

Variability in Symbiont Chlorophyll of Hawaiian Corals from Field and Airborne Spectroscopy

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites
2.2. Field Measurements
2.3. Laboratory Assays
Chl c2 = 27.09 ((A630 − A750)/PL) − 3.63 ((A663 − A750)/PL)
2.4. Airborne Remote Sensing
2.5. Chemometric Analyses
3. Results
3.1. Field and Airborne Spectroscopy
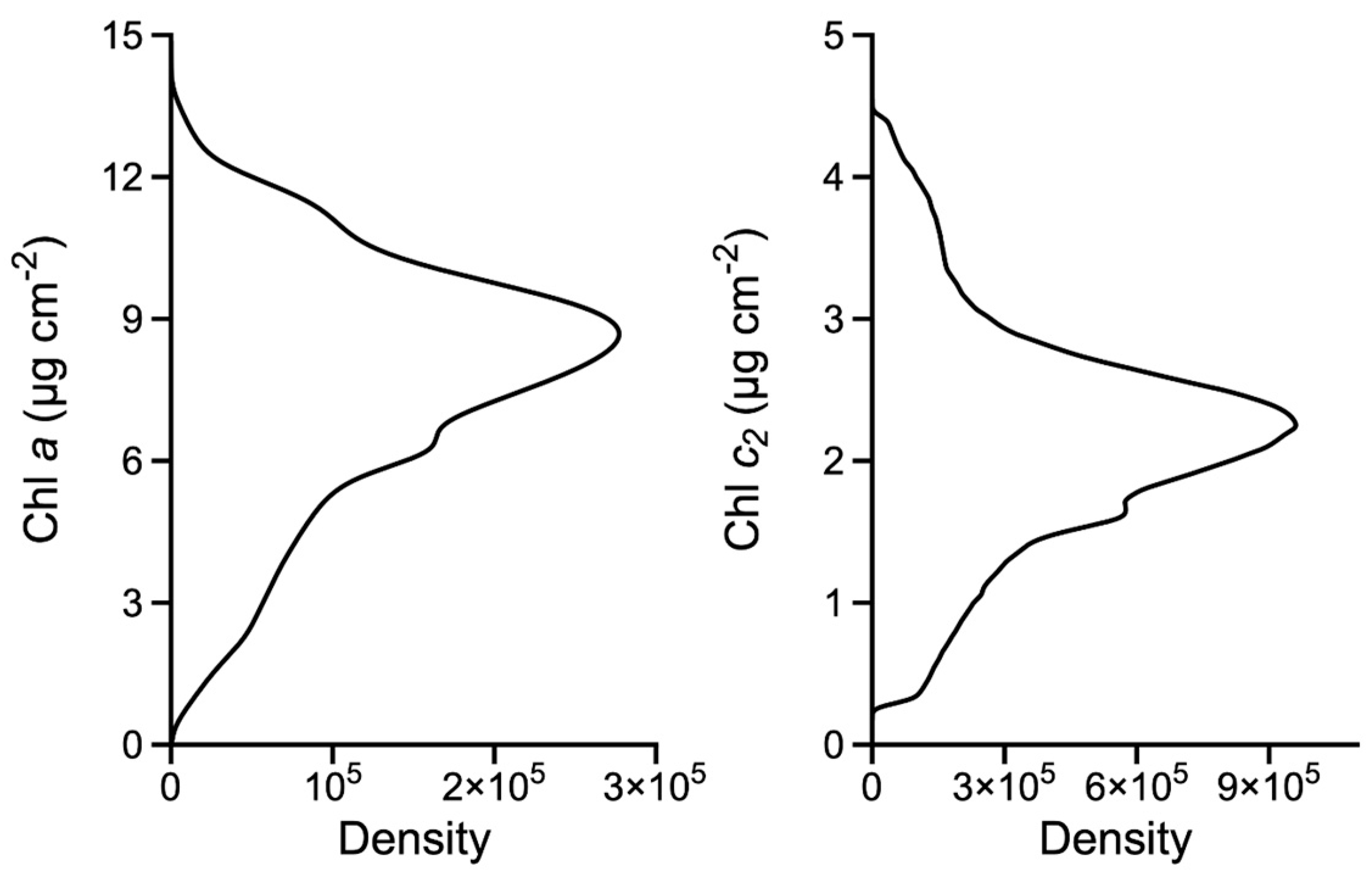
3.2. Chlorophyll Chemistry
3.3. Spectroscopic Estimation of Chlorophyll in Corals
3.4. Application to Airborne Imaging Spectroscopy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veron, J.E.; Hoegh-Guldberg, O.; Lenton, T.M.; Lough, J.M.; Obura, D.O.; Pearce-Kelly, P.; Sheppard, C.R.; Spalding, M.; Stafford-Smith, M.G.; Rogers, A.D. The coral reef crisis: The critical importance of <350 ppm CO2. Mar. Pollut. Bull. 2009, 58, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef]
- Obura, D.O.; Aeby, G.; Amornthammarong, N.; Appeltans, W.; Bax, N.; Bishop, J.; Brainard, R.E.; Chan, S.; Fletcher, P.; Gordon, T.A. Coral reef monitoring, reef assessment technologies, and ecosystem-based management. Front. Mar. Sci. 2019, 6, 580. [Google Scholar] [CrossRef]
- Eddy, T.D.; Cheung, W.W.L.; Bruno, J.F. Historical baselines of coral cover on tropical reefs as estimated by expert opinion. PeerJ 2018, 6, e4308. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef]
- Mumby, P.J.; Skirving, W.; Strong, A.E.; Hardy, J.T.; LeDrew, E.F.; Hochberg, E.J.; Stumpf, R.P.; David, L.T. Remote sensing of coral reefs and their physical environment. Mar. Pollut. Bull. 2004, 48, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Drury, C.; Martin, R.E.; Knapp, D.E.; Heckler, J.; Levy, J.; Gates, R.D.; Asner, G.P. Ecosystem-scale mapping of coral species and thermal tolerance. Front. Ecol. Environ. 2022, 20, 285–291. [Google Scholar] [CrossRef]
- Drury, C.; Lirman, D. Genotype by environment interactions in coral bleaching. Proc. R. Soc. B 2021, 288, 20210177. [Google Scholar] [CrossRef]
- Lundgren, P.; Vera, J.C.; Peplow, L.; Manel, S.; van Oppen, M.J. Genotype–environment correlations in corals from the Great Barrier Reef. BMC Genet. 2013, 14, 9. [Google Scholar] [CrossRef]
- Schweinsberg, M.; Weiss, L.C.; Striewski, S.; Tollrian, R.; Lampert, K.P. More than one genotype: How common is intracolonial genetic variability in scleractinian corals? Mol. Ecol. 2015, 24, 2673–2685. [Google Scholar] [CrossRef]
- Baird, A.H.; Bhagooli, R.; Ralph, P.J.; Takahashi, S. Coral bleaching: The role of the host. Trends Ecol. Evol. 2009, 24, 16–20. [Google Scholar] [CrossRef]
