Spatio-Temporal Variability of the Habitat Suitability Index for the Todarodes pacificus (Japanese Common Squid) around South Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fishery Data
2.2. Satellite Dataset
2.3. HSI Model
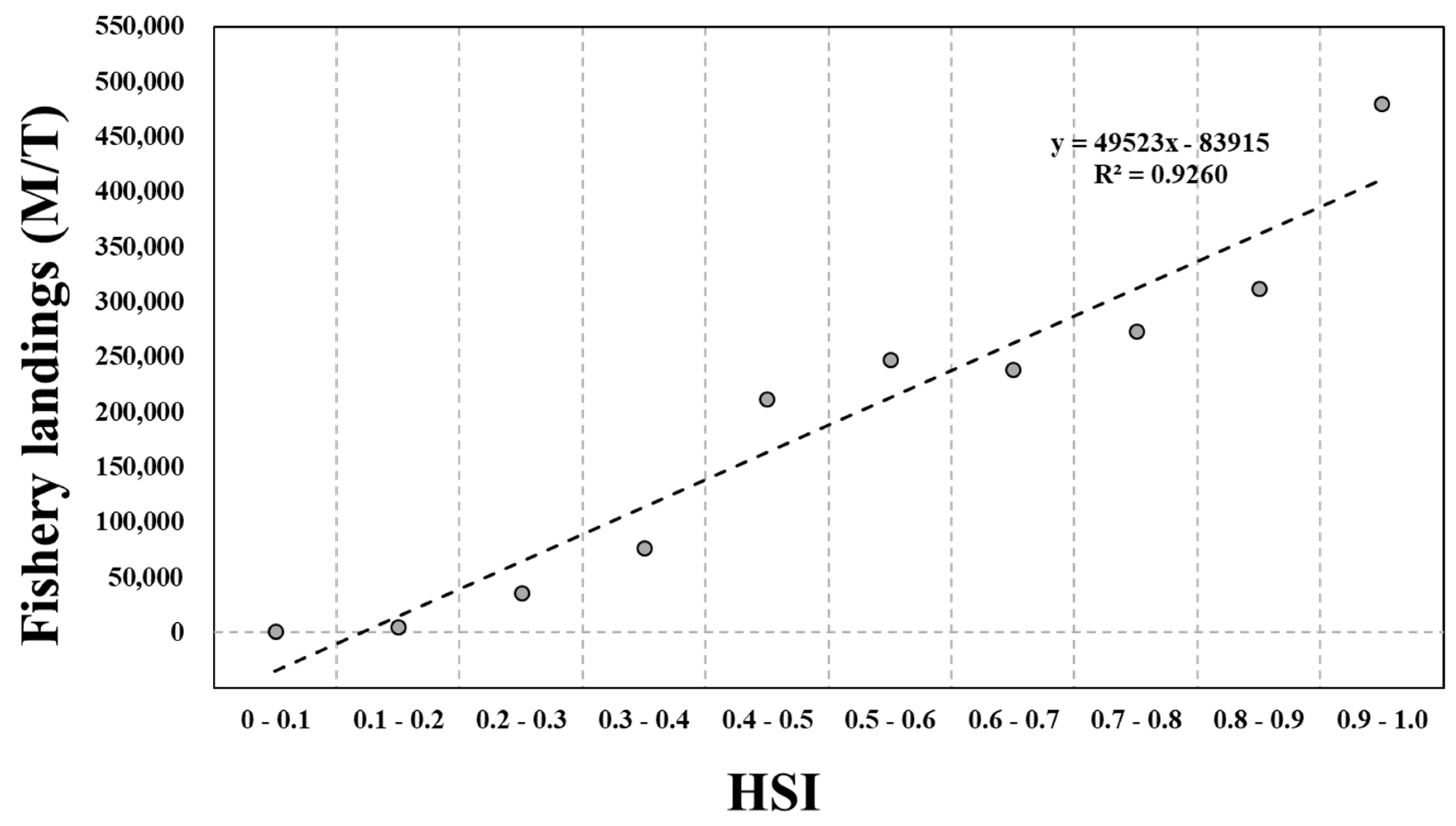
3. Results
3.1. Preferred Environmental Conditions of the Todarodes Pacificus
3.2. HSI Model Derivation
3.3. HSI Model Evaluation
4. Discussion
4.1. Environmental Conditions
4.2. Seasonal Variation of the HSI
4.3. Regions with High Catch Probability
5. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Korea Statistics; Korean Statistical Information Service (KOSIS). Fishery Production Survey; Korean Statistical Information Service (KOSIS): Daejeon, Korea, 2019. [Google Scholar]
- Kiyofuji, H.; Saitoh, S. Use of Nighttime Visible Images to Detect Japanese Common Squid Todarodes Pacificus Fishing Areas and Potential Migration Routes in the Sea of Japan. Mar. Ecol. Prog. Ser. 2004, 276, 173–186. [Google Scholar] [CrossRef]
- Kim, G.B.; Stapleton, H.M. PBDEs, Methoxylated PBDEs and HBCDs in Japanese Common Squid (Todarodes Pacificus) from Korean Offshore Waters. Mar. Pollut. Bull. 2010, 60, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Waska, H.; Kim, G.; Kim, G.B. Comparison of S, Se, and 210 Po Accumulation Patterns in Common Squid Todarodes Pacificus from the Yellow Sea and East/Japan Sea. Ocean Sci. J. 2013, 48, 215–224. [Google Scholar] [CrossRef]
- Alabia, I.; Dehara, M.; Saitoh, S.; Hirawake, T. Seasonal Habitat Patterns of Japanese Common Squid (Todarodes Pacificus) Inferred from Satellite-Based Species Distribution Models. Remote Sens. 2016, 8, 921. [Google Scholar] [CrossRef]
- Park, C.; Kim, Y.; Park, J.; Kim, Z.; Choi, Y.; Lee, D.; Choi, K.; Kim, S.; Hwang, K. Ecology and Fishing Grounds of Major Commercial Fish Species in the Coastal and Offshore Korean Waters; National Fisheries Research and Development Institute Busan: Busan, Korea, 1998. (In Korean) [Google Scholar]
- Kang, Y.S.; Kim, J.Y.; Kim, H.G.; Park, J.H. Long-term Changes in Zooplankton and its Relationship with Squid, Todarodes Pacificus, Catch in Japan/East Sea. Fish. Oceanogr. 2002, 11, 337–346. [Google Scholar] [CrossRef]
- Jones, D.D.; Walters, C.J. Catastrophe Theory and Fisheries Regulation. J. Fish. Board Can. 1976, 33, 2829–2833. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts Across Natural Systems. Nature 2003, 421, 37. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate Change and Distribution Shifts in Marine Fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef]
- Daskalov, G.M.; Grishin, A.N.; Rodionov, S.; Mihneva, V. Trophic Cascades Triggered by Overfishing Reveal Possible Mechanisms of Ecosystem Regime Shifts. Proc. Natl. Acad. Sci. USA 2007, 104, 10518–10523. [Google Scholar] [CrossRef]
- Brooks, R.P. Improving Habitat Suitability Index Models. Wildl. Soc. Bull. 1997, 25, 163–167. [Google Scholar]
- Morris, L.; Ball, D. Habitat Suitability Modelling of Economically Important Fish Species with Commercial Fisheries Data. ICES J. Mar. Sci. 2006, 63, 1590–1603. [Google Scholar] [CrossRef]
