

The Seasonality of Eddy-Induced Chlorophyll-a Anomalies in the Kuroshio Extension System
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. In Situ Data
2.1.2. Satellite Observations
2.1.3. Model Data
2.2. Methods
2.2.1. Composite Analysis
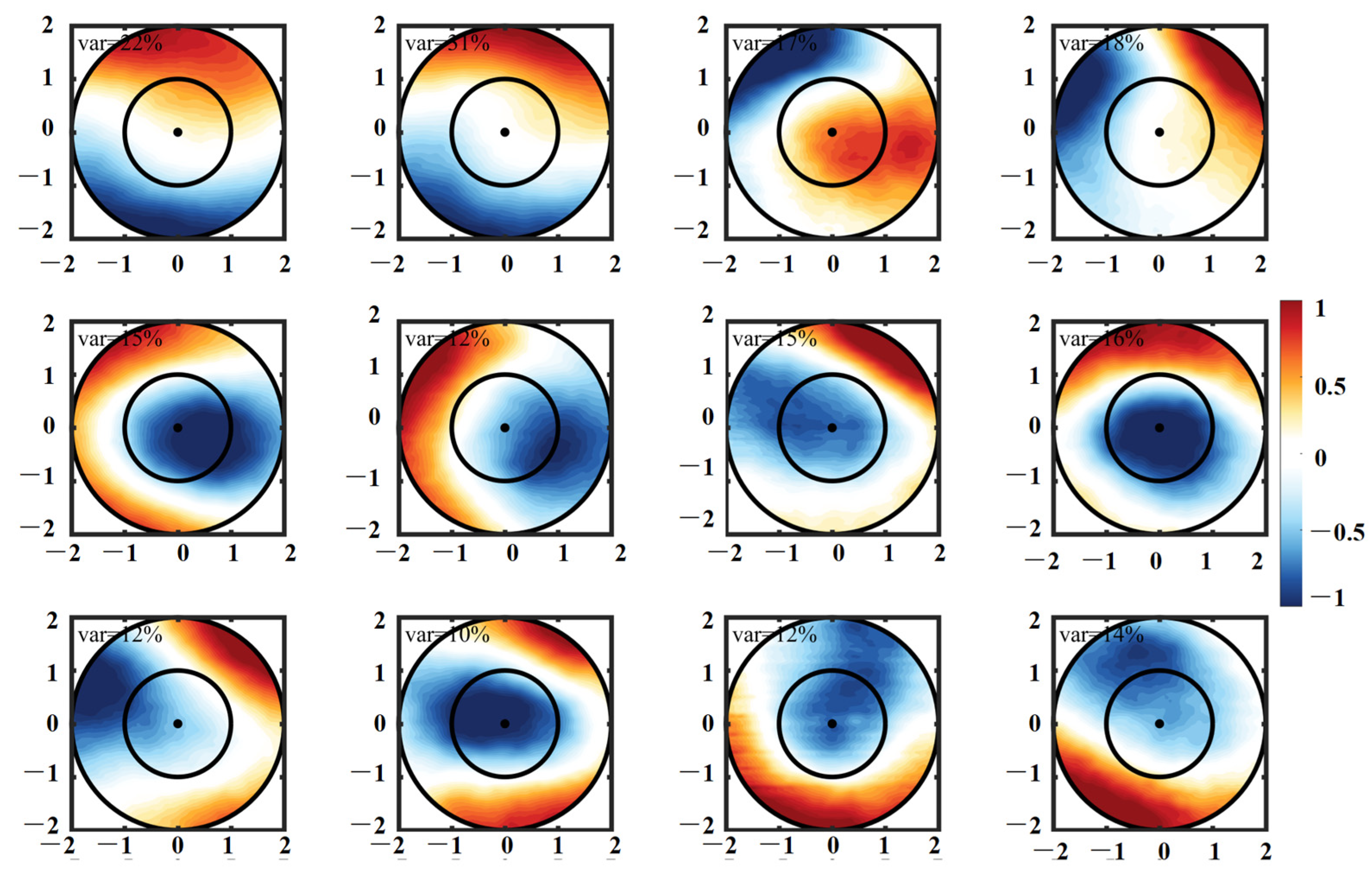
2.2.2. EOF Analysis
2.2.3. Mixed Layer Depth
2.2.4. Mixed Layer Nutrient Budget
3. Results
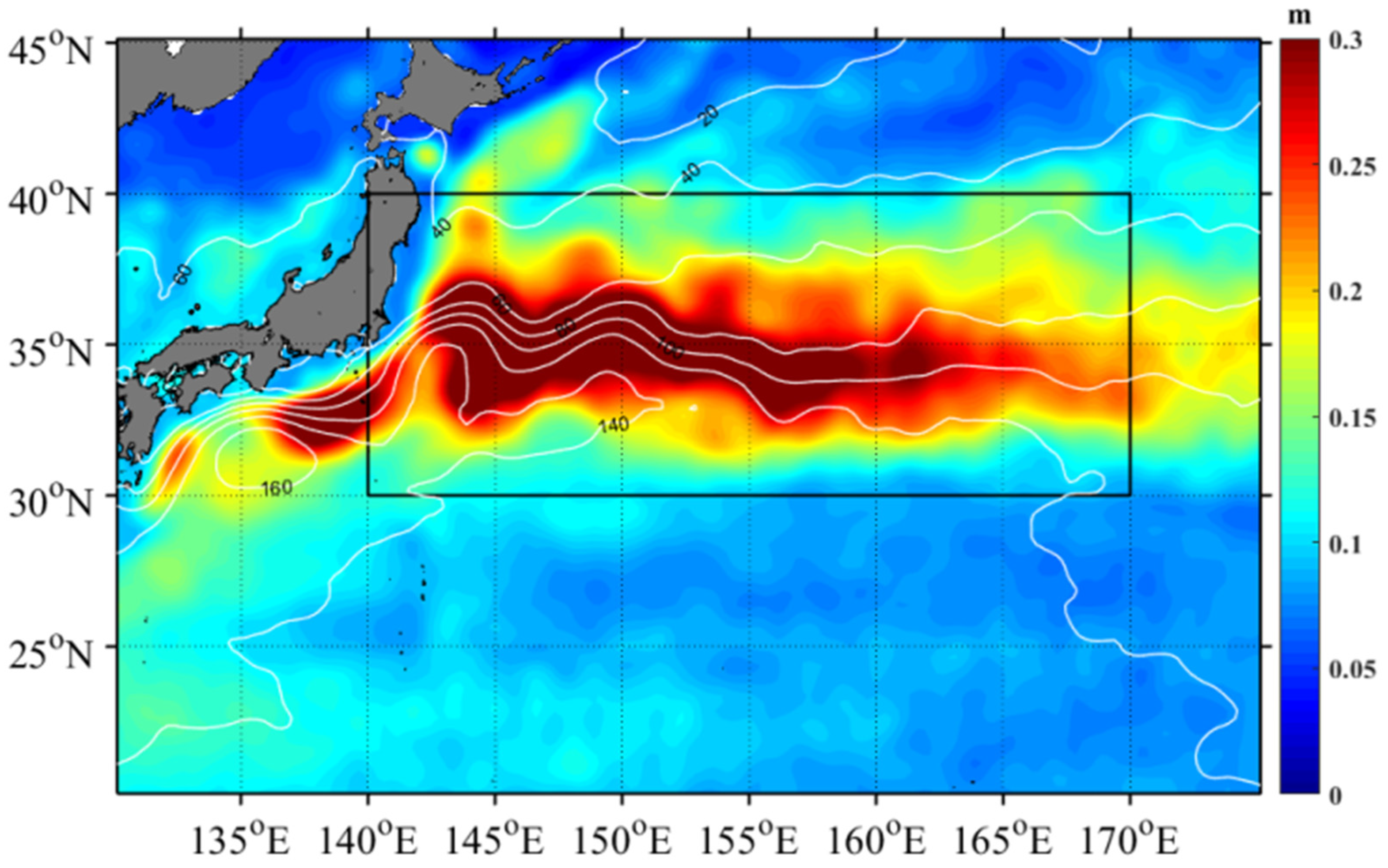
3.1. Eddy Properties
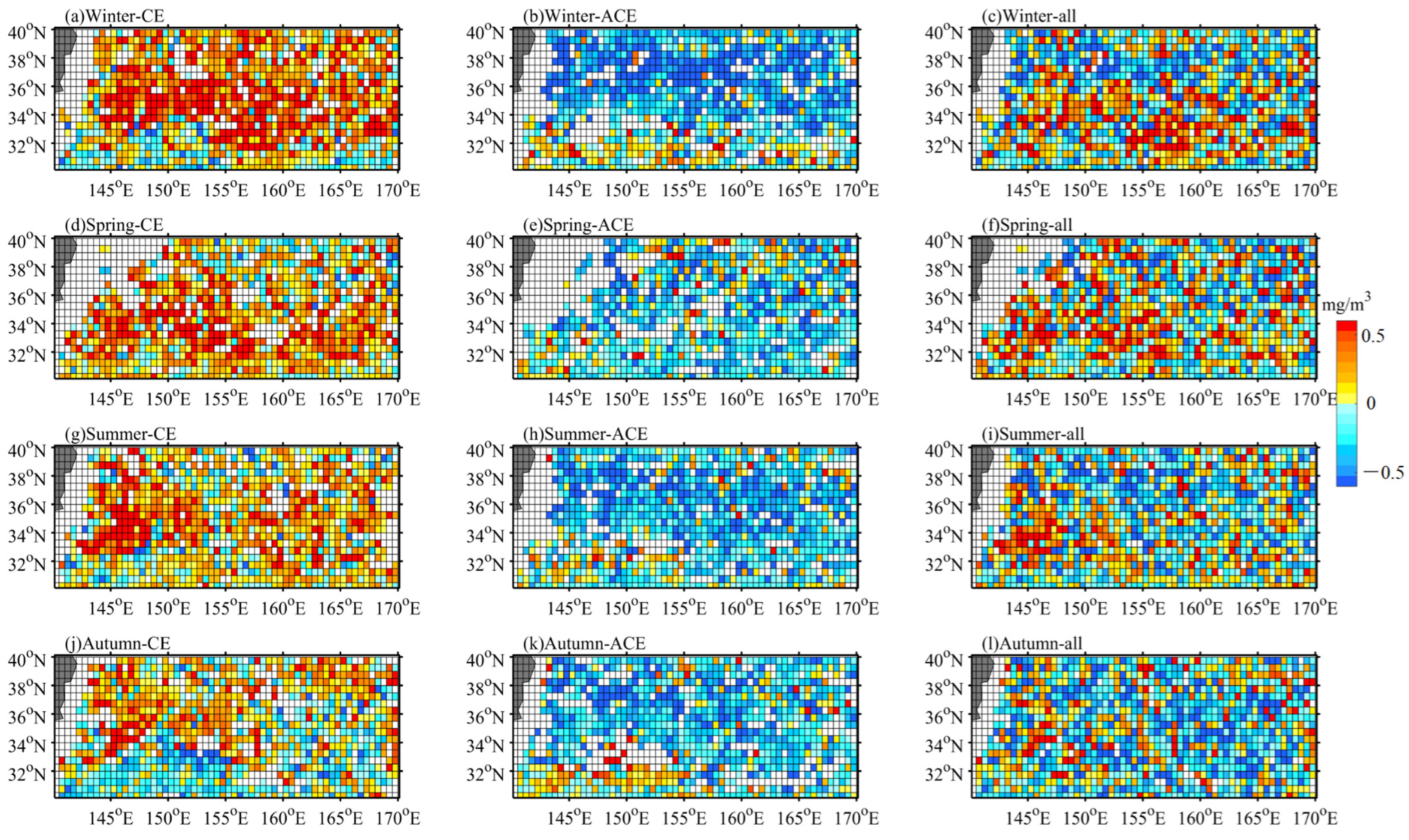
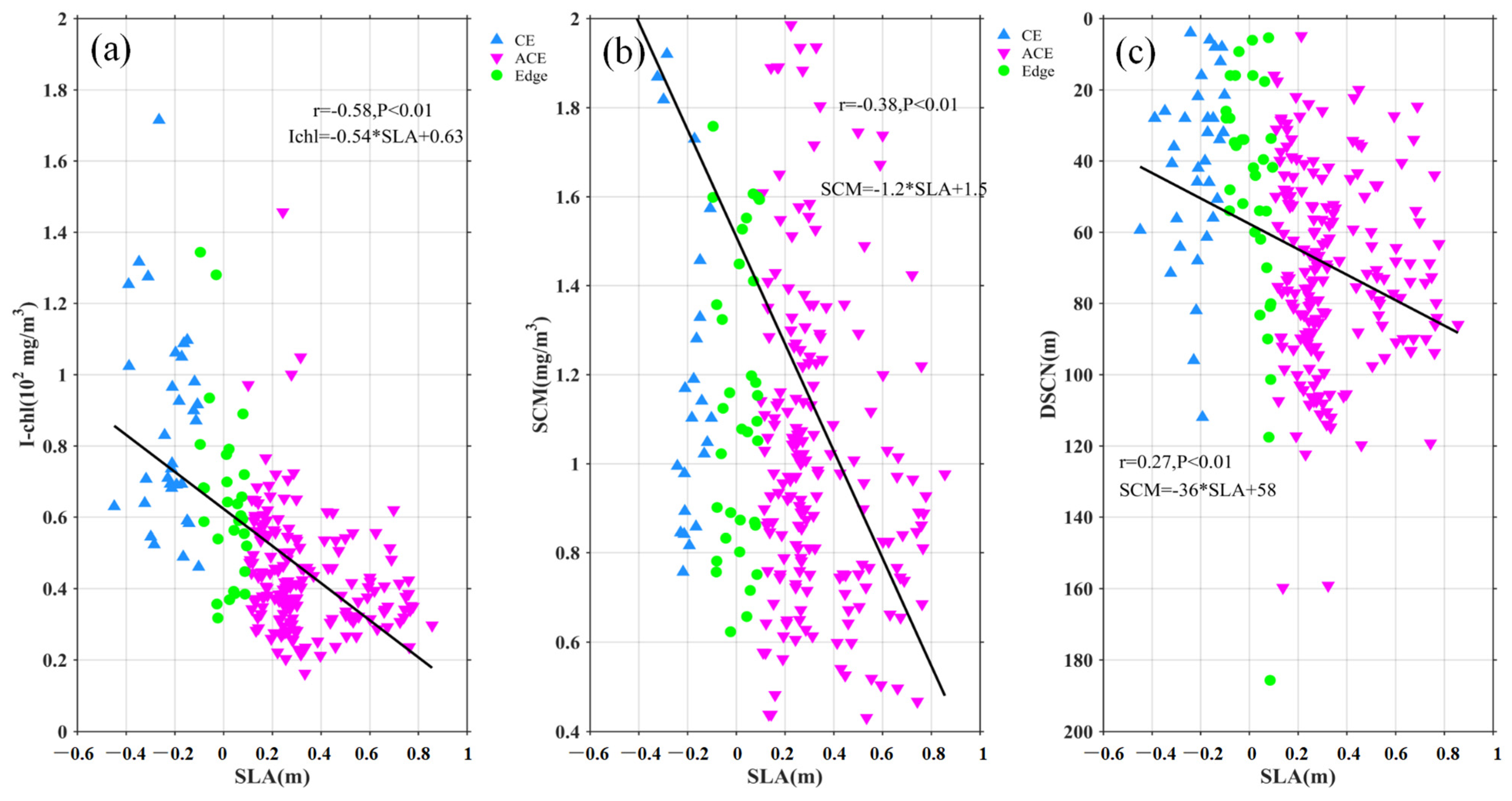
3.2. Relationship between Eddy and Surface Chl-a
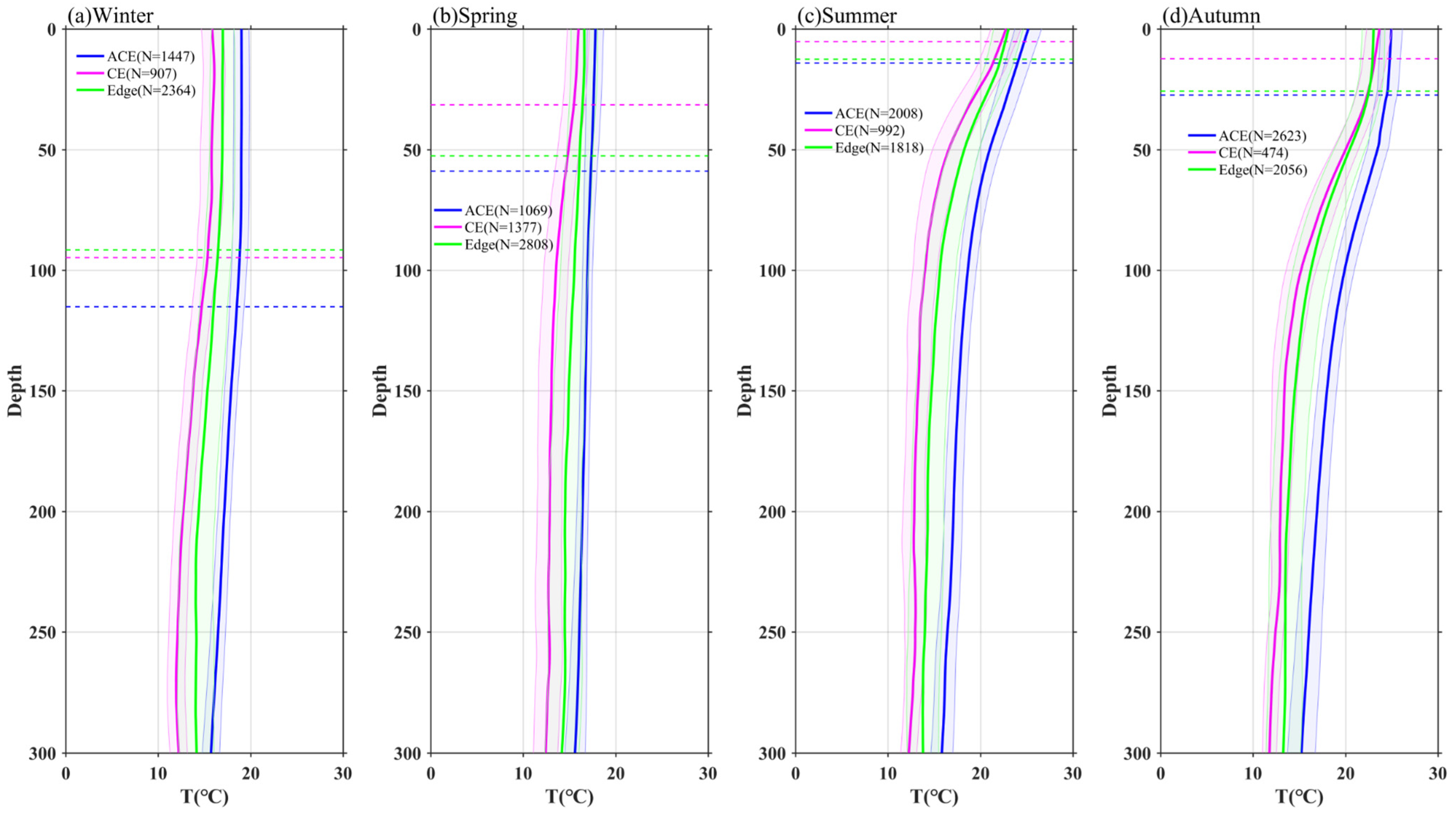
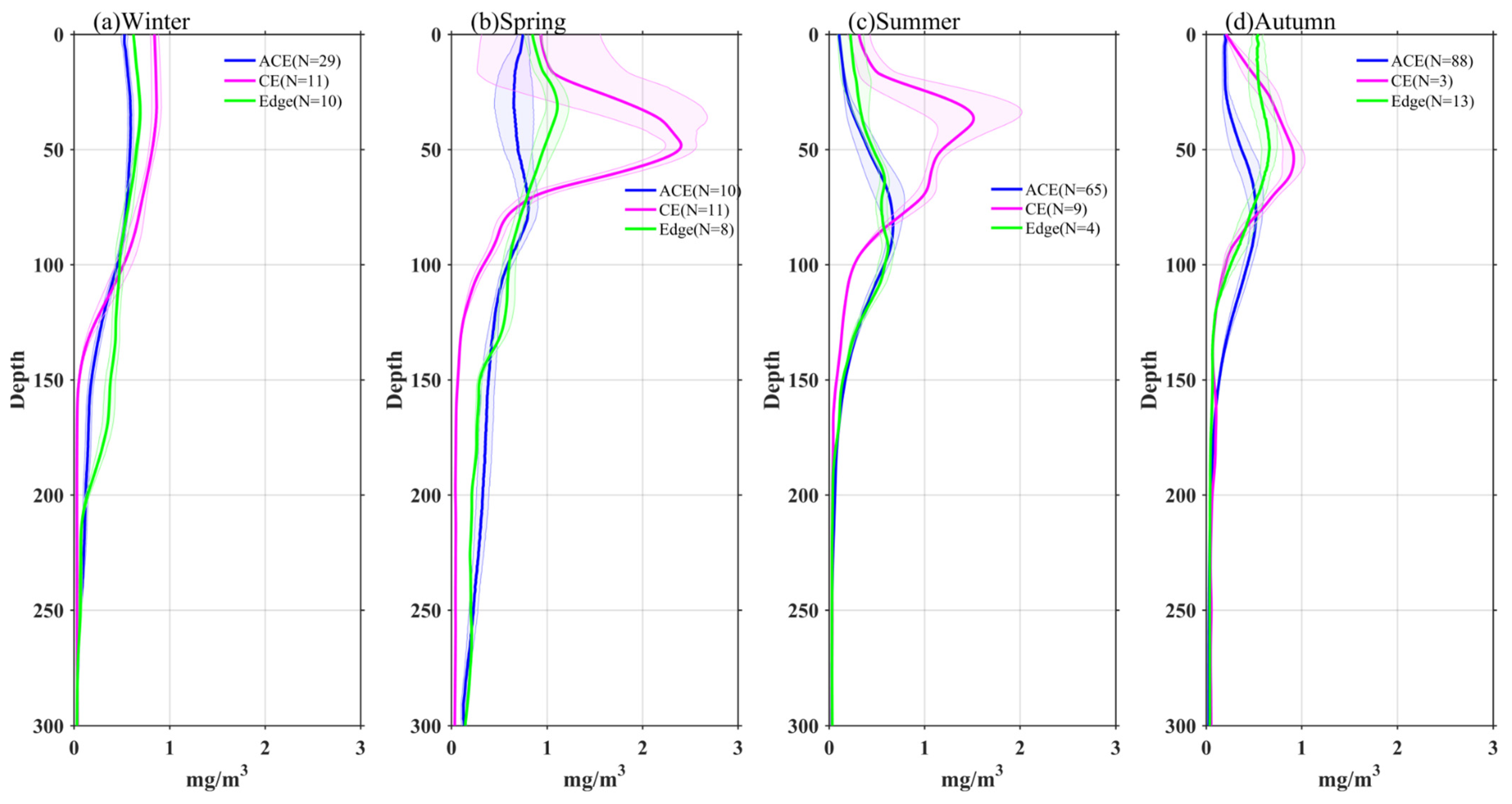
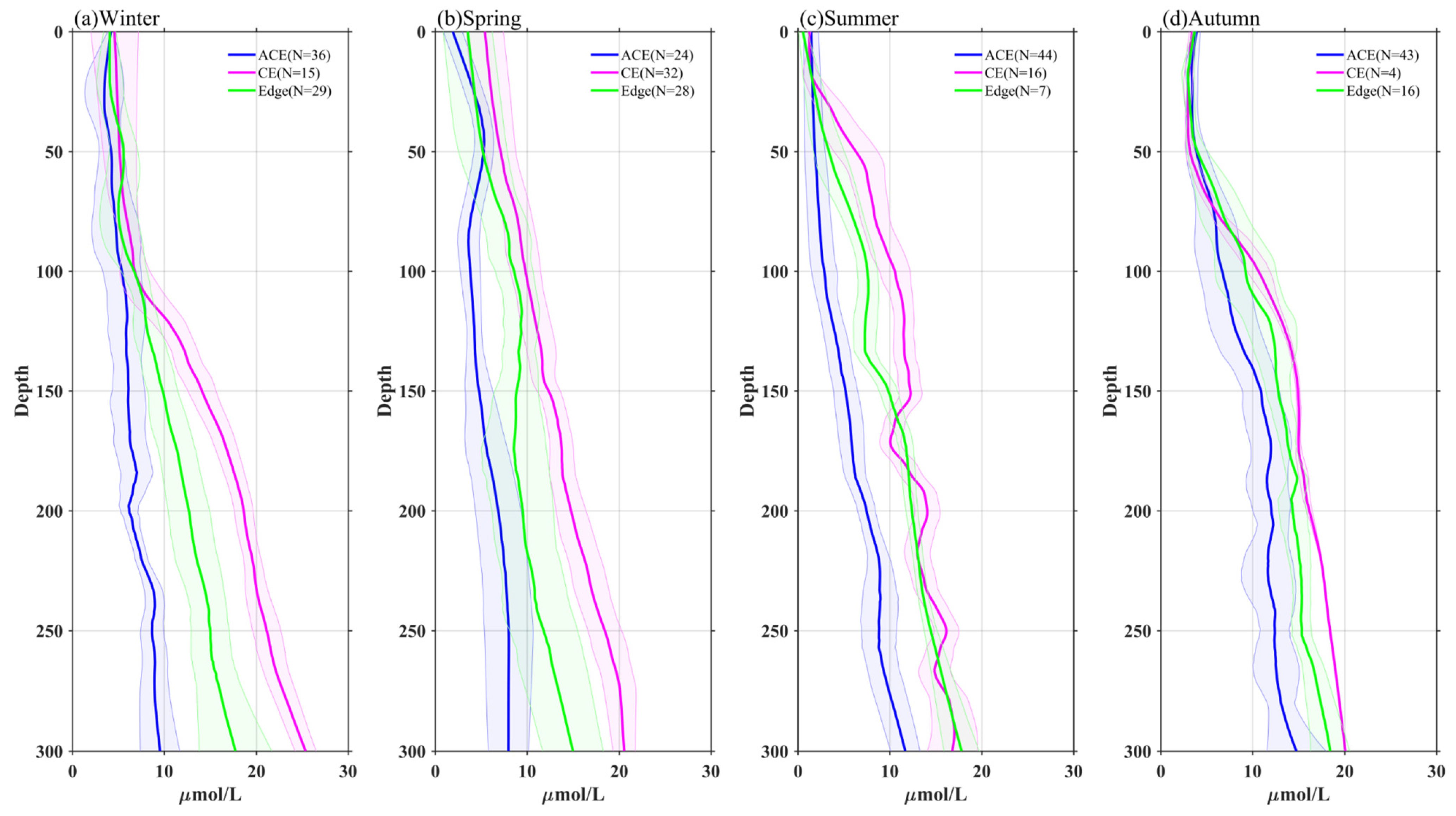
3.3. Vertical Profiles of Temperature, Chl-a, and Nitrate in Eddies
4. Discussion
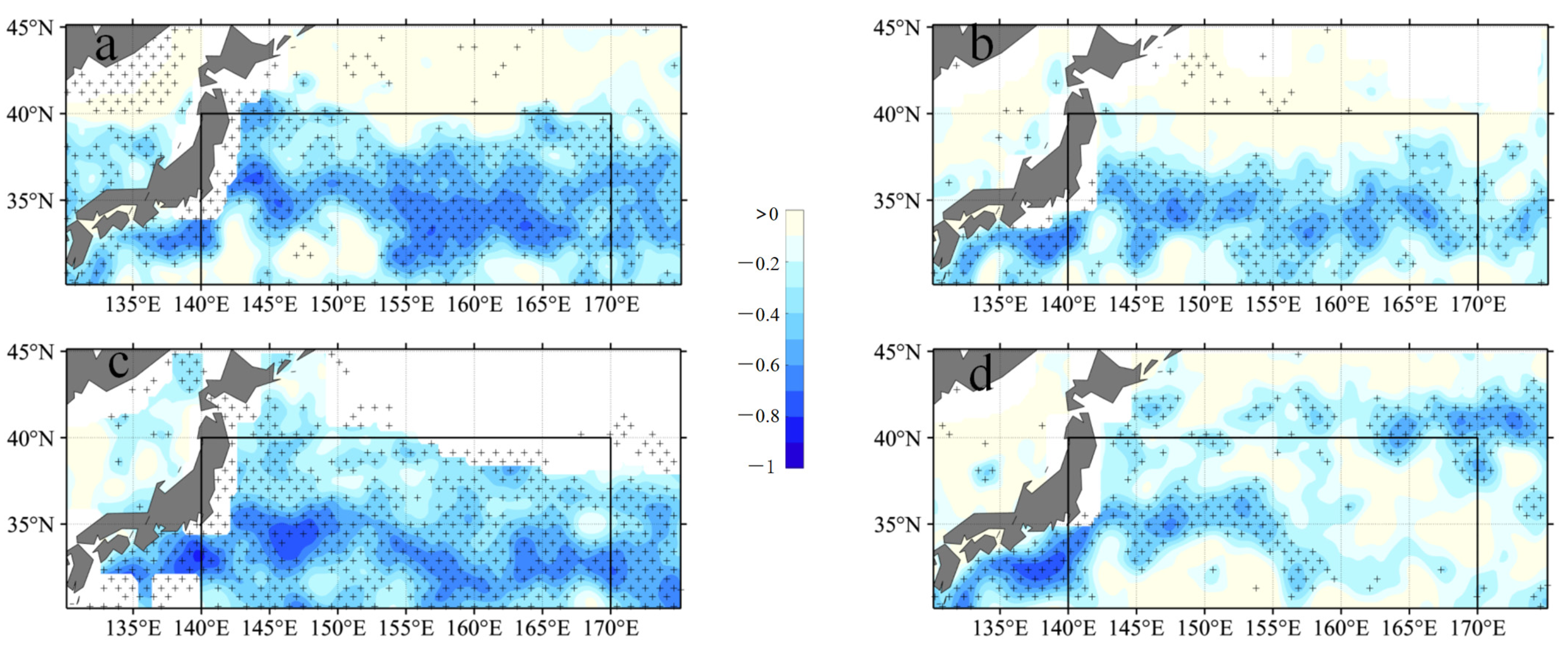
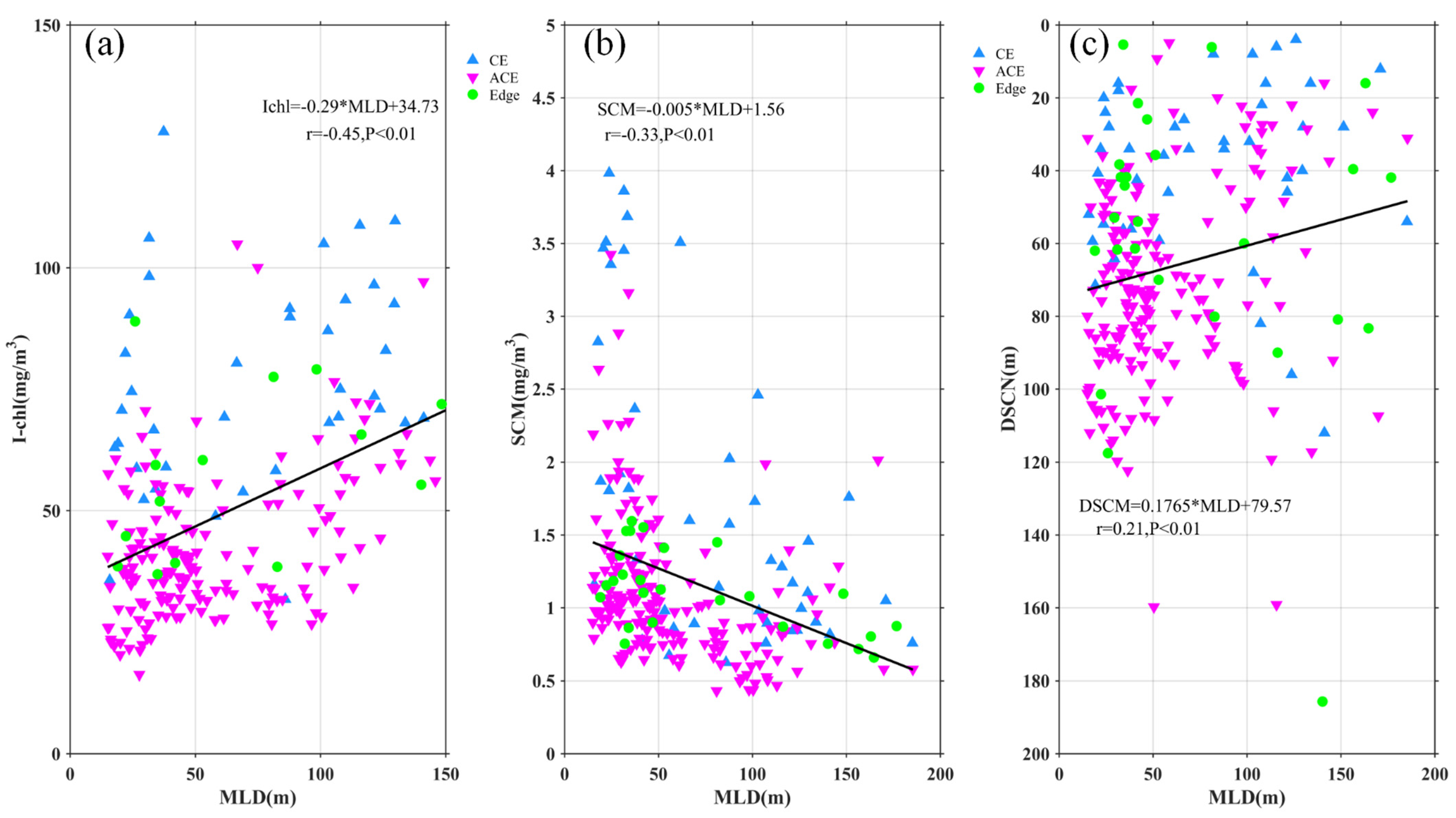
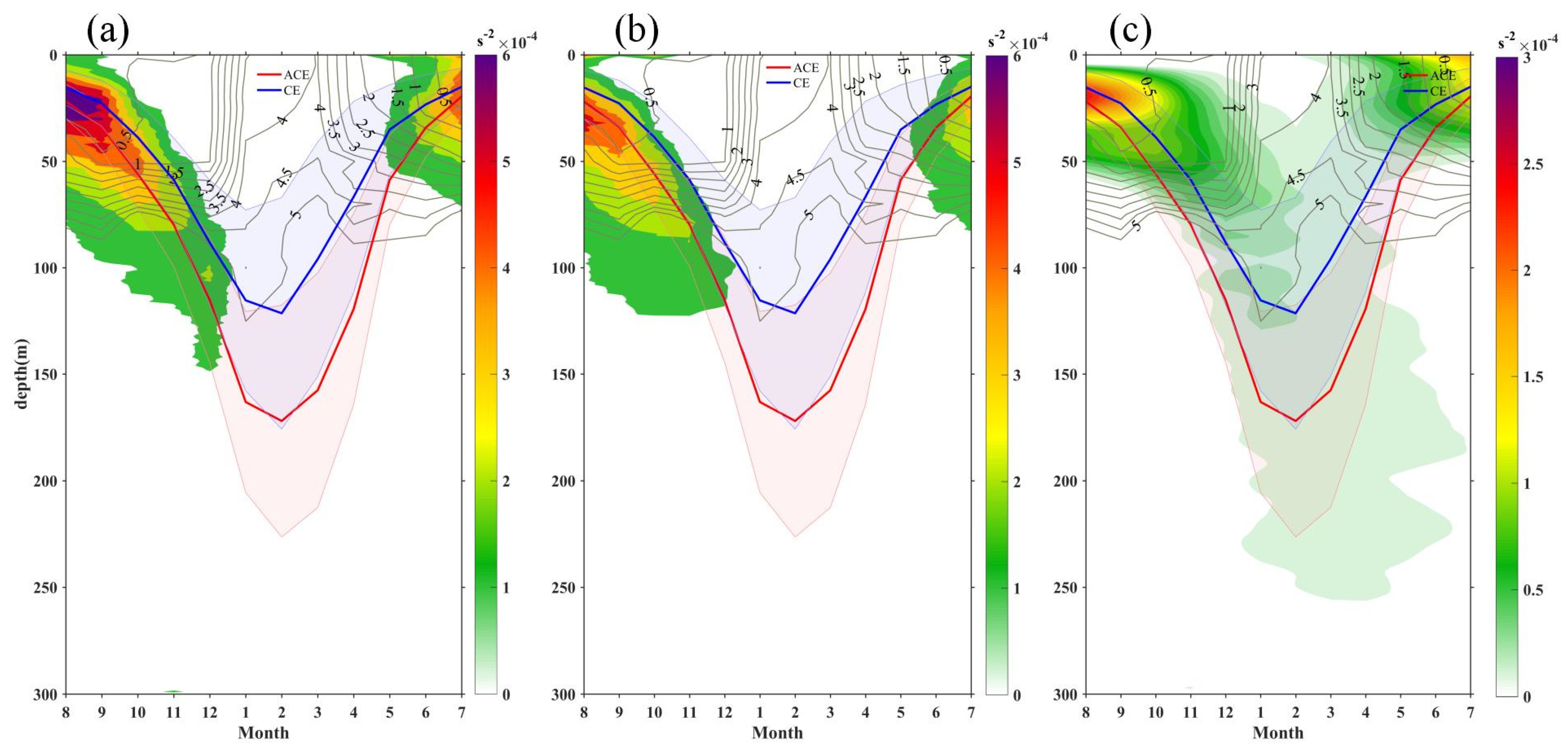
4.1. Modulation of Mixed Layer Depth by Eddies
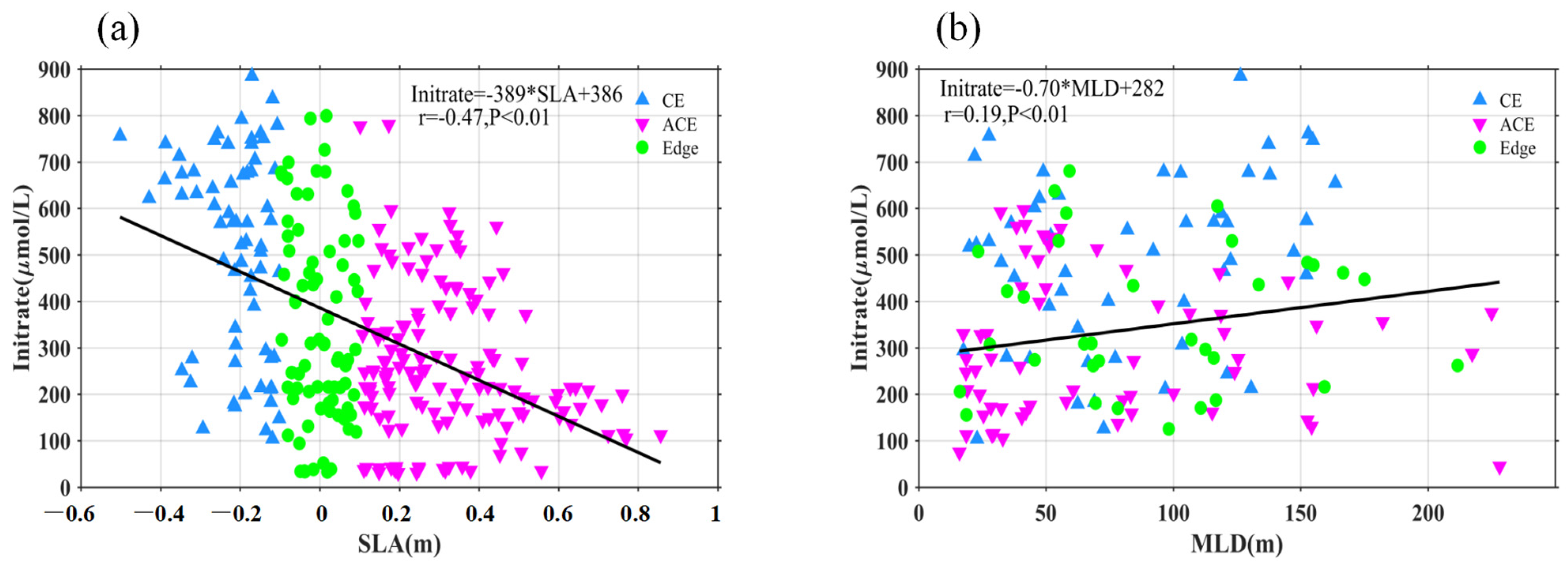
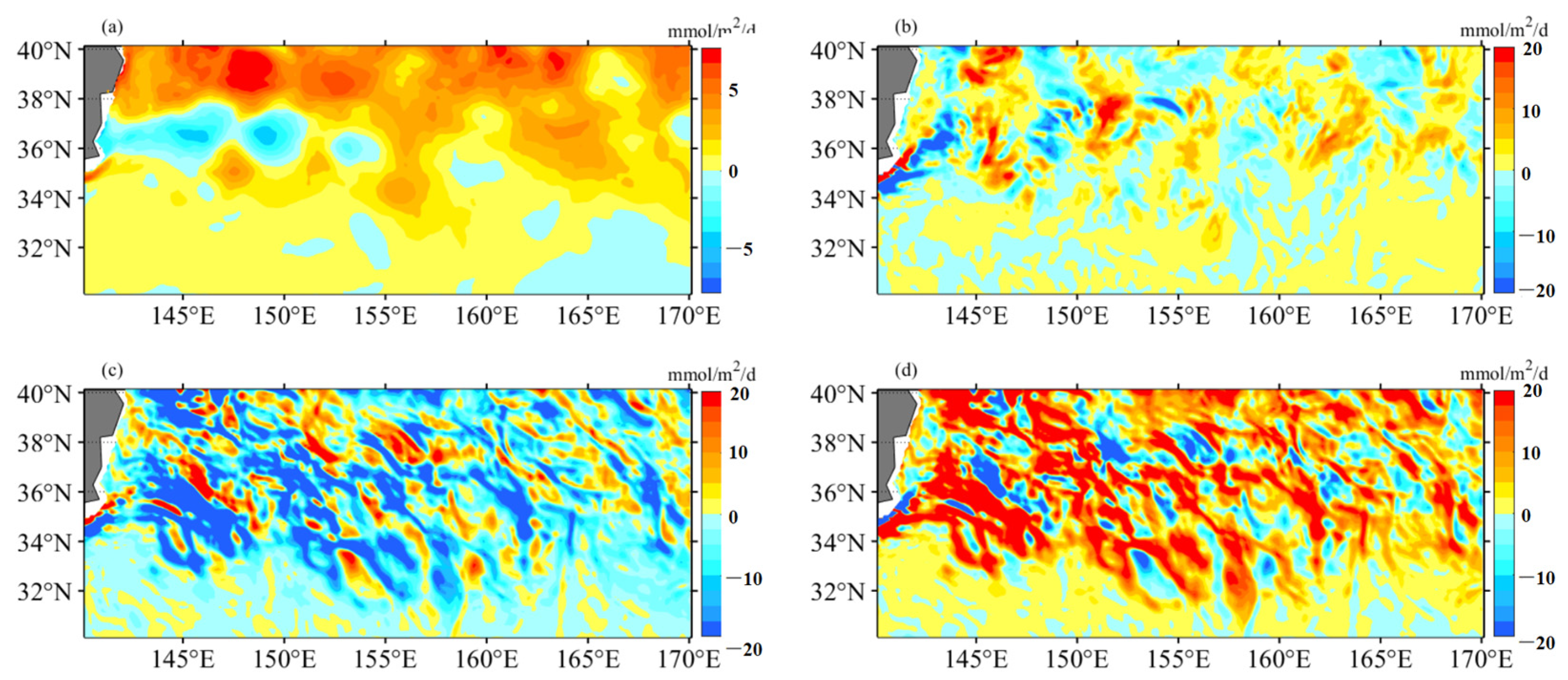
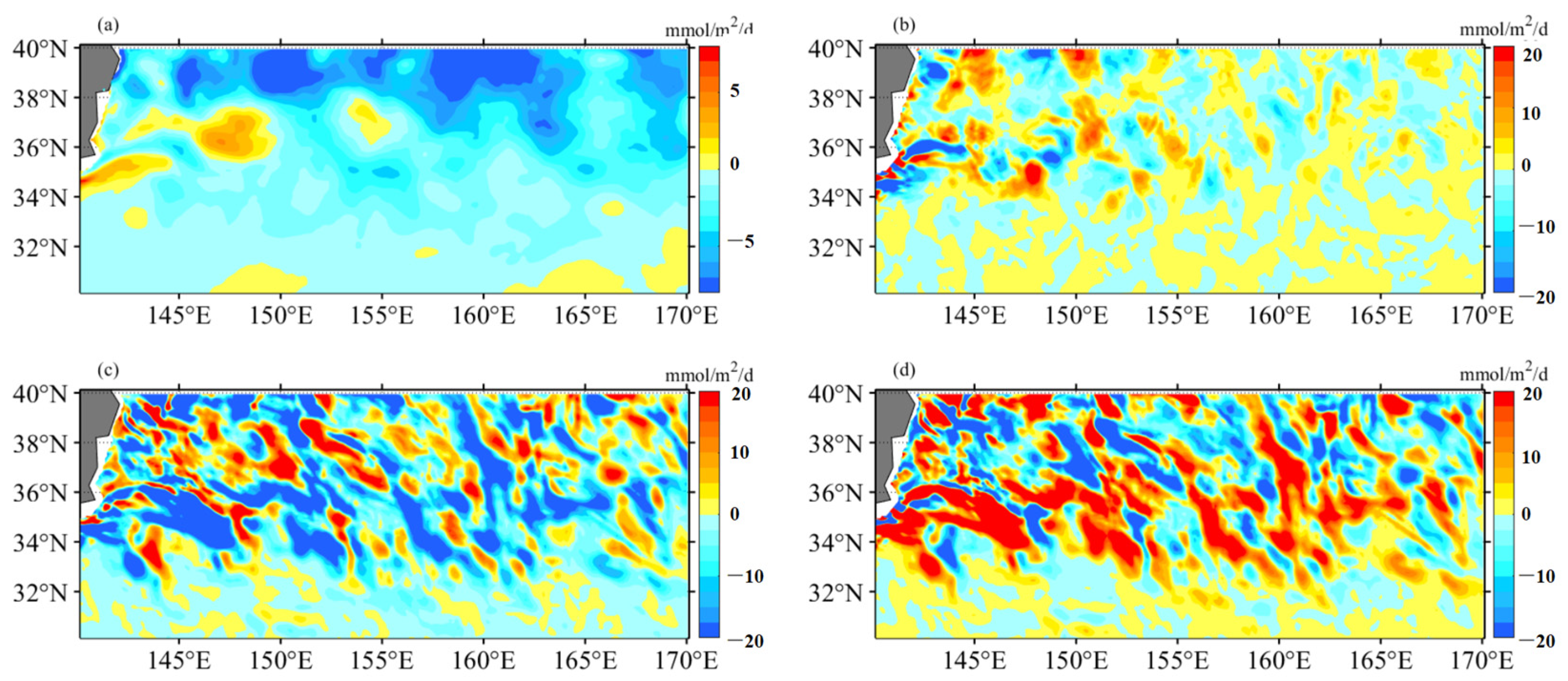
4.2. Mesoscale Influence on Nutrient Budget
4.3. Consideration of Submesoscale Processes
5. Conclusions
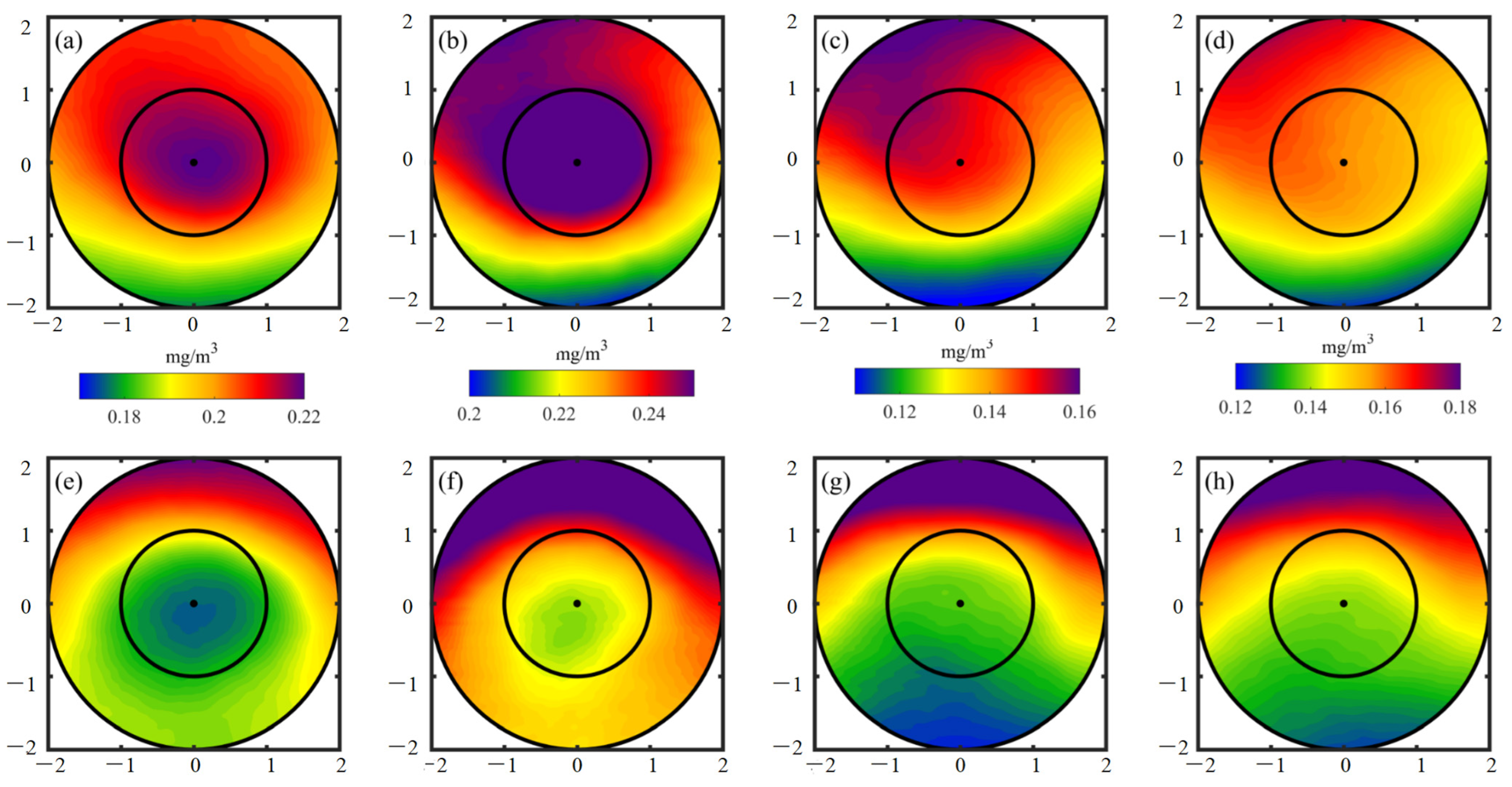
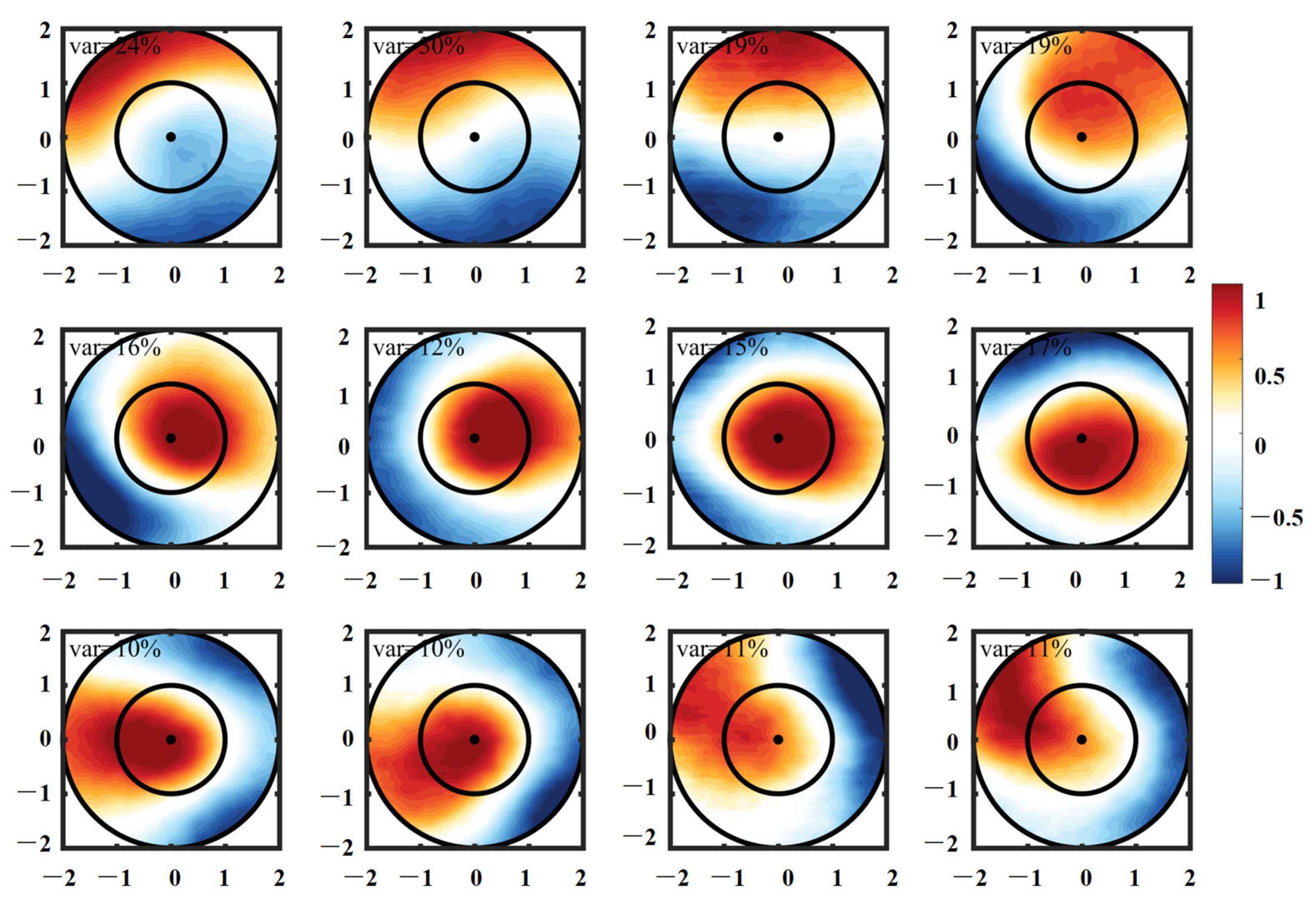
- CEs and ACEs exhibit opposite surface Chl-a anomalies, with CEs inducing positive anomalies and ACEs causing negative anomalies, particularly during winter. The monopole Chl-a patterns within the centers of the eddies correspond to positive or negative anomalies, depending on the sign of the principal component. These Chl-a anomalies account for approximately 26% and 18% of the total CEs and ACEs, respectively, across all seasons. These anomalies result from the uplifting or deepening of isopycnals and nitrate, stimulating or suppressing phytoplankton growth. Consequently, CEs and ACEs lead to variations in SCM depth-integrated Chl-a and nitrate, predominantly near the main axis of the KE.