- Knowlton, N. The future of coral reefs. Proc. Natl. Acad. Sci. USA 2001, 98, 5419–5425. [Google Scholar] [CrossRef]
- Bahr, K.D.; Severino, S.J.; Tsang, A.O.; Han, J.J.; Richards Dona, A.; Stender, Y.O.; Weible, R.M.; Graham, A.; McGowan, A.E.; Rodgers, K.S. The Hawaiian Ko‘a Card: Coral health and bleaching assessment color reference card for Hawaiian corals. SN Appl. Sci. 2020, 2, 1706. [Google Scholar] [CrossRef]
- Siebeck, U.; Marshall, N.; Klüter, A.; Hoegh-Guldberg, O. Monitoring coral bleaching using a colour reference card. Coral Reefs 2006, 25, 453–460. [Google Scholar] [CrossRef]
- Roth, M.S. The engine of the reef: Photobiology of the coral-algal symbiosis. Front. Microbiol. 2014, 5, 422. [Google Scholar] [CrossRef]
- Dellisanti, W.; Chung, J.T.; Chow, C.F.; Wu, J.; Wells, M.L.; Chan, L.L. Experimental techniques to assess coral physiology in situ under global and local stressors: Current approaches and novel insights. Front. Physiol. 2021, 12, 594. [Google Scholar] [CrossRef]
- Donovan, M.K.; Alves, C.; Burns, J.; Drury, C.; Meier, O.W.; Ritson-Williams, R.; Cunning, R.; Dunn, R.P.; Goodbody-Gringley, G.; Henderson, L.M. From polyps to pixels: Understanding coral reef resilience to local and global change across scales. Landsc. Ecol. 2023, 38, 737–752. [Google Scholar] [CrossRef]
- Hedley, J.D.; Mumby, P.J. Biological and remote sensing perspectives of pigmentation in coral reef organisms. Adv. Mar. Biol. 2002, 43, 277–317. [Google Scholar] [PubMed]
- Torres-Perez, J.L.; Guild, L.S.; Armstrong, R.A.; Corredor, J.; Zuluaga-Montero, A.; Polanco, R. Relative Pigment Composition and Remote Sensing Reflectance of Caribbean Shallow-Water Corals. PLoS ONE 2015, 10, e0143709. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, E.J.; Apprill, A.M.; Atkinson, M.J.; Bidigare, R.R. Bio-optical modeling of photosynthetic pigments in corals. Coral Reefs 2006, 25, 99–109. [Google Scholar] [CrossRef]
- Joyce, K.E.; Phinn, S.R. Hyperspectral analysis of chlorophyll content and photosynthetic capacity of coral reef substrates. Limnol. Oceanogr. 2003, 48, 489–496. [Google Scholar] [CrossRef]
- Torres-Pérez, J.; Guild, L.; Armstrong, R. Hyperspectral Distinction of Two Caribbean Shallow-Water Corals Based on Their Pigments and Corresponding Reflectance. Remote Sens. 2012, 4, 3813–3832. [Google Scholar] [CrossRef]
- Hochberg, E. Spectral reflectance of coral reef bottom-types worldwide and implications for coral reef remote sensing. Remote Sens. Environ. 2003, 85, 159–173. [Google Scholar] [CrossRef]
