
Sensitivity of L-Band SAR Backscatter to Aboveground Biomass of Global Forests


Abstract
:
1. Introduction
- What is the overall sensitivity of L-band backscatter to AGB over global forests?
- How many forest specific algorithms are required for global estimation of AGB?
- What is the minimum number of radar observations required to estimate AGB annually?
2. Materials and Methods
2.1. Remote Sensing Data
2.1.1. ALOS PALSAR
2.1.2. ICESat GLAS LiDAR
2.1.3. Landcover Maps
2.2. AGB Estimation from GLAS
2.3. Methodology
2.3.1. Radar Biomass Models
2.3.2. Radar Sensitivity
2.3.3. Radar Detection of Biomass
3. Results
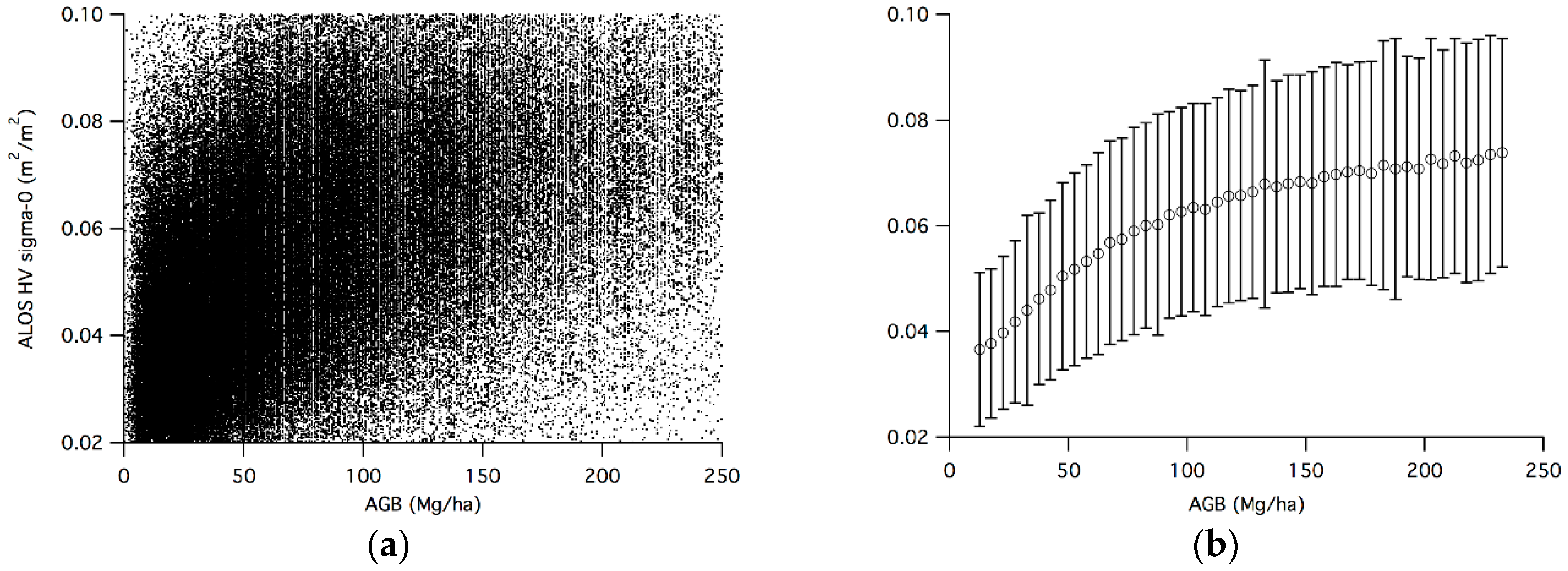
3.1. GLAS-Based AGB and ALOS Backscatter
3.2. Sensitivity Analysis
3.3. Required Number of Observations
4. Discussion
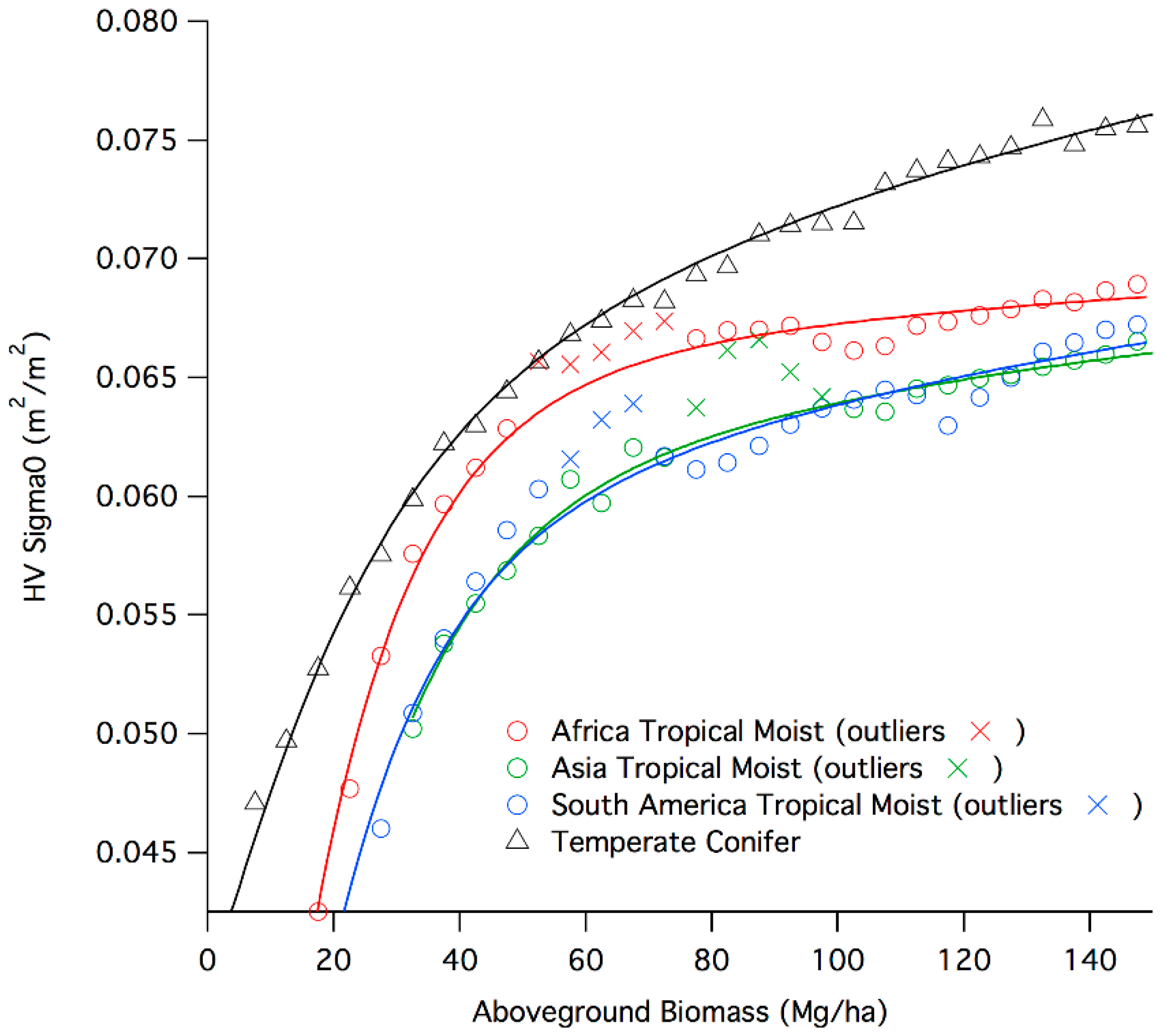
4.1. Radar Backscatter Model
4.2. Backscatter Sensitivity to Biomass
4.3. Anomaly in Radar Backscatter Sensitivity to AGB
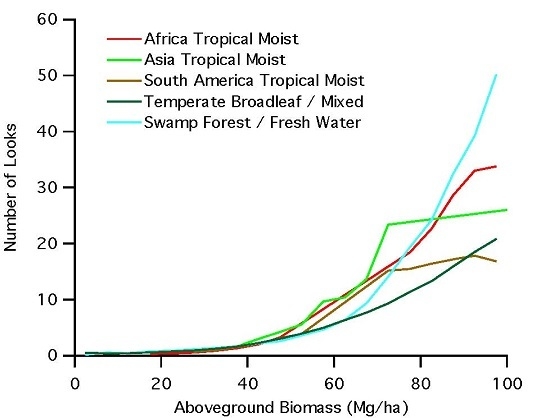
- Assuming no errors in the GLAS LiDAR measurements over swamp forests, the results suggest a bi-modal distribution in terms of AGB. Since we can generally assume that GLAS is a systematic measurement of the surface, then the histogram of point distribution within each AGB bin can be interpreted as the frequency of