- The vertical distribution of Chl-a within eddies exhibits distinct patterns. Above the SCM layer, Chl-a concentrations are higher within CEs and lower within ACEs compared to the edge values, irrespective of winter variations. Conversely, below the SCM layer, Chl-a concentrations are lower within CEs and higher within ACEs than the edge values. Nutrient supply resulting from stratification differences under convective mixing and eddy stirring may contribute to these anomalies.
- Additionally, another study examined the adjustment of MLD in eddies, revealing the influence of eddy-induced upwelling and downwelling in CEs and ACEs on nutrient supply and Chl-a concentrations. The differences between CEs and ACEs are more pronounced in winter due to deeper mixing, enhanced nutrient supply, and the redistribution of Chl-a. The shallow mixed layer and stratification during summer affect nutrient injection and contribute to variations in Chl-a concentrations. Convective mixing processes also play a role in nutrient increase or decrease during winter and summer, respectively.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chelton, D.B.; Schlax, M.G.; Samelson, R.M. Global observations of nonlinear mesoscale eddies. Prog. Oceanogr. 2011, 91, 167–216. [Google Scholar] [CrossRef]
- Qiu, B. Kuroshio Extension variability and forcing of the Pacific decadal oscillations: Responses and potential feedback. J. Phys. Oceanogr. 2003, 33, 2465–2482. [Google Scholar] [CrossRef]
- Qiu, B.; Chen, S. Variability of the Kuroshio Extension jet, recirculation gyre, and mesoscale eddies on decadal time scales. J. Phys. Oceanogr. 2005, 35, 2090–2103. [Google Scholar] [CrossRef]