- Hedley, J.D.; Roelfsema, C.M.; Chollett, I.; Harborne, A.R.; Heron, S.F.; Weeks, S.; Skirving, W.J.; Strong, A.E.; Eakin, C.M.; Christensen, T.R. Remote sensing of coral reefs for monitoring and management: A review. Remote Sens. 2016, 8, 118. [Google Scholar] [CrossRef]
- Thompson, D.R.; Hochberg, E.J.; Asner, G.P.; Green, R.O.; Knapp, D.E.; Gao, B.-C.; Garcia, R.; Gierach, M.; Lee, Z.; Maritorena, S. Airborne mapping of benthic reflectance spectra with Bayesian linear mixtures. Remote Sens. Environ. 2017, 200, 18–30. [Google Scholar] [CrossRef]
- Asner, G.P.; Vaughn, N.R.; Heckler, J.; Knapp, D.E.; Balzotti, C.; Shafron, E.; Martin, R.E.; Neilson, B.J.; Gove, J.M. Large-scale mapping of live corals to guide reef conservation. Proc. Natl. Acad. Sci. USA 2020, 117, 33711–33718. [Google Scholar] [CrossRef]
- Asner, G.P.; Vaughn, N.R.; Martin, R.E.; Foo, S.A.; Heckler, J.; Neilson, B.J.; Gove, J.M. Mapped coral mortality and refugia in an archipelago-scale marine heat wave. Proc. Natl. Acad. Sci. USA 2022, 119, e2123331119. [Google Scholar] [CrossRef]
- Caruso, C.; Rocha de Souza, M.; Ruiz-Jones, L.; Conetta, D.; Hancock, J.; Hobbs, C.; Hobbs, C.; Kahkejian, V.; Kitchen, R.; Marin, C. Genetic patterns in Montipora capitata across an environmental mosaic in Kāne’ohe Bay, O’ahu, Hawai’i. Mol. Ecol. 2022, 31, 5201–5213. [Google Scholar] [CrossRef]
- Fitt, W.K.; McFarland, F.K.; Warner, M.E.; Chilcoat, G.C. Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnol. Oceanogr. 2000, 45, 677–685. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, H.; Kang, Y.; Bina, D.; Lo, C.S.; Blankenship, R.E. Characterization of the peridinin-chlorophyll a-protein complex in the dinoflagellate Symbiodinium. Biochim. Biophys. Acta 2012, 1817, 983–989. [Google Scholar] [CrossRef]
- Niedzwiedzki, D.M.; Jiang, J.; Lo, C.S.; Blankenship, R.E. Spectroscopic properties of the Chlorophyll a-Chlorophyll c 2-Peridinin-Protein-Complex (acpPC) from the coral symbiotic dinoflagellate Symbiodinium. Photosynth. Res. 2014, 120, 125–139. [Google Scholar] [CrossRef]
- Jeffrey, S.t.; Humphrey, G. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Veal, C.; Holmes, G.; Nunez, M.; Hoegh-Guldberg, O.; Osborn, J. A comparative study of methods for surface area and three-dimensional shape measurement of coral skeletons. Limnol. Oceanogr. Methods 2010, 8, 241–253. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Boardman, J.; Green, R.O.; Kennedy-Bowdoin, T.; Eastwood, M.; Martin, R.E.; Anderson, C.; Field, C.B. Carnegie Airborne Observatory-2: Increasing science data dimensionality via high-fidelity multi-sensor fusion. Remote Sens. Environ. 2012, 124, 454–465. [Google Scholar] [CrossRef]