occurrence of this particular AGB-class within the given forest type. The bi-modal distribution suggests that there may be two types of swamp forests with low and high average biomass density, and with very few areas transitioning between the two forest types. Although each type has a distribution around the mean value (~40 Mg·ha−1 for low biomass swamps and ~175 Mg·ha−1 for high biomass swamps), the two swamp forests are distinct in their biomass values.
- The discontinuity in radar backscatter between these two modes also points to some different physical and/or environmental conditions between the two forest types giving rise to significantly different scattering responses. The responses may be associated with two different inundation cycles or seasonal variations in water level with the low biomass swamp forests coinciding with high inundation state and high biomass swamps under a lower inundation state, creating a bi-modal behavior in the radar backscatter. In addition, here, we are only considering the HV backscatter with the low sensitivity to inundation state. There is a high possibility that HV backscatter is capturing a strong volume-surface scattering in low biomass swamps and then transitioning to the regular HV biomass relationship to high biomass swamps.
- There is also the possibility of GLAS measurements are erroneous in swamp forests with high-level inundation. Water has a strong absorption of the near-infrared wavelength used by GLAS. If enough of the surface under the forest is inundated with water, the GLAS LiDAR observation may experience very weak return from the surface and erroneously assume sub-canopy returns to be ground returns, and hence underestimating the canopy height. This would have an effect of shifting points in the affected areas into bins to the left (smaller AGB). The result of such shift would be an increase in the average sigma-0 of the lower bins (HV backscatter is not affected by the same mechanism), and a decrease in the number of points in the bins that the GLAS shots were shifted away from. However, this would also suggest that the same effect is not observed or is as pronounced in high biomass swamp forests. Otherwise, everything would shift to the left together there would not be a discontinuity in the AGB values that suggest that this may not be the right explanation of the discontinuity in the AGB values. To further explore the cause of this behavior in the radar backscatter, studies that include ground measurements of forest biomass and multi-temporal radar observations at different inundation state may be required.