- Chen, X.; Li, G.; Feng, B.; Tian, S. Habitat Suitability Index of Chub Mackerel (Scomber Japonicus) from July to September in the East China Sea. J. Oceanogr. 2009, 65, 93–102. [Google Scholar] [CrossRef]
- Galparsoro, I.; Borja, Á.; Bald, J.; Liria, P.; Chust, G. Predicting Suitable Habitat for the European Lobster (Homarus Gammarus), on the Basque Continental Shelf (Bay of Biscay), using Ecological-Niche Factor Analysis. Ecol. Model. 2009, 220, 556–567. [Google Scholar] [CrossRef]
- Li, G.; Chen, X.; Lei, L.; Guan, W. Distribution of Hotspots of Chub Mackerel Based on Remote-Sensing Data in Coastal Waters of China. Int. J. Remote Sens. 2014, 35, 4399–4421. [Google Scholar] [CrossRef]
- Lee, D.; Son, S.; Kim, W.; Park, J.; Joo, H.; Lee, S. Spatio-Temporal Variability of the Habitat Suitability Index for Chub Mackerel (Scomber Japonicus) in the East/Japan Sea and the South Sea of South Korea. Remote Sens. 2018, 10, 938. [Google Scholar] [CrossRef]
- Rubec, P.J.; Bexley, J.C.; Norris, H.; Coyne, M.; Monaco, M.; Smith, S.; Ault, J. Suitability Modeling to Delineate Habitat Essential. Am. Fish. Soc. Symp. 1999, 22, 108–133. [Google Scholar]
- Vinagre, C.; Fonseca, V.; Cabral, H.; Costa, M.J. Habitat Suitability Index Models for the Juvenile Soles, Solea Solea and Solea Senegalensis, in the Tagus Estuary: Defining Variables for Species Management. Fish. Res. 2006, 82, 140–149. [Google Scholar] [CrossRef]
- Attrill, M.J.; Power, M. Climatic Influence on a Marine Fish Assemblage. Nature 2002, 417, 275. [Google Scholar] [CrossRef]
- Boeuf, G.; Payan, P. How should Salinity Influence Fish Growth? Com. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 411–423. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Knust, R. Climate Change Affects Marine Fishes through the Oxygen Limitation of Thermal Tolerance. Science 2007, 315, 95–97. [Google Scholar] [CrossRef]
- Zainuddin, M.; Kiyofuji, H.; Saitoh, K.; Saitoh, S. Using Multi-Sensor Satellite Remote Sensing and Catch Data to Detect Ocean Hot Spots for Albacore (Thunnus Alalunga) in the Northwestern North Pacific. Deep Sea Res. II 2006, 53, 419–431. [Google Scholar] [CrossRef]
- Robinson, C.J.; Gómez-Gutiérrez, J.; de León, D.A.S. Jumbo Squid (Dosidicus Gigas) Landings in the Gulf of California Related to Remotely Sensed SST and Concentrations of Chlorophyll a (1998–2012). Fish. Res. 2013, 137, 97–103. [Google Scholar] [CrossRef]
- Tian, Y.; Akamine, T.; Suda, M. Variations in the Abundance of Pacific Saury (Cololabis Saira) from the Northwestern Pacific in Relation to Oceanic-Climate Changes. Fish. Res. 2003, 60, 439–454. [Google Scholar] [CrossRef]
- NASA Goddard Space Flight Center; Ocean Ecology Laboratory; Ocean Biology Processing Group. Moderate-Resolution Imaging Spectroradiometer (MODIS) Aqua 11µm Day/Night Sea Surface Temperature Data; 2018 Reprocessing; NASA OB.DAAC: Greenbelt, MD, USA, 2018. Available online: https://oceandata.sci.gsfc.nasa.gov/MODIS-Aqua/Mapped/Monthly/4km/sst/ (accessed on 11 September 2018). [CrossRef]
