Quantifying Plant Species α-Diversity Using Normalized Difference Vegetation Index and Climate Data in Alpine Grasslands


Abstract
:1. Introduction
2. Materials and Methods
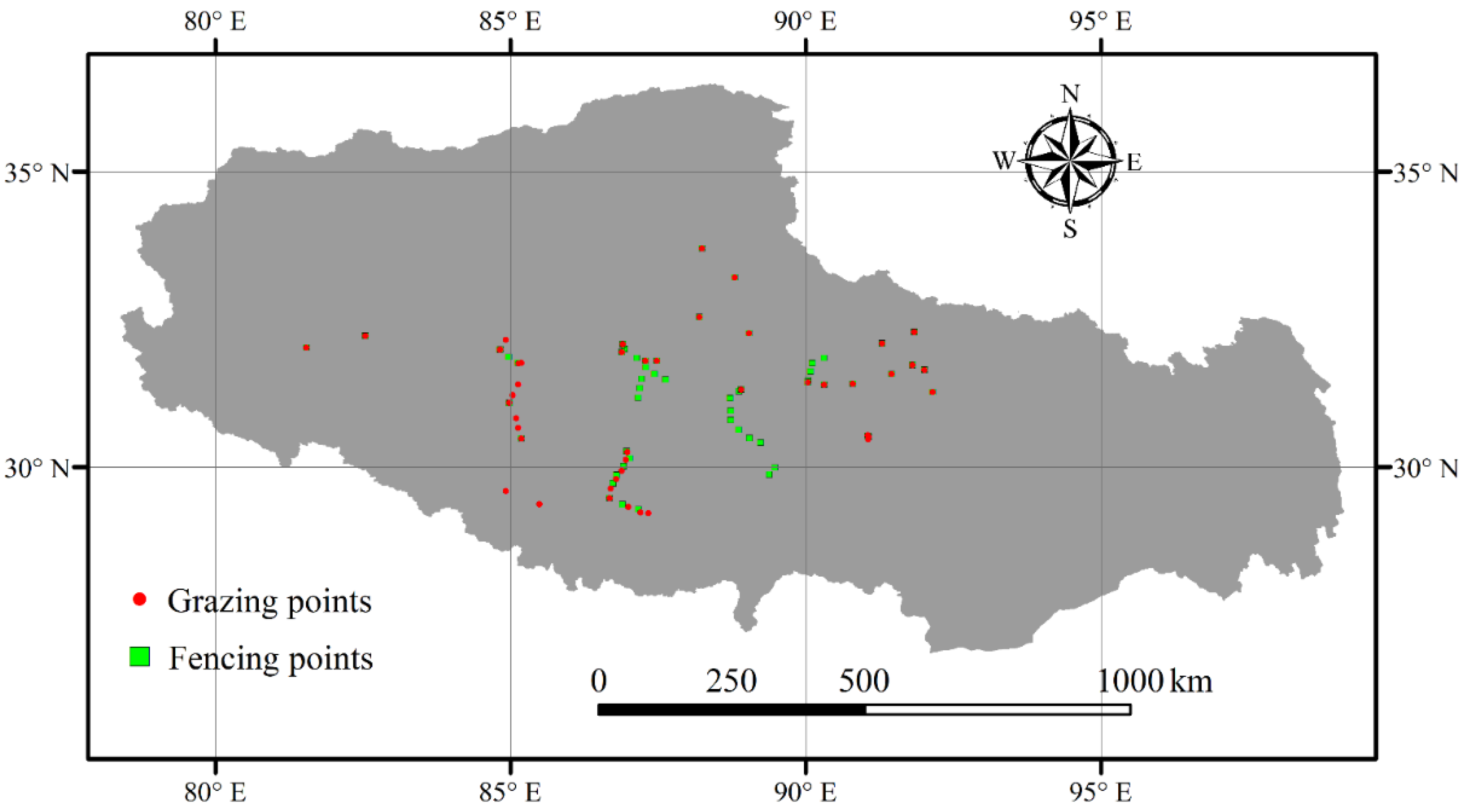
2.1. Data
2.2. Statistical Analysis
3. Results
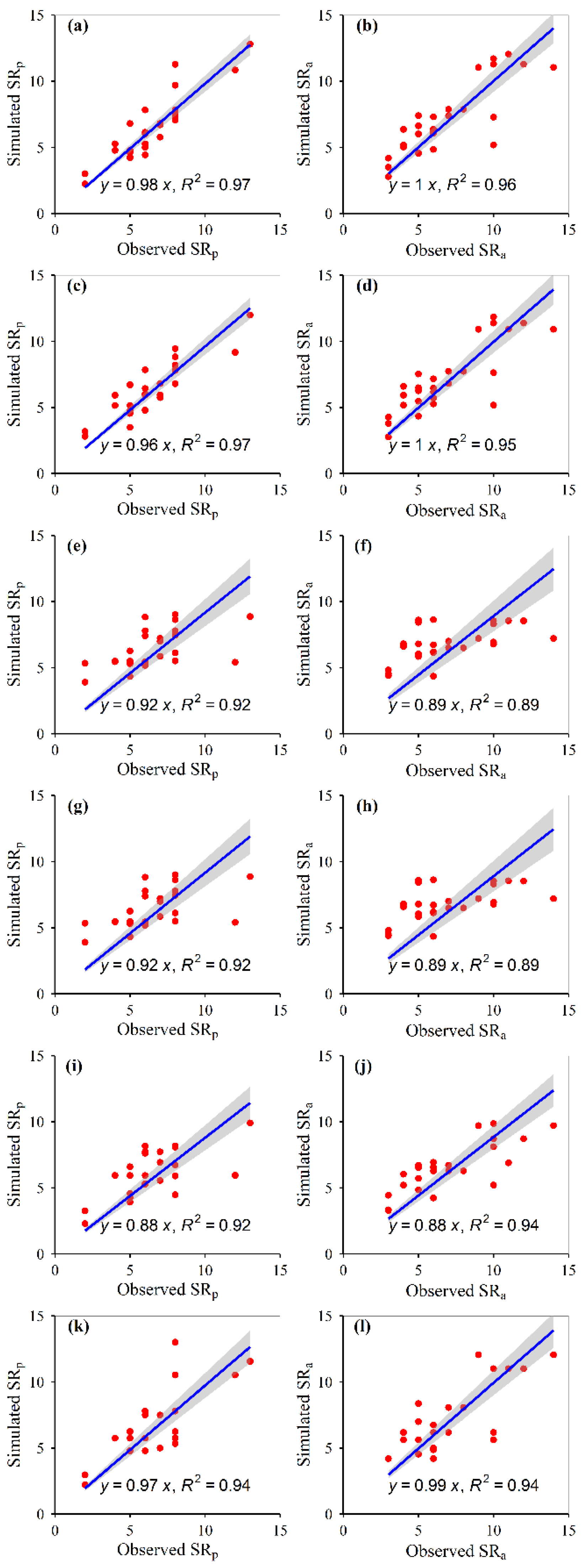
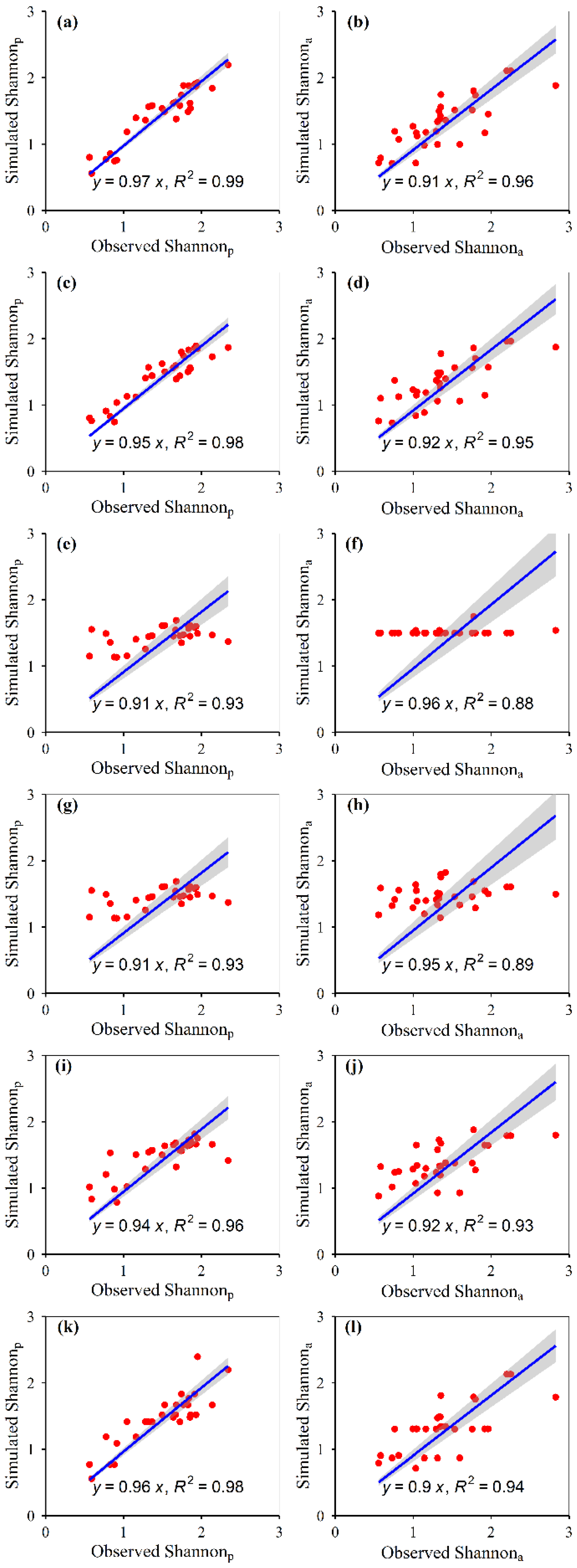
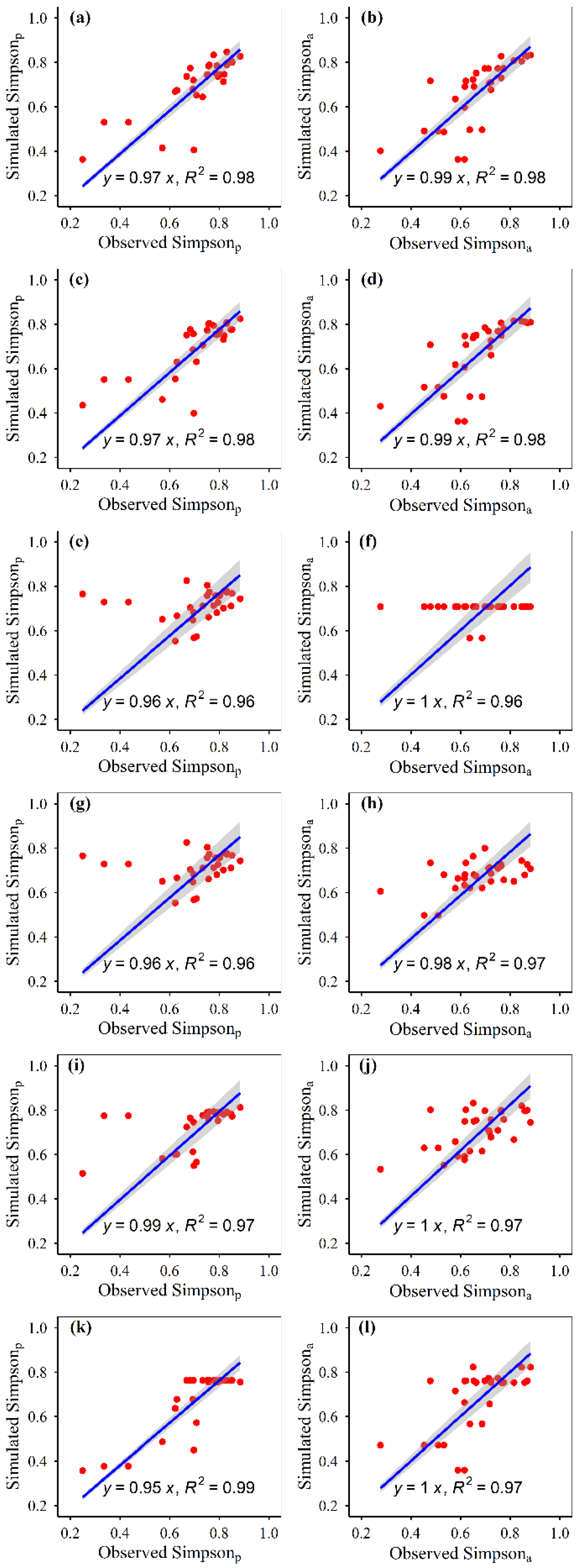
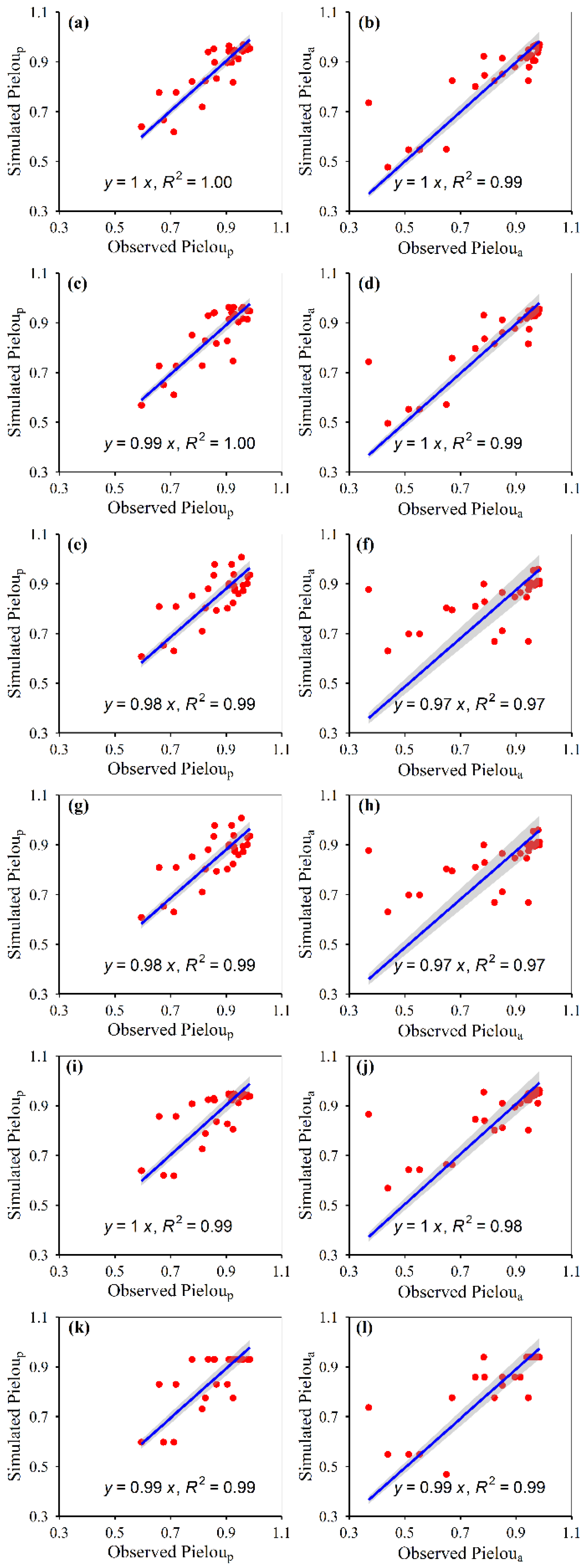
3.1. Model Construction
3.2. Model Accuracies
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berendse, F.; Van Ruijven, J.; Jongejans, E.; Keesstra, S. Loss of Plant Species Diversity Reduces Soil Erosion Resistance. Ecosystems 2015, 18, 881–888. [Google Scholar] [CrossRef] [Green Version]
- Brummitt, N.; Araujo, A.C.; Harris, T. Areas of plant diversity-What do we know? Plants People Planet 2021, 3, 33–44. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Zak, D.R.; Blackwood, C.B.; Curtis, C.D.; Tilman, D. Resource availability controls fungal diversity across a plant diversity gradient. Ecol. Lett. 2006, 9, 1127–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welti, E.; Helzer, C.; Joern, A. Impacts of plant diversity on arthropod communities and plant-herbivore network architecture. Ecosphere 2017, 8, e01983. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.A.; Harte, J.; Zhao, X.Q. Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecol. Lett. 2004, 7, 1170–1179. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Lemmens, C.; Gielen, B.; Bossuyt, H.; Malchair, S.; Carnol, M.; Merckx, R.; Nijs, I. Combined effects of climate warming and plant diversity loss on above- and below-ground grassland productivity. Environ. Exp. Bot. 2007, 60, 95–104. [Google Scholar] [CrossRef]