4.4. GLAS Lidar Derived AGB
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ALOS | Advanced Land Observing Satellite |
| PALSAR | Phased Array L-band Synthetic Aperture Radar |
| GLAS | Geoscience Laser Altimeter System |
| AGB | Aboveground Biomass |
| SAR | Synthetic Aperture Radar |
| LiDAR | Light Detection and Ranging |
| WWF | World Wildlife Fund |
| DRL | Discrete Return LiDAR |
| NASA | National Aeronautics and Space Administration |
| ISRO | Indian Space Research Organization |
| ESA | European Space Agency |
| NISAR | NASA ISRO SAR |
References
- Houghton, R.A.; Hall, F.; Goetz, S.J. Importance of biomass in the global carbon cycle. J. Geophys. Res. 2009, 114, G00E03. [Google Scholar] [CrossRef]
- Le Quéré, C.; Moriarty, R.; Andrew, R.M.; Canadell, J.G.; Sitch, S.; Korsbakken, J.I.; Friedlingstein, P.; Peters, G.P.; Andres, R.J.; Boden, T.A.; et al. Global carbon budget 2015. Earth Syst. Sci. Data 2015, 7, 349–396. [Google Scholar] [CrossRef] [Green Version]
- Frolking, S.; Palace, M.W.; Clark, D.B.; Chambers, J.Q.; Shugart, H.H.; Hurtt, G.C. Forest disturbance and recovery: A general review in the context of spaceborne remote sensing of impacts on aboveground biomass and canopy structure. J. Geophys. Res. 2009, 114, G00E02. [Google Scholar] [CrossRef]
- Harris, N.L.; Brown, S.; Hagen, S.C.; Saatchi, S.S.; Petrova, S. Baseline map of carbon emissions from deforestation in tropical regions. Science 2012, 336, 1573–1576. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and estimating tropical forest carbon stocks: Making REDD a reality. Environ. Res. Lett. 2007, 2, 045023. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.A.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Forest Resources Assessment 2010—Main Report; FAO Forestry: Rome, Italy, 2010. [Google Scholar]
- Keith, H.; Mackey, B.G.; Lindenmayer, D.B. Re-evaluation of forest biomass carbon stocks and lessons from the world’s most carbon-dense forests. Proc. Natl. Acad. Sci. USA 2009, 106, 11635–11640. [Google Scholar] [CrossRef] [PubMed]
- Schimel, D.; Stephens, B.B.; Fisher, J.B. Effect of increasing CO2 on the terrestrial carbon cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Saatchi, S.; Mascaro, J.; Xu, L.; Keller, M.; Yang, Y.; Duffy, P.; Espírito Santo, F.; Baccini, A.; Chambers, J.; Schimel, D. Seeing the forest beyond the trees. Glob. Ecol. Biogeogr. 2015, 24, 606–610. [Google Scholar] [CrossRef]
- Woodall, C.W.; Heath, L.S.; Domke, D.M.; Nichols, M.C. Methods and Equations for Estimating Aboveground Volume, Biomass, and Carbon for Trees in the US Forest Inventory, 2010; U.S. Forest Service: Washington, DC, USA, 2011. [Google Scholar]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Thurner, M.; Beer, C.; Santoro, M.; Carvalhais, N.; Wutzler, T.; Schepaschenko, D.; Shvidenko, A.; Kompter, E.; Ahrens, B.; Levick, S.R.; et al. Carbon stock and density of northern boreal and temperate forests. Glob. Ecol. Biogeogr. 2014, 23, 297–310. [Google Scholar] [CrossRef]
- Le Toan, T.; Quegan, S.; Davidson, M.W.J.; Balzter, H.; Paillou, P.; Papathanassiou, K.; Plummer, S.; Rocca, F.; Saatchi, S.; Shugart, H.; et al. The BIOMASS mission: Mapping global forest biomass to better understand the terrestrial carbon cycle. Remote Sens. Environ. 2011, 115, 2850–2860. [Google Scholar] [CrossRef]