- NASA Goddard Space Flight Center; Ocean Ecology Laboratory; Ocean Biology Processing Group. Moderate-Resolution Imaging Spectroradiometer (MODIS) Aqua Chlorophyll Data; 2018 Reprocessing; NASA OB.DAAC: Greenbelt, MD, USA, 2018. Available online: https://oceandata.sci.gsfc.nasa.gov/MODIS-Aqua/Mapped/Monthly/4km/chlor_a/ (accessed on 11 September 2018). [CrossRef]
- NASA Goddard Space Flight Center; Ocean Ecology Laboratory; Ocean Biology Processing Group. Moderate-Resolution Imaging Spectroradiometer (MODIS) Aqua Photosynthetically Available Radiation Data; 2018 Reprocessing; NASA OB.DAAC: Greenbelt, MD, USA, 2018. Available online: https://oceandata.sci.gsfc.nasa.gov/MODIS-Aqua/Mapped/Monthly/4km/par/ (accessed on 11 September 2018). [CrossRef]
- NASA Goddard Space Flight Center; Ocean Ecology Laboratory; Ocean Biology Processing Group. Moderate-Resolution Imaging Spectroradiometer (MODIS) Aqua Downwelling Diffuse Attenuation Coefficient Data; 2018 Reprocessing; NASA OB.DAAC: Greenbelt, MD, USA, 2018. Available online: https://oceandata.sci.gsfc.nasa.gov/MODIS-Aqua/Mapped/Monthly/4km/Kd_490/ (accessed on 11 September 2018). [CrossRef]
- Behrenfeld, M.J.; Falkowski, P.G. Photosynthetic Rates Derived from satellite-based Chlorophyll Concentration. Limnol. Oceanogr. 1997, 42, 1–20. [Google Scholar] [CrossRef]
- Kirk, J.T. Light and Photosynthesis in Aquatic Ecosystems, 2nd ed.; Cambridge University Press: Cambridge, UK, 1994; pp. 129–169. [Google Scholar]
- Yamada, K.; Ishizaka, J.; Nagata, H. Spatial and Temporal Variability of Satellite Primary Production in the Japan Sea from 1998 to 2002. J. Oceanogr. 2005, 61, 857–869. [Google Scholar] [CrossRef]
- Yen, K.; Lu, H.; Chang, Y.; Lee, M. Using Remote-Sensing Data to Detect Habitat Suitability for Yellowfin Tuna in the Western and Central Pacific Ocean. Int. J. Remote Sens. 2012, 33, 7507–7522. [Google Scholar] [CrossRef]
- Chen, X.; Feng, B.; Xu, L. A Comparative Study on Habitat Suitability Index of Bigeye Tuna, Thunnus obesus in the Indian Ocean. J. Fish. Sci. China 2008, 15, 269–278. [Google Scholar]
- Grebenkov, A.; Lukashevich, A.; Linkov, I.; Kapustka, L.A. A habitat suitability evaluation technique and its application to environmental risk assessment. In Ecotoxicology, Ecological Risk Assessment and Multiple Stressors; Springer: Berlin/Heidelberg, Germany, 2006; pp. 191–201. [Google Scholar]
- Hess, G.R.; Bay, J.M. A Regional Assessment of Windbreak Habitat Suitability. Environ. Monit. Assess. 2000, 61, 239–256. [Google Scholar] [CrossRef]
- Lauver, C.L.; Busby, W.H.; Whistler, J.L. Testing a GIS Model of Habitat Suitability for a Declining Grassland Bird. Environ. Manag. 2002, 30, 88–97. [Google Scholar] [CrossRef]
- Van der Lee, G.E.M.; Van der Molen, D.T.; Van den Boogaard, H.F.P.; Van der Klis, H. Uncertainty Analysis of a Spatial Habitat Suitability Model and Implications for Ecological Management of Water Bodies. Landsc. Ecol. 2006, 21, 1019–1032. [Google Scholar] [CrossRef]
- Kaschner, K.; Kesner-Reyes, K.; Garilao, C.; Rius-Barile, J.; Rees, T.; Froese, R. AquaMaps: Predicted Range Maps for Aquatic Species; Version August 2016; World Wide Web Electronic Publication; Available online: www.aquamaps.org (accessed on 2 January 2019).