- Scheiner, S.M.; Reybenayas, J.M. Global patterns of plant diversity. Evol. Ecol. 1994, 8, 331–347. [Google Scholar] [CrossRef]
- Zha, X.J.; Tian, Y.; Ou, Z.; Fu, G. Response of forage nutrient storages to grazing in alpine grasslands. Front. Plant Sci. 2022, 3505. [Google Scholar] [CrossRef]
- Zhang, H.; Fu, G. Responses of plant, soil bacterial and fungal communities to grazing vary with pasture seasons and grassland types, northern Tibet. Land Degrad. Dev. 2021, 32, 1821–1832. [Google Scholar] [CrossRef]
- Wang, J.W.; Yu, C.Q.; Fu, G. Asymmetrical warming between elevations may result in similar plant community composition between elevations in alpine grasslands. Front. Ecol. Evol. 2021, 702. [Google Scholar] [CrossRef]
- Wang, J.W.; Yu, C.Q.; Fu, G. Warming reconstructs the elevation distributions of aboveground net primary production, plant species and phylogenetic diversity in alpine grasslands. Ecol. Indic. 2021, 133, 108355. [Google Scholar] [CrossRef]
- Sun, W.; Li, S.W.; Wang, J.H.; Fu, G. Effects of grazing on plant species and phylogenetic diversity in alpine grasslands, Northern Tibet. Ecol. Eng. 2021, 170, 106331. [Google Scholar] [CrossRef]
- Bastazini, V.A.G.; Galiana, N.; Hillebrand, H.; Estiarte, M.; Ogaya, R.; Peñuelas, J.; Sommer, U.; Montoya, J.M. The impact of climate warming on species diversity across scales: Lessons from experimental meta-ecosystems. Glob. Ecol. Biogeogr. 2021, 30, 1545–1554. [Google Scholar] [CrossRef]