- Mitchard, E.T.A.; Saatchi, S.S.; Woodhouse, I.H.; Nangendo, G.; Ribeiro, N.S.; Williams, M.; Ryan, C.M.; Lewis, S.L.; Feldpausch, T.R.; Meir, P. Using satellite radar backscatter to predict above-ground woody biomass: A consistent relationship across four different African landscapes. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Saatchi, S.S.; McDonald, K.C. Coherent effects in microwave backscattering models for forest canopies. IEEE Trans. Geosci. Remote Sens. 1997, 35, 1032–1044. [Google Scholar] [CrossRef]
- Du, Y.; Ulaby, F.T.; Dobson, M.C. Sensitivity to soil moisture by active and passive microwave sensors. IEEE Trans. Geosci. Remote Sens. 2000, 38, 105–114. [Google Scholar] [CrossRef]
- Santos, J.R.; Freitas, C.C.; Araujo, L.S.; Dutra, L.V.; Mura, J.C.; Gama, F.F.; Soler, L.S.; SantAnna, S.J.S. Airborne P-band SAR applied to the aboveground biomass studies in the Brazilian tropical rainforest. Remote Sens. Environ. 2003, 87, 482–493. [Google Scholar] [CrossRef]
- Sandberg, G.; Ulander, L.M.H.; Fransson, J.E.S.; Holmgren, J.; Le Toan, T. L- and P-band backscatter intensity for biomass retrieval in hemiboreal forest. Remote Sens. Environ. 2011, 115, 2874–2886. [Google Scholar] [CrossRef]
- Saatchi, S.; Marlier, M.; Chazdon, R.L.; Clark, D.B.; Russell, A.E. Impact of spatial variability of tropical forest structure on radar estimation of aboveground biomass. Remote Sens. Environ. 2011, 115, 2836–2849. [Google Scholar] [CrossRef]
- Lucas, R.; Armston, J.; Fairfax, R.; Fensham, R.; Accad, A.; Carreiras, J.; Kelley, J.; Bunting, P.; Clewley, D.; Bray, S. An evaluation of the ALOS PALSAR L-band backscatter—Above ground biomass relationship Queensland, Australia: Impacts of surface moisture condition and vegetation structure. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2010, 3, 576–593. [Google Scholar] [CrossRef]
- Cartus, O.; Santoro, M.; Kellndorfer, J. Mapping forest aboveground biomass in the Northeastern United States with ALOS PALSAR dual-polarization L-band. Remote Sens. Environ. 2012, 124, 466–478. [Google Scholar] [CrossRef]
- Dobson, M.C.; Ulaby, F.T.; LeToan, T.; Beaudoin, A.; Kasischke, E.S.; Christensen, N. Dependence of radar backscatter on coniferous forest biomass. IEEE Trans. Geosci. Remote Sens. 1992, 30, 412–415. [Google Scholar] [CrossRef]
- Pulliainen, J.T.; Kurvonen, L.; Hallikainen, M.T. Multitemporal behavior of L- and C-band SAR observations of boreal forests. IEEE Trans. Geosci. Remote Sens. 1999, 37, 927–937. [Google Scholar] [CrossRef]
- Robinson, C.; Saatchi, S.; Neumann, M.; Gillespie, T. Impacts of spatial variability on aboveground biomass estimation from L-band radar in a temperate forest. Remote Sens. 2013, 5, 1001–1023. [Google Scholar] [CrossRef]
- Neumann, M.; Saatchi, S.S.; Ulander, L.M.H.; Fransson, J.E.S. Assessing Performance of L- and P-Band Polarimetric Interferometric SAR Data in Estimating Boreal Forest Above-Ground Biomass. IEEE Trans. Geosci. Remote Sens. 2012, 50, 714–726. [Google Scholar] [CrossRef]
- Mermoz, S.; Réjou-Méchain, M.; Villard, L.; Le Toan, T.; Rossi, V.; Gourlet-Fleury, S. Decrease of L-band SAR backscatter with biomass of dense forests. Remote Sens. Environ. 2015, 159, 307–317. [Google Scholar] [CrossRef]
- Hoekman, D.H.; Quiriones, M.J. Land cover type and biomass classification using AirSAR data for evaluation of monitoring scenarios in the Colombian Amazon. IEEE Trans. Geosci. Remote Sens. 2000, 38, 685–696. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Houghton, R.A.; Alvala, R.C.; Soares, J.V.; Yu, Y. Distribution of aboveground live biomass in the Amazon basin. Glob. Chang. Biol. 2007, 13, 816–837. [Google Scholar] [CrossRef]