- Siegel, D.A.; Peterson, P.; McGillicuddy, D.J., Jr.; Maritorena, S.; Nelson, N.B. Bio-optical footprints created by mesoscale eddies in the Sargasso Sea. Geophys. Res. Lett. 2011, 38, 47660. [Google Scholar] [CrossRef] [Green Version]
- Gaube, P.; Chelton, D.B.; Strutton, P.G.; Behrenfeld, M.J. Satellite observations of chlorophyll, phytoplankton biomass, and Ekman pumping in nonlinear mesoscale eddies. J. Geophys. Res. Ocean. 2013, 118, 6349–6370. [Google Scholar] [CrossRef] [Green Version]
- Jose, Y.S.; Aumont, O.; Machu, E.; Penven, P.; Moloney, C.L.; Maury, O. Influence of mesoscale eddies on biological production in the Mozambique Channel: Several contrasted examples from a coupled ocean-biogeochemistry model. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2014, 100, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Dufois, F.; Hardman-Mountford, N.J.; Fernandes, M.; Wojtasiewicz, B.; Shenoy, D.; Slawinski, D.; Toresen, R. Observational insights into chlorophyll distributions of subtropical South Indian Ocean eddies. Geophys. Res. Lett. 2017, 44, 3255–3264. [Google Scholar] [CrossRef] [Green Version]
- Dufois, F.; Hardman-Mountford, N.J.; Greenwood, J.; Richardson, A.J.; Feng, M.; Matear, R.J. Anticyclonic eddies are more productive than cyclonic eddies in subtropical gyres because of winter mixing. Sci. Adv. 2016, 2, e1600282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Du, Y.; Liao, X.; Xiang, C. Evidence of eddy-enhanced winter chlorophyll-a blooms in northern Arabian Sea: 2017 cruise expedition. J. Geophys. Res. Ocean. 2020, 125, e2019JC015582. [Google Scholar] [CrossRef]
- He, Q.; Zhan, H.; Xu, J.; Cai, S.; Zhan, W.; Zhou, L.; Zha, G. Eddy-induced chlorophyll anomalies in the western South China Sea. J. Geophys. Res. Ocean. 2019, 124, 9487–9506. [Google Scholar] [CrossRef]