- Gao, B.C.; Goetz, A.F. Column atmospheric water vapor and vegetation liquid water retrievals from airborne imaging spectrometer data. J. Geophys. Res. Atmos. 1990, 95, 3549–3564. [Google Scholar] [CrossRef]
- Thompson, D.R.; Gao, B.-C.; Green, R.O.; Roberts, D.A.; Dennison, P.E.; Lundeen, S.R. Atmospheric correction for global mapping spectroscopy: ATREM advances for the HyspIRI preparatory campaign. Remote Sens. Environ. 2015, 167, 64–77. [Google Scholar] [CrossRef]
- Feilhauer, H.; Asner, G.P.; Martin, R.E.; Schmidtlein, S. Brightness-normalized partial least squares regression for hyperspectral data. J. Quant. Spectrosc. Radiat. Transf. 2010, 111, 1947–1957. [Google Scholar] [CrossRef]
- Martens, H. Reliable and relevant modelling of real world data: A personal account of the development of PLS Regression. Chemom. Intell. Lab. Syst. 2001, 58, 85–95. [Google Scholar] [CrossRef]
- Chen, S.; Hong, X.; Harris, C.J.; Sharkey, P.M. Sparse modeling using orthogonal forest regression with PRESS statistic and regularization. IEEE Trans. Syst. Man. Cybern. 2004, 34, 898–911. [Google Scholar] [CrossRef]
- Harrison, D.E.; Asner, G.P. Sensitivity of spectral communities to shifts in benthic composition in Hawai‘i. Remote Sens. Environ. 2024. [Google Scholar] [CrossRef]
- Tulloch, V.J.; Klein, C.J.; Jupiter, S.D.; Tulloch, A.I.; Roelfsema, C.; Possingham, H.P. Trade-offs between data resolution, accuracy, and cost when choosing information to plan reserves for coral reef ecosystems. J. Environ. Manag. 2017, 188, 108–119. [Google Scholar] [CrossRef]
- Mumby, P.; Green, E.; Edwards, A.; Clark, C. Coral reef habitat mapping: How much detail can remote sensing provide? Mar. Biol. 1997, 130, 193–202. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.; Carranza, L.; Martinez, P.; Houcheime, M.; Sinca, F.; Weiss, P. Spectroscopy of canopy chemicals in humid tropical forests. Remote Sens. Environ. 2011, 115, 3587–3598. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E. Quantifying forest canopy traits: Imaging spectroscopy versus field survey. Remote Sens. Environ. 2015, 158, 15–27. [Google Scholar] [CrossRef]
- Lesser, M.; Mobley, C. Bathymetry, water optical properties, and benthic classification of coral reefs using hyperspectral remote sensing imagery. Coral Reefs 2007, 26, 819–829. [Google Scholar] [CrossRef]