- Papathanassiou, K.P.; Cloude, S.R. Single-baseline polarimetric SAR interferometry. IEEE Trans. Geosci. Remote Sens. 2001, 39, 2352–2363. [Google Scholar] [CrossRef]
- Cloude, S.R. Dual-baseline coherence tomography. IEEE Geosci. Remote Sens. Lett. 2007, 4, 127–131. [Google Scholar] [CrossRef]
- Balzter, H. Forest mapping and monitoring with interferometric synthetic aperture radar (InSAR). Prog. Phys. Geogr. 2001, 25, 159–177. [Google Scholar] [CrossRef]
- Hajnsek, I.; Kugler, F.; Lee, S.K.; Papathanassiou, K.P. Tropical-forest-parameter estimation by means of Pol-InSAR: The INDREX-II campaign. IEEE Trans. Geosci. Remote Sens. 2009, 47, 481–493. [Google Scholar] [CrossRef]
- Mitchard, E.T.A.; Saatchi, S.S.; White, L.J.T.; Abernethy, K.A.; Jeffery, K.J.; Lewis, S.L.; Collins, M.; Lefsky, M.A.; Leal, M.E.; Woodhouse, I.H.; et al. Mapping tropical forest biomass with radar and spaceborne LiDAR: Overcoming problems of high biomass and persistent cloud. Biogeosci. Discuss. 2011, 8, 8781–8815. [Google Scholar] [CrossRef]
- Shimada, M.; Ohtaki, T. Generating large-scale high-quality SAR Mosaic datasets: Application to PALSAR data for global monitoring. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2010, 3, 637–656. [Google Scholar] [CrossRef]
- Lefsky, M.A. A global forest canopy height map from the moderate resolution imaging spectroradiometer and the geoscience laser altimeter system. Geophys. Res. Lett. 2010, 37, L15401. [Google Scholar] [CrossRef]
- Los, S.O.; Rosette, J.A.B.; Kljun, N.; North, P.R.J.; Chasmer, L.; Suárez, J.C.; Hopkinson, C.; Hill, R.A.; van Gorsel, E.; Mahoney, C.; et al. Vegetation height and cover fraction between 60°S and 60°N from ICESat GLAS data. Geosci. Model. Dev. 2012, 5, 413–432. [Google Scholar] [CrossRef]
- Simard, M.; Pinto, N.; Fisher, J.B.; Baccini, A. Mapping forest canopy height globally with spaceborne lidar. J. Geophys. Res. 2011, 116, G04021. [Google Scholar] [CrossRef]
- Neigh, C.S.R.; Nelson, R.F.; Ranson, K.J.; Margolis, H.A.; Montesano, P.M.; Sun, G.; Kharuk, V.; Næsset, E.; Wulder, M.A.; Andersen, H.-E. Taking stock of circumboreal forest carbon with ground measurements, airborne and spaceborne LiDAR. Remote Sens. Environ. 2013, 137, 274–287. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Keller, M.; Pang, Y.; De Camargo, P.B.; Hunter, M.O. Revised method for forest canopy height estimation from Geoscience Laser Altimeter System waveforms. J. Appl. Remote Sens. 2007, 1, 013537. [Google Scholar]
- Popescu, S.C.; Zhao, K.; Neuenschwander, A.; Lin, C. Satellite LIDAR vs. small footprint airborne LIDAR: Comparing the accuracy of aboveground biomass estimates and forest structure metrics at footprint level. Remote Sens. Environ. 2011, 115, 2786–2797. [Google Scholar] [CrossRef]
- Arino, O.; Bicheron, P.; Achard, F.; Latham, J.; Witt, R.; Weber, J.-L. GLOBCOVER—The most detailed portrait of Earth. ESA Bull. Eur. Space Agency 2008, 136, 24–31. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C. Terrestrial ecoregions of the world: A new map of life on earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Yu, Y. Global Distribution of Carbon Stock in Live Woody Vegetation; University of California, Los Angeles: Los Angeles, CA, USA.