- Kouketsu, S.; Kaneko, H.; Okunishi, T.; Sasaoka, K.; Itoh, S.; Inoue, R.; Ueno, H. Mesoscale eddy effects on temporal variability of surface chlorophyll a in the Kuroshio Extension. J. Oceanogr. 2016, 72, 439–451. [Google Scholar] [CrossRef]
- Huang, J.; Xu, F. Observational evidence of subsurface chlorophyll response to mesoscale eddies in the North Pacific. Geophys. Res. Lett. 2018, 45, 8462–8470. [Google Scholar] [CrossRef] [Green Version]
- Siswanto, E.; Sasai, Y.; Matsumoto, K.; Honda, M.C. Winter–Spring Phytoplankton Phenology Associated with the Kuroshio Extension Instability. Remote Sens. 2022, 14, 1186. [Google Scholar] [CrossRef]
- McGillicuddy, D.J., Jr.; Robinson, A.R. Eddy-induced nutrient supply and new production in the Sargasso Sea. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1997, 44, 1427–1450. [Google Scholar] [CrossRef]
- McGillicuddy, D.J., Jr.; Robinson, A.R.; Siegel, D.A.; Jannasch, H.W.; Johnson, R.; Dickey, T.D.; Knap, A.H. Influence of mesoscale eddies on new production in the Sargasso Sea. Nature 1998, 394, 263–266. [Google Scholar] [CrossRef]
- He, Q.; Zhan, H.; Cai, S.; Zha, G. On the asymmetry of eddy-induced surface chlorophyll anomalies in the southeastern Pacific: The role of eddy-Ekman pumping. Prog. Oceanogr. 2016, 141, 202–211. [Google Scholar] [CrossRef]
- Wang, T.; Chen, F.; Zhang, S.; Pan, J.; Devlin, A.T.; Ning, H.; Zeng, W. Remote Sensing and Argo Float Observations Reveal Physical Processes Initiating a Winter-Spring Phytoplankton Bloom South of the Kuroshio Current Near Shikoku. Remote Sens. 2020, 12, 4065. [Google Scholar] [CrossRef]
- Dufois, F.; Hardman-Mountford, N.J.; Greenwood, J.; Richardson, A.J.; Feng, M.; Herbette, S.; Matear, R. Impact of eddies on surface chlorophyll in the South Indian Ocean. J. Geophys. Res. Ocean. 2014, 119, 8061–8077. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Xu, F.; Zhou, K.; Xiu, P.; Lin, Y. Temporal evolution of near-surface chlorophyll over cyclonic eddy lifecycles in the southeastern Pacific. J. Geophys. Res. Ocean. 2017, 122, 6165–6179. [Google Scholar] [CrossRef]