- Marco Agustín, L.-C.; Laura, F.-R.; Juan Sebastián, Z.-S.; Oscar, H.-R.; Carlos, L.-C. Correlation of chlorophyll a and total carotenoid concentrations with coral bleaching from locations on the Pacific coast of Mexico. Mar. Freshw. Behav. Physiol. 2006, 39, 279–291. [Google Scholar] [CrossRef]
- Zhao, M.; Yu, K. Application of chlorophyll fluorescence technique in the study of coral symbiotic zooxanthellae micro-ecology. Acta Ecol. Sin. 2014, 34, 165–169. [Google Scholar] [CrossRef]
- Wangpraseurt, D.; Larkum, A.W.; Ralph, P.J.; Kuhl, M. Light gradients and optical microniches in coral tissues. Front. Microbiol. 2012, 3, 316. [Google Scholar] [CrossRef] [PubMed]
- Cunning, R.; Ritson-Williams, R.; Gates, R.D. Patterns of bleaching and recovery of Montipora capitata in Kāne ‘ohe Bay, Hawai ‘i, USA. Mar. Ecol. Prog. Ser. 2016, 551, 131–139. [Google Scholar] [CrossRef]
- Wall, C.B.; Ritson-Williams, R.; Popp, B.N.; Gates, R.D. Spatial variation in the biochemical and isotopic composition of corals during bleaching and recovery. Limnol. Oceanogr. 2019, 64, 2011–2028. [Google Scholar] [CrossRef] [PubMed]
- Drury, C.; Bean, N.K.; Harris, C.I.; Hancock, J.R.; Huckeba, J.; Roach, T.N.; Quinn, R.A.; Gates, R.D. Intrapopulation adaptive variance supports thermal tolerance in a reef-building coral. Commun. Biol. 2022, 5, 486. [Google Scholar] [CrossRef] [PubMed]
- Pegau, W.S.; Gray, D.; Zaneveld, J.R.V. Absorption and attenuation of visible and near-infrared light in water: Dependence on temperature and salinity. Appl. Opt. 1997, 36, 6035–6046. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Xu, Z.; Yang, Y.; Liu, Y.; Zhao, H.; Zhang, Y.; Xie, B.; Zhou, W.; Li, C.; Cao, W. In situ hyperspectral characteristics and the discriminative ability of remote sensing to coral species in the South China Sea. GIScience Remote Sens. 2022, 59, 272–294. [Google Scholar] [CrossRef]
- Kok, J.; Bainbridge, S.; Olsen, M.; Rigby, P. Towards effective aerial drone-based hyperspectral remote sensing of coral reefs. In Proceedings of the Global Oceans 2020: Singapore–US Gulf Coast, Biloxi, MS, USA, 5–30 October 2020; pp. 1–6. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R2 | RMSE | %RMSE | ||
|---|---|---|---|---|
| Chl a (µg cm−2) | ||||
| Field | Training | 0.73 (0.57–0.82) | 1.82 (1.50–2.43) | 30.08 (25.39–38.38) |
| Testing | 0.64 (0.54–0.75) | 2.17 (1.80–2.75) | 34.31 (25.59–41.35) | |
| Model | 0.81 | 1.53 | 24.88 | |
| Airborne | Training | 0.76 (0.57–0.89) | 1.67 (1.18–2.39) | 27.65 (19.28–38.48) |
| Testing | 0.61 (0.51–0.73) | 2.21 (1.86–2.73) | 34.92 (26.01–42.50) | |