- Saatchi, S.; Moghaddam, M. Estimation of crown and stem water content and biomass of boreal forest using polarimetric SAR imagery. IEEE Trans. Geosci. Remote Sens. 2000, 38, 697–709. [Google Scholar] [CrossRef]
- Askne, J.; Santoro, M.; Smith, G.; Fransson, J.E.S. Multitemporal repeat-pass SAR interferometry of boreal forests. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1540–1550. [Google Scholar] [CrossRef]
- Hensley, S.; Oveisgharan, S.; Saatchi, S.; Simard, M.; Ahmed, R.; Haddad, Z. An error model for biomass estimates derived from polarimetric radar backscatter. IEEE Trans. Geosci. Remote Sens. 2014, 52, 4065–4082. [Google Scholar] [CrossRef]
- Chauhan, N.S.; Lang, R.H.; Ranson, K.J. Radar modeling of a boreal forest. IEEE Trans. Geosci. Remote Sens. 1991, 29, 627–638. [Google Scholar] [CrossRef]
- Karam, M.A.; Fung, A.K.; Lang, R.H.; Chauhan, N.S. A microwave scattering model for layered vegetation. IEEE Trans. Geosci. Remote Sens. 1992, 30, 767–784. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WWF Biome | GlobCover * | Category |
|---|---|---|
| Tropical Moist Broadleaf | 40 | Americas Tropical Moist |
| Tropical Moist Broadleaf | 40 | Africa Tropical Moist |
| Tropical Moist Broadleaf | 40 | Asia Tropical Moist |
| Temperate Broadleaf/Mixed | 50, 60 | Temperate Broadleaf |
| Temperate Conifer | 70, 90 | Temperate Conifer |
| Tropical Savanna/Shrub | 110, 120, 130 | Tropical Shrublands |
| Tropical Dry Broadleaf | 110, 120, 130 | Tropical Dry Broadleaf |
| Boreal + Tundra | 70, 90 | America Boreal |
| Boreal + Tundra | 70, 90 | Eurasia Boreal |
| Tropical Moist Broadleaf | 160 | Swamp Forest/Fresh Water |
| Tropical Moist Broadleaf | 170 | Mangrove/Saline Water |
| Category | A | B | C | α |
|---|---|---|---|---|
| Africa Tropical Moist | 0.056492 | 0.064689 | 0 | 0.038247 |
| Asia Tropical Moist | 0.045409 | 0.060518 | 0 | 0.060518 |
| America Tropical Moist | 0.040546 | 0.068784 | 0 | 0.098841 |
| Temperate Conifer | 0.0092565 | 0.057336 | 0.04 | 0.27162 |
| Temperate Broadleaf/Mixed | 0.041469 | 0.034296 | 0.026406 | 0.012282 |
| Tropical Shrubland | 0.016429 | 0.11013 | 0 | 0.2675 |
| Tropical Dry Broadleaf | 0.021563 | 0.042324 | 0.027519 | 0.1117 |
| North America Boreal | 0.018911 | 0.019744 | 0.029106 | 0.15723 |
| Eurasia Boreal | 0.0091605 | 0.038506 | 0.04 | 0.26141 |
| Fresh Water Flooded | 0.047845 | 0.045581 | 0.022164 | 0.0058592 |
| Saline Water flooded | 0.013682 | 0.051846 | 0.02192 | 0.21116 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Saatchi, S. Sensitivity of L-Band SAR Backscatter to Aboveground Biomass of Global Forests. Remote Sens. 2016, 8, 522. https://doi.org/10.3390/rs8060522
Yu Y, Saatchi S. Sensitivity of L-Band SAR Backscatter to Aboveground Biomass of Global Forests. Remote Sensing. 2016; 8(6):522. https://doi.org/10.3390/rs8060522
Chicago/Turabian StyleYu, Yifan, and Sassan Saatchi. 2016. "Sensitivity of L-Band SAR Backscatter to Aboveground Biomass of Global Forests" Remote Sensing 8, no. 6: 522. https://doi.org/10.3390/rs8060522