- Castillo, J.; Barbieri, M.; Gonzalez, A. Relationships between Sea Surface Temperature, Salinity, and Pelagic Fish Distribution off Northern Chile. ICES J. Mar. Sci. 1996, 53, 139–146. [Google Scholar] [CrossRef]
- Jaureguizar, A.J.; Menni, R.; Guerrero, R.; Lasta, C. Environmental Factors Structuring Fish Communities of the Rıo de la Plata Estuary. Fish. Res. 2004, 66, 195–211. [Google Scholar] [CrossRef]
- Sutcliffe, W., Jr.; Drinkwater, K.; Muir, B. Correlations of Fish Catch and Environmental Factors in the Gulf of Maine. J. Fish. Board Can. 1977, 34, 19–30. [Google Scholar] [CrossRef]
- Smith, R.; Dustan, P.; Au, D.; Baker, K.; Dunlap, E. Distribution of Cetaceans and Sea-Surface Chlorophyll Concentrations in the California Current. Mar. Biol. 1986, 91, 385–402. [Google Scholar] [CrossRef]
- Lee, D.; An, Y.R.; Park, K.J.; Kim, H.W.; Lee, D.; Joo, H.T.; Oh, Y.G.; Kim, S.M.; Kang, C.K.; Lee, S.H. Spatial Distribution of Common Minke Whale (Balaenoptera acutorostrata) as an Indication of a Biological Hotspot in the East Sea. Deep Sea Res. II 2017, 143, 91–99. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Reported Catches | No. of Matchable Catches |
|---|---|---|
| 2010 | 797 | 486 |
| 2011 | 1593 | 956 |
| 2012 | 1479 | 918 |
| 2013 | 1372 | 834 |
| 2014 | 1219 | 775 |
| 2015 | 1640 | 1099 |
| 2016 | 1301 | 892 |
| Total | 9401 | 5960 |
| Variables | SI models | RMSE | R2 |
|---|---|---|---|
| SSTWS | exp(−0.2929(XSST − 15.44)2) | 0.0500 | 0.9654 |
| SSTSA | exp(−0.0657(XSST − 24.22)2) | 0.0745 | 0.9461 |
| SSHAWS | exp(−184.8(XSST + 0.01325)2) | 0.0871 | 0.9063 |
| SSHASA | exp(−157(XSST − 0.1514)2) | 0.0667 | 0.9468 |
| Chl-a | exp(−16.98(ln(XChl) + 0.5997)2) | 0.0640 | 0.9165 |
| PP | exp(−0.00004637(XPP − 658.7)2) | 0.0876 | 0.9072 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Son, S.H.; Lee, C.-I.; Kang, C.-K.; Lee, S.H. Spatio-Temporal Variability of the Habitat Suitability Index for the Todarodes pacificus (Japanese Common Squid) around South Korea. Remote Sens. 2019, 11, 2720. https://doi.org/10.3390/rs11232720
Lee D, Son SH, Lee C-I, Kang C-K, Lee SH. Spatio-Temporal Variability of the Habitat Suitability Index for the Todarodes pacificus (Japanese Common Squid) around South Korea. Remote Sensing. 2019; 11(23):2720. https://doi.org/10.3390/rs11232720
Chicago/Turabian StyleLee, Dabin, Seung Hyun Son, Chung-Il Lee, Chang-Keun Kang, and Sang Heon Lee. 2019. "Spatio-Temporal Variability of the Habitat Suitability Index for the Todarodes pacificus (Japanese Common Squid) around South Korea" Remote Sensing 11, no. 23: 2720. https://doi.org/10.3390/rs11232720