- Bongaarts, J. Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Popul. Dev. Rev. 2019, 45, 680–681. [Google Scholar] [CrossRef] [Green Version]
- Elgersma, A.; Søegaard, K. Effects of species diversity on seasonal variation in herbage yield and nutritive value of seven binary grass-legume mixtures and pure grass under cutting. Eur. J. Agron. 2016, 78, 73–83. [Google Scholar] [CrossRef]
- Prieto, P.; Penuelas, J.; Lloret, F.; Llorens, L.; Estiarte, M. Experimental drought and warming decrease diversity and slow down post-fire succession in a Mediterranean shrubland. Ecography 2009, 32, 623–636. [Google Scholar] [CrossRef]
- Fu, G.; Sun, W. Temperature sensitivities of vegetation indices and aboveground biomass are primarily linked with warming magnitude in high-cold grasslands. Sci. Total Environ. 2022, 843, 157002. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X. Asymmetrical warming of growing/non-growing season increases soil respiration during growing season in an alpine meadow. Sci. Total Environ. 2022, 812, 152591. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X.; Zhang, X.Z. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau. Agricult. For. Meteorol. 2018, 249, 11–21. [Google Scholar] [CrossRef]
- Han, F.S.; Fu, G.; Yu, C.Q.; Wang, S.H. Modeling nutrition quality and storage of forage using climate data and normalized-difference vegetation index in alpine grasslands. Remote Sens. 2022, 14, 3410. [Google Scholar] [CrossRef]
- Mantas, C.J.; Castellano, J.G.; Moral-García, S.; Abellán, J. A comparison of random forest based algorithms: Random credal random forest versus oblique random forest. Soft Comput. 2019, 23, 10739–10754. [Google Scholar] [CrossRef]