- Sasai, Y.; Sasaoka, K.; Sasaki, H.; Ishida, A. Seasonal and intra-seasonal variability of chlorophyll-a in the North Pacific: Model and satellite data. J. Earth Simulator 2007, 8, 3–11. [Google Scholar] [CrossRef]
- Lin, P.; Ma, J.; Chai, F.; Xiu, P.; Liu, H. Decadal variability of nutrients and biomass in the southern region of Kuroshio Extension. Prog. Oceanogr. 2020, 188, 102441. [Google Scholar] [CrossRef]
- Rii, Y.M.; Brown, S.L.; Nencioli, F.; Kuwahara, V.; Dickey, T.; Karl, D.M.; Bidigare, R.R. The transient oasis: Nutrient-phytoplankton dynamics and particle export in Hawaiian lee cyclones. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 1275–1290. [Google Scholar] [CrossRef]
- Bidigare, R.R.; Benitez-Nelson, C.; Leonard, C.L.; Quay, P.D.; Parsons, M.L.; Foley, D.G.; Seki, M.P. Influence of a cyclonic eddy on microheterotroph biomass and carbon export in the lee of Hawaii. Geophys. Res. Lett. 2003, 30, 16393. [Google Scholar] [CrossRef]
- Maritorena, S.; d’Andon, O.H.F.; Mangin, A.; Siegel, D.A. Merged satellite ocean color data products using a bio-optical model: Characteristics, benefits and issues. Remote Sens. Environ. 2010, 114, 1791–1804. [Google Scholar] [CrossRef]
- Kaihatu, J.M.; Handler, R.A.; Marmorino, G.O.; Shay, L.K. Empirical orthogonal function analysis of ocean surface currents using complex and real-vector methods. J. Atmos. Ocean. Technol. 1998, 15, 927–941. [Google Scholar] [CrossRef]
- Lorenz, E.N. Empirical Orthogonal Functions and Statistical Weather Prediction; Department of Meteorology, Massachusetts Institute of Technology: Cambridge, UK, 1956; Volume 1, p. 52. [Google Scholar]
