent LaF3: Yb, Er in early development of zebrafish embryos. Theranostics 2013, 3, 258. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Zebrafish: A complete animal model to enumerate the nanoparticle toxicity. J. Nanobiotechnol. 2016, 14, 65. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Gao, C.; Wang, P.; Zhang, G.-J.; Chen, Z. Fish-on-a-chip: Microfluidics for zebrafish research. Lab Chip 2016, 16, 1106–1125. [Google Scholar] [CrossRef]
- Mani, K.; Chien, T.-C.C.; Panigrahi, B.; Chen, C.-Y. Manipulation of zebrafish’s orientation using artificial cilia in a microchannel with actively adaptive wall design. Sci. Rep. 2016, 6, 36385. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, D.; van Noort, D.; Iliescu, C.; Zheng, B.X.; Poon, K.L.; Korzh, S.; Korzh, V.; Yu, H. Fish and chips: A microfluidic perfusion platform for monitoring zebrafish development. Lab Chip 2012, 12, 892–900. [Google Scholar] [CrossRef]
- Chang, T.Y.; Pardo-Martin, C.; Allalou, A.; Wahlby, C.; Yanik, M.F. Fully automated cellular-resolution vertebrate screening platform with parallel animal processing. Lab Chip 2012, 12, 711–716. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Chien, T.-C.C.; Mani, K.; Tsai, H.-Y. Axial orientation control of zebrafish larvae using artificial cilia. Microfluid. Nanofluid. 2016, 20, 12. [Google Scholar] [CrossRef]
- Olive, R.; Wolf, S.; Dubreuil, A.; Bormuth, V.; Debrégeas, G.; Candelier, R. Rheotaxis of larval zebrafish: Behavioral study of a multi-sensory process. Front. Sys. Neurosci. 2016, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Oteiza, P.; Odstrcil, I.; Lauder, G.; Portugues, R.; Engert, F. A novel mechanism for mechanosensory-based rheotaxis in larval zebrafish. Nature 2017, 547, 445. [Google Scholar] [CrossRef] [Green Version]
- Orger, M.B.; de Polavieja, G.G. Zebrafish behavior: Opportunities and challenges. Annu. Rev. Neurosci. 2017, 40, 125–147. [Google Scholar] [CrossRef] [Green Version]
- Bak-Coleman, J.; Smith, D.; Coombs, S. Going with, then against the flow: Evidence against the optomotor hypothesis of fish rheotaxis. Anim. Behav. 2015, 107, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, J.C.; Baker, C.F.; Carton, A.G. The lateral line can mediate rheotaxis in fish. Nature 1997, 389, 960. [Google Scholar] [CrossRef]
- Suli, A.; Watson, G.M.; Rubel, E.W.; Raible, D.W. Rheotaxis in larval zebrafish is mediated by lateral line mechanosensory hair cells. PLoS ONE 2012, 7, e29727. [Google Scholar] [CrossRef] [Green Version]
- Van Trump, W.J.; McHenry, M.J. The morphology and mechanical sensitivity of lateral line receptors in zebrafish larvae (danio rerio). J. Exp. Biol. 2008, 211, 2105–2115. [Google Scholar] [CrossRef] [Green Version]
- Higham, T.E.; Stewart, W.J.; Wainwright, P.C. Turbulence, temperature, and turbidity: The ecomechanics of predator—Prey interactions in fishes. Integr. Comp. Biol. 2015, 55, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Orger, M.B.; Smear, M.C.; Anstis, S.M.; Baier, H. Perception of fourier and non-fourier motion by larval zebrafish. Nat. Neurosci. 2000, 3, 1128. [Google Scholar] [CrossRef]
- Bilotta, J.; Saszik, S. The zebrafish as a model visual system. Int. J. Dev. Neurosci. 2001, 19, 621–629. [Google Scholar] [CrossRef]
- Orger, M.B.; Kampff, A.R.; Severi, K.E.; Bollmann, J.H.; Engert, F. Control of visually guided behavior by distinct populations of spinal projection neurons. Nat. Neurosci. 2008, 11, 327. [Google Scholar] [CrossRef]
- Mani, K.; Hsieh, Y.-C.; Panigrahi, B.; Chen, C.-Y. A noninvasive light driven technique integrated microfluidics for zebrafish larvae transportation. Biomicrofluidics 2018, 12, 021101. [Google Scholar] [CrossRef] [Green Version]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Brachydanio Rerio); University of Oregon Press: Corvallis, OR, USA, 1993. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to Imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Neuhauss, S.C. Behavioral genetic approaches to visual system development and function in zebrafish. J. Neurobiol. 2003, 54, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Peimani, A.R.; Zoidl, G.; Rezai, P. A microfluidic device for quantitative investigation of zebrafish larvae’s rheotaxis. Biomed. Microdevices 2017, 19, 99. [Google Scholar] [CrossRef]
- Symvoulidis, P.; Lauri, A.; Stefanoiu, A.; Cappetta, M.; Schneider, S.; Jia, H.; Stelzl, A.; Koch, M.; Perez, C.C.; Myklatun, A. Neubtracker—imaging neurobehavioral dynamics in freely behaving fish. Nat. Methods 2017, 14, 1079. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, J.; Marques, J.C.; Grama, A.; Hildebrand, D.G.; Gu, W.; Li, J.M.; Robson, D.N. Pan-neuronal calcium imaging with cellular resolution in freely swimming zebrafish. Nat. Methods 2017, 14, 1107. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panigrahi, B.; Chen, C.-Y. Microfluidic Transportation Control of Larval Zebrafish through Optomotor Regulations under a Pressure-Driven Flow. Micromachines 2019, 10, 880. https://doi.org/10.3390/mi10120880
Panigrahi B, Chen C-Y. Microfluidic Transportation Control of Larval Zebrafish through Optomotor Regulations under a Pressure-Driven Flow. Micromachines. 2019; 10(12):880. https://doi.org/10.3390/mi10120880
Chicago/Turabian StylePanigrahi, Bivas, and Chia-Yuan Chen. 2019. "Microfluidic Transportation Control of Larval Zebrafish through Optomotor Regulations under a Pressure-Driven Flow" Micromachines 10, no. 12: 880. https://doi.org/10.3390/mi10120880