- Mishina, Y.; Murata, R.; Yamauchi, Y.; Yamashita, T.; Fujiyoshi, H. Boosted Random Forest. IEICE Trans. Inf. Syst. 2015, 98, 1630–1636. [Google Scholar] [CrossRef] [Green Version]
- Mishina, Y.; Tsuchiya, M.; Fujiyoshi, H. Boosted Random Forest. In Proceedings of the 9th International Conference on Computer Vision Theory and Applications (VISAPP), Lisbon, Portugal, 5–8 January 2014. [Google Scholar]
- Paul, A.; Mukherjee, D.P. Reinforced Random Forest. In Proceedings of the 10th Indian Conference on Computer Vision, Graphics and Image Processing (ICVGIP), Guwahati, India, 18–22 December 2016; Indian Institute of Technology: Delhi, India, 2016. [Google Scholar]
- Sun, J.Y.; Zhong, G.Q.; Huang, K.Z.; Dong, J. Banzhaf random forests: Cooperative game theory based random forests with consistency. Neural Netw. 2018, 106, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.M.; Li, C.H.; Shi, H.; Zou, Q. Material Classification Using Random Forest. In Proceedings of the 2nd International Conference on Advanced Measurement and Test (AMT 2011), Nanchang, China, 24–26 June 2011; Peoples Republic of China: Beijing, China, 2011. [Google Scholar]
- Fu, G.; Wang, J.H.; Li, S.W. Response of forage nutritional quality to climate change and human activities in alpine grasslands. Sci. Total Environ. 2022, 845, 157552. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.S.; Shen, Z.X.; Shi, P.L.; Zhou, Y.T.; Zhang, Z.X. Effects of grazing exclusion on plant functional group diversity alpine grasslands along a precipitation gradient on the Northern Tibetan Plateau. Arct. Antarct. Alp. Res. 2014, 46, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.S.; Zhang, X.Z.; Shen, Z.X.; Shi, P.L.; Yu, C.Q.; Song, M.H.; Li, X.J. Species richness and diversity of alpine grasslands on the Northern Tibetan Plateau:effects of grazing exclusion and growing season precipitation. J. Resour. Ecol. 2012, 3, 236–242. [Google Scholar]
- Fu, G.; Wang, J.H.; Li, S.W.; He, P. Responses of forage nutrient quality to grazing in the alpine grassland of Northern Tibet. Acta Pratacult. Sin. 2021, 30, 38–50. [Google Scholar]
- Fu, G.; Shen, Z.X.; Sun, W.; Zhong, Z.M.; Zhang, X.Z.; Zhou, Y.T. A meta-analysis of the effects of experimental warming on plant physiology and growth on the Tibetan Plateau. J. Plant Growth Regul. 2015, 34, 57–65. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X. Effects of enhanced UV-B radiation on plant physiology and growth on the Tibetan Plateau: A meta-analysis. Acta Physiol. Plant. 2017, 39, 85. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X. Response of alpine plants to nitrogen addition on the Tibetan Plateau: A meta-analysis. J. Plant Growth Regul. 2016, 35, 974–979. [Google Scholar] [CrossRef]
- Zhong, Z.M.; Fu, G. Response of soil fungal species, phylogenetic and functional diversity to diurnal asymmetric warming in an alpine agricultural ecosystem. Agric. Ecosyst. Environ. 2022, 335, 107993. [Google Scholar] [CrossRef]
- Yu, C.Q.; Han, F.S.; Fu, G. Effects of 7 years experimental warming on soil bacterial and fungal community structure in the Northern Tibet alpine meadow at three elevations. Sci. Total Environ. 2019, 655, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Sun, W.; Li, S.W.; Zhang, J.; Yu, C.Q.; Shen, Z.X. Modeling aboveground biomass using MODIS images and climatic data in grasslands on the Tibetan Plateau. J. Resour. Ecol. 2017, 8, 42–49. [Google Scholar]