- Damerell, G.M.; Heywood, K.J.; Thompson, A.F.; Binetti, U.; Kaiser, J. The vertical structure of upper ocean variability at the Porcupine Abyssal Plain during 2012–2013. J. Geophys. Res. Ocean. 2016, 121, 3075–3089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caniaux, G.; Planton, S. A three-dimensional ocean mesoscale simulation using data from the SEMAPHORE experiment: Mixed layer heat budget. J. Geophys. Res. Ocean. 1998, 103, 25081–25099. [Google Scholar] [CrossRef]
- Radenac, M.H.; Jouanno, J.; Tchamabi, C.C.; Awo, M.; Bourlès, B.; Arnault, S.; Aumont, O. Physical drivers of the nitrate seasonal variability in the Atlantic cold tongue. Biogeosciences 2020, 17, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Resplandy, L.; Lévy, M.; Madec, G.; Pous, S.; Aumont, O.; Kumar, D. Contribution of mesoscale processes to nutrient budgets in the Arabian Sea. J. Geophys. Res. Ocean. 2011, 116, 7006. [Google Scholar] [CrossRef] [Green Version]
- Flierl, G.R. Particle motions in large-amplitude wave fields. Geophys. Astrophys. Fluid Dyn. 1981, 18, 39–74. [Google Scholar] [CrossRef]
- Itoh, S.; Yasuda, I. Characteristics of mesoscale eddies in the Kuroshio–Oyashio Extension region detected from the distribution of the sea surface height anomaly. J. Phys. Oceanogr. 2010, 40, 1018–1034. [Google Scholar] [CrossRef]
- Karl, D.M.; Church, M.J. Microbial oceanography and the Hawaii Ocean Time-series programme. Nat. Rev. Microbiol. 2014, 12, 699–713. [Google Scholar] [CrossRef]
- Karl, D.M.; Church, M.J. Ecosystem structure and dynamics in the North Pacifific subtropical gyre: New views of an old ocean. Ecosystems 2017, 20, 433–457. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Long, M.C.; Gaube, P.; Frenger, I.; Marshall, J.; McGillicuddy, D.J. Seasonal variation in the correlation between anomalies of sea level and chlorophyll in the Antarctic Circumpolar Current. Geophys. Res. Lett. 2018, 45, 5011–5019. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Wang, Z.; Zhang, K. Controlling effects of mesoscale eddies on thermohaline structure and in situ chlorophyll distribution in the western North Pacifific. J. Mar. Syst. 2017, 175, 24–35. [Google Scholar] [CrossRef]