| Model | 0.84 | 1.40 | 22.76 | |
| Chl c2 (µg cm−2) | ||||
| Field | Training | 0.72 (0.54–0.81) | 0.51 (0.42–0.70) | 28.69 (22.95–37.63) |
| Testing | 0.56 (0.37–0.73) | 0.65 (0.53–0.82) | 35.39 (25.96–44.26) | |
| Model | 0.81 | 1.53 | 24.88 | |
| Airborne | Training | 0.72 (0.47–0.90) | 0.51 (0.31–0.76) | 28.61 (17.92–40.91) |
| Testing | 0.53 (0.36–0.71) | 0.67 (0.51–0.87) | 36.68 (28.22–47.90) | |
| Model | 0.82 | 0.42 | 23.20 | |
| Chl a (µg cm−2) | Chl c2 (µg cm−2) | Chl c2:a | |||||
|---|---|---|---|---|---|---|---|
| No. Samples (Pixels) | Mean ± SD | Median | Mean ± SD | Median | Mean ± SD | Median | |
| All | 1,442,258 | 7.68 ± 2.49 | 8.04 | 2.16 ± 0.78 | 2.17 | 0.28 ± 0.06 | 0.27 |
| By Depth (m) | |||||||
| 0–1 | 595,210 | 7.64 ± 2.26 | 8.06 | 2.08 ± 0.65 | 2.15 | 0.27 ± 0.05 | 0.26 |
| 1–2 | 518,693 | 7.41 ± 2.58 | 7.73 | 2.03 ± 0.79 | 2.05 | 0.27 ± 0.06 | 0.26 |
| 2–3 | 190,046 | 7.97 ± 2.40 | 8.40 | 2.30 ± 0.76 | 2.29 | 0.29 ± 0.07 | 0.28 |
| 3–4 | 82,901 | 8.48 ± 2.73 | 9.14 | 2.78 ± 0.85 | 2.79 | 0.34 ± 0.09 | 0.33 |
| 4–5 | 37,112 | 8.40 ± 3.14 | 9.30 | 2.97 ± 0.95 | 3.13 | 0.38 ± 0.10 | 0.35 |
| ≥5 | 18,296 | 8.27 ± 3.34 | 9.14 | 3.00 ± 1.01 | 3.20 | 0.39 ± 0.11 | 0.35 |
| By Reef | |||||||
| Reef 1 | 326,341 | 7.87 ± 2.75 | 8.30 | 2.41 ± 0.91 | 2.36 | 0.31 ± 0.08 | 0.30 |
| Reef 4 and 5 | 330,740 | 8.03 ± 2.16 | 8.57 | 2.23 ± 0.67 | 2.29 | 0.28 ± 0.06 | 0.27 |
| Reef 13 | 394,160 | 7.55 ± 2.30 | 7.98 | 2.02 ± 0.66 | 2.09 | 0.27 ± 0.05 | 0.26 |
| Reef 25 | 105,985 | 7.13 ± 1.78 | 7.13 | 1.94 ± 0.49 | 1.91 | 0.28 ± 0.05 | 0.26 |
| Reef 42 | 65,712 | 6.76 ± 2.70 | 7.10 | 1.99 ± 0.92 | 1.97 | 0.30 ± 0.08 | 0.27 |
| Reef 44 | 219,320 | 7.62 ± 2.91 | 7.69 | 2.12 ± 0.90 | 2.08 | 0.28 ± 0.06 | 0.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asner, G.P.; Drury, C.; Vaughn, N.R.; Hancock, J.R.; Martin, R.E. Variability in Symbiont Chlorophyll of Hawaiian Corals from Field and Airborne Spectroscopy. Remote Sens. 2024, 16, 732. https://doi.org/10.3390/rs16050732
Asner GP, Drury C, Vaughn NR, Hancock JR, Martin RE. Variability in Symbiont Chlorophyll of Hawaiian Corals from Field and Airborne Spectroscopy. Remote Sensing. 2024; 16(5):732. https://doi.org/10.3390/rs16050732
Chicago/Turabian StyleAsner, Gregory P., Crawford Drury, Nicholas R. Vaughn, Joshua R. Hancock, and Roberta E. Martin. 2024. "Variability in Symbiont Chlorophyll of Hawaiian Corals from Field and Airborne Spectroscopy" Remote Sensing 16, no. 5: 732. https://doi.org/10.3390/rs16050732