- Fu, G.; Shen, Z.X.; Zhang, X.Z.; Shi, P.L.; Zhang, Y.J.; Wu, J.S. Estimating air temperature of an alpine meadow on the Northern Tibetan Plateau using MODIS land surface temperature. Acta Ecol. Sin. 2011, 31, 8–13. [Google Scholar] [CrossRef]
- Wu, J.S.; Fu, G. Modelling aboveground biomass using MODIS FPAR/LAI data in alpine grasslands of the Northern Tibetan Plateau. Remote Sens. Lett. 2018, 9, 150–159. [Google Scholar] [CrossRef]
- Fu, G.; Wu, J.S. Validation of MODIS Collection 6 FPAR/LAI in the alpine grassland of the Northern Tibetan Plateau. Remote Sens. Lett. 2017, 8, 831–838. [Google Scholar] [CrossRef]
- Cortez, P. Data Mining with Neural Networks and Support Vector Machines Using the R/rminer Tool. In Proceedings of the 10th Industrial Conference on Data Mining, Berlin, Germany, 12–14 July 2010. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Freund, Y.; Schapire, R.E. A decision-theoretic generalization of on-line learning and an application to boosting. J. Comput. Syst. Sci. 1997, 55, 119–139. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity | Scenes | Random Forest | Generalized Boosted Regression | Artificial Neural Network | Multiple Linear Regression | Support Vector Machines | Recursive Regression Trees | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | Mean Square Errors | ntree | mtry | Trees | Mean train Error | Mean cv Error | Error | Size | Intercept | Temperature | Precipitation | Radiation | NDVI | R2 | Mean residuals | Mean Decision Values | gamma | rho | Support Vector Nos | R2 | ||