- Xiu, P.; Chai, F. Eddies affect subsurface phytoplankton and oxygen distributions in the North Pacific Subtropical Gyre. Geophys. Res. Lett. 2020, 47, e2020GL087037. [Google Scholar] [CrossRef]
- Oschlies, A.; Garçon, V. Eddy-induced enhancement of primary production in a model of the North Atlantic Ocean. Nature 1998, 394, 266–269. [Google Scholar] [CrossRef]
- Chelton, D.B.; Gaube, P.; Schlax, M.G.; Early, J.J.; Samelson, R.M. The influence of nonlinear mesoscale eddies on near-surface oceanic chlorophyll. Science 2011, 334, 328–332. [Google Scholar] [CrossRef]
- Dong, J.; Fox-Kemper, B.; Zhang, H.; Dong, C. The seasonality of submesoscale energy production, content, and cascade. Geophys. Res. Lett. 2020, 47, e2020GL087388. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, S.; Claret, M.; Pascual, A.; Olita, A.; Troupin, C.; Capet, A.; Mahadevan, A. Effects of oceanic mesoscale and submesoscale frontal processes on the vertical transport of phytoplankton. J. Geophys. Res. Ocean. 2019, 124, 5999–6014. [Google Scholar] [CrossRef] [Green Version]
- Uchida, T.; Balwada, D.; Abernathey, R.P.; McKinley, G.A.; Smith, S.K.; Lévy, M. Vertical eddy iron fluxes support primary production in the open Southern Ocean. Nat. Commun. 2020, 11, 1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévy, M.; Ferrari, R.; Franks, P.J.; Martin, A.P.; Rivière, P. Bringing physics to life at the submesoscale. Geophys. Res. Lett. 2012, 39, e2012GL052756. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Qiu, B. Surface chlorophyll enhancement in mesoscale eddies by submesoscale spiral bands. Geophys. Res. Lett. 2020, 47, e2020GL088820. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CE | ACE | Edge | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winter | Spring | Summer | Autumn | All | Winter | Spring | Summer | Autumn | All | Winter | Spring | Summer | Autumn | All | |
| Chl-a | 11 | 11 | 9 | 3 | 34 | 29 | 10 | 65 | 88 | 192 | 10 | 8 | 4 | 13 | 35 |
| Nitrate | 15 | 32 | 16 | 4 | 67 | 36 | 24 | 44 | 43 | 147 | 29 | 28 | 7 | 16 | 80 |
| Temperature and Salinity | 907 | 1377 | 922 | 474 | 3680 | 1447 | 1069 | 2008 | 2623 | 7147 | 2364 | 2808 | 1818 | 2056 | 9046 |
| CE | ACE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Winter | Spring | Summer | Autumn | All | Winter | Spring | Summer | Autumn | All | |
| Number | 574 | 630 | 594 | 547 | 1223 | 556 | 619 | 555 | 526 | 1180 |
| Amplitude (cm) | 14.04 | 13.71 | 14.55 | 15.2 | 14.38 | 13.12 | 13.1 | 13.77 | 13.66 | 13.41 |
| Radius (km) | 79.93 | 78.27 | 78.42 | 80.74 | 79.34 | 84.42 | 82.67 | 85.21 | 86.36 | 84.67 |
| Rotational Speed (m/s) | 0.36 | 0.37 | 0.39 | 0.39 | 0.38 | 0.32 | 0.33 | 0.33 | 0.33 | 0.33 |
| Chl-a anomaly (mg/m3) | 0.48 | 0.41 | 0.31 | 0.11 | 0.33 | −0.5 | −0.37 | −0.36 | −0.29 | −0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Zhang, S.; Chen, F.; Xiao, L. The Seasonality of Eddy-Induced Chlorophyll-a Anomalies in the Kuroshio Extension System. Remote Sens. 2023, 15, 3865. https://doi.org/10.3390/rs15153865
Wang T, Zhang S, Chen F, Xiao L. The Seasonality of Eddy-Induced Chlorophyll-a Anomalies in the Kuroshio Extension System. Remote Sensing. 2023; 15(15):3865. https://doi.org/10.3390/rs15153865
Chicago/Turabian StyleWang, Tongyu, Shuwen Zhang, Fajin Chen, and Luxing Xiao. 2023. "The Seasonality of Eddy-Induced Chlorophyll-a Anomalies in the Kuroshio Extension System" Remote Sensing 15, no. 15: 3865. https://doi.org/10.3390/rs15153865