| SR | Potential | 0.73 | 1.94 | 134 | 3 | 987 | 2.70 | 3.49 | 286.74 | 0 | 5.26 | 0.00 | 0.01 | 0.00 | 0.25 | 0.10 | −0.04 | 0.33 | 0 | 441 | 0.57 | |
| Actual | 0.62 | 3.03 | 124 | 4 | 953 | 3.21 | 4.57 | 263.34 | 0 | −2.96 | 0.23 | 0.01 | 0.00 | 0.00 | 0.19 | 0.23 | −0.08 | 0.25 | 0 | 335 | 0.43 | |
| Shannon | Potential | 0.72 | 0.06 | 117 | 2 | 993 | 0.09 | 0.11 | 50.37 | 0 | −1.45 | 0.04 | 0.00 | 0.00 | 0.09 | −0.03 | 0.07 | 0.33 | 1 | 432 | 0.45 | |
| Actual | 0.61 | 0.09 | 118 | 1 | 969 | 0.10 | 0.12 | 43.30 | 8 | −0.99 | 0.06 | 0.00 | 0.00 | 0.00 | 0.10 | −0.01 | 0.03 | 0.25 | 1 | 323 | 0.53 | |
| Simpson | Potential | 0.72 | 0.01 | 196 | 1 | 991 | 0.01 | 0.02 | 17.96 | 0 | −0.82 | 0.02 | 0.00 | 0.00 | 0.17 | −0.03 | 0.18 | 0.33 | 1 | 420 | 0.45 | |
| Actual | 0.62 | 0.01 | 163 | 3 | 942 | 0.01 | 0.01 | 13.58 | 8 | −0.41 | 0.02 | 0.00 | 0.00 | 0.00 | 0.13 | −0.02 | 0.14 | 0.25 | 0 | 311 | 0.52 | |
| Pielou | Potential | 0.71 | 0.01 | 448 | 1 | 969 | 0.01 | 0.01 | 13.10 | 0 | −0.82 | 0.02 | 0.00 | 0.00 | 0.37 | −0.02 | 0.13 | 0.33 | 1 | 378 | 0.67 | |
| Actual | 0.73 | 0.01 | 210 | 3 | 912 | 0.01 | 0.01 | 12.35 | 0 | −0.38 | 0.02 | 0.00 | 0.00 | 0.00 | 0.39 | −0.02 | 0.10 | 0.25 | 0 | 287 | 0.70 | |
| Models | Potential α-Diversity | Actual α-Diversity | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Species Richness | Shannon | Simpson | Pielou | Species Richness | Shannon | Simpson | Pielou | ||
| Relative bias | Random forest | −1.00 | −1.09 | −1.81 | 0.70 | 4.39 | −4.49 | −0.59 | 1.17 |
| Generalized boosted regression | −1.40 | −2.80 | −1.40 | −0.90 | 4.61 | −2.54 | −0.15 | 0.94 | |
| Artificial neural network | −1.23 | −1.07 | −0.03 | −1.71 | 0.49 | 9.14 | 4.37 | 0.09 | |
| Multiple linear regression | −1.23 | −1.07 | −0.03 | −1.71 | 0.49 | 6.53 | 0.88 | 0.09 | |
| Support vector machines | −6.53 | −0.19 | 2.01 | 0.79 | −3.92 | −0.28 | 5.69 | 2.88 | |
| Recursive regression trees | 0.01 | −1.32 | −4.02 | −0.47 | 4.85 | −4.13 | 1.55 | 0.30 | |
| RMSE | Random forest | 1.10 | 0.17 | 0.09 | 0.05 | 1.58 | 0.32 | 0.10 | 0.09 |
| Generalized boosted regression | 1.14 | 0.20 | 0.09 | 0.06 | 1.60 | 0.34 | 0.11 | 0.09 | |
| Artificial neural network | 1.93 | 0.41 | 0.15 | 0.07 | 2.37 | 0.52 | 0.14 | 0.14 | |
| Multiple linear regression | 1.93 | 0.41 | 0.15 | 0.07 | 2.37 | 0.50 | 0.12 | 0.14 | |
| Support vector machines | 1.89 | 0.31 | 0.13 | 0.07 | 1.88 | 0.40 | 0.12 | 0.11 | |
| Recursive regression trees | 1.70 | 0.23 | 0.08 | 0.07 | 1.80 | 0.37 | 0.12 | 0.10 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Fu, G. Quantifying Plant Species α-Diversity Using Normalized Difference Vegetation Index and Climate Data in Alpine Grasslands. Remote Sens. 2022, 14, 5007. https://doi.org/10.3390/rs14195007
Tian Y, Fu G. Quantifying Plant Species α-Diversity Using Normalized Difference Vegetation Index and Climate Data in Alpine Grasslands. Remote Sensing. 2022; 14(19):5007. https://doi.org/10.3390/rs14195007
Chicago/Turabian StyleTian, Yuan, and Gang Fu. 2022. "Quantifying Plant Species α-Diversity Using Normalized Difference Vegetation Index and Climate Data in Alpine Grasslands" Remote Sensing 14, no. 19: 5007. https://doi.org/10.3390